
Molecular Characteristics and Prevalence of Rifampin Resistance in Staphylococcus aureus Isolates from Patients with Bacteremia in South Korea

 , ,
, ,
Abstract
:1. Introduction
2. Results
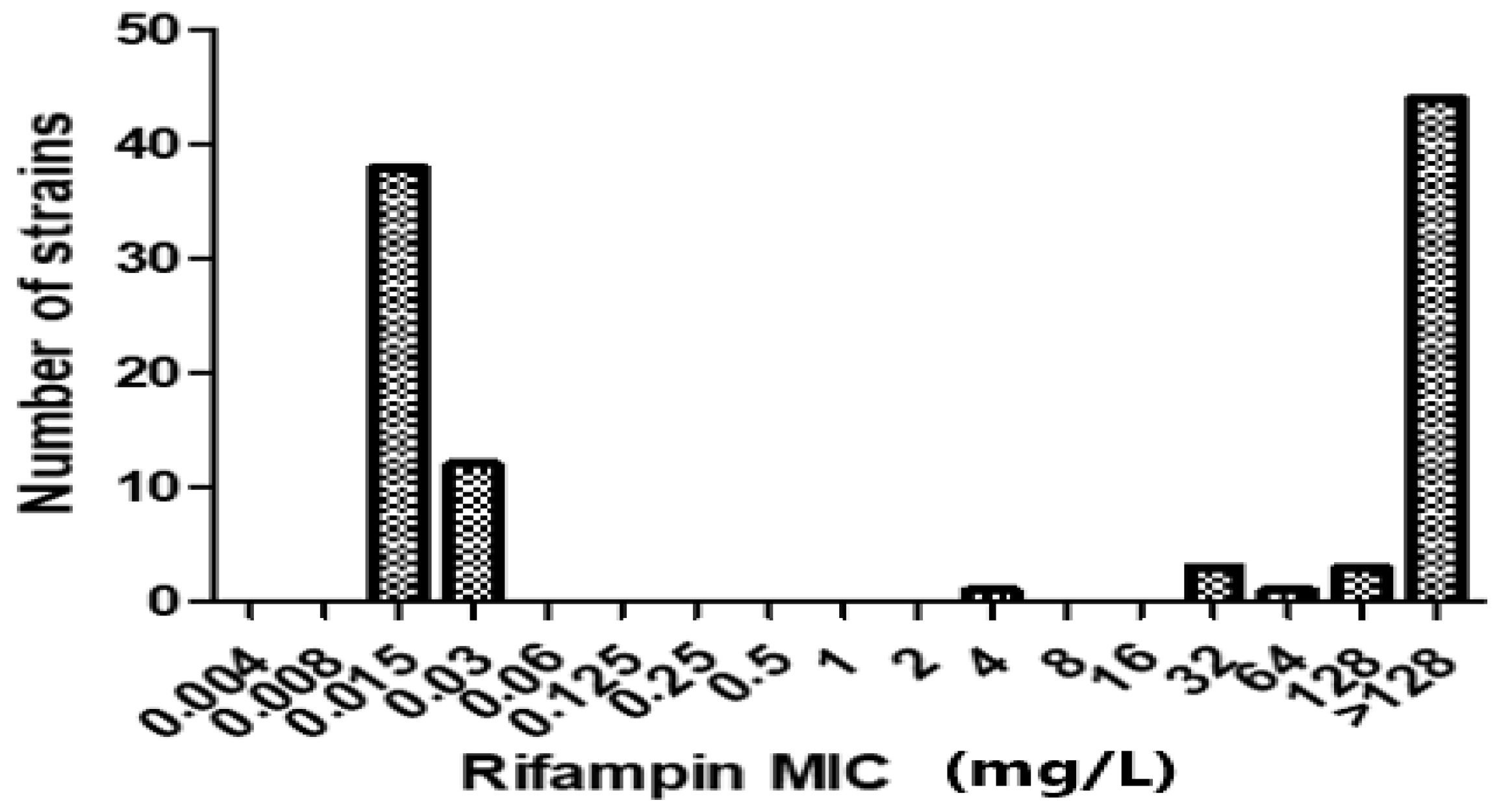
2.1. Characteristics of Study Isolates and Profile of rpoB Gene Mutations
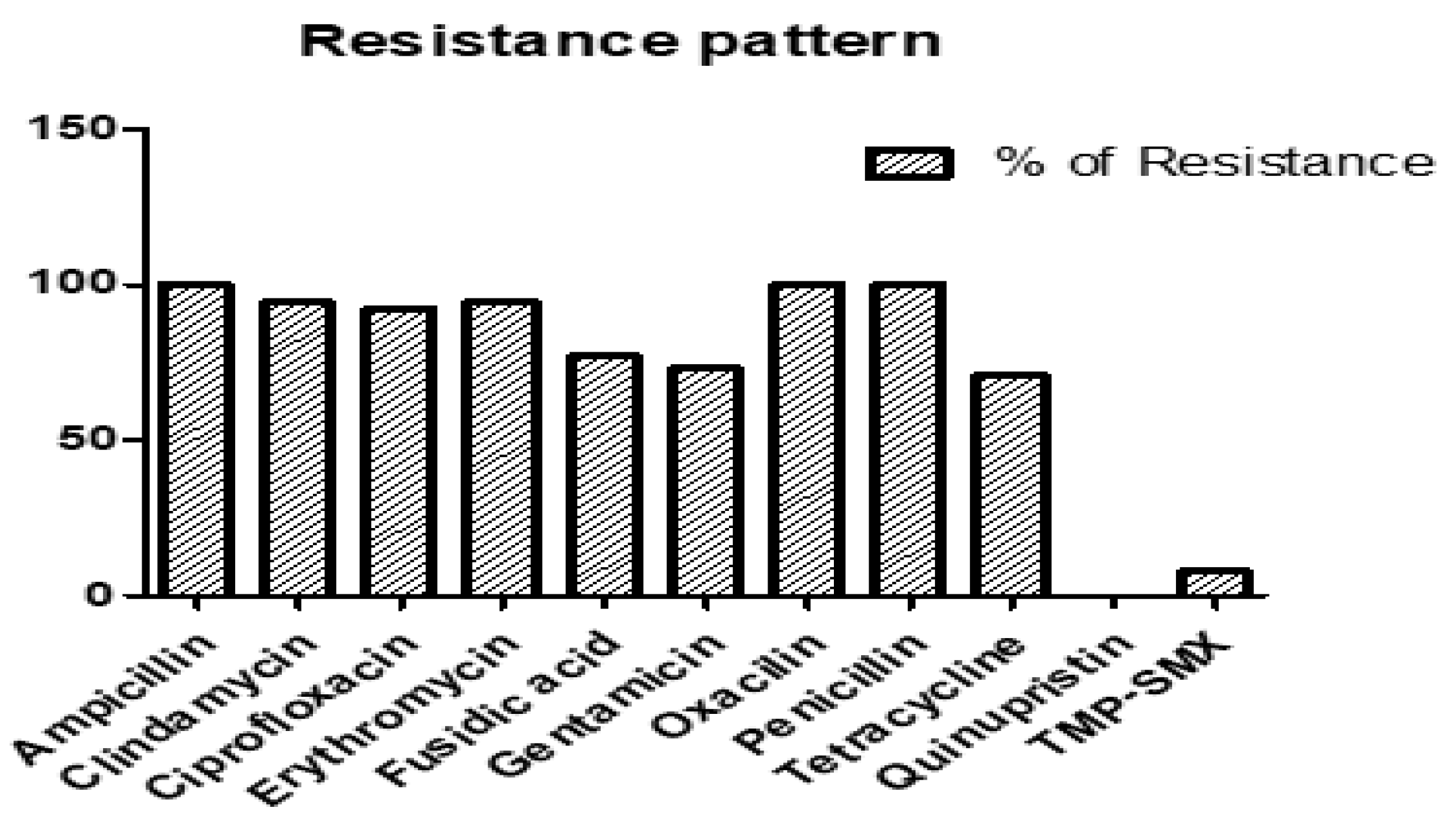
2.2. Antimicrobial Resistance Profiles of RIF-R MRSA Isolates
2.3. Molecular Typing
2.4. δ-Hemolysin Activities As Genotypic Characteristics of S. aureus Isolates by Rifampin
3. Discussion
4. Materials and Methods
4.1. Collection of S. aureus Isolates
4.2. Agr Functionality Test
4.3. Antimicrobial Susceptibility Tests
4.4. Molecular Typing
4.4.1. Detection of the mecA Gene
4.4.2. Multilocus Sequence Typing
4.4.3. SCCmec Typing
4.4.4. Spa Typing
4.5. PCR Detection of Rifampin Resistance-Associated Mutations
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lowy, F.D. Staphylococcus aureus infections. N. Engl. J. Med. 1998, 339, 520–532. [Google Scholar] [CrossRef] [PubMed]
- Guérillot, R.; da Silva, A.G.; Monk, I.; Giulieri, S.; Tomita, T.; Alison, E.; Porter, J.; Pidot, S.; Gao, W.; Peleg, A.Y.; et al. Convergent evolution driven by rifampin exacerbates the global burden of drug-resistant Staphylococcus aureus. mSphere 2018, 3, e00550-17. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Bayer, A.; Cosgrove, S.E.; Daum, R.S.; Fridkin, S.K.; Gorwitz, R.J.; Kaplan, S.L.; Karchmer, A.W.; Levine, D.P.; Murray, B.E.; et al. Clinical practice guidelines by the infectious diseases society of america for the treatment of methicillin-resistant Staphylococcus aureus infections in adults and children. Clin. Infect. Dis. 2011, 52, e18–e55. [Google Scholar] [CrossRef] [PubMed]
- Osmon, D.R.; Berbari, E.F.; Berendt, A.R.; Lew, D.; Zimmerli, W.; Steckelberg, J.M.; Rao, N.; Hanssen, A.; Wilson, W.R. Diagnosis and management of prosthetic joint infection: Clinical practice guidelines by the Infectious Diseases Society of America. Clin. Infect. Dis. 2013, 56, e1–e25. [Google Scholar] [CrossRef] [PubMed]
- Habib, G.; Lancellotti, P.; Antunes, M.J.; Bongiorni, M.G.; Casalta, J.P.; Del Zotti, F.; Dulgheru, R.; El Khoury, G.; Erba, P.A.; Iung, B.; et al. 2015 ESC Guidelines for the management of infective endocarditis: The Task Force for the Management of Infective Endocarditis of the European Society of Cardiology (ESC). Endorsed by: European Association for Cardio-Thoracic Surgery (EACTS), the European Association of Nuclear Medicine (EANM). Eur. Heart J. 2015, 36, 3075–3128. [Google Scholar] [CrossRef] [PubMed]
- Aubry-Damon, H.; Soussy, C.J.; Courvalin, P. Characterization of mutations in the rpoB gene that confer rifampin resistance in Staphylococcus aureus. Antimicrob. Agents Chemother. 1998, 42, 2590–2594. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Shan, W.; Ma, X.; Chang, W.; Zhou, X.; Lu, H.; Dai, Y. Molecular characterization of rifampicin-resistant Staphylococcus aureus isolates in a Chinese teaching hospital from Anhui, China. BMC Microbiol. 2012, 12, 240. [Google Scholar] [CrossRef] [PubMed]
- Bongiorno, D.; Mongelli, G.; Stefani, S.; Campanile, F. Burden of rifampicin- and methicillin-resistant Staphylococcus aureus in Italy. Microb. Drug Resist. 2018, 24, 732–738. [Google Scholar] [CrossRef]
- Guo, Y.; Wang, B.; Rao, L.; Wang, X.; Zhao, H.; Li, M.; Yu, F. Molecular characteristics of rifampin-sensitive and -resistant isolates and characteristics of rpoB gene mutations in methicillin-resistant Staphylococcus aureus. Infect. Drug Resist. 2021, 14, 4591–4600. [Google Scholar] [CrossRef]
- Vazquez-Rosas, G.J.; Merida-Vieyra, J.; Aparicio-Ozores, G.; Lara-Hernandez, A.; De Colsa, A.; Aquino-Andrade, A. Molecular characterization of Staphylococcus aureus obtained from blood cultures of paediatric patients treated in a tertiary care hospital in Mexico. Infect. Drug Resist. 2021, 14, 1545–1556. [Google Scholar] [CrossRef]
- Goudarzi, M.; Seyedjavadi, S.S.; Nasiri, M.J.; Goudarzi, H.; Nia, R.S.; Dabiri, H. Molecular characteristics of methicillin-resistant Staphylococcus aureus (MRSA) strains isolated from patients with bacteremia based on MLST, SCCmec, spa, and agr locus types analysis. Microb. Pathog. 2017, 104, 328–335. [Google Scholar] [CrossRef] [PubMed]
- Tekeli, A.; Ocal, D.N.; Ozmen, B.B.; Karahan, Z.C.; Dolapci, I. Molecular characterization of methicillin-resistant Staphylococcus aureus bloodstream isolates in a Turkish university hospital between 2002 and 2012. Microb. Drug Resist. 2016, 22, 564–569. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Kim, S.S.; Choi, S.M.; Bae, C.J.; Oh, T.H.; Kim, S.E.; Kim, U.J.; Kang, S.J.; Jung, S.I.; Park, K.H. Rifamycin resistance, rpoB gene mutation and clinical outcomes of Staphylococcus species isolates from prosthetic joint infections in Republic of Korea. J. Glob. Antimicrob. Resist. 2022, 28, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.; Wang, X.; Wang, X.; Du, B.; Xu, K.; Zhang, F.; Jiang, C.; Zhao, Y.; Zhu, Y. Molecular characterization and virulence gene profiling of methicillin-resistant Staphylococcus aureus associated with bloodstream infections in southern China. Front. Microbiol. 2022, 13, 1008052. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lin, D.; Huang, Z.; Zhang, J.; Xie, W.; Liu, P.; Jing, H.; Wang, J. Clonality, virulence genes, and antibiotic resistance of Staphylococcus aureus isolated from blood in Shandong, China. BMC Microbiol. 2021, 21, 281. [Google Scholar] [CrossRef] [PubMed]
- Gu, F.; He, W.; Xiao, S.; Wang, S.; Li, X.; Zeng, Q.; Ni, Y.; Han, L. Antimicrobial resistance and molecular epidemiology of Staphylococcus aureus causing bloodstream infections at Ruijin Hospital in Shanghai from 2013 to 2018. Sci. Rep. 2020, 10, 6019. [Google Scholar] [CrossRef] [PubMed]
- Alfouzan, W.; Udo, E.E.; Modhaffer, A.; Alosaimi, A. Molecular characterization of methicillin- resistant Staphylococcus aureus in a tertiary care hospital in Kuwait. Sci. Rep. 2019, 9, 18527. [Google Scholar] [CrossRef]
- Soo Ko, K.; Kim, Y.S.; Song, J.H.; Yeom, J.S.; Lee, H.; Jung, S.I.; Jeong, D.R.; Kim, S.W.; Chang, H.H.; Ki, H.K.; et al. Genotypic diversity of methicillin-resistant Staphylococcus aureus isolates in Korean hospitals. Antimicrob. Agents Chemother. 2005, 49, 3583–3585. [Google Scholar] [CrossRef]
- Cha, H.Y.; Moon, D.C.; Choi, C.H.; Oh, J.Y.; Jeong, Y.S.; Lee, Y.C.; Seol, S.Y.; Cho, D.T.; Chang, H.H.; Kim, S.W.; et al. Prevalence of the ST239 clone of methicillin-resistant Staphylococcus aureus and differences in antimicrobial susceptibilities of ST239 and ST5 clones identified in a Korean hospital. J. Clin. Microbiol. 2005, 43, 3610–3614. [Google Scholar] [CrossRef]
- Kwon, J.C.; Kim, S.H.; Park, S.H.; Choi, S.M.; Lee, D.G.; Choi, J.H.; Park, C.; Shin, N.Y.; Yoo, J.H. Molecular epidemiologic analysis of methicillin-resistant Staphylococcus aureus isolates from bacteremia and nasal colonization at 10 intensive care units: Multicenter prospective study in Korea. J. Korean Med. Sci. 2011, 26, 604–611. [Google Scholar] [CrossRef]
- Park, S.H.; Park, C.; Yoo, J.H.; Choi, S.M.; Choi, J.H.; Shin, H.H.; Lee, D.G.; Lee, S.; Kim, J.; Choi, S.E.; et al. Emergence of community-associated methicillin-resistant Staphylococcus aureus strains as a cause of healthcare-associated bloodstream infections in Korea. Infect. Control Hosp. Epidemiol. 2009, 30, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.Y.; Lee, H.J.; Lee, M.S. Molecular characteristics of methicillin-resistant Staphylococcus aureus blood isolates: Clonal spread of staphylococcal cassette chromosome mec type IVA between the community and the hospital. Microb. Drug Resist. 2010, 16, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Fang, R.; Wang, C.; Tian, X.; Lin, J.; Zeng, W.; Zhou, T.; Xu, C. Resistance profiles and biological characteristics of rifampicin-resistant Staphylococcus aureus small-colony variants. Infect. Drug Resist. 2021, 14, 1527–1536. [Google Scholar] [CrossRef] [PubMed]
- Qazi, O.; Rahman, H.; Tahir, Z.; Qasim, M.; Khan, S.; Ahmad Anjum, A.; Yaqub, T.; Tayyab, M.; Ali, N.; Firyal, S. Mutation pattern in rifampicin resistance determining region of rpoB gene in multidrug-resistant Mycobacterium tuberculosis isolates from Pakistan. Int. J. Mycobacteriol. 2014, 3, 173–177. [Google Scholar] [CrossRef] [PubMed]
- Adikaram, C.P.; Perera, J.; Wijesundera, S.S. Geographical profile of rpoB gene mutations in rifampicin resistant Mycobacterium tuberculosis isolates in Sri Lanka. Microb. Drug Resist. 2012, 18, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Frénay, H.M.; Bunschoten, A.E.; Schouls, L.M.; van Leeuwen, W.J.; Vandenbroucke-Grauls, C.M.; Verhoef, J.; Mooi, F.R. Molecular typing of methicillin-resistant Staphylococcus aureus on the basis of protein A gene polymorphism. Eur. J. Clin. Microbiol. Infect. Dis. 1996, 15, 60–64. [Google Scholar] [CrossRef] [PubMed]
- Heep, M.; Brandstätter, B.; Rieger, U.; Lehn, N.; Richter, E.; Rüsch-Gerdes, S.; Niemann, S. Frequency of rpoB mutations inside and outside the cluster I region in rifampin-resistant clinical Mycobacterium tuberculosis isolates. J. Clin. Microbiol. 2001, 39, 107–110. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.P.; Crawford, J.T.; Shinnick, T.M. The rpoB gene of Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 1994, 38, 805–811. [Google Scholar] [CrossRef]
- Kapur, V.; Li, L.L.; Iordanescu, S.; Hamrick, M.R.; Wanger, A.; Kreiswirth, B.N.; Musser, J.M. Characterization by automated DNA sequencing of mutations in the gene (rpoB) encoding the RNA polymerase beta subunit in rifampin-resistant Mycobacterium tuberculosis strains from New York City and Texas. J. Clin. Microbiol. 1994, 32, 1095–1098. [Google Scholar] [CrossRef]
- Musser, J.M. Antimicrobial agent resistance in mycobacteria: Molecular genetic insights. Clin. Microbiol. Rev. 1995, 8, 496–514. [Google Scholar] [CrossRef]
- Chuang, Y.Y.; Huang, Y.C. Molecular epidemiology of community-associated meticillin-resistant Staphylococcus aureus in Asia. Lancet Infect. Dis. 2013, 13, 698–708. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, A.J.; Huovinen, T.; Fishwick, C.W.; Chopra, I. Molecular genetic and structural modeling studies of Staphylococcus aureus RNA polymerase and the fitness of rifampin resistance genotypes in relation to clinical prevalence. Antimicrob. Agents Chemother. 2006, 50, 298–309. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, Y.; Cui, L.; Katayama, Y.; Kozue, K.; Hiramatsu, K. Impact of rpoB mutations on reduced vancomycin susceptibility in Staphylococcus aureus. J. Clin. Microbiol. 2011, 49, 2680–2684. [Google Scholar] [CrossRef] [PubMed]
- Bæk, K.T.; Thøgersen, L.; Mogenssen, R.G.; Mellergaard, M.; Thomsen, L.E.; Petersen, A.; Skov, S.; Cameron, D.R.; Peleg, A.Y.; Frees, D. Stepwise decrease in daptomycin susceptibility in clinical Staphylococcus aureus isolates associated with an initial mutation in rpoB and a compensatory inactivation of the clpX gene. Antimicrob. Agents Chemother. 2015, 59, 6983–6991. [Google Scholar] [CrossRef]
- Howden, B.P.; Davies, J.K.; Johnson, P.D.; Stinear, T.P.; Grayson, M.L. Reduced vancomycin susceptibility in Staphylococcus aureus, including vancomycin-intermediate and heterogeneous vancomycin-intermediate strains: Resistance mechanisms, laboratory detection, and clinical implications. Clin. Microbiol. Rev. 2010, 23, 99–139. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.T.; Liao, C.H.; Chen, S.Y.; Yang, C.J.; Hsu, H.S.; Teng, L.J.; Hsueh, P.R. Characterization of rifampin-resistant Staphylococcus aureus nasal carriage in patients receiving rifampin-containing regimens for tuberculosis. Infect. Drug Resist. 2018, 11, 1175–1182. [Google Scholar] [CrossRef] [PubMed]
- Traber, K.E.; Lee, E.; Benson, S.; Corrigan, R.; Cantera, M.; Shopsin, B.; Novick, R.P. agr function in clinical Staphylococcus aureus isolates. Microbiology 2008, 154, 2265–2274. [Google Scholar] [CrossRef]
- Wayne, P.A. Performance Standards for Antimicrobial Susceptibility Testing: Twenty-Six Informational Supplement M100-S26; Clinical and Laboratory Standard Institute: Wayne, PA, USA, 2016. [Google Scholar]
- Enright, M.C.; Day, N.P.; Davies, C.E.; Peacock, S.J.; Spratt, B.G. Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones of Staphylococcus aureus. J. Clin. Microbiol. 2000, 38, 1008–1015. [Google Scholar] [CrossRef]
- Koreen, L.; Ramaswamy, S.V.; Graviss, E.A.; Naidich, S.; Musser, J.M.; Kreiswirth, B.N. spa typing method for discriminating among Staphylococcus aureus isolates: Implications for use of a single marker to detect genetic micro- and macrovariation. J. Clin. Microbiol. 2004, 42, 792–799. [Google Scholar] [CrossRef]
- Zhang, K.; McClure, J.A.; Elsayed, S.; Louie, T.; Conly, J.M. Novel multiplex PCR assay for characterization and concomitant subtyping of staphylococcal cassette chromosome mec types I to V in methicillin-resistant Staphylococcus aureus. J. Clin. Microbiol. 2005, 43, 5026–5033. [Google Scholar] [CrossRef]
- Harmsen, D.; Claus, H.; Witte, W.; Rothgänger, J.; Claus, H.; Turnwald, D.; Vogel, U. Typing of methicillin-resistant Staphylococcus aureus in a university hospital setting by using novel software for spa repeat determination and database management. J. Clin. Microbiol. 2003, 41, 5442–5448. [Google Scholar] [CrossRef]
- Mick, V.; Domínguez, M.A.; Tubau, F.; Liñares, J.; Pujol, M.; Martín, R. Molecular characterization of resistance to Rifampicin in an emerging hospital-associated Methicillin-resistant Staphylococcus aureus clone ST228, Spain. BMC Microbiol. 2010, 10, 68. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| MRSA rpoB Mutations | Rifampicin MIC | ||
|---|---|---|---|
| Nucleotide Mutation | Amino Acid Substitution | MIC μg/mL | Number of Isolates |
| gct/gat | A477D | >128 | 17 |
| cat/tat | H481Y | >128 | 4 |
| agc/aac + tct/cct | S463N, S464P | >128 | 2 |
| caa/cta | Q468L | >128 | 2 |
| gct/gat + act/gct | A477D, T801A | >128 | 2 |
| cgt/cat | R484H | >128 | 1 |
| tct/cct + att/ctt | S464P, I527L | >128 | 1 |
| caa/cga | Q468R | >128 | 1 |
| cat/aat + att/atg | H481N, I527M | >128 | 1 |
| cat/aat + att/atg + gaa/aaa | H481N, I527M, E568K | >128 | 1 |
| cat/aat + tca/tta | H481N, S529L | >128 | 1 |
| cat/cgt | H481R | >128 | 1 |
| gct/act + cat/aat | A477T, H481N | >128 | 1 |
| tca/tta | S486L | >128 | 1 |
| gac/ggc | D471G | >128 | 1 |
| gac/gaa + gct/gat | D471E, A477D | >128 | 1 |
| caa/cta + gat/gaa | Q468L, D668E | >128 | 1 |
| caa/cta + aaa/ata | Q468L, K1166I | >128 | 1 |
| att/ctt + gct/gat | I448L, A477D | >128 | 1 |
| gaa/aaa | E568K | >128 | 1 |
| agc/aac + tct/cct + aaa/ata | S463N, S464P, K1584I | 128 | 1 |
| cgt/ctt | R197L | 128 | 1 |
| cta/ata + acg/aag | L485I, T480K | 64 | 1 |
| tct/cct | S464P | 32 | 2 |
| att/cat | I527H | 32 | 1 |
| MLST | SCCmec | Spa Type | Number of Isolates | ||||
|---|---|---|---|---|---|---|---|
| Ic | II | IIa | IIb | IVa | |||
| ST5 | 1 | 25 | t2460 | 26 | |||
| 2 | 2 | t002 | 4 | ||||
| 3 | t9353 | 3 | |||||
| 2 | t324 | 2 | |||||
| 1 | t1228 | 1 | |||||
| 1 | t2461 | 1 | |||||
| 1 | t9363 | 1 | |||||
| 1 | t264 | 1 | |||||
| 1 | t564 | 1 | |||||
| 1 | t18239 | 1 | |||||
| 2 | 1 | unknown | 3 | ||||
| ST72 | 1 | t324 | 1 | ||||
| 1 | t2431 | 1 | |||||
| 1 | t664 | 1 | |||||
| 1 | t148 | 1 | |||||
| 1 | unknown | 1 | |||||
| ST254 | 1 | t324 | 1 | ||||
| 1 | t688 | 1 | |||||
| ST1 | 1 | t2460 | 1 | ||||
| Genotype | Number (%) of Isolates | |
|---|---|---|
| Rifampin Resistance (n = 57) | Rifampin Susceptible (n = 1558) | |
| n = 1615 | ||
| MRSA | 52 (91.2) * | 791 (48.9) |
| MSSA | 5 (8.8) * | 767 (47.5) |
| n = 1615 | ||
| agr function | 14 (24.6) * | 835 (53.6) |
| agr dysfunction | 43 (75.4) * | 723 (46.4) |
| n =1615 | ||
| ST5 | 44 (77.2) * | 499 (32.0) |
| Non-ST5 | 13 (22.8) * | 1059 (68.0) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.K.; Eom, Y.; Kim, E.; Chang, E.; Bae, S.; Jung, J.; Kim, M.J.; Chong, Y.P.; Kim, S.-H.; Choi, S.-H.; et al. Molecular Characteristics and Prevalence of Rifampin Resistance in Staphylococcus aureus Isolates from Patients with Bacteremia in South Korea. Antibiotics 2023, 12, 1511. https://doi.org/10.3390/antibiotics12101511
Kim YK, Eom Y, Kim E, Chang E, Bae S, Jung J, Kim MJ, Chong YP, Kim S-H, Choi S-H, et al. Molecular Characteristics and Prevalence of Rifampin Resistance in Staphylococcus aureus Isolates from Patients with Bacteremia in South Korea. Antibiotics. 2023; 12(10):1511. https://doi.org/10.3390/antibiotics12101511
Chicago/Turabian StyleKim, Yong Kyun, Yewon Eom, Eunsil Kim, Euijin Chang, Seongman Bae, Jiwon Jung, Min Jae Kim, Yong Pil Chong, Sung-Han Kim, Sang-Ho Choi, and et al. 2023. "Molecular Characteristics and Prevalence of Rifampin Resistance in Staphylococcus aureus Isolates from Patients with Bacteremia in South Korea" Antibiotics 12, no. 10: 1511. https://doi.org/10.3390/antibiotics12101511
APA StyleKim, Y. K., Eom, Y., Kim, E., Chang, E., Bae, S., Jung, J., Kim, M. J., Chong, Y. P., Kim, S.-H., Choi, S.-H., Lee, S.-O., & Kim, Y. S. (2023). Molecular Characteristics and Prevalence of Rifampin Resistance in Staphylococcus aureus Isolates from Patients with Bacteremia in South Korea. Antibiotics, 12(10), 1511. https://doi.org/10.3390/antibiotics12101511