
National Surveillance of Tetracycline, Erythromycin, and Clindamycin Resistance in Invasive Streptococcus pyogenes: A Retrospective Study of the Situation in Spain, 2007–2020

 , , and
, , and
Abstract
1. Introduction
2. Results
2.1. Clinical and Epidemiological Data for Resistant S. pyogenes Isolates
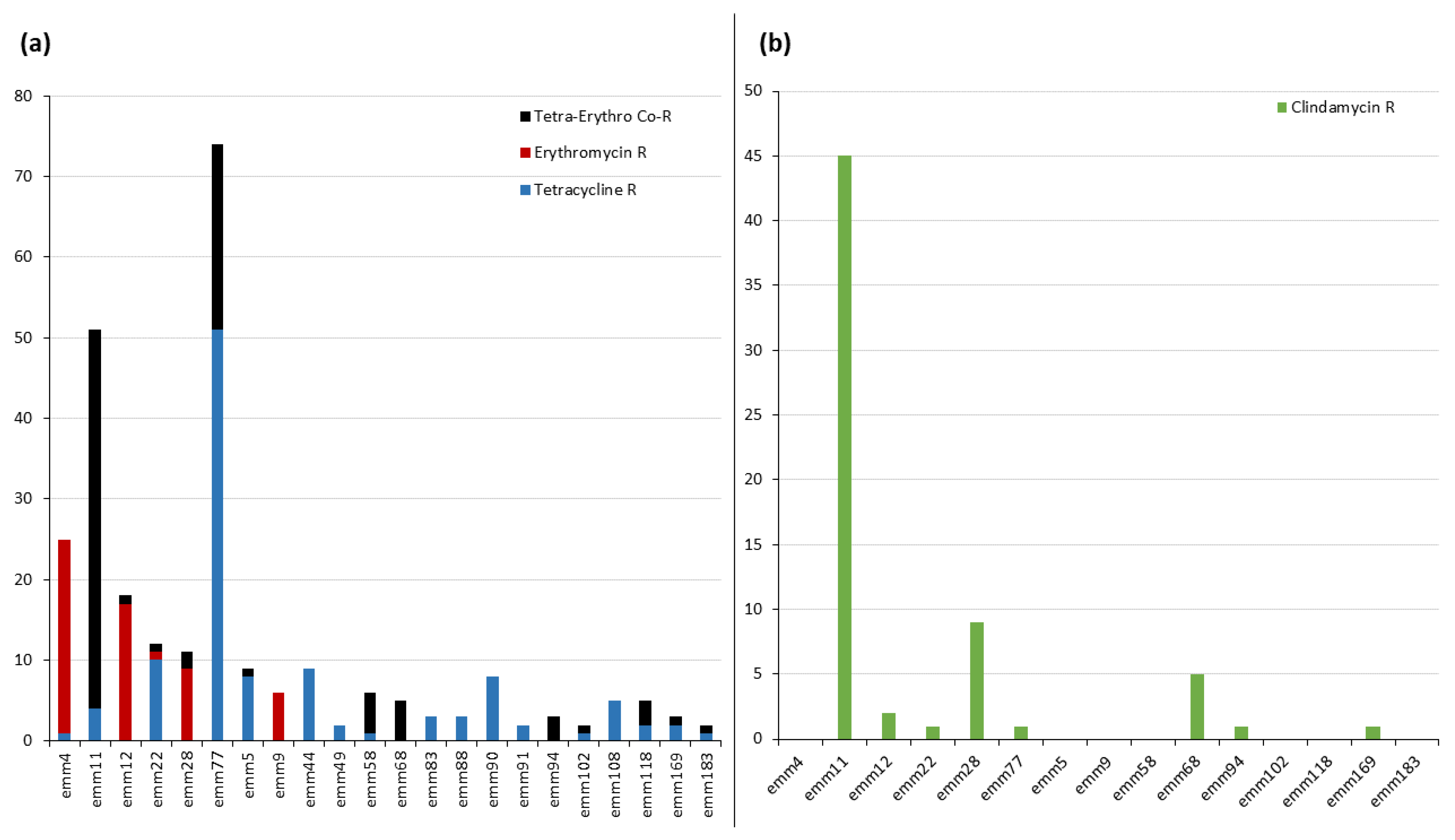
2.2. emm Types Resistant to Tetracycline, Erythromycin, and Clindamycin
2.3. Tetracycline and Erythromycin Resistance: Genotype Analysis
2.4. Macrolide and Lincosamide Resistance: Phenotype Analysis
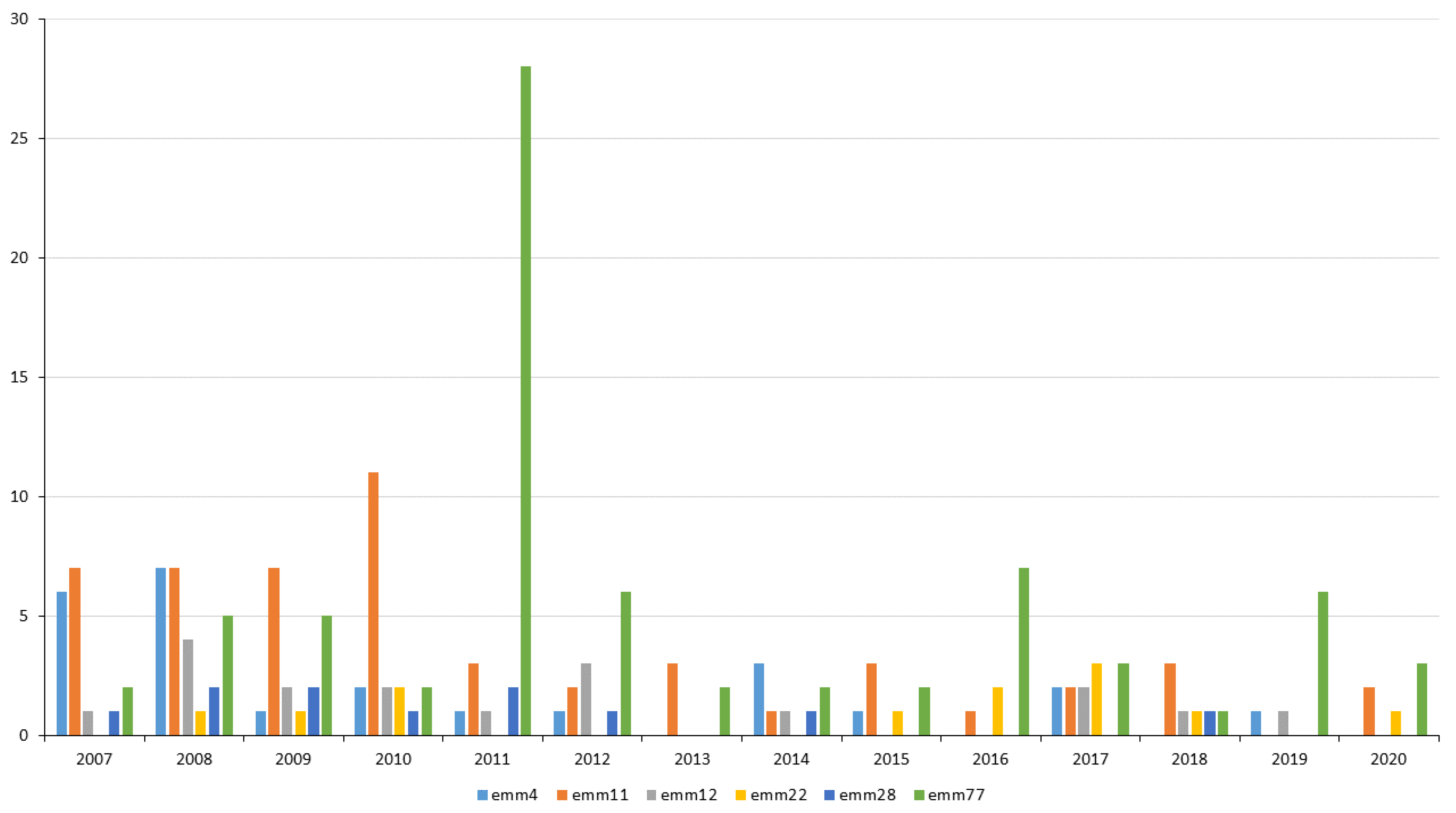
2.5. Changes in Resistance in 2007–2020
2.6. Resistant Clones
3. Discussion
4. Materials and Methods
4.1. Basic Microbiological Typing
4.2. Tetracycline, Erythromycin, and Clindamycin Resistance: Phenotype and Genotype Analyses
4.3. Multilocus Sequence Typing and Description of Resistant Clones
4.4. Statistical Analysis
4.5. Ethical Approval
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carapetis, J.R.; Steer, A.C.; Mulholland, E.K.; Weber, M. The global burden of group A streptococcal diseases. Lancet Infect. Dis. 2005, 5, 685–694. [Google Scholar] [CrossRef]
- Walker, M.J.; Barnett, T.C.; McArthur, J.D.; Cole, J.N.; Gillen, C.M.; Henningham, A.; Sriprakash, K.S.; Sanderson-Smith, M.L.; Nizet, V. Disease manifestations and pathogenic mechanisms of group A Streptococcus. Clin. Microbiol. Rev. 2014, 27, 264–301. [Google Scholar] [CrossRef]
- Bisno, A.L.; Brito, M.O.; Collins, C.M. Molecular basis of group A streptococcal virulence. Lancet Infect. Dis. 2003, 3, 191–200. [Google Scholar] [CrossRef]
- Center for Disease Control and Prevention (CDC). Group A Streptococcal (GAS) Disease. Available online: https://www.cdc.gov/groupastrep/diseases-hcp/index.html (accessed on 28 December 2022).
- Johnson, A.F.; LaRock, C.N. Antibiotic treatment, mechanisms for failure, and adjunctive therapies for infections by group A Streptococcus. Front. Microbiol. 2021, 12, 760255. [Google Scholar] [CrossRef]
- Varaldo, P.E.; Montanari, M.P.; Giovanetti, E. Genetic elements responsible for erythromycin resistance in streptococci. Antimicrob. Agents Chemother. 2009, 53, 343–353. [Google Scholar] [CrossRef]
- Brenciani, A.; Bacciaglia, A.; Vecchi, M.; Vitali, L.A.; Varaldo, P.E.; Giovanetti, E. Genetic elements carrying erm(B) in Streptococcus pyogenes and association with tet(M) tetracycline resistance gene. Antimicrob. Agent. Chemother. 2007, 51, 1209–1216. [Google Scholar] [CrossRef]
- Brenciani, A.; Tiberi, E.; Bacciaglia, A.; Petrelli, D.; Varaldo, P.E.; Giovanetti, E. Two distinct genetic elements are responsible for erm(TR)-mediated erythromycin resistance in tetracycline-susceptible and tetracycline-resistant strains of Streptococcus pyogenes. Antimicrob. Agent. Chemother. 2011, 55, 2106–2112. [Google Scholar] [CrossRef]
- Berbel, D.; Càmara, J.; González-Díaz, A.; Cubero, M.; López de Egea, G.; Martí, S.; Tubau, F.; Domínguez, A.; Ardanuy, C. Deciphering mobile genetic elements disseminating macrolide resistance in Streptococcus pyogenes over a 21 year period in Barcelona, Spain. J. Antimicrob. Chemother. 2021, 76, 1991–2003. [Google Scholar] [CrossRef]
- Pérez-Trallero, E.; Montes, M.; Orden, B.; Tamayo, E.; García-Arenzana, J.M.; Marimón, J.M. Phenotypic and genotypic characterization of Streptococcus pyogenes isolates displaying the MLSB phenotype of macrolide resistance in Spain, 1999 to 2005. Antimicrob. Agent. Chemother. 2007, 51, 1228–1233. [Google Scholar] [CrossRef]
- Ardanuy, C.; Domenech, A.; Rolo, D.; Calatayud, L.; Tubau, F.; Ayats, J.; Martín, R.; Liñares, J. Molecular characterization of macrolide- and multidrug-resistant Streptococcus pyogenes isolated from adult patients in Barcelona, Spain (1993–2008). J. Antimicrob. Chemother. 2010, 65, 634–643. [Google Scholar] [CrossRef]
- Rubio-López, V.; Valdezate, S.; Álvarez, D.; Villalón, P.; Medina, M.J.; Salcedo, C.; Sáez-Nieto, J.-A. Molecular epidemiology, antimicrobial susceptibilities and resistance mechanisms of Streptococcus pyogenes isolates resistant to erythromycin and tetracycline in Spain (1994-2006). BMC Microbiol. 2012, 12, 215. [Google Scholar] [CrossRef]
- Montes, M.; Ardanuy, C.; Tamayo, E.; Domènech, A.; Liñares, J.; Pérez-Trallero, E. Epidemiological and molecular analysis of Streptococcus pyogenes isolates causing invasive disease in Spain (1998-2009): Comparison with non invasive isolates. Eur. J. Clin. Microbiol. Infect. Dis. 2011, 30, 1295–1302. [Google Scholar] [CrossRef]
- Leclerq, R. Mechanisms of resistance to macrolides and lincosamides: Nature of the resistance elements and their clinical implications. Clin. Infect. Dis. 2002, 34, 482–492. [Google Scholar] [CrossRef]
- Roberts, M.C. Tetracycline and MLS Nomenclature. 2022. Available online: http://faculty.washington.edu/marilynr/ (accessed on 8 November 2022).
- Seppälä, H.; Skurnik, M.; Soini, H.; 5, M.C.; Huovinen, P. A novel erythromycin resistance methylase gene (ermTR) in Streptococcus pyogenes. Antimicrob. Agents Chemother. 1998, 42, 257–262. [Google Scholar] [CrossRef]
- Woodbury, R.L.; Klammer, K.A.; Xiong, Y.; Bailiff, T.; Glennen, A.; Bartkus, J.M.; Lynfield, R.; Van Beneden, C.; Beall, B.W. Plasmid-borne erm(T) from invasive, macrolide-resistant Streptococcus pyogenes strains. Antimicrob. Agents Chemother. 2008, 52, 1140–1143. [Google Scholar] [CrossRef]
- Instituto de Salud Carlos III. Centro Nacional de Microbiología. Área de Orientación Diagnóstica. Programas de Vigilancia de Bacterias. Vigilancia de la Infección Estreptocócica. Available online: https://www.12.es/QueHacemos/Servicios/DiagnosticoMicrobiol%C3%B3gicoyProgramasVigilancia/Paginas/default.aspx (accessed on 15 December 2022).
- Villalón, P.; Sáez-Nieto, J.A.; Rubio-López, V.; Medina-Pascual, M.J.; Garrido, N.; Carrasco, G.; Pino-Rosa, S.; Valdezate, S. Invasive Streptococcus pyogenes disease in Spain: A microbiological and epidemiological study covering the period 2007–2019. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 2295–2303. [Google Scholar] [CrossRef]
- The European Committee of Antimicrobial Susceptibility Testing. Clinical Breakpoints–Bacteria (V12.0). 2022. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Breakpoint_tables/v_12.0_Breakpoint_Tables.pdf (accessed on 8 November 2022).
- European Centre for Disease Prevention and Control (ECDC). Available online: https://www.ecdc.europa.eu/en/publications-data/directory-online-resources-prevention-and-control-antimicrobial-resistance-amr (accessed on 28 December 2022).
- Plan Nacional de Resistencia a Antibióticos (PRAN). Available online: https://www.resistenciaantibioticos.es/es (accessed on 28 December 2022).
- Vannice, K.S.; Ricaldi, J.; Nanduri, S.; Fang, F.C.; Lynch, J.B.; Bryson-Cahn, C.; Wright, T.; Duchin, J.; Kay, M.; Chochua, S.; et al. Streptococcus pyogenes pbp2x mutation confers reduced susceptibilit to β–lactam antibiotics. Clin. Infect. Dis. 2019, 71, 201–204. [Google Scholar] [CrossRef]
- Montes, M.; Tamayo, E.; Mojica, C.; García-Arenzana, J.M.; Esnal, O.; Pérez-Trallero, E. What causes erythromycin resistance in Streptococcus pyogenes? Dynamics of four clones in a southern European región from 2005 to 2012. J. Antimicrob. Chemother. 2014, 69, 1474–1482. [Google Scholar] [CrossRef]
- Giovanetti, E.; Montanari, M.P.; Mingoia, M.; Varaldo, P.E. Phenotypes and genotypes of erythromycin-resistant Streptococcus pyogenes strains in Italy and heterogeneity of inducibly resistant strains. Antimicrob. Agents Chemother. 1999, 43, 1935–1940. [Google Scholar] [CrossRef][Green Version]
- Willems, R.J.L.; Hanage, W.P.; Bessen, D.E.; Feil, E.J. Population biology of Gram-positive Pathogens: High-risk clones for dissemination of antibiotic resistance. FEMS Microbiol. Rev. 2011, 35, 872–900. [Google Scholar] [CrossRef]
- Center for Disease Control and Prevention. Streptococcus Laboratory. 2022. Available online: https://www.cdc.gov/streplab/groupa-strep/index.html (accessed on 8 November 2022).
- Schmitz, F.J.; Beyer, A.; Charpentier, E.; Henriques Normark, B.; Schade, M.; Fluit, A.C.; Hafner, D.; Novak, R. Toxin-gene profile heterogeneity among endemic invasive European group A streptococcal isolates. J. Infect. Dis. 2003, 188, 1578–1586. [Google Scholar] [CrossRef]
- Poyart, C.; Jardy, L.; Quesne, G.; Berche, P.; Trieu-Cuot, P. Genetic basis of antibiotic resistance in Streptococcus agalactiae strains isolated in a French hospital. Antimicrob. Agents Chemother. 2003, 47, 794–797. [Google Scholar] [CrossRef]
- Malhotra-Kumar, S.; Lammens, C.; Piessens, J.; Goossens, H. Multiplex PCR for simultaneous detection of macrolide and tetracycline resistance determinants in streptococci. Antimicrob. Agents Chemother. 2005, 49, 4798–4800. [Google Scholar] [CrossRef]
- Luthje, P.; Schwarz, S. Molecular basis of resistance to macrolides and lincosamides among staphylococci and streptococci from various animal sources collected in the resistance monitoring program BfT-GermVet. Int. J. Antimicrob. Agents 2007, 29, 528–535. [Google Scholar] [CrossRef]
- Sutcliffe, J.; Grebe, T.; Tait-Kamradt, A.; Wondrack, L. Detection of erythromycin-resistant determinants by PCR. Antimicrob. Agents Chemother. 1996, 40, 2562–2566. [Google Scholar] [CrossRef]
- Jolley, K.A.; Brain, J.E.; Maiden, M.C.J. Open-access bacterial population genomics: BIGSdb software, the PubMLST.org website. Wellcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef]
- Enright, M.C.; Spratt, B.G.; Kalia, A.; Cross, J.H.; Bessen, D. Multilocus sequence typing of Streptococcus pyogenes and the relationships betweem emm type and clone. Infect. Immun. 2001, 69, 2416–2427. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type a | N b | Tetracycline c | Erythromycin c | Clindamycin c | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| n | % | MIC50 | MIC90 | n | % | MIC50 | MIC90 | n | % | MIC50 | MIC90 | ||
| emm | |||||||||||||
| Total | 1983 | 237 | 12.0 | 0.25 | 12 | 172 | 8.7 | 0.125 | 0.25 | 78 | 3.9 | 0.125 | 0.25 |
| emm4 | 161 | 1 | 0.6 | 0.25 | 0.5 | 24 | 14.9 | 0.125 | 16 | 0 | 0.0 | 0.125 | 0.25 |
| emm11 | 63 | 51 | 81.0 | 16 | 32 | 47 | 74.6 | >256 | >256 | 45 | 71.4 | >256 | >256 |
| emm12 | 115 | 1 | 0.86 | 0.25 | 0.5 | 18 | 15.7 | 0.19 | 16 | 2 | 1.7 | 0.125 | 0.25 |
| emm22 | 36 | 11 | 30.6 | 0.25 | 16 | 2 | 5.6 | 0.125 | 0.25 | 1 | 2.8 | 0.125 | 0.25 |
| emm28 | 97 | 2 | 2.1 | 0.25 | 0.5 | 11 | 11.3 | 0.125 | >256 | 9 | 9.2 | 0.125 | 0.5 |
| emm77 | 81 | 74 | 91.4 | 32 | 32 | 23 | 28.4 | 0.19 | 16 | 1 | 1.2 | 0.125 | 0.25 |
| Genotype | |||||||||||||
| tetM | - | 152 | - | 16 | 48 | - | - | - | - | - | - | - | - |
| tetO | - | 48 | - | 32 | 48 | - | - | - | - | - | - | - | - |
| tetM-tetO | - | 33 | - | 24 | 32 | - | - | - | - | - | - | - | - |
| mefA-msrD | - | - | - | - | - | 40 | - | 16 | 32 | 0 | 0 | 0.125 | 0.25 |
| ermB | - | - | - | - | - | 61 | - | >256 | >256 | 57 | - | >256 | >256 |
| ermTR | - | - | - | - | - | 24 | - | 12 | >256 | 1 | - | 0.125 | 0.25 |
| ermT | - | - | - | - | - | 1 | - | >256 | >256 | 0 | 0 | 0.125 | 0.125 |
| emm Type | No. Isolates a | MLST b | Genotype c | Phenotype d | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| ST | n | tetM | tetO | msrD | mefA | ermB | ermTR | ermT | |||
| Only tetracycline resistance | |||||||||||
| emm22 | 10/12 | 46 | 2 | + | - | ||||||
| emm77 | 41/74 | 63 | 3 | - | + | ||||||
| Only erythromycin resistance | |||||||||||
| emm4 | 19/25 | 39 | 5 | + | + | - | - | - | M | ||
| emm12 | 12/18 | 36 | 4 | + | + | - | - | - | M | ||
| emm28 | 6/11 | 52 | 2 | - | - | + | - | - | cMLSB | ||
| Tetracycline–erythromycin co-resistance | |||||||||||
| emm11 | 33/51 | 403 | 5 | + | - | - | - | + | - | - | cMLSB |
| emm77 | 8/74 | 63 | 2 | - | + | - | - | - | + | - | iMLSB |
| emm77 | 8/74 | 63 | 2 | + | + | - | - | - | + | - | iMLSB |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villalón, P.; Bárcena, M.; Medina-Pascual, M.J.; Garrido, N.; Pino-Rosa, S.; Carrasco, G.; Valdezate, S. National Surveillance of Tetracycline, Erythromycin, and Clindamycin Resistance in Invasive Streptococcus pyogenes: A Retrospective Study of the Situation in Spain, 2007–2020. Antibiotics 2023, 12, 99. https://doi.org/10.3390/antibiotics12010099
Villalón P, Bárcena M, Medina-Pascual MJ, Garrido N, Pino-Rosa S, Carrasco G, Valdezate S. National Surveillance of Tetracycline, Erythromycin, and Clindamycin Resistance in Invasive Streptococcus pyogenes: A Retrospective Study of the Situation in Spain, 2007–2020. Antibiotics. 2023; 12(1):99. https://doi.org/10.3390/antibiotics12010099
Chicago/Turabian StyleVillalón, Pilar, Marta Bárcena, María José Medina-Pascual, Noelia Garrido, Silvia Pino-Rosa, Gema Carrasco, and Sylvia Valdezate. 2023. "National Surveillance of Tetracycline, Erythromycin, and Clindamycin Resistance in Invasive Streptococcus pyogenes: A Retrospective Study of the Situation in Spain, 2007–2020" Antibiotics 12, no. 1: 99. https://doi.org/10.3390/antibiotics12010099
APA StyleVillalón, P., Bárcena, M., Medina-Pascual, M. J., Garrido, N., Pino-Rosa, S., Carrasco, G., & Valdezate, S. (2023). National Surveillance of Tetracycline, Erythromycin, and Clindamycin Resistance in Invasive Streptococcus pyogenes: A Retrospective Study of the Situation in Spain, 2007–2020. Antibiotics, 12(1), 99. https://doi.org/10.3390/antibiotics12010099