
The Antibacterial Properties of Polish Honey against Streptococcus mutans—A Causative Agent of Dental Caries


Abstract
:1. Introduction
2. Results and Discussion
2.1. Antioxidant Properties
2.1.1. Total Phenolic Content
2.1.2. Antioxidant Properties Measured by FRAP Method
2.2. GOX Activity
2.3. Hydrogen Peroxide Content
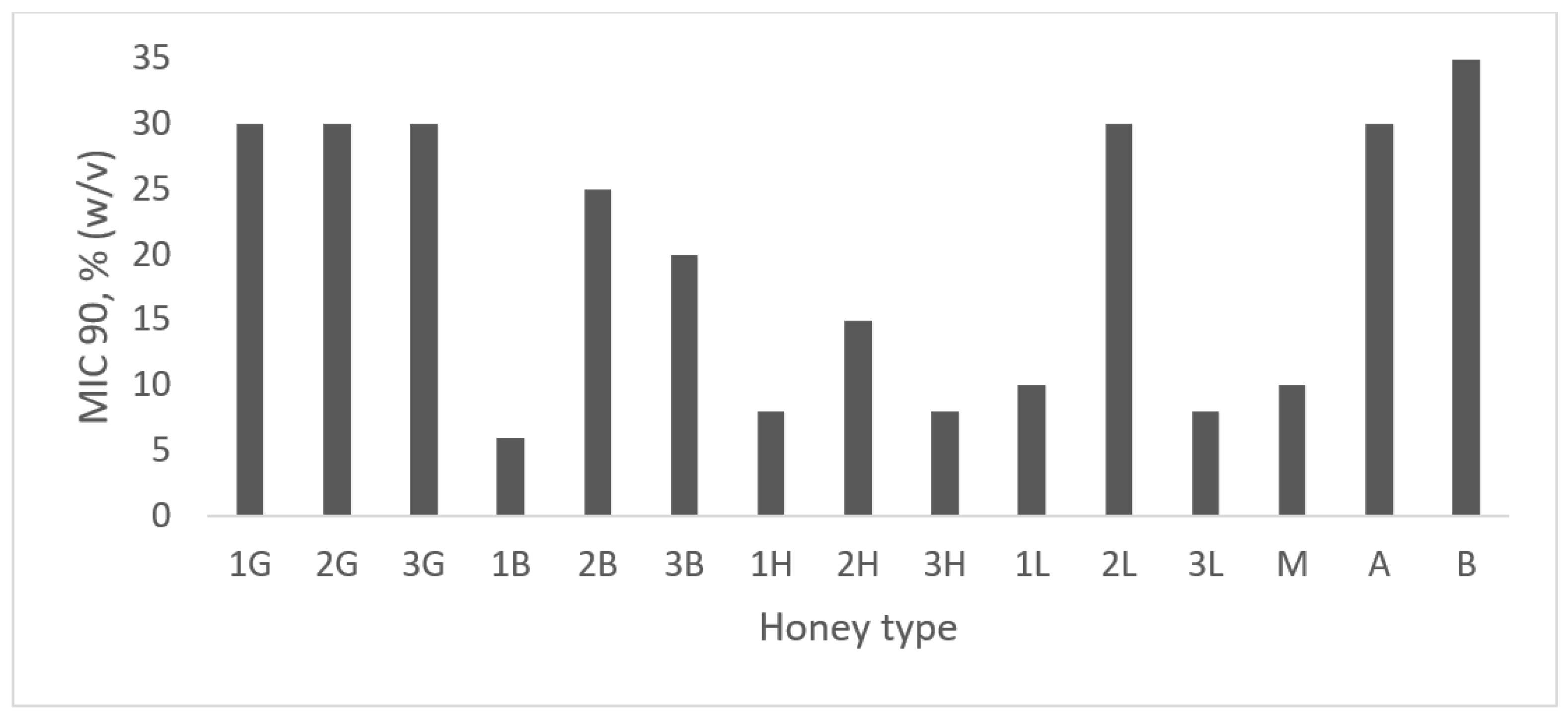
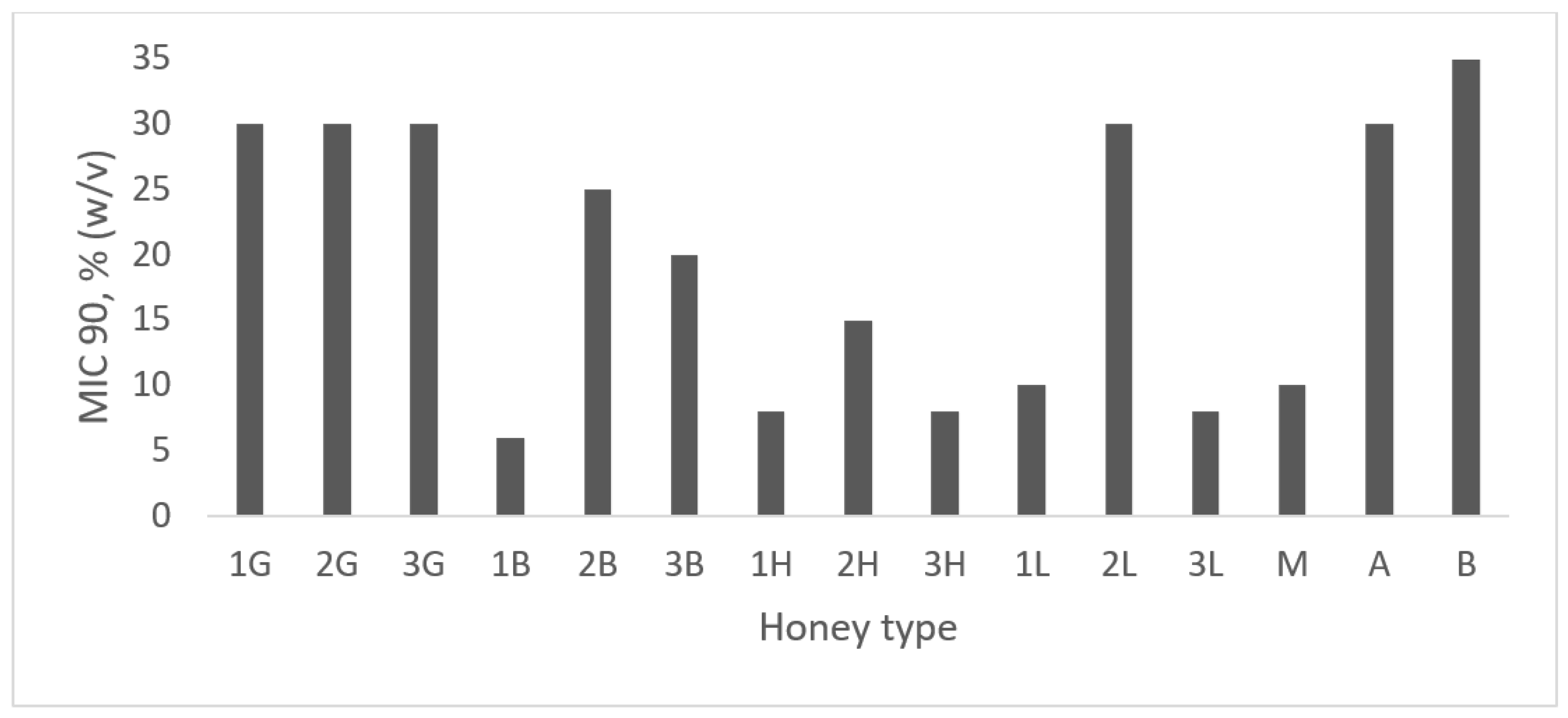
2.4. Antibacterial Activity
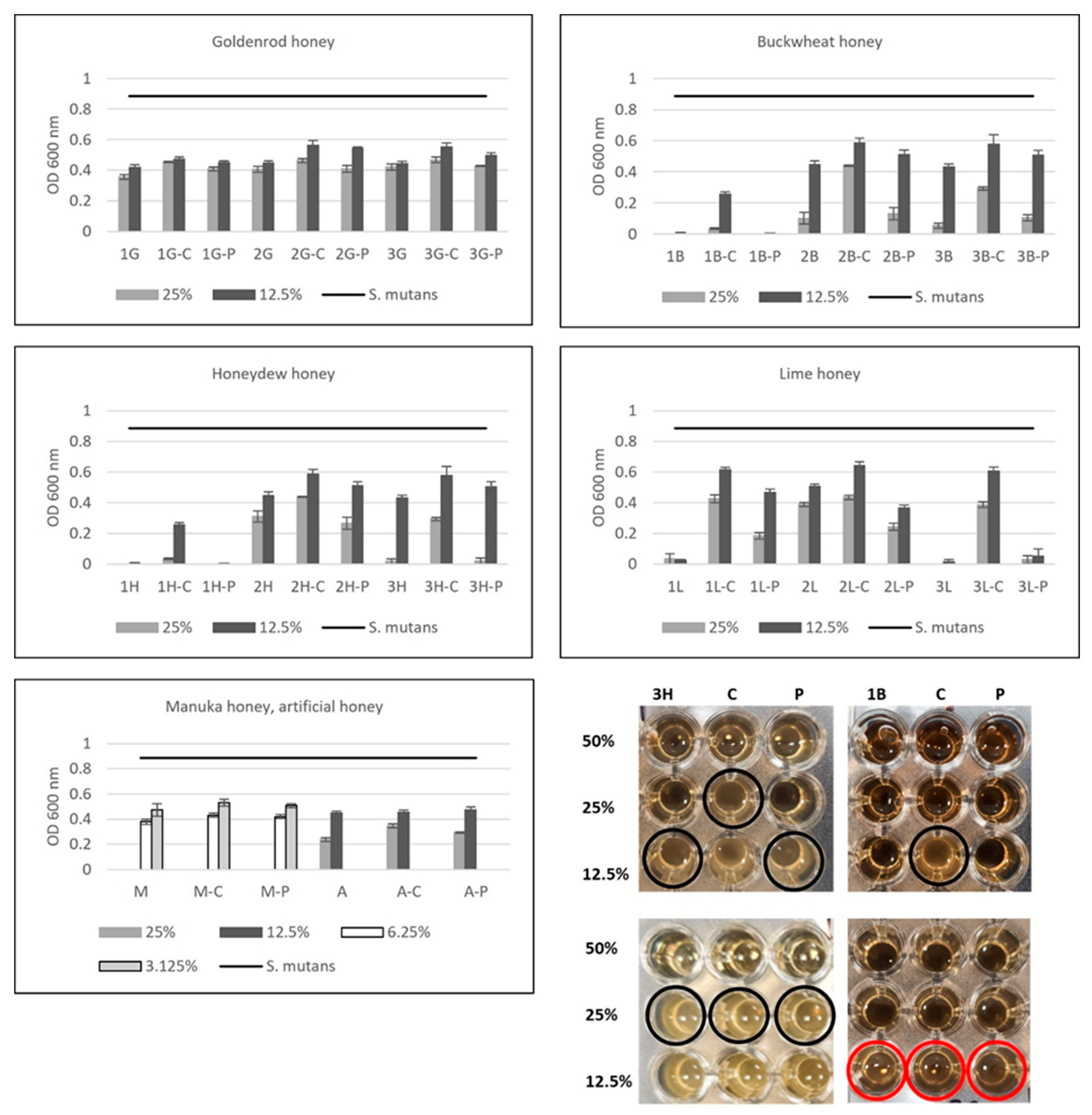
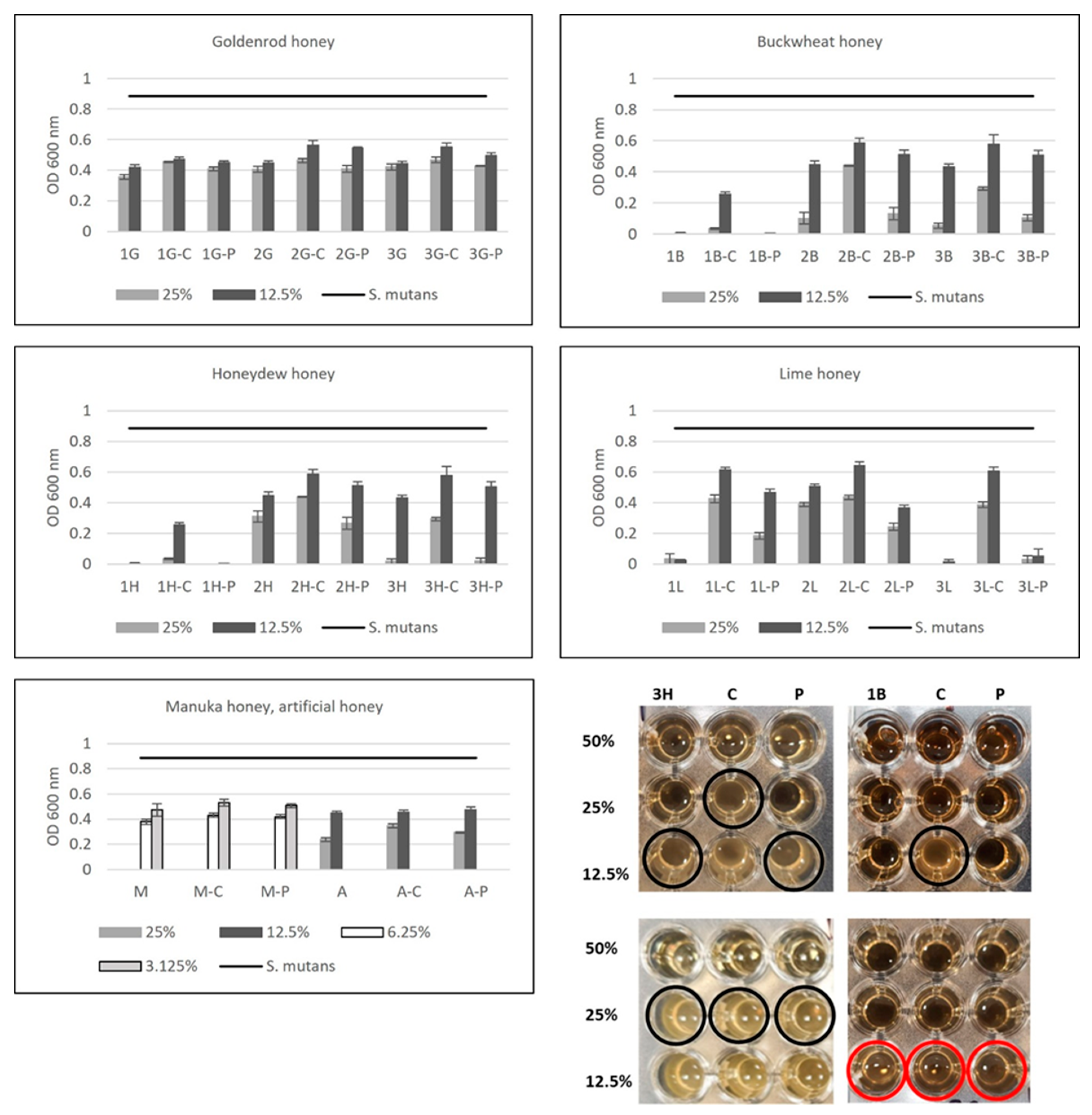
2.5. Antibacterial Activity of Honey after Enzymatic Treatment with Catalase and Proteinase K—The Role of Proteins and Hydrogen Peroxide in S. mutans Growth Inhibition
2.6. Antibiofilm Activity
3. Materials and Methods
3.1. Antioxidant Potential
3.1.1. Total Polyphenol Content (TPC)
3.1.2. Antioxidant Potential Measured by FRAP
3.2. Glucose Oxidase Activity Determination
3.3. Hydrogen Peroxide Determination
3.4. Antibacterial Properties of Honey—Minimum Inhibitory Concentration (MIC)
3.5. Antibacterial Activity of Honey after Enzymatic Treatment with Catalase and Proteinase K
3.6. Biofilm Inhibition Activity
3.7. Biofilm Eradication Assay
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, X.; Daliri, E.B.; Kim, N.; Kim, J.R.; Yoo, D.; Oh, D.H. Microbial Etiology and Prevention of Dental Caries: Exploiting Natural Products to Inhibit Cariogenic Biofilms. Pathogens 2020, 9, 569. [Google Scholar] [CrossRef] [PubMed]
- WHO. Global Oral Health Status Report: Towards Universal Health Coverage for Oral Health by 2030; WHO: Geneva, Switzerland, 2022; ISBN 978-92-4-006148-4. [Google Scholar]
- Deglovic, J.; Majtanova, N.; Majtan, J. Antibacterial and Antibiofilm Effect of Honey in the Prevention of Dental Caries: A Recent Perspective. Foods 2022, 11, 2670. [Google Scholar] [CrossRef] [PubMed]
- Fabres-Klein, M.H.; Caizer Santos, M.J.; Contelli Klein, R.; Nunes de Souza, G.; de Oliveira Barros Ribon, A. An association between milk and slime increases biofilm production by bovine Staphylococcus aureus. BMC Vet. Res. 2015, 11, 3. [Google Scholar] [CrossRef]
- Matsumoto-Nakano, M. Role of Streptococcus mutans surface proteins for biofilm formation. Jpn. Dent. Sci. Rev. 2018, 54, 22–29. [Google Scholar] [CrossRef]
- Kamarehei, F.; Mehdiabadi, M.; Naderi, F. Antibacterial effects of natural compoundson biofilm formation of Streptococcus mutans. Clin. Exp. Dent. Res. 2022, 8, 1426–1433. [Google Scholar] [CrossRef]
- Kawakita, E.R.H.; Ré, A.C.S.; Peixoto, M.P.G.; Ferreira, M.P.; Ricomini-Filho, A.P.; Freitas, O.; Aires, C.P. Effect of Chitosan Dispersion and Microparticles on Older Streptococcus mutans Biofilms. Molecules 2019, 24, 1808. [Google Scholar] [CrossRef]
- Mishra, R.; Panda, A.K.; De Mandal, S.; Shakeel, M.; Bisht, S.S.; Khan, J. Natural anti-biofilm agents: Strategies to control Biofilm-forming pathogens. Front. Microbiol. 2020, 11, 566325. [Google Scholar] [CrossRef]
- Mehrishi, P.; Agarwal, P.; Shobha Broor, S.; Sharma, A. Antibacterial and Antibiofilm Properties of Medicinal Plant Extracts against Multi Drug Resistant Staphylococcus Species and Non Fermenter Bacteria. J. Pure Appl. Microbiol. 2020, 14, 403–413. [Google Scholar] [CrossRef]
- Nijampatnam, B.; Zhang, H.; Cai, X.; Michalek, S.M.; Wu, H.; Velu, S.E. Inhibition of Streptococcus mutans Biofilms by the Natural Stilbene Piceatannol Through the Inhibition of Glucosyltransferases. ACS Omega 2018, 3, 8378–8385. [Google Scholar] [CrossRef]
- Aqawi, M.; Sionov, R.V.; Gallily, R.; Friedman, M.; Steinberg, D. Anti-Biofilm Activity of Cannabigerol against Streptococcus mutans. Microorganisms 2021, 9, 2031. [Google Scholar] [CrossRef]
- Koo, H.; Nino de Guzman, P.; Schobel, B.D.; Vacca Smith, A.V.; Bowen, W.H. Influence of cranberry juice on glucan-mediated processes involved in Streptococcus mutans biofilm development. Caries Res. 2006, 40, 20–27. [Google Scholar] [CrossRef]
- Thimothe, J.; Bonsi, I.A.; Padilla-Zakour, O.I.; Koo, H. Chemical characterization of red wine grape (Vitis vinifera and Vitis interspecific hybrids) and pomace phenolic extracts and their biological activity against Streptococcus mutans. J. Agric. Food Chem. 2007, 55, 10200–10207. [Google Scholar] [CrossRef]
- Cianciosi, D.; Forbes-Hernández, T.Y.; Afrin, S.; Gasparrini, M.; Reboredo-Rodriguez, P.; Manna, P.P.; Zhang, J.; Bravo Lamas, L.; Martínez Flórez, S.; Agudo Toyos, P.; et al. Phenolic Compounds in Honey and Their Associated Health Benefits: A Review. Molecules 2018, 23, 2322. [Google Scholar] [CrossRef]
- Grabek-Lejko, D.; Miłek, M.; Sidor, E.; Puchalski, C.; Dżugan, M. Antiviral and Antibacterial Effect of Honey Enriched with Rubus spp. as a Functional Food with Enhanced Antioxidant Properties. Molecules 2022, 27, 4859. [Google Scholar] [CrossRef]
- Almasaudi, S. The antibacterial activities of honey. Saudi J. Biol. Sci. 2021, 28, 2188–2196. [Google Scholar] [CrossRef]
- Dżugan, M.; Tomczyk, M.; Sowa, P.; Grabek-Lejko, D. Antioxidant Activity as Biomarker of Honey Variety. Molecules 2018, 23, 2069. [Google Scholar] [CrossRef]
- Albaridi, N.A. Antibacterial Potency of Honey. Int. J. Microbiol. 2019, 2019, 2464507. [Google Scholar] [CrossRef]
- Grabek-Lejko, D.; Słowik, J.; Kasprzyk, I. Activity of selected honey types against Staphylococcus aureus methicillin susceptible (MSSA) and methicillin resistant (MRSA) bacteria and its correlation with hydrogen peroxide, phenolic content and antioxidant capacity. Farmacia 2018, 66, 37–43. [Google Scholar]
- Nassar, H.M.; Li, M.; Gregory, R.L. Effect of honey on Streptococcus mutans growth and biofilm formation. Appl. Environ. Microbiol. 2012, 78, 536–540. [Google Scholar] [CrossRef]
- Patel, H.R.; Krishnan, C.A.; Thanveer, K. Antimicrobial effect of honey on Streptococcus mutans—An in vitro study. Int. J. Dent. Sci. Res. 2013, 1, 46–49. [Google Scholar] [CrossRef]
- Schmidlin, P.R.; English, H.; Duncan, W.; Belibasakis, G.N.; Thurnheer, T. Antibacterial potential of Manuka honey against three oral bacteria in vitro. Swiss Dent. J. 2014, 124, 922–924. [Google Scholar]
- Habluetzel, A.; Schmid, C.; Carvalho, T.S.; Lussi, A.; Eick, S. Impact of honey on dental erosion and adhesion of early bacterial colonizers. Sci. Rep. 2018, 8, 10936. [Google Scholar] [CrossRef]
- Voidarou, C.; Antoniadou, M.; Rozos, G.; Alexopoulos, A.; Giorgi, E.; Tzora, A.; Skoufos, I.; Varzakas, T.; Bezirtzoglou, E. An in vitro study of different types of Greek honey as potential natural antimicrobials against dental caries and other oral pathogenic microorganisms. Case study simulation of oral cavity conditions. Appl. Sci. 2021, 11, 6318. [Google Scholar] [CrossRef]
- Romário-Silva, D.; Alencar, S.M.; Bueno-Silva, B.; Sardi, J.d.C.O.; Franchin, M.; Carvalho, R.D.P.d.; Ferreira, T.E.d.S.A.; Rosalen, P.L. Antimicrobial Activity of Honey against Oral Microorganisms: Current Reality, Methodological Challenges and Solutions. Microorganisms 2022, 10, 2325. [Google Scholar] [CrossRef] [PubMed]
- Moynihan, P. Sugars and Dental Caries: Evidence for Setting a Recommended Threshold for Intake. Adv. Nutr. 2016, 7, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Tomczyk, M.; Bocian, A.; Sidor, E.; Miłek, M.; Zaguła, G.; Dżugan, M. The Use of HPTLC and SDS-PAGE Methods for Coniferous Honeydew Honey Fingerprinting Compiled with Mineral Content and Antioxidant Activity. Molecules 2022, 27, 720. [Google Scholar] [CrossRef]
- Directive, E.U. Directive 2014/63/EU of the European Parliament and of the Council amending Council Directive 2001/110/EC relating to honey. Off. J. Eur. Communities 2014, 164, 1–5. [Google Scholar]
- Gośliński, M.; Nowak, D.; Szwengiel, A. Multidimensional Comparative Analysis of Bioactive Phenolic Compounds of Honeys of Various Origin. Antioxidants 2021, 10, 530. [Google Scholar] [CrossRef] [PubMed]
- Bucekova, M.; Jardekova, L.; Juricova, V.; Bugarova, V.; Di Marco, G.; Gismondi, A.; Leonardi, D.; Farkasovska, J.; Godocikova, J.; Laho, M.; et al. Antibacterial Activity of Different Blossom Honeys: New Findings. Molecules 2019, 24, 1573. [Google Scholar] [CrossRef]
- Wilczyńska, A. Phenolic content and antioxidant activity of different types of Polish honey—A short report. Pol. J. Food Nutr. Sci. 2010, 60, 309–313. [Google Scholar]
- Puścion-Jakubik, A.; Karpińska, E.; Moskwa, J.; Socha, K. Content of Phenolic Acids as a Marker of Polish Honey Varieties and Relationship with Selected Honey-Quality-Influencing Variables. Antioxidants 2022, 11, 1312. [Google Scholar] [CrossRef] [PubMed]
- Gośliński, M.; Nowak, D.; Kłębukowska, L. Antioxidant properties and antimicrobial activity of manuka honey versus Polish honeys. J. Food Sci. Technol. 2020, 57, 1269–1277. [Google Scholar] [CrossRef] [PubMed]
- Socha, R.; Juszczak, L.; Pietrzyk, S.; Gałkowska, D.; Fortuna, T.; Witczak, T. Phenolic profile and antioxidant properties of Polish honeys. Int. J. Food Sci. Tech. 2011, 46, 528–534. [Google Scholar] [CrossRef]
- Becerril-Sánchez, A.L.; Quintero-Salazar, B.; Dublán-García, O.; Escalona-Buendía, H.B. Phenolic Compounds in Honey and Their Relationship with Antioxidant Activity, Botanical Origin, and Color. Antioxidants 2021, 10, 1700. [Google Scholar] [CrossRef] [PubMed]
- Alshareef, R.M.H.; Al-Farhan, B.S.; Mohammed, M.E.A. Glucose Oxidase and Catalase Activities in Honey Samples from the Southwestern Region of Saudi Arabia. Appl. Sci. 2022, 12, 7584. [Google Scholar] [CrossRef]
- Strelec, I.; Crevar, B.; Kovač, T.; Rajs, B.B.; Primorac, L.; Flanjak, I. Glucose oxidase activity and hydrogen peroxide accumulation in Croatian honeys. Croat. J. Food Sci. Technol. 2018, 10, 33–41. [Google Scholar] [CrossRef]
- Bucekova, M.; Buriova, M.; Pekarik, L.; Majtan, V.; Majtan, J. Phytochemicals-mediated production of hydrogen peroxide is crucial for high antibacterial activity of honeydew honey. Sci. Rep. 2018, 8, 9061. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi-Motamayel, F.; Hendi, S.S.; Alikhani, M.Y.; Khamverdi, Z. Antibacterial activity of honey on cariogenic bacteria. J. Dent. 2013, 10, 10–15. [Google Scholar]
- Ghabanchi, J.; Bazargani, A.; Afkar, M.D.; Foroushan, S.B.; Aein, S.D. In vitro assessment of anti-Streptococcus mutans potential of honey. Iran. Red. Crescent Med. J. 2010, 12, 61–64. [Google Scholar]
- Basson, N.J.; Grobler, S.R. Antimicrobial activity of two South African honeys produced from indigenous Leucospermum cordifolium and Erica species on selected micro-organisms. BMC Complement. Altern. Med. 2008, 8, 41. [Google Scholar] [CrossRef]
- Brudzynski, K.; Lannigan, R. Mechanism of Honey Bacteriostatic Action Against MRSA and VRE Involves Hydroxyl Radicals Generated from Honey’s Hydrogen Peroxide. Front. Microbiol. 2012, 3, 36. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, D.M.; Krishnakumar, K.; Batres, M.A.; Hakola-Parry, A.; Cokcetin, N.; Harry, E.; Carter, D.A. A cost-effective colourimetric assay for quantifying hydrogen peroxide in honey. Access Microbiol. 2019, 1, e000065. [Google Scholar] [CrossRef]
- Roy, R.; Tiwari, M.; Donelli, G.; Tiwari, V. Strategies for combating bacterial biofilms: A focus on anti-biofilm agents and their mechanisms of action. Virulence 2018, 9, 522–554. [Google Scholar] [CrossRef]
- Johnston, M.; McBride, M.; Dahiya, D.; Owusu-Apenten, R.; Nigam, P.S. Antibacterial activity of Manuka honey and its components: An overview. AIMS Microbiol. 2018, 4, 655–664. [Google Scholar] [CrossRef]
- Haney, E.F.; Trimble, M.J.; Cheng, J.T.; Vallé, Q.; Hancock, R.E.W. Critical Assessment of Methods to Quantify Biofilm Growth and Evaluate Antibiofilm Activity of Host Defence Peptides. Biomolecules 2018, 8, 29. [Google Scholar] [CrossRef] [PubMed]
- Zayed, S.M.; Aboulwafa, M.M.; Hashem, A.M.; Saleh, S.E. Biofilm formation by Streptococcus mutans and its inhibition by green tea extracts. AMB Express. 2021, 11, 73. [Google Scholar] [CrossRef] [PubMed]
- Kwasny, S.; Opperman, T. Static biofilm cultures of gram positive Pathogens Grown in a Microtiter Format Used for Anti-Biofilm Drug Discovery. Curr. Protoc. Pharmacol. 2010, 50, 13A-8. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Honey Type | Honey Sample | TPC (mg GAE/100 g) | FRAP (µmol TE/100 g) |
|---|---|---|---|
| Goldenrod | 1G | 35.43 ± 1.52 c,d | 91.23 ± 1.41 a,b |
| 2G | 33.42 ± 0.60 b,c | 97.73 ± 5.66 a,b,c | |
| 3G | 30.27 ± 0.71 a,b | 75.07 ± 3.61 a | |
| Buckwheat | 1B | 139.73 ± 3.23 j | 796.40 ± 43.24 k |
| 2B | 97.99 ± 5.00 i | 525.60 ± 7.63 i | |
| 3B | 88.72 ± 4.43 h | 354.40 ± 35.36 g | |
| Honeydew | 1H | 64.25 ± 2.44 f | 343.60 ± 5.60 g |
| 2H | 69.27 ± 5.36 g | 343.60 ± 21.90 g | |
| 3H | 98.72 ± 3.44 i | 584,13 ± 11.21 j | |
| Lime | 1L | 36.03 ± 2.93 c,d | 141.60 ± 4.92 d,e |
| 2L | 48.22 ± 2.38 e | 236.33 ± 9.97 f | |
| 3L | 40.05 ± 2.00 d | 167.07 ± 16.15 e | |
| Manuka | M | 84.75 ± 1.33 h | 403.60 ± 21.14 h |
| Artificial | A | 27.58 ± 0.42 a | 125.73 ± 0.83 c,d |
| B | 28.22 ± 0.76 a | 122.00 ± 2.12 b,c,d |
| Honey Type | Honey Sample | H2O2 (umol/mL *) | GOX (mU/mL) |
|---|---|---|---|
| Goldenrod | 1G | 39.57 ± 0.49 c | 23.17 ± 0.35 c |
| 2G | 17.74 ± 1.12 b | 16.74 ± 0.01 b | |
| 3G | 100.32 ± 17.33 d | 26.39 ± 0.58 c | |
| Buckwheat | 1B | 114.73 ± 4.52 e | 87.26 ± 2.92 g |
| 2B | 23.12 ± 0.99 b | 47.38 ± 0.94 e | |
| 3B | 161.30 ± 1.05 f | 36.91 ± 1.93 d | |
| Honeydew | 1H | 161.29 ± 1.61 f | 70.73 ± 2.23 f |
| 2H | 164.52 ± 15.97 f | 84.53 ± 2.60 g | |
| 3H | 16.67 ± 0.37 b | 139.14 ± 3.19 i | |
| Lime | 1L | 17.74 ± 0.32 b | 71.80 ± 4.44 f |
| 2L | 11.83 ± 0.93 b | 120.20 ± 4.53 h | |
| 3L | 215.81 ± 7.38 g | 25.71 ± 0.59 c | |
| Manuka | M | 16.88 ± 0.49 b | 17.22 ± 0.40 b |
| Artificial | A | 0.00 ± 0.00 a | 3.38 ± 0.40 a |
| B | 0.00 ± 0.00 a | 3.45 ± 0.32 a |
| Honey Type | Honey Sample | Biofilm Formation MBIC 90 | Biofilm Eradication MBIC 50 |
|---|---|---|---|
| Goldenrod | 1G | 30 | 45 |
| 2G | 30 | >50 | |
| 3G | 30 | 50 | |
| Buckwheat | 1B | 15 | 10 |
| 2B | 30 | 40 | |
| 3B | 30 | 45 | |
| Honeydew | 1H | 20 | 25 |
| 2H | 15 | 50 | |
| 3H | 20 | 45 | |
| Lime | 1L | 25 | 25 |
| 2L | 30 | >50 | |
| 3L | 20 | >50 | |
| Manuka | M | 20 | 50 |
| Artificial | A | 30 | >50 |
| B | 35 | >50 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grabek-Lejko, D.; Hyrchel, T. The Antibacterial Properties of Polish Honey against Streptococcus mutans—A Causative Agent of Dental Caries. Antibiotics 2023, 12, 1640. https://doi.org/10.3390/antibiotics12111640
Grabek-Lejko D, Hyrchel T. The Antibacterial Properties of Polish Honey against Streptococcus mutans—A Causative Agent of Dental Caries. Antibiotics. 2023; 12(11):1640. https://doi.org/10.3390/antibiotics12111640
Chicago/Turabian StyleGrabek-Lejko, Dorota, and Tomasz Hyrchel. 2023. "The Antibacterial Properties of Polish Honey against Streptococcus mutans—A Causative Agent of Dental Caries" Antibiotics 12, no. 11: 1640. https://doi.org/10.3390/antibiotics12111640
APA StyleGrabek-Lejko, D., & Hyrchel, T. (2023). The Antibacterial Properties of Polish Honey against Streptococcus mutans—A Causative Agent of Dental Caries. Antibiotics, 12(11), 1640. https://doi.org/10.3390/antibiotics12111640