
Evolution and Emergence of Antibiotic Resistance in Given Ecosystems: Possible Strategies for Addressing the Challenge of Antibiotic Resistance

 ,
,  ,
,  ,
,  and
and
Abstract
1. Introduction
2. History of Antibiotics
3. Rise of Antimicrobial Resistance
4. What Caused These Organisms in the Environment to Develop Resistance to Multiple Drugs?

{kind=link}
{kind=link}
{kind=link}
| Antibiotic Resistance | Plasmid-Borne Genes | Resistant Organisms | References |
|---|---|---|---|
| Beta-lactams | blaIMP encoding imipenem resistance; blaVIM (Verona integron- encoded metallo-β-lactamases) | P. aeruginosa | [38,39] |
| blaOXA encoding oxacillin resistance | S. aureus | [40] | |
| blaNDM encoding metallo-β-lactamase | E. coli | [41] | |
| blaNDM-1 gene; blaOXA-23 | A. baumannii | [42] | |
| blaIMP-9, blaSIM-2, and blaVIM-2 | P. aeruginosa | [42,43] | |
| blaNDM, blaIMP, blaIMP-27, blaVIM, and blaKPC | Enterobacteriaceae | [44,45] | |
| blaNDM-1 | E. coli | [46] | |
| blaIMP | Metagenome | [39] | |
| blaOXA-23 | A. baumannii | [42] | |
| blaTEM; blaSHV, blaOXA | H. influenzae; E. coli; K. pneumoniae | [47,48] | |
| blaTEM | S. pneumoniae | [49] | |
| PBP2a | S. pneumoniae; E. coli | [50,51] | |
| CTX-M, OXA-30 | E. coli | [52] | |
| mecA, | S. aureus | [53] | |
| blaOXA-48 | Enterobacteriaceae | [54] | |
| Fluoroquinolones | qnrA, qnrB, qnrC, qnrD, qnrS, and aac(6′)-lb-cr | Campylobacter spp., Salmonella spp., and Shigella sp., K. pneumoniae, E. coli | [55,56] |
| gyrA | E. coli | [57,58] | |
| parC and parE | E. coli; K. pneumoniae | [57,59] | |
| NorC, NorA and MepA | S. aureus | [60] | |
| Rv1634 | Mycobacterium tuberculosis | [61] | |
| MfpA | Mycobacterium | [62] | |
| qnrS2 | Aeromonas | [63,64] | |
| QepA | E. coli | [65] | |
| OqxAB | E. coli | [66] | |
| parC and gyrA | S. pneumoniae | [67,68] | |
| SmeVWX | S. maltophilia | [69] | |
| Smqnr | S. maltophilia | [70] | |
| SmeDEF | Stenotrophomonas maltophilia | [71] | |
| pqsA | P. aeruginosa | [72] | |
| glpD, ygfA, and yigB | E. coli | [73] | |
| Glycopeptides | vanA, vanB, vanC, vanD, vanE, vanG, vanL, vanM, and vanN | Enterococci | [74,75,76] |
| vanRSHAX | S. aureus | [76] | |
| sarA | [77] | ||
| vanA and ermB | Enterococci | [78] | |
| Aminoglycosides | aac(3)-IV | E. coli | [79] |
| Polymyxins | mcr-1 | E. coli | [80] |
| Tetracyclines | tet genes | Streptomyces | [81] |
| Tn916 | B. subtilis | [82] | |
| Tet38 | S. aureus | [83] | |
| Lipopetides | pitA | S. aureus | [84] |
| Rifampicin | purB and purM | S. aureus | [85] |
| Cephalosporins | blaCTX-M, blaCMY | Kluyvera ascorbata; Kluyvera georgiana | [86] |
| blaCTX-M-1 and blaCMY-2 | E. coli | [87] | |
| Vancomycin | vanA, vanB, vanH, vanR, vanS, vanW, vanX, vanY, and vanZ | S. aureus | [88] |
| Multidrug resistance (MDR) | acrB | E. coli | [89] |
| SGI1 | S. enterica | [90] | |
| blaNDM-1 | P. aeruginosa | [91] |
5. Mechanisms of Antibiotic Resistance
6. Antibiotic Resistance in Different Environments
6.1. Aquatic Environments
6.1.1. Wastewater
Wastewater Treatment Plants
Hospital and Pharmaceutical Wastewater
6.1.2. Rivers and Groundwater
6.1.3. Marine System
6.1.4. Factors Affecting Antibiotic Resistance in the Aquatic Environment
6.2. Terrestrial Environments
6.2.1. Sludge Manure
6.2.2. Agricultural System
6.2.3. Manure from Livestock and Pesticides
6.2.4. Factors Affecting Antibiotic Persistence in the Terrestrial Environment
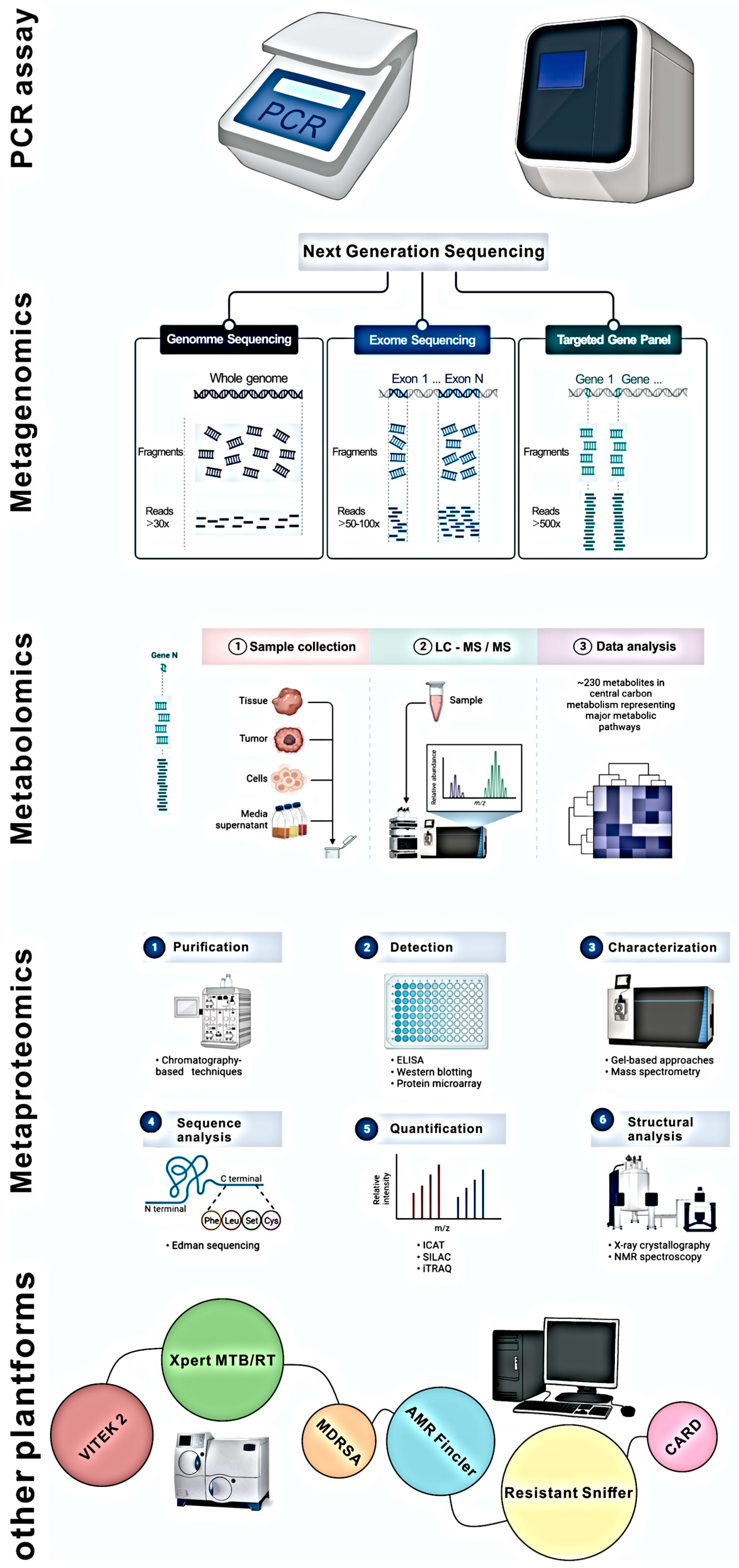
6.3. Tools Used for Antibiotic Resistance Studies
6.4. Ecotoxicological Impact of Antibiotics and Antibiotic-Resistant Bacteria
7. Antibiotic-Resistant Bacteria and Human Health Concerns
8. Strategies for Addressing the Challenge of Antibiotic Resistance
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kourkouta, L.; Tsaloglidou, A.; Koukourikos, K.; Iliadis, C.; Plati, P.; Dimitriadou, A. History of Antibiotics. Sumer. J. Med. Healthc. 2018, 1, 51–54. [Google Scholar] [CrossRef]
- Hutchings, M.I.; Truman, A.W.; Wilkinson, B. Antibiotics: Past, Present and Future. Curr. Opin. Microbiol. 2019, 51, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Zaffiri, L.; Gardner, J.; Toledo-Pereyra, L.H. History of Antibiotics. from Salvarsan to Cephalosporins. J. Investig. Surg. 2012, 25, 67–77. [Google Scholar] [CrossRef]
- Alnemri, A.R.; Almaghrabi, R.H.; Alonazi, N.; Alfrayh, A.R. Current Paediatric Research. Curr. Pediatr. Res. 2016, 20, 169–173. [Google Scholar]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-Resistant, Extensively Drug-Resistant and Pandrug-Resistant Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed]
- Bosch, F.; Rosich, L. The Contributions of Paul Ehrlich to Pharmacology: A Tribute on the Occasion of the Centenary of His Nobel Prize. Pharmacology 2008, 82, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Walsh, C.T.K.; Wright, G. Introduction: Antibiotic Resistance. Chem. Rev. 2005, 105, 391–394. [Google Scholar] [CrossRef]
- Derry, J.E. Evaluation of Cefaclor. Am. J. Hosp. Pharm. 1981, 38, 54–58. [Google Scholar] [CrossRef]
- Drehobl, M.; Bianchi, P.; Keyserling, C.H.; Tack, K.J.; Griffin, T.J. Comparison of Cefdinir and Cefaclor in Treatment of Community-Acquired Pneumonia. Antimicrob. Agents Chemother. 1997, 41, 1579–1583. [Google Scholar] [CrossRef][Green Version]
- Schaad, U.B.; Suter, S.; Gianella-Borradori, A.; Pfenninger, J.; Auckenthaler, R.; Bernath, O.; Cheseaux, J.-J.; Wedgwood, J. A Comparison of Ceftriaxone and Cefuroxime for the Treatment of Bacterial Meningitis in Children. N. Engl. J. Med. 1990, 322, 141–147. [Google Scholar] [CrossRef]
- Deal, E.N.; Micek, S.T.; Reichley, R.M.; Ritchie, D.J. Effects of an Alternative Cefepime Dosing Strategy in Pulmonary and Bloodstream Infections Caused by Enterobacter Spp, Citrobacter Freundii, and Pseudomonas Aeruginosa: A Single-Center, Open-Label, Prospective, Observational Study. Clin. Ther. 2009, 31, 299–310. [Google Scholar] [CrossRef] [PubMed]
- McNabb, J.; Quintiliani, R.; Nightingale, C.H.; Nicolau, D.P. Comparison of the Bactericidal Activity of Trovafloxacin and Ciprofloxacin, Alone and in Combination with Cefepime, against Pseudomonas Aeruginosa. Chemotherapy 2000, 46, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Endimiani, A.; Perez, F.; Bonomo, R.A. Cefepime: A Reappraisal in an Era of Increasing Antimicrobial Resistance. Expert Rev. Anti. Infect. Ther. 2008, 6, 805–824. [Google Scholar] [CrossRef] [PubMed]
- Conly, J.M.; Johnston, B.L. Where Are All the New Antibiotics? The New Antibiotic Paradox. Can. J. Infect. Dis. Med. Microbiol. 2005, 16, 159–160. [Google Scholar] [CrossRef] [PubMed]
- Zaman, S.B.; Hussain, M.A.; Nye, R.; Mehta, V.; Mamun, K.T.; Hossain, N. A Review on Antibiotic Resistance: Alarm Bells Are Ringing. Cureus 2017, 9, e1403. [Google Scholar] [CrossRef] [PubMed]
- Dodds, D.R. Antibiotic Resistance: A Current Epilogue. Biochem. Pharmacol. 2017, 134, 139–146. [Google Scholar] [CrossRef]
- Levy, S.B.; Marshall, B. Antibacterial Resistance Worldwide: Causes, Challenges and Responses. Nat. Med. 2004, 10, S122–S129. [Google Scholar] [CrossRef] [PubMed]
- Durand, G.A.; Raoult, D.; Dubourg, G. Antibiotic Discovery: History, Methods and Perspectives. Int. J. Antimicrob. Agents 2019, 53, 371–382. [Google Scholar] [CrossRef]
- Lowy, F.D. Antimicrobial Resistance: The Example of Staphylococcus Aureus. J. Clin. Investig. 2003, 111, 1265–1273. [Google Scholar] [CrossRef]
- Appelbaum, P.C. The Emergence of Vancomycin-intermediate and Vancomycin-resistant Staphylococcus Aureus. Clin. Microbiol. Infect. 2006, 12, 16–23. [Google Scholar] [CrossRef]
- Aminov, R.I.; Mackie, R.I. Evolution and Ecology of Antibiotic Resistance Genes. FEMS Microbiol. Lett. 2007, 271, 147–161. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control. Antimicrobial Resistance in the EU/EEA (EARS-Net)—Annual Epidemiological Report 2019; ECDC: Stockholm, Sweden, 2020; Available online: https://www.ecdc.europa.eu/en/publications-data/surveillance-antimicrobial-resistance-europe-2019#:~:text=EARS%2DNet%20data%20for%202019,aureus%20(20.6%25)%2C%20K (accessed on 1 November 2022).
- Indian Council of Medical Research. AMR and Surveillance Network Annual Report. January 2019 to December 2019; ICMR: New Delhi, India, 2020; Available online: https://main.icmr.nic.in/sites/default/files/guidelines/AMRSN_annual_report_2020.pdf (accessed on 14 October 2022).
- WHO. WHO Global Antimicrobial Resistance and Use Surveillance System (GLASS) Report: 2021; World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]
- Hu, F.; Zhu, D.; Wang, F.; Wang, M. Current Status and Trends of Antibacterial Resistance in China. Clin. Infect. Dis. 2018, 67, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Ndihokubwayo, J.B.; Yahaya, A.A.; Desta, A.T.; Ki-Zerbo, G.; Odei, E.A.; Keita, B.; Pana, A.P.; Nkhoma, W. Antimicrobial Resistance in the African Region: Issues, Challenges and Actions Proposed. African Health Monit. 2013, 16, 27–30. [Google Scholar]
- Tadesse, B.T.; Ashley, E.A.; Ongarello, S.; Havumaki, J.; Wijegoonewardena, M.; Gonzalez, I.J.; Dittrich, S. Antimicrobial Resistance in Africa: A Systematic Review. BMC Infect. Dis. 2017, 17, 616. [Google Scholar] [CrossRef] [PubMed]
- Uddin, T.M.; Chakraborty, A.J.; Khusro, A.; Zidan, B.M.R.M.; Mitra, S.; Emran, T.B.; Dhama, K.; Ripon, M.K.H.; Gajdács, M.; Sahibzada, M.U.K. Antibiotic Resistance in Microbes: History, Mechanisms, Therapeutic Strategies and Future Prospects. J. Infect. Public Health 2021, 14, 1750–1766. [Google Scholar] [CrossRef]
- Benveniste, R.; Davies, J. Aminoglycoside Antibiotic-Inactivating Enzymes in Actinomycetes Similar to Those Present in Clinical Isolates of Antibiotic-Resistant Bacteria. Proc. Natl. Acad. Sci. USA 1973, 70, 2276–2280. [Google Scholar] [CrossRef]
- Bennett, P.M. Plasmid Encoded Antibiotic Resistance: Acquisition and Transfer of Antibiotic Resistance Genes in Bacteria. Br. J. Pharmacol. 2008, 153, S347–S357. [Google Scholar] [CrossRef]
- Firth, N.; Skurray, R.A. Mobile Elements in the Evolution and Spread of Multiple-Drug Resistance in Staphylococci. Drug Resist. Updat. 1998, 1, 49–58. [Google Scholar] [CrossRef]
- Cloeckaert, A.; Schwarz, S. Molecular Characterization, Spread and Evolution of Multidrug Resistance in Salmonella Enterica Typhimurium DT104. Vet. Res. 2001, 32, 301–310. [Google Scholar] [CrossRef]
- Palmer, K.L.; Kos, V.N.; Gilmore, M.S. Horizontal Gene Transfer and the Genomics of Enterococcal Antibiotic Resistance. Curr. Opin. Microbiol. 2010, 13, 632–639. [Google Scholar] [CrossRef]
- Schultsz, C.; Geerlings, S. Plasmid-Mediated Resistance in Enterobacteriaceae. Drugs 2012, 72, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Huang, P.; Lu, P. The Resistance Mechanisms and Clinical Impact of Resistance to the Third Generation Cephalosporins in Species of Enterobacter Cloacae Complex in Taiwan. Antibiotics 2022, 11, 1153. [Google Scholar] [CrossRef]
- Robicsek, A.; Jacoby, G.A.; Hooper, D.C. The Worldwide Emergence of Plasmid-Mediated Quinolone Resistance. Lancet Infect. Dis. 2006, 6, 629–640. [Google Scholar] [CrossRef] [PubMed]
- Strahilevitz, J.; Jacoby, G.A.; Hooper, D.C.; Robicsek, A. Plasmid-Mediated Quinolone Resistance: A Multifaceted Threat. Clin. Microbiol. Rev. 2009, 22, 664–689. [Google Scholar] [CrossRef] [PubMed]
- Pagano, M.; Martins, A.F.; Barth, A.L. Mobile Genetic Elements Related to Carbapenem Resistance in Acinetobacter Baumannii. Braz. J. Microbiol. 2016, 47, 785–792. [Google Scholar] [CrossRef]
- Berglund, F.; Marathe, N.P.; Osterlund, T.; Bengtsson-Palme, J.; Kotsakis, S.; Flach, C.F.; Larsson, D.G.J.; Kristiansson, E. Identification of 76 Novel B1 Metallo-Beta-Lactamases through Large-Scale Screening of Genomic and Metagenomic Data. Microbiome 2017, 5, 134. [Google Scholar] [CrossRef]
- Armin, S.; Fallah, F.; Navidinia, M.; Vosoghian, S. Prevalence of BlaOXA-1 and BlaDHA-1 AmpC β-Lactamase-Producing and Methicillin-Resistant Staphylococcus Aureus in Iran. Arch. Pediatr. Infect. Dis. 2017, 5, e36778. [Google Scholar]
- Shaheen, B.W.; Nayak, R.; Boothe, D.M. Emergence of a New Delhi Metallo-β-Lactamase (NDM-1)-Encoding Gene in Clinical Escherichia Coli Isolates Recovered from Companion Animals in the United States. Antimicrob. Agents Chemother. 2013, 57, 2902–2903. [Google Scholar] [CrossRef]
- Partridge, S.R.; Kwong, S.M.; Firth, N.; Jensen, S.O. Mobile Genetic Elements Associated with Antimicrobial Resistance. Clin. Microbiol. Rev. 2018, 31, e00088-17. [Google Scholar] [CrossRef]
- Moosavian, M.; Rahimzadeh, M. Molecular Detection of Metallo-β-Lactamase Genes, BlaIMP-1, BlaVIM-2 and BlaSPM-1 in Imipenem Resistant Pseudomonas Aeruginosa Isolated from Clinical Specimens in Teaching Hospitals of Ahvaz, Iran. Iran. J. Microbiol. 2015, 7, 2. [Google Scholar]
- Mollenkopf, D.F.; Stull, J.W.; Mathys, D.A.; Bowman, A.S.; Feicht, S.M.; Grooters, S.V.; Daniels, J.B.; Wittum, T.E. Carbapenemase-Producing Enterobacteriaceae Recovered from the Environment of a Swine Farrow-to-Finish Operation in the United States. Antimicrob. Agents Chemother. 2017, 61, e01298-16. [Google Scholar] [CrossRef]
- Mathys, D.A.; Mathys, B.A.; Mollenkopf, D.F.; Daniels, J.B.; Wittum, T.E. Enterobacteriaceae Harboring AmpC (Bla CMY) and ESBL (Bla CTX-M) in Migratory and Nonmigratory Wild Songbird Populations on Ohio Dairies. Vector-Borne Zoonotic Dis. 2017, 17, 254–259. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.P.; Woodford, N. Global Spread of Antibiotic Resistance: The Example of New Delhi Metallo-β-Lactamase (NDM)-Mediated Carbapenem Resistance. J. Med. Microbiol. 2013, 62, 499–513. [Google Scholar] [CrossRef] [PubMed]
- Fleury, C.; Resman, F.; Rau, J.; Riesbeck, K. Prevalence, Distribution and Transfer of Small β-Lactamase-Containing Plasmids in Swedish Haemophilus Influenzae. J. Antimicrob. Chemother. 2014, 69, 1238–1242. [Google Scholar] [CrossRef][Green Version]
- Tayh, G.; Al Laham, N.; Fhoula, I.; Abedelateef, N.; El-Laham, M.; Elkader Elottol, A.; Ben Slama, K. Frequency and Antibiotics Resistance of Extended-Spectrum Beta-Lactamase (ESBLs) Producing Escherichia Coli and Klebsiella Pneumoniae Isolated from Patients in Gaza Strip, Palestine. J. Med. Microbiol. Infect. Dis. 2021, 9, 133–141. [Google Scholar] [CrossRef]
- Von Wintersdorff, C.J.H.; Penders, J.; Van Niekerk, J.M.; Mills, N.D.; Majumder, S.; Van Alphen, L.B.; Savelkoul, P.H.M.; Wolffs, P.F.G. Dissemination of Antimicrobial Resistance in Microbial Ecosystems through Horizontal Gene Transfer. Front. Microbiol. 2016, 7, 173. [Google Scholar] [CrossRef] [PubMed]
- Bassetti, M.; Merelli, M.; Temperoni, C.; Astilean, A. New Antibiotics for Bad Bugs: Where Are We? Ann. Clin. Microbiol. Antimicrob. 2013, 12, 22. [Google Scholar] [CrossRef] [PubMed]
- Shirley, J.D.; Nauta, K.M.; Carlson, E.E. Live-Cell Profiling of Penicillin-Binding Protein Inhibitors in Escherichia Coli MG1655. ACS Infect. Dis. 2022, 8, 1241–1252. [Google Scholar] [CrossRef]
- Pai, H.; Kim, M.R.; Seo, M.-R.; Choi, T.Y.; Oh, S.H. A Nosocomial Outbreak of Escherichia Coli Producing CTX-M-15 and OXA-30 β-Lactamase. Infect. Control Hosp. Epidemiol. 2006, 27, 312–314. [Google Scholar] [CrossRef]
- Jamrozy, D.; Coll, F.; Mather, A.E.; Harris, S.R.; Harrison, E.M.; MacGowan, A.; Karas, A.; Elston, T.; Estée Török, M.; Parkhill, J. Evolution of Mobile Genetic Element Composition in an Epidemic Methicillin-Resistant Staphylococcus Aureus: Temporal Changes Correlated with Frequent Loss and Gain Events. BMC Genomics 2017, 18, 684. [Google Scholar] [CrossRef]
- Evans, B.A.; Amyes, S.G.B. OXA β-Lactamases. Clin. Microbiol. Rev. 2014, 27, 241–263. [Google Scholar] [CrossRef] [PubMed]
- Roshani, M.; Goodarzi, A.; Hashemi, A.; Afrasiabi, F.; Goudarzi, H.; Arabestani, M. Detection of QnrA, QnrB, and QnrS Genes in Klebsiella Pneumoniae and Escherichia Coli Isolates from Leukemia Patients. Rev. Med. Microbiol. 2022, 33, 14–19. [Google Scholar] [CrossRef]
- Ghobadi, N.; HakimiAleni, R. Evaluation of Plasmid-Mediated Quinolone Resistance Genes And Biofilm Formation in Clinical Isolates of Escherichia Coli. Exp. Anim. Biol. 2022, 10, 77–86. [Google Scholar]
- Redgrave, L.S.; Sutton, S.B.; Webber, M.A.; Piddock, L.J. V Fluoroquinolone Resistance: Mechanisms, Impact on Bacteria, and Role in Evolutionary Success. Trends Microbiol. 2014, 22, 438–445. [Google Scholar] [CrossRef]
- Haque, T.A.; Urmi, U.L.; Islam, A.B.M.M.K.; Ara, B.; Nahar, S.; Mosaddek, A.S.M.; Lugova, H.; Kumar, S.; Jahan, D.; Rahman, N.A.A. Detection of Qnr Genes and GyrA Mutation to Quinolone Phenotypic Resistance of UTI Pathogens in Bangladesh and the Implications. J. Appl. Pharm. Sci. 2022, 12, 185–198. [Google Scholar] [CrossRef]
- Yakout, M.A.; Ali, G.H. A Novel ParC Mutation Potentiating Fluoroquinolone Resistance in Klebsiella Pneumoniae and Escherichia Coli Clinical Isolates. J. Infect. Dev. Ctries. 2022, 16, 314–319. [Google Scholar] [CrossRef]
- Truong-Bolduc, Q.C.; Wang, Y.; Reedy, J.L.; Vyas, J.M.; Hooper, D.C. Staphylococcus Aureus Efflux Pumps and Tolerance to Ciprofloxacin and Chlorhexidine Following Induction by Mupirocin. Antimicrob. Agents Chemother. 2022, 66, e01845-21. [Google Scholar] [CrossRef]
- De Rossi, E.; Arrigo, P.; Bellinzoni, M.; Silva, P.E.A.; Martín, C.; Aínsa, J.A.; Guglierame, P.; Riccardi, G. The Multidrug Transporters Belonging to Major Facilitator Superfamily (MFS) in Mycobacterium Tuberculosis. Mol. Med. 2002, 8, 714–724. [Google Scholar] [CrossRef]
- Montero, C.; Mateu, G.; Rodriguez, R.; Takiff, H. Intrinsic Resistance of Mycobacterium Smegmatis to Fluoroquinolones May Be Influenced by New Pentapeptide Protein MfpA. Antimicrob. Agents Chemother. 2001, 45, 3387–3392. [Google Scholar] [CrossRef]
- Cattoir, V.; Poirel, L.; Aubert, C.; Soussy, C.-J.; Nordmann, P. Unexpected Occurrence of Plasmid-Mediated Quinolone Resistance Determinants in Environmental Aeromonas Spp. Emerg. Infect. Dis. 2008, 14, 231. [Google Scholar] [CrossRef]
- Hu, X.; Yu, X.; Shang, Y.; Xu, H.; Guo, L.; Liang, Y.; Kang, Y.; Song, L.; Sun, J.; Yue, F. Emergence and Characterization of a Novel IncP-6 Plasmid Harboring Bla Aeromonas KPC-2. In “One Health” Approach for Revealing Reservoirs and Transmission of Antimicrobial Resistance; Frontiers Media SA: Lausanne, Switzerland, 2022. [Google Scholar]
- Yamane, K.; Wachino, J.; Suzuki, S.; Kimura, K.; Shibata, N.; Kato, H.; Shibayama, K.; Konda, T.; Arakawa, Y. New Plasmid-Mediated Fluoroquinolone Efflux Pump, QepA, Found in an Escherichia Coli Clinical Isolate. Antimicrob. Agents Chemother. 2007, 51, 3354–3360. [Google Scholar] [CrossRef] [PubMed]
- Hansen, L.H.; Jensen, L.B.; Sørensen, H.I.; Sørensen, S.J. Substrate Specificity of the OqxAB Multidrug Resistance Pump in Escherichia Coli and Selected Enteric Bacteria. J. Antimicrob. Chemother. 2007, 60, 145–147. [Google Scholar] [CrossRef] [PubMed]
- Yokota, S.; Sato, K.; Kuwahara, O.; Habadera, S.; Tsukamoto, N.; Ohuchi, H.; Akizawa, H.; Himi, T.; Fujii, N. Fluoroquinolone-Resistant Streptococcus Pneumoniae Strains Occur Frequently in Elderly Patients in Japan. Antimicrob. Agents Chemother. 2002, 46, 3311–3315. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ferrándiz, M.J.; Fenoll, A.; Liñares, J.; De La Campa, A.G. Horizontal Transfer of ParC and GyrA in Fluoroquinolone-Resistant Clinical Isolates of Streptococcus Pneumoniae. Antimicrob. Agents Chemother. 2000, 44, 840–847. [Google Scholar] [CrossRef]
- Zając, O.M.; Tyski, S.; Laudy, A.E. The Contribution of Efflux Systems to Levofloxacin Resistance in Stenotrophomonas Maltophilia Clinical Strains Isolated in Warsaw, Poland. Biology 2022, 11, 1044. [Google Scholar] [CrossRef]
- Wu, C.-J.; Lu, H.-F.; Lin, Y.-T.; Zhang, M.-S.; Li, L.-H.; Yang, T.-C. Substantial Contribution of SmeDEF, SmeVWX, SmQnr, and Heat Shock Response to Fluoroquinolone Resistance in Clinical Isolates of Stenotrophomonas Maltophilia. Front. Microbiol. 2019, 10, 822. [Google Scholar] [CrossRef]
- Sánchez, M.B.; Martínez, J.L. The Efflux Pump SmeDEF Contributes to Trimethoprim-Sulfamethoxazole Resistance in Stenotrophomonas Maltophilia. Antimicrob. Agents Chemother. 2015, 59, 4347–4348. [Google Scholar] [CrossRef]
- Häussler, S.; Becker, T. The Pseudomonas Quinolone Signal (PQS) Balances Life and Death in Pseudomonas Aeruginosa Populations. PLoS Pathog. 2008, 4, e1000166. [Google Scholar] [CrossRef]
- Hansen, S.; Lewis, K.; Vulic, M. Role of Global Regulators and Nucleotide Metabolism in Antibiotic Tolerance in Escherichia Coli. Antimicrob. Agents Chemother. 2008, 52, 2718–2726. [Google Scholar] [CrossRef]
- Binda, E.; Marinelli, F.; Marcone, G.L. Old and New Glycopeptide Antibiotics: Action and Resistance. Antibiotics 2014, 3, 572–594. [Google Scholar] [CrossRef]
- Yushchuk, O.; Binda, E.; Marinelli, F. Glycopeptide Antibiotic Resistance Genes: Distribution and Function in the Producer Actinomycetes. Front. Microbiol. 2020, 11, 1173. [Google Scholar] [CrossRef]
- Kahne, D.; Leimkuhler, C.; Lu, W.; Walsh, C. Glycopeptide and Lipoglycopeptide Antibiotics. Chem. Rev. 2005, 105, 425–448. [Google Scholar] [CrossRef] [PubMed]
- Abdelhady, W.; Bayer, A.S.; Seidl, K.; Moormeier, D.E.; Bayles, K.W.; Cheung, A.; Yeaman, M.R.; Xiong, Y.Q. Impact of Vancomycin on SarA-Mediated Biofilm Formation: Role in Persistent Endovascular Infections Due to Methicillin-Resistant Staphylococcus Aureus. J. Infect. Dis. 2014, 209, 1231–1240. [Google Scholar] [CrossRef] [PubMed]
- Novais, C.; Coque, T.M.; Boerlin, P.; Herrero, I.; Moreno, M.A.; Dominguez, L.; Peixe, L. Vancomycin-Resistant Enterococcus Faecium Clone in Swine, Europe. Emerg. Infect. Dis. 2005, 11, 1985. [Google Scholar] [CrossRef] [PubMed]
- Xiong, W.; Sun, Y.; Zhang, T.; Ding, X.; Li, Y.; Wang, M.; Zeng, Z. Antibiotics, Antibiotic Resistance Genes, and Bacterial Community Composition in Fresh Water Aquaculture Environment in China. Microb. Ecol. 2015, 70, 425–432. [Google Scholar] [CrossRef]
- Schwarz, S.; Johnson, A.P. Transferable Resistance to Colistin: A New but Old Threat. J. Antimicrob. Chemother. 2016, 71, 2066–2070. [Google Scholar] [CrossRef]
- Pang, Y.; Bosch, T.; Roberts, M.C. Single Polymerase Chain Reaction for the Detection of Tetracycline-Resistant Determinants Tet K and Tet L. Mol. Cell. Probes 1994, 8, 417–422. [Google Scholar] [CrossRef]
- Showsh, S.A.; Andrews, R.E., Jr. Tetracycline Enhances Tn916-Mediated Conjugal Transfer. Plasmid 1992, 28, 213–224. [Google Scholar] [CrossRef]
- Truong-Bolduc, Q.C.; Wang, Y.; Hooper, D.C. Role of Staphylococcus aureus Tet38 in Transport of Tetracycline and Its Regulation in a Salt Stress Environment. J. Bacteriol. 2022, 204, e00142-22. [Google Scholar] [CrossRef]
- Mechler, L.; Herbig, A.; Paprotka, K.; Fraunholz, M.; Nieselt, K.; Bertram, R. A Novel Point Mutation Promotes Growth Phase-Dependent Daptomycin Tolerance in Staphylococcus Aureus. Antimicrob. Agents Chemother. 2015, 59, 5366–5376. [Google Scholar] [CrossRef]
- Yee, R.; Cui, P.; Shi, W.; Feng, J.; Zhang, Y. Genetic Screen Reveals the Role of Purine Metabolism in Staphylococcus Aureus Persistence to Rifampicin. Antibiotics 2015, 4, 627–642. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, R. Growing Group of Extended-Spectrum β-Lactamases: The CTX-M Enzymes. Antimicrob. Agents Chemother. 2004, 48, 1–14. [Google Scholar] [CrossRef] [PubMed]
- de Been, M.; Lanza, V.F.; de Toro, M.; Scharringa, J.; Dohmen, W.; Du, Y.; Hu, J.; Lei, Y.; Li, N.; Tooming-Klunderud, A. Dissemination of Cephalosporin Resistance Genes between Escherichia Coli Strains from Farm Animals and Humans by Specific Plasmid Lineages. PLoS Genet. 2014, 10, e1004776. [Google Scholar] [CrossRef] [PubMed]
- Kristich, C.J.; Rice, L.B.; Arias, C.A. Enterococcal Infection—Treatment and Antibiotic Resistance. In Enterococci from Commensals to Leading Causes of Drug Resistant Infection; Gilmore, M.S., Clewell, D.B., Ike, Y., Shankar, N., Eds.; Massachusetts Eye and Ear Infirmary: Boston, MA, USA, 2014. [Google Scholar]
- Bailey, A.M.; Webber, M.A.; Piddock, L.J. V Medium Plays a Role in Determining Expression of AcrB, MarA, and SoxS in Escherichia Coli. Antimicrob. Agents Chemother. 2006, 50, 1071–1074. [Google Scholar] [CrossRef] [PubMed]
- Douard, G.; Praud, K.; Cloeckaert, A.; Doublet, B. The Salmonella Genomic Island 1 Is Specifically Mobilized in Trans by the IncA/C Multidrug Resistance Plasmid Family. PLoS One 2010, 5, e15302. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, A.F.; Shahin, M.; Shokoohizadeh, L.; Ghanbari, F.; Solgi, H.; Shahcheraghi, F. Emerge of NDM-1-Producing Multidrug-Resistant Pseudomonas Aeruginosa and Co-Harboring of Carbapenemase Genes in South of Iran. Iran. J. Public Health 2020, 49, 959. [Google Scholar]
- He, J.; Li, C.; Cui, P.; Wang, H. Detection of Tn7-Like Transposons and Antibiotic Resistance in Enterobacterales from Animals Used for Food Production with Identification of Three Novel Transposons Tn6813, Tn6814, and Tn6765. Front. Microbiol. 2020, 11, 2049. [Google Scholar] [CrossRef]
- Akrami, F.; Rajabnia, M.; Pournajaf, A. Resistance Integrons; A Mini Review. Casp. J. Intern. Med. 2019, 10, 370–376. [Google Scholar] [CrossRef]
- Jones, R.N.; Kugler, K.C.; Pfaller, M.A.; Winokur, P.L. Characteristics of Pathogens Causing Urinary Tract Infections in Hospitals in North America: Results from the SENTRY Antimicrobial Surveillance Program, 1997. Diagn. Microbiol. Infect. Dis. 1999, 35, 55–63. [Google Scholar] [CrossRef]
- Arakawa, Y.; Murakami, M.; Suzuki, K.; Ito, H.; Wacharotayankun, R.; Ohsuka, S.; Kato, N.; Ohta, M. A Novel Integron-like Element Carrying the Metallo-Beta-Lactamase Gene BlaIMP. Antimicrob. Agents Chemother. 1995, 39, 1612–1615. [Google Scholar] [CrossRef]
- Lin, M.-F.; Liou, M.-L.; Tu, C.-C.; Yeh, H.-W.; Lan, C.-Y. Molecular Epidemiology of Integron-Associated Antimicrobial Gene Cassettes in the Clinical Isolates of Acinetobacter Baumannii from Northern Taiwan. Ann. Lab. Med. 2013, 33, 242–247. [Google Scholar] [CrossRef] [PubMed]
- Guérin, E.; Jové, T.; Tabesse, A.; Mazel, D.; Ploy, M.-C. High-Level Gene Cassette Transcription Prevents Integrase Expression in Class 1 Integrons. J. Bacteriol. 2011, 193, 5675–5682. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Bao, X.; Ji, L.; Chen, L.; Liu, J.; Miao, J.; Chen, D.; Bian, H.; Li, Y.; Yu, G. Resistance Integrons: Class 1, 2 and 3 Integrons. Ann. Clin. Microbiol. Antimicrob. 2015, 14, 45. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Carattoli, A.; Bernabeu, S.; Bruderer, T.; Frei, R.; Nordmann, P. A Novel IncQ Plasmid Type Harbouring a Class 3 Integron from Escherichia Coli. J. Antimicrob. Chemother. 2010, 65, 1594–1598. [Google Scholar] [CrossRef] [PubMed]
- Jamali, S. Integrons and Insertion Sequences Associated with Beta-Lactamases. In Beta-Lactam Resistance in Gram-Negative Bacteria; Springer: Berlin/Heidelberg, Germany, 2022; pp. 179–189. [Google Scholar]
- Deylam Salehi, M.; Ferdosi-Shahandashti, E.; Yahyapour, Y.; Khafri, S.; Pournajaf, A.; Rajabnia, R. Integron-Mediated Antibiotic Resistance in Acinetobacter Baumannii Isolated from Intensive Care Unit Patients, Babol, North of Iran. Biomed Res. Int. 2017, 2017, 7157923. [Google Scholar] [CrossRef]
- Chokshi, A.; Sifri, Z.; Cennimo, D.; Horng, H. Global Contributors to Antibiotic Resistance. J. Glob. Infect. Dis. 2019, 11, 36. [Google Scholar]
- Sreeja, M.K.; Gowrishankar, N.L.; Adisha, S.; Divya, K.C. Antibiotic Resistance-Reasons and the Most Common Resistant Pathogens–a Review. Res. J. Pharm. Technol. 2017, 10, 1886. [Google Scholar] [CrossRef]
- Blair, J.M.A.; Webber, M.A.; Baylay, A.J.; Ogbolu, D.O.; Piddock, L.J. V Molecular Mechanisms of Antibiotic Resistance. Nat. Rev. 2015, 13, 42–51. [Google Scholar] [CrossRef]
- Decousser, J.W.; Poirel, L.; Nordmann, P. Characterization of a Chromosomally Encoded Extended-Spectrum Class A β-Lactamase from Kluyvera Cryocrescens. Antimicrob. Agents Chemother. 2001, 45, 3595–3598. [Google Scholar] [CrossRef]
- Li, R.; Chan, E.W.-C.; Chen, S. Characterisation of a Chromosomally-Encoded Extended-Spectrum β-Lactamase Gene BlaPER-3 in Aeromonas Caviae of Chicken Origin. Int. J. Antimicrob. Agents 2016, 47, 103–105. [Google Scholar] [CrossRef]
- Wellington, E.M.H.; Boxall, A.B.A.; Cross, P.; Feil, E.J.; Gaze, W.H.; Hawkey, P.M.; Johnson-Rollings, A.S.; Jones, D.L.; Lee, N.M.; Otten, W.; et al. The Role of the Natural Environment in the Emergence of Antibiotic Resistance in Gram-Negative Bacteria. Lancet Infect. Dis. 2013, 13, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Forsberg, K.J.; Reyes, A.; Wang, B.; Selleck, E.M.; Sommer, M.O.A.; Dantas, G. The Shared Antibiotic Resistome of Soil Bacteria and Human Pathogens. Science 2012, 337, 1107–1111. [Google Scholar] [CrossRef] [PubMed]
- Perry, J.A.; Wright, G.D. The Antibiotic Resistance “Mobilome”: Searching for the Link between Environment and Clinic. Front. Microbiol. 2013, 4, 138. [Google Scholar] [CrossRef] [PubMed]
- Le, T.H.; Truong, T.; Tran, L.-T.; Nguyen, D.-H.; Pham, T.P.T.; Ng, C. Antibiotic Resistance in the Aquatic Environments: The Need for an Interdisciplinary Approach. Int. J. Environ. Sci. Technol. 2022, 244. [Google Scholar] [CrossRef]
- Marti, E.; Balcázar, J.L. Antibiotic Resistance in the Aquatic Environment. In Comprehensive Analytical Chemistry; Elsevier: Amsterdam, The Netherlands, 2013; Volume 62, pp. 671–684. ISBN 0166-526X. [Google Scholar]
- Munita, J.M.; Arias, C.A. Mechanisms of Antibiotic Resistance. Microbiol. Spectr. 2016, 4, 2. [Google Scholar] [CrossRef]
- Pagès, J.M.; Masi, M.; Barbe, J. Inhibitors of Efflux Pumps in Gram-Negative Bacteria. Trends Mol. Med. 2005, 11, 382–389. [Google Scholar] [CrossRef]
- Srikumar, R.; Kon, T.; Gotoh, N.; Poole, K. Expression of Pseudomonas Aeruginosa Multidrug Efflux Pumps MexA-MexB-OprM and MexC-MexD-OprJ in a Multidrug-Sensitive Escherichia Coli Strain. Antimicrob. Agents Chemother. 1998, 42, 65–71. [Google Scholar] [CrossRef]
- Huovinen, P. Trimethoprim Resistance. Antimicrob. Agents Chemother. 1987, 31, 1451–1456. [Google Scholar] [CrossRef]
- Yun, M.-K.; Wu, Y.; Li, Z.; Zhao, Y.; Waddell, M.B.; Ferreira, A.M.; Lee, R.E.; Bashford, D.; White, S.W. Catalysis and Sulfa Drug Resistance in Dihydropteroate Synthase. Science 2012, 335, 1110–1114. [Google Scholar] [CrossRef]
- Livermore, D.M. Mechanisms of Resistance to Cephalosporin Antibiotics. Drugs 1987, 34, 64–88. [Google Scholar] [CrossRef]
- Livermore, D.M. Mechanisms of Resistance To-Lactam Antibiotics. Scand. J. Infect. Dis. 1991, 78, 7–16. [Google Scholar]
- MacGowan, A.; Macnaughton, E. Antibiotic Resistance. Medicine 2017, 45, 622–628. [Google Scholar] [CrossRef]
- Neu, H.C. The Crisis in Antibiotic Resistance. Science 1992, 257, 1064–1073. [Google Scholar] [CrossRef] [PubMed]
- Brooun, A.; Liu, S.; Lewis, K. A Dose-Response Study of Antibiotic Resistance in Pseudomonas Aeruginosa Biofilms. Antimicrob. Agents Chemother. 2000, 44, 640–646. [Google Scholar] [CrossRef]
- Oliveira, M.; Leonardo, I.C.; Nunes, M.; Silva, A.F.; Barreto Crespo, M.T. Environmental and Pathogenic Carbapenem Resistant Bacteria Isolated from a Wastewater Treatment Plant Harbour Distinct Antibiotic Resistance Mechanisms. Antibiotics 2021, 10, 1118. [Google Scholar] [CrossRef]
- Suay-García, B.; Pérez-Gracia, M.T. Drug-Resistant Neisseria Gonorrhoeae: Latest Developments. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 1065–1071. [Google Scholar] [CrossRef]
- Singh, A.; Shahid, M.; Khan, P.A.; Khan, H.M.; Sami, H. An Overview on Antibiotic Resistance in Gram-Negative Bacteria. In Beta-Lactam Resistance in Gram-Negative Bacteria; Springer Nature: Cham, Switzerland, 2022; pp. 3–15. [Google Scholar]
- Schiavano, G.F.; Carloni, E.; Andreoni, F.; Magi, S.; Chironna, M.; Brandi, G.; Amagliani, G. Prevalence and Antibiotic Resistance of Pseudomonas Aeruginosa in Water Samples in Central Italy and Molecular Characterization of Opr D in Imipenem Resistant Isolates. PLoS ONE 2017, 12, e0189172. [Google Scholar] [CrossRef]
- Ma, L.; Lu, P.-L.; Siu, L.K.; Hsieh, M.-H. Molecular Typing and Resistance Mechanisms of Imipenem-Non-Susceptible Klebsiella Pneumoniae in Taiwan: Results from the Taiwan Surveillance of Antibiotic Resistance (TSAR) Study, 2002–2009. J. Med. Microbiol. 2013, 62, 101–107. [Google Scholar] [CrossRef]
- Hiramatsu, K. Vancomycin-Resistant Staphylococcus Aureus: A New Model of Antibiotic Resistance. Lancet Infect. Dis. 2001, 1, 147–155. [Google Scholar] [CrossRef]
- Arias, C.A.; Murray, B.E. The Rise of the Enterococcus: Beyond Vancomycin Resistance. Nat. Rev. Microbiol. 2012, 10, 266–278. [Google Scholar] [CrossRef]
- Schweizer, H.P. Mechanisms of Antibiotic Resistance in Burkholderia Pseudomallei: Implications for Treatment of Melioidosis. Future Microbiol. 2012, 7, 1389–1399. [Google Scholar] [CrossRef] [PubMed]
- Brochet, M.; Couvé, E.; Zouine, M.; Poyart, C.; Glaser, P. A Naturally Occurring Gene Amplification Leading to Sulfonamide and Trimethoprim Resistance in Streptococcus Agalactiae. J. Bacteriol. 2008, 190, 672–680. [Google Scholar] [CrossRef] [PubMed]
- Lauxen, A.I.; Kobauri, P.; Wegener, M.; Hansen, M.J.; Galenkamp, N.S.; Maglia, G.; Szymanski, W.; Feringa, B.L.; Kuipers, O.P. Mechanism of Resistance Development in E. Coli against TCAT, a Trimethoprim-Based Photoswitchable Antibiotic. Pharmaceuticals 2021, 14, 392. [Google Scholar] [CrossRef] [PubMed]
- Sköld, O. Resistance to Trimethoprim and Sulfonamides. Vet. Res. 2001, 32, 261–273. [Google Scholar] [CrossRef]
- Parulekar, R.S.; Sonawane, K.D. Insights into the Antibiotic Resistance and Inhibition Mechanism of Aminoglycoside Phosphotransferase from Bacillus Cereus: In Silico and in Vitro Perspective. J. Cell. Biochem. 2018, 119, 9444–9461. [Google Scholar] [CrossRef]
- Pantosti, A.; Sanchini, A.; Monaco, M. Mechanisms of Antibiotic Resistance in Staphylococcus Aureus. Futur. Microbiol. 2007, 2, 323–334. [Google Scholar] [CrossRef]
- Bryan, L.E.; Kowand, S.K.; Van Den Elzen, H.M. Mechanism of Aminoglycoside Antibiotic Resistance in Anaerobic Bacteria: Clostridium Perfringens and Bacteroides Fragilis. Antimicrob. Agents Chemother. 1979, 15, 7–13. [Google Scholar] [CrossRef]
- Moreira, M.A.S.; Oliveira, J.A.; Teixeira, L.M.; Moraes, C.A. Detection of a Chloramphenicol Efflux System in Escherichia Coli Isolated from Poultry Carcass. Vet. Microbiol. 2005, 109, 75–81. [Google Scholar] [CrossRef]
- Wolter, N.; Smith, A.M.; Farrell, D.J.; Schaffner, W.; Moore, M.; Whitney, C.G.; Jorgensen, J.H.; Klugman, K.P. Novel Mechanism of Resistance to Oxazolidinones, Macrolides, and Chloramphenicol in Ribosomal Protein L4 of the Pneumococcus. Antimicrob. Agents Chemother. 2005, 49, 3554–3557. [Google Scholar] [CrossRef]
- Mosher, R.H.; Camp, D.J.; Yang, K.; Brown, M.P.; Shaw, W.V.; Vining, L.C. Inactivation of Chloramphenicol by O-Phosphorylation: A Novel Resistance Mechanism in Streptomyces Venezuelae Isp5230, a Chloramphenicol Producer. J. Biol. Chem. 1995, 270, 27000–27006. [Google Scholar] [CrossRef]
- Fernández, M.; Conde, S.; de la Torre, J.; Molina-Santiago, C.; Ramos, J.-L.; Duque, E. Mechanisms of Resistance to Chloramphenicol in Pseudomonas Putida KT2440. Antimicrob. Agents Chemother. 2012, 56, 1001–1009. [Google Scholar] [CrossRef] [PubMed]
- Shortridge, V.D.; Doern, G.V.; Brueggemann, A.B.; Beyer, J.M.; Flamm, R.K. Prevalence of Macrolide Resistance Mechanisms in Streptococcus Pneumoniae Isolates from a Multicenter Antibiotic Resistance Surveillance Study Conducted in the United States in 1994–1995. Clin. Infect. Dis. 1999, 29, 1186–1188. [Google Scholar] [CrossRef]
- Bolukaoto, J.Y.; Monyama, C.M.; Chukwu, M.O.; Lekala, S.M.; Nchabeleng, M.; Maloba, M.R.B.; Mavenyengwa, R.T.; Lebelo, S.L.; Monokoane, S.T.; Tshepuwane, C.; et al. Antibiotic Resistance of Streptococcus Agalactiae Isolated from Pregnant Women in Garankuwa, South Africa. BMC Res. Notes 2015, 8, 6–12. [Google Scholar] [CrossRef]
- Kyriakidis, I.; Vasileiou, E.; Pana, Z.D.; Tragiannidis, A. Acinetobacter Baumannii Antibiotic Resistance Mechanisms. Pathogens 2021, 10, 373. [Google Scholar] [CrossRef]
- De Francesco, V.; Zullo, A.; Hassan, C.; Giorgio, F.; Rosania, R.; Ierardi, E. Mechanisms of Helicobacter Pylori Antibiotic Resistance: An Updated Appraisal. World J. Gastrointest. Pathophysiol. 2011, 2, 35. [Google Scholar] [CrossRef] [PubMed]
- Roberts, M.C. Mechanisms of Bacterial Antibiotic Resistance and Lessons Learned from Environmental Tetracycline-Resistant Bacteria. In Antimicrobial Resistance in the Environment; John Wiley & Sons, Inc.: New York, NY, USA, 2011; pp. 93–121. [Google Scholar]
- Nordmann, P.; Poirel, L. Emergence of Plasmid-Mediated Resistance to Quinolones in Enterobacteriaceae. J. Antimicrob. Chemother. 2005, 56, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, M.B. Antibiotic Resistance in the Opportunistic Pathogen Stenotrophomonas Maltophilia. Front. Microbiol. 2015, 6, 658. [Google Scholar] [CrossRef]
- Prince, A. Antibiotic Resistance of Pseudomonas Species. J. Pediatr. 1986, 108, 830–834. [Google Scholar] [CrossRef]
- Nnadozie, C.F.; Odume, O.N. Freshwater Environments as Reservoirs of Antibiotic Resistant Bacteria and Their Role in the Dissemination of Antibiotic Resistance Genes. Environ. Pollut. 2019, 254, 113067. [Google Scholar] [CrossRef]
- Marti, E.; Variatza, E.; Balcazar, J.L. The Role of Aquatic Ecosystems as Reservoirs of Antibiotic Resistance. Trends Microbiol. 2014, 22, 36–41. [Google Scholar] [CrossRef]
- Naquin, A.; Shrestha, A.; Sherpa, M.; Nathaniel, R.; Boopathy, R. Presence of Antibiotic Resistance Genes in a Sewage Treatment Plant in Thibodaux, Louisiana, USA. Bioresour. Technol. 2015, 188, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Rowan, N.J. Defining Established and Emerging Microbial Risks in the Aquatic Environment: Current Knowledge, Implications, and Outlooks. Int. J. Microbiol. 2011, 2011, 462832. [Google Scholar] [CrossRef] [PubMed]
- Michael, I.; Rizzo, L.; Mcardell, C.S.; Manaia, C.M.; Merlin, C.; Schwartz, T.; Dagot, C.; Fatta-kassinos, D. Urban Wastewater Treatment Plants as Hotspots for the Release of Antibiotics in the Environment: A Review. Water Res. 2012, 47, 957–995. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.-F.; Lin, C.-F.; Lin, A.Y.-C.; Hong, P.-K.A. Sorption and Biodegradation of Sulfonamide Antibiotics by Activated Sludge: Experimental Assessment Using Batch Data Obtained under Aerobic Conditions. Water Res. 2011, 45, 3389–3397. [Google Scholar] [CrossRef]
- Ben, W.; Wang, J.; Cao, R.; Yang, M.; Zhang, Y.; Qiang, Z. Distribution of Antibiotic Resistance in the Effluents of Ten Municipal Wastewater Treatment Plants in China and the Effect of Treatment Processes. Chemosphere 2017, 172, 392–398. [Google Scholar] [CrossRef]
- Dorival-García, N.; Zafra-Gómez, A.; Navalón, A.; González, J.; Vílchez, J.L. Removal of Quinolone Antibiotics from Wastewaters by Sorption and Biological Degradation in Laboratory-Scale Membrane Bioreactors. Sci. Total Environ. 2013, 442, 317–328. [Google Scholar] [CrossRef]
- Li, B.; Zhang, T. Biodegradation and Adsorption of Antibiotics in the Activated Sludge Process. Environ. Sci. Technol. 2010, 44, 3468–3473. [Google Scholar] [CrossRef]
- Rowe, W.P.M.; Baker-Austin, C.; Verner-Jeffreys, D.W.; Ryan, J.J.; Micallef, C.; Maskell, D.J.; Pearce, G.P. Overexpression of Antibiotic Resistance Genes in Hospital Effluents over Time. J. Antimicrob. Chemother. 2017, 72, 1617–1623. [Google Scholar] [CrossRef]
- Kümmerer, K. Antibiotics in the Aquatic Environment–a Review–Part I. Chemosphere 2009, 75, 417–434. [Google Scholar] [CrossRef]
- Sabaté, M.; Prats, G.; Moreno, E.; Ballesté, E.; Blanch, A.R.; Andreu, A. Virulence and Antimicrobial Resistance Profiles among Escherichia Coli Strains Isolated from Human and Animal Wastewater. Res. Microbiol. 2008, 159, 288–293. [Google Scholar] [CrossRef]
- Da Costa, P.M.; Vaz-Pires, P.; Bernardo, F. Antimicrobial Resistance in Enterococcus Spp. Isolated in Inflow, Effluent and Sludge from Municipal Sewage Water Treatment Plants. Water Res. 2006, 40, 1735–1740. [Google Scholar] [CrossRef] [PubMed]
- da Costa, P.M.; Oliveira, M.; Bica, A.; Vaz-Pires, P.; Bernardo, F. Antimicrobial Resistance in Enterococcus Spp. and Escherichia Coli Isolated from Poultry Feed and Feed Ingredients. Vet. Microbiol. 2007, 120, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Mao, D.; Yu, S.; Rysz, M.; Luo, Y.; Yang, F.; Li, F.; Hou, J.; Mu, Q.; Alvarez, P.J.J. Prevalence and Proliferation of Antibiotic Resistance Genes in Two Municipal Wastewater Treatment Plants. Water Res. 2015, 85, 458–466. [Google Scholar] [CrossRef] [PubMed]
- Cacace, D.; Fatta-Kassinos, D.; Manaia, C.M.; Cytryn, E.; Kreuzinger, N.; Rizzo, L.; Karaolia, P.; Schwartz, T.; Alexander, J.; Merlin, C.; et al. Antibiotic Resistance Genes in Treated Wastewater and in the Receiving Water Bodies: A Pan-European Survey of Urban Settings. Water Res. 2019, 162, 320–330. [Google Scholar] [CrossRef]
- Lien, L.; Lan, P.; Chuc, N.; Hoa, N.; Nhung, P.; Thoa, N.; Diwan, V.; Tamhankar, A.; Stålsby Lundborg, C. Antibiotic Resistance and Antibiotic Resistance Genes in Escherichia Coli Isolates from Hospital Wastewater in Vietnam. Int. J. Environ. Res. Public Health 2017, 14, 699. [Google Scholar] [CrossRef]
- Lundborg, C.S.; Tamhankar, A.J. Antibiotic Residues in the Environment of South East Asia. BMJ 2017, 358, 42–45. [Google Scholar] [CrossRef]
- Selvarajan, R.; Sibanda, T.; Pandian, J.; Mearns, K. Taxonomic and Functional Distribution of Bacterial Communities in Domestic and Hospital Wastewater System: Implications for Public and Environmental Health. Antibiotics 2021, 10, 1059. [Google Scholar] [CrossRef]
- Kunhikannan, S.; Thomas, C.J.; Franks, A.E.; Mahadevaiah, S.; Kumar, S.; Petrovski, S. Environmental Hotspots for Antibiotic Resistance Genes. Microbiologyopen 2021, 10, e1197. [Google Scholar] [CrossRef]
- Rozman, U.; Duh, D.; Cimerman, M.; Turk, S.Š. Hospital Wastewater Effluent: Hot Spot for Antibiotic Resistant Bacteria. J. Water Sanit. Hyg. Dev. 2020, 10, 171–178. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, P.; Yang, Q. Occurrence and Diversity of Antibiotic Resistance in Untreated Hospital Wastewater. Sci. Total Environ. 2018, 621, 990–999. [Google Scholar] [CrossRef]
- Obayiuwana, A.; Ibekwe, A.M. Antibiotic Resistance Genes Occurrence in Wastewaters from Selected Pharmaceutical Facilities in Nigeria. Water 2020, 12, 1897. [Google Scholar] [CrossRef]
- Zhang, S.; Huang, J.; Zhao, Z.; Cao, Y.; Li, B. Hospital Wastewater as a Reservoir for Antibiotic Resistance Genes: A Meta-Analysis. Front. Public Health 2020, 8, 574968. [Google Scholar] [CrossRef] [PubMed]
- Obinwanne, C.; Nyaruaba, R.; Ekeng, R.; Okon, U.; Izuma, C.; Ebido, C.C.; Oluwole, A.; Sunday, E.; Ikechukwu, K. Antibiotic Resistance in the Aquatic Environment: Analytical Techniques and Interactive Impact of Emerging Contaminants. Environ. Toxicol. Pharmacol. 2022, 96, 103995. [Google Scholar] [CrossRef]
- Baquero, F.; Martínez, J.L.; Cantón, R. Antibiotics and Antibiotic Resistance in Water Environments. Curr. Opin. Biotechnol. 2008, 19, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Ji, M.; Zhai, H.; Guo, Y.; Liu, Y. Science of the Total Environment Occurrence of Antibiotics and Antibiotic Resistance Genes in WWTP Ef Fl Uent-Receiving Water Bodies and Reclaimed Wastewater Treatment Plants. Sci. Total Environ. 2021, 796, 148919. [Google Scholar] [CrossRef]
- Park, J.C.; Lee, J.C.; Oh, J.Y.; Jeong, Y.W.; Cho, J.W.; Joo, H.S.; Lee, W.K.; Lee, W.B. Antibiotic Selective Pressure for the Maintenance of Antibiotic Resistant Genes in Coliform Bacteria Isolated from the Aquatic Environment. Water Sci. Technol. 2003, 47, 249–253. [Google Scholar] [CrossRef]
- Zhu, B. Abundance Dynamics and Sequence Variation of Neomycin Phosphotransferase Gene (NptII) Homologs in River Water. Aquat. Microb. Ecol. 2007, 48, 131–140. [Google Scholar] [CrossRef][Green Version]
- Mohapatra, H.; Mohapatra, S.S.; Mantri, C.K.; Colwell, R.R.; Singh, D. V Vibrio Cholerae Non-O1, Non-O139 Strains Isolated before 1992 from Varanasi, India Are Multiple Drug Resistant, Contain IntSXT, Dfr18 and AadA5 Genes. Environ. Microbiol. 2008, 10, 866–873. [Google Scholar] [CrossRef]
- Pei, R.; Kim, S.-C.; Carlson, K.H.; Pruden, A. Effect of River Landscape on the Sediment Concentrations of Antibiotics and Corresponding Antibiotic Resistance Genes (ARG). Water Res. 2006, 40, 2427–2435. [Google Scholar] [CrossRef]
- Mukherjee, S.; Chakraborty, R. Incidence of Class 1 Integrons in Multiple Antibiotic-Resistant Gram-Negative Copiotrophic Bacteria from the River Torsa in India. Res. Microbiol. 2006, 157, 220–226. [Google Scholar] [CrossRef]
- Bergeron, S.; Boopathy, R.; Nathaniel, R.; Corbin, A.; LaFleur, G. Presence of Antibiotic Resistant Bacteria and Antibiotic Resistance Genes in Raw Source Water and Treated Drinking Water. Int. Biodeterior. Biodegrad. 2015, 102, 370–374. [Google Scholar] [CrossRef]
- Vulliet, E.; Cren-olivé, C. Screening of Pharmaceuticals and Hormones at the Regional Scale, in Surface and Groundwaters Intended to Human Consumption. Environ. Pollut. 2011, 159, 2929–2934. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, R.A.; Polk, J.S.; Datta, T.; Parekh, R.R.; Agga, G.E. Occurrence of Antibiotic Resistant Bacteria in Urban Karst Groundwater Systems. Water 2022, 14, 960. [Google Scholar] [CrossRef]
- Andrade, L.; Kelly, M.; Hynds, P.; Weatherill, J.; Majury, A.; Dwyer, O. Groundwater Resources as a Global Reservoir for Antimicrobial-Resistant Bacteria. Water Res. 2020, 170, 115360. [Google Scholar] [CrossRef]
- Zou, H.Y.; He, L.Y.; Gao, F.Z.; Zhang, M.; Chen, S.; Wu, D.L.; Liu, Y.S.; He, L.X.; Bai, H.; Ying, G.G. Antibiotic Resistance Genes in Surface Water and Groundwater from Mining Affected Environments. Sci. Total Environ. 2021, 772, 145516. [Google Scholar] [CrossRef]
- Kampouris, I.D.; Alygizakis, N.; Klümper, U.; Agrawal, S.; Lackner, S.; Cacace, D.; Kunze, S.; Thomaidis, N.S.; Slobdonik, J.; Berendonk, T.U. Elevated Levels of Antibiotic Resistance in Groundwater during Treated Wastewater Irrigation Associated with Infiltration and Accumulation of Antibiotic Residues. J. Hazard. Mater. 2022, 423, 127155. [Google Scholar] [CrossRef]
- Murphy, H.M.; Prioleau, M.D.; Borchardt, M.A.; Hynds, P.D. Epidemiological Evidence of Groundwater Contribution to Global Enteric Disease, 1948–2015. Hydrogeol. J. 2017, 25, 981–1001. [Google Scholar] [CrossRef]
- Hatosy, S.M.; Martiny, A.C. The Ocean as a Global Reservoir of Antibiotic Resistance Genes. Appl. Environ. Microbiol. 2015, 81, 7593–7599. [Google Scholar] [CrossRef]
- Habibi, N.; Uddin, S.; Lyons, B.; Al-Sarawi, H.A.; Behbehani, M.; Shajan, A.; Razzack, N.A.; Zakir, F.; Alam, F. Antibiotic Resistance Genes Associated with Marine Surface Sediments: A Baseline from the Shores of Kuwait. Sustainability 2022, 14, 8029. [Google Scholar] [CrossRef]
- Feng-xia, Y.; Da-qing, M.A.O.; Yi, L.U.O.; Qing, W.; Quan-Hua, M.U. Horizontal Transfer of Antibiotic Resistance Genes in the Environment. Yingyong Shengtai Xuebao 2013, 24, 2993–3002. [Google Scholar]
- Song, L.; Li, L.; Yang, S.; Lan, J.; He, H.; McElmurry, S.P.; Zhao, Y. Sulfamethoxazole, Tetracycline and Oxytetracycline and Related Antibiotic Resistance Genes in a Large-Scale Landfill, China. Sci. Total Environ. 2016, 551, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Xu, Y.; Wang, H.; Guo, C.; Qiu, H.; He, Y.; Zhang, Y.; Li, X.; Meng, W. Occurrence of Antibiotics and Antibiotic Resistance Genes in a Sewage Treatment Plant and Its Effluent-Receiving River. Chemosphere 2015, 119, 1379–1385. [Google Scholar] [CrossRef] [PubMed]
- Zou, S.; Xu, W.; Zhang, R.; Tang, J.; Chen, Y.; Zhang, G. Occurrence and Distribution of Antibiotics in Coastal Water of the Bohai Bay, China: Impacts of River Discharge and Aquaculture Activities. Environ. Pollut. 2011, 159, 2913–2920. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Yuan, T.; Zhou, L.; Cheng, S.; Qu, X.; Lu, P.; Feng, Q. Impact Factors of the Accumulation, Migration and Spread of Antibiotic Resistance in the Environment. Environ. Geochem. Health 2021, 43, 1741–1758. [Google Scholar] [CrossRef]
- Seiler, C.; Berendonk, T.U. Heavy Metal Driven Co-Selection of Antibiotic Resistance in Soil and Water Bodies Impacted by Agriculture and Aquaculture. Front. Microbiol. 2012, 3, 399. [Google Scholar] [CrossRef]
- Qiu, Z.; Shen, Z.; Qian, D.; Jin, M.; Yang, D.; Wang, J.; Zhang, B.; Yang, Z.; Chen, Z.; Wang, X. Effects of Nano-TiO2 on Antibiotic Resistance Transfer Mediated by RP4 Plasmid. Nanotoxicology 2015, 9, 895–904. [Google Scholar] [CrossRef]
- Wang, X.; Yang, F.; Zhao, J.; Xu, Y.; Mao, D.; Zhu, X.; Luo, Y.; Alvarez, P.J.J. Bacterial Exposure to ZnO Nanoparticles Facilitates Horizontal Transfer of Antibiotic Resistance Genes. NanoImpact 2018, 10, 61–67. [Google Scholar] [CrossRef]
- Qiu, X.; Zhou, G.; Wang, H. Nanoscale Zero-Valent Iron Inhibits the Horizontal Gene Transfer of Antibiotic Resistance Genes in Chicken Manure Compost. J. Hazard. Mater. 2022, 422, 126883. [Google Scholar] [CrossRef]
- Imran, M.; Das, K.R.; Naik, M.M. Co-Selection of Multi-Antibiotic Resistance in Bacterial Pathogens in Metal and Microplastic Contaminated Environments: An Emerging Health Threat. Chemosphere 2019, 215, 846–857. [Google Scholar] [CrossRef]
- Bondarczuk, K.; Markowicz, A.; Piotrowska-Seget, Z. The Urgent Need for Risk Assessment on the Antibiotic Resistance Spread via Sewage Sludge Land Application. Environ. Int. 2016, 87, 49–55. [Google Scholar] [CrossRef]
- Yang, Y.; Xu, C.; Cao, X.; Lin, H.; Wang, J. Antibiotic Resistance Genes in Surface Water of Eutrophic Urban Lakes Are Related to Heavy Metals, Antibiotics, Lake Morphology and Anthropic Impact. Ecotoxicology 2017, 26, 831–840. [Google Scholar] [CrossRef] [PubMed]
- Andersson, D.I.; Balaban, N.Q.; Baquero, F.; Courvalin, P.; Glaser, P.; Gophna, U.; Kishony, R.; Molin, S.; Tønjum, T. Antibiotic Resistance: Turning Evolutionary Principles into Clinical Reality. FEMS Microbiol. Rev. 2020, 44, 171–188. [Google Scholar] [CrossRef] [PubMed]
- Baker-Austin, C.; Wright, M.S.; Stepanauskas, R.; McArthur, J.V. Co-Selection of Antibiotic and Metal Resistance. Trends Microbiol. 2006, 14, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Ouvrard, S.; Barnier, C.; Bauda, P.; Beguiristain, T.; Biache, C.; Bonnard, M.; Caupert, C.; Cebron, A.; Cortet, J.; Cotelle, S.; et al. In Situ Assessment of Phytotechnologies for Multicontaminated Soil Management. Int. J. Phytoremediation 2011, 13, 245–263. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.; He, L.; Li, T.; Dai, Z.; Sun, S.; Ren, L.; Liang, Y.-Q.; Zhang, Y.; Li, C. Impact of the Surrounding Environment on Antibiotic Resistance Genes Carried by Microplastics in Mangroves. Sci. Total Environ. 2022, 837, 155771. [Google Scholar] [CrossRef]
- Aslam, B.; Wang, W.; Arshad, M.I.; Khurshid, M.; Muzammil, S.; Rasool, M.H.; Nisar, M.A.; Alvi, R.F.; Aslam, M.A.; Qamar, M.U.; et al. Antibiotic Resistance: A Rundown of a Global Crisis. Infect. Drug Resist. 2018, 11, 1645. [Google Scholar] [CrossRef] [PubMed]
- Aydin, S.; Ünlü, İ.D.; Arabacı, D.N.; Duru, Ö.A. Evaluating the Effect of Microalga Haematococcus Pluvialis Bioaugmentation on Aerobic Membrane Bioreactor in Terms of Performance, Membrane Fouling and Microbial Community Structure. Sci. Total Environ. 2022, 807, 149908. [Google Scholar] [CrossRef] [PubMed]
- Pu, M.; Ailijiang, N.; Mamat, A.; Chang, J.; Zhang, Q.; Liu, Y.; Li, N. Occurrence of Antibiotics in the Different Biological Treatment Processes, Reclaimed Wastewater Treatment Plants and Effluent-Irrigated Soils. J. Environ. Chem. Eng. 2022, 10, 107715. [Google Scholar] [CrossRef]
- Munir, M.; Wong, K.; Xagoraraki, I. Release of Antibiotic Resistant Bacteria and Genes in the Effluent and Biosolids of Five Wastewater Utilities in Michigan. Water Res. 2010, 45, 681–693. [Google Scholar] [CrossRef]
- Yan, Q.; Xu, Y.; Chen, L.; Cao, Z.; Shao, Y.; Xu, Y.; Yu, Y.; Fang, C.; Zhu, Z.; Feng, G.; et al. Irrigation with Secondary Municipal-Treated Wastewater: Potential Effects, Accumulation of Typical Antibiotics and Grain Quality Responses in Rice (Oryza Sativa L.). J. Hazard. Mater. 2021, 410, 124655. [Google Scholar] [CrossRef]
- Dungan, R.S.; Strausbaugh, C.A.; Leytem, A.B. Survey of Selected Antibiotic Resistance Genes in Agricultural and Non-Agricultural Soils in South-Central Idaho. FEMS Microbiol. Ecol. 2019, 95, fiz071. [Google Scholar] [CrossRef] [PubMed]
- Bougnom, B.P.; Thiele-Bruhn, S.; Ricci, V.; Zongo, C.; Piddock, L.J.V. Raw Wastewater Irrigation for Urban Agriculture in Three African Cities Increases the Abundance of Transferable Antibiotic Resistance Genes in Soil, Including Those Encoding Extended Spectrum β-Lactamases (ESBLs). Sci. Total Environ. 2020, 698, 134201. [Google Scholar] [CrossRef] [PubMed]
- Szczepanowski, R.; Linke, B.; Krahn, I.; Gartemann, K.-H.; Guetzkow, T.; Eichler, W.; Pühler, A.; Schlueter, A. Detection of 140 Clinically Relevant Antibiotic-Resistance Genes in the Plasmid Metagenome of Wastewater Treatment Plant Bacteria Showing Reduced Susceptibility to Selected Antibiotics. Microbiology 2009, 155, 2306–2319. [Google Scholar] [CrossRef] [PubMed]
- Gudda, F.O.; Waigi, M.G.; Odinga, E.S.; Yang, B.; Carter, L.; Gao, Y. Antibiotic-Contaminated Wastewater Irrigated Vegetables Pose Resistance Selection Risks to the Gut Microbiome. Environ. Pollut. 2020, 264, 114752. [Google Scholar] [CrossRef] [PubMed]
- Onalenna, O.; Rahube, T.O. Assessing Bacterial Diversity and Antibiotic Resistance Dynamics in Wastewater Effluent-Irrigated Soil and Vegetables in a Microcosm Setting. Heliyon 2022, 8, e09089. [Google Scholar] [CrossRef]
- Sun, C.; Zhang, B.; Ning, D.; Zhang, Y.; Dai, T.; Wu, L.; Li, T.; Liu, W.; Zhou, J.; Wen, X. Seasonal Dynamics of the Microbial Community in Two Full-Scale Wastewater Treatment Plants: Diversity, Composition, Phylogenetic Group Based Assembly and Co-Occurrence Pattern. Water Res. 2021, 200, 117295. [Google Scholar] [CrossRef] [PubMed]
- Abdalla, S.E.; Abia, A.L.K.; Amoako, D.G.; Perrett, K.; Bester, L.A.; Essack, S.Y. From Farm-to-Fork: E. Coli from an Intensive Pig Production System in South Africa Shows High Resistance to Critically Important Antibiotics for Human and Animal Use. Antibiotics 2021, 10, 178. [Google Scholar] [CrossRef]
- Molechan, C.; Amoako, D.G.; Abia, A.L.K.; Somboro, A.M.; Bester, L.A.; Essack, S.Y. Molecular Epidemiology of Antibiotic-Resistant Enterococcus Spp. from the Farm-to-Fork Continuum in Intensive Poultry Production in KwaZulu-Natal, South Africa. Sci. Total Environ. 2019, 692, 868–878. [Google Scholar] [CrossRef]
- Chereau, F.; Opatowski, L.; Tourdjman, M.; Vong, S. Risk Assessment for Antibiotic Resistance in South East Asia. BMJ 2017, 358, j3393. [Google Scholar] [CrossRef]
- Fatoba, D.O.; Abia, A.L.K.; Amoako, D.G.; Essack, S.Y. Rethinking Manure Application: Increase in Multidrug-Resistant Enterococcus Spp. in Agricultural Soil Following Chicken Litter Application. Microorganisms 2021, 9, 885. [Google Scholar] [CrossRef]
- Zhang, L.; Li, L.; Sha, G.; Liu, C.; Wang, Z.; Wang, L. Aerobic Composting as an Effective Cow Manure Management Strategy for Reducing the Dissemination of Antibiotic Resistance Genes: An Integrated Meta-Omics Study. J. Hazard. Mater. 2020, 386, 121895. [Google Scholar] [CrossRef] [PubMed]
- Skandalis, N.; Maeusli, M.; Papafotis, D.; Miller, S.; Lee, B.; Theologidis, I.; Luna, B. Environmental Spread of Antibiotic Resistance. Antibiotics 2021, 10, 640. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Du, Z.; Guo, T.; Wu, J.; Wang, B.; Wei, Z.; Jia, L.; Kang, K. Effects of Heavy Metals Stress on Chicken Manures Composting via the Perspective of Microbial Community Feedback. Environ. Pollut. 2022, 294, 118624. [Google Scholar] [CrossRef]
- Mu, M.; Yang, F.; Han, B.; Tian, X.; Zhang, K. Manure Application: A Trigger for Vertical Accumulation of Antibiotic Resistance Genes in Cropland Soils. Ecotoxicol. Environ. Saf. 2022, 237, 113555. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.; Zhang, H.; Song, D.; Chen, H.; Lin, X.; Wang, Y.; Ji, L. Distribution of Antibiotic, Heavy Metals and Antibiotic Resistance Genes in Livestock and Poultry Feces from Different Scale of Farms in Ningxia, China. J. Hazard. Mater. 2022, 440, 129719. [Google Scholar] [CrossRef] [PubMed]
- Patricia Macías Farrera, G.; Tenorio Borroto, E.; Rivera Ramírez, F.; Vázquez Chagoyán, J.; Talavera Rojas, M.; Yong Angel, G.; Montes de Oca Jimenez, R. Detection of Quinolone Resistance in Salmonella Typhimurium Pig Isolates Determined by GyrA Gene Mutation Using PCR-and Sequence-Based Techniques within the GyrA Gene. Curr. Pharm. Des. 2016, 22, 5079–5084. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishnan, B.; Venkateswarlu, K.; Sethunathan, N.; Megharaj, M. Local Applications but Global Implications: Can Pesticides Drive Microorganisms to Develop Antimicrobial Resistance? Sci. Total Environ. 2019, 654, 177–189. [Google Scholar] [CrossRef]
- Xing, Y.; Herrera, D.; Zhang, S.; Kang, X.; Men, Y. Site-Specific Target-Modification Mutations Exclusively Induced by the Coexposure to Low Levels of Pesticides and Streptomycin Caused Strong Streptomycin Resistance in Clinically Relevant Escherichia Coli. J. Hazard. Mater. Adv. 2022, 7, 100141. [Google Scholar] [CrossRef]
- Shahid, M.; Khan, M.S. Tolerance of Pesticides and Antibiotics among Beneficial Soil Microbes Recovered from Contaminated Rhizosphere of Edible Crops. Curr. Res. Microb. Sci. 2022, 3, 100091. [Google Scholar] [CrossRef]
- Komijani, M.; Shamabadi, N.S.; Shahin, K.; Eghbalpour, F.; Tahsili, M.R.; Bahram, M. Heavy Metal Pollution Promotes Antibiotic Resistance Potential in the Aquatic Environment. Environ. Pollut. 2021, 274, 116569. [Google Scholar] [CrossRef]
- Wang, Y.; Lu, S.; Liu, X.; Chen, J.; Han, M.; Wang, Z.; Guo, W. Profiles of Antibiotic Resistance Genes in an Inland Salt-Lake Ebinur Lake, Xinjiang, China: The Relationship with Antibiotics, Environmental Factors, and Microbial Communities. Ecotoxicol. Environ. Saf. 2021, 221, 112427. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Sreekrishnan, T.R.; Ahammad, S.Z. Effects of Heavy Metals on the Development and Proliferation of Antibiotic Resistance in Urban Sewage Treatment Plants. Environ. Pollut. 2022, 308, 119649. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Zhang, J.; Lu, T.; Zhang, K. Metagenomics Revealed the Mobility and Hosts of Antibiotic Resistance Genes in Typical Pesticide Wastewater Treatment Plants. Sci. Total Environ. 2022, 817, 153033. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Cheng, D.; Xue, J.; Feng, Y.; Wakelin, S.A.; Weaver, L.; Shehata, E.; Li, Z. Fate of Bacterial Community, Antibiotic Resistance Genes and Gentamicin Residues in Soil after Three-Year Amendment Using Gentamicin Fermentation Waste. Chemosphere 2022, 291, 132734. [Google Scholar] [CrossRef] [PubMed]
- Niu, Q.; Li, K.; Yang, H.; Zhu, P.; Huang, Y.; Wang, Y.; Li, X.; Li, Q. Exploring the Effects of Heavy Metal Passivation under Fenton-like Reaction on the Removal of Antibiotic Resistance Genes during Composting. Bioresour. Technol. 2022, 359, 127476. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhu, J.; Liu, S.; Luo, Y.; Zhao, R.; Guo, F.; Li, B. Estuarine Salinity Gradient Governs Sedimentary Bacterial Community but Not Antibiotic Resistance Gene Profile. Sci. Total Environ. 2022, 806, 151390. [Google Scholar] [CrossRef]
- Manaia, C.M.; Rocha, J.; Scaccia, N.; Marano, R.; Radu, E.; Biancullo, F.; Cerqueira, F.; Fortunato, G.; Iakovides, I.C.; Zammit, I.; et al. Antibiotic Resistance in Wastewater Treatment Plants: Tackling the Black Box. Environ. Int. 2018, 115, 312–324. [Google Scholar] [CrossRef]
- Li, Z.; Sun, A.; Liu, X.; Chen, Q.-L.; Bi, L.; Ren, P.-X.; Shen, J.-P.; Jin, S.; He, J.-Z.; Hu, H.-W.; et al. Climate Warming Increases the Proportions of Specific Antibiotic Resistance Genes in Natural Soil Ecosystems. J. Hazard. Mater. 2022, 430, 128442. [Google Scholar] [CrossRef]
- Schmeisser, C.; Steele, H.; Streit, W.R. Metagenomics, Biotechnology with Non-Culturable Microbes. Appl. Microbiol. Biotechnol. 2007, 75, 955–962. [Google Scholar] [CrossRef]
- Liu, H.; Taylor, T.H.; Pettus, K.; Johnson, S.; Papp, J.R.; Trees, D. Comparing the Disk-Diffusion and Agar Dilution Tests for Neisseria Gonorrhoeae Antimicrobial Susceptibility Testing. Antimicrob. Resist. Infect. Control. 2016, 5, 46. [Google Scholar] [CrossRef]
- Kuchibiro, T.; Komatsu, M.; Yamasaki, K.; Nakamura, T.; Niki, M.; Nishio, H.; Kida, K.; Ohama, M.; Nakamura, A.; Nishi, I. Evaluation of the VITEK2 AST—XN17 Card for the Detection of Carbapenemase—Producing Enterobacterales in Isolates Primarily Producing Metallo β—Lactamase. Eur. J. Clin. Microbiol. Infect. Dis. 2022, 41, 723–732. [Google Scholar] [CrossRef] [PubMed]
- Zaheer, R.; Noyes, N.; Ortega Polo, R.; Cook, S.R.; Marinier, E.; Van Domselaar, G.; Belk, K.E.; Morley, P.S.; McAllister, T.A. Impact of Sequencing Depth on the Characterization of the Microbiome and Resistome. Sci. Rep. 2018, 8, 5890. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Zhou, X.; Chen, Z.; Deng, X.; Gehring, A.; Ou, H.; Zhang, L.; Shi, X. PRAP: Pan Resistome Analysis Pipeline. BMC Bioinform. 2020, 21, 20. [Google Scholar] [CrossRef]
- Zhuang, M.; Achmon, Y.; Cao, Y.; Liang, X.; Chen, L.; Wang, H.; Siame, B.A.; Leung, K.Y. Distribution of Antibiotic Resistance Genes in the Environment. Environ. Pollut. 2021, 285, 117402. [Google Scholar] [CrossRef] [PubMed]
- Perry, J.A.; Wright, G.D. Forces Shaping the Antibiotic Resistome. BioEssays 2014, 36, 1179–1184. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, D.F.K.; Istvan, P.; Quirino, B.F.; Kruger, R.H. Functional Metagenomics as a Tool for Identification of New Antibiotic Resistance Genes from Natural Environments. Microb. Ecol. 2017, 73, 479–491. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Feng, M.; Yang, K.; Cao, Y.; Zhang, J.; Xu, J.; Hernández, S.H.; Wei, X.; Fan, M. Transcriptomic and Metabolomic Analyses Reveal Antibacterial Mechanism of Astringent Persimmon Tannin against Methicillin-Resistant Staphylococcus Aureus Isolated from Pork. Food Chem. 2020, 309, 125692. [Google Scholar] [CrossRef] [PubMed]
- Yu, K.; Zhang, T. Metagenomic and Metatranscriptomic Analysis of Microbial Community Structure and Gene Expression of Activated Sludge. PLoS ONE 2012, 7, e38183. [Google Scholar] [CrossRef]
- Su, C.; Lei, L.; Duan, Y.; Zhang, K.Q.; Yang, J. Culture-Independent Methods for Studying Environmental Microorganisms: Methods, Application, and Perspective. Appl. Microbiol. Biotechnol. 2012, 93, 993–1003. [Google Scholar] [CrossRef]
- Martinez, J.L. Environmental Pollution by Antibiotics and by Antibiotic Resistance Determinants. Environ. Pollut. 2009, 157, 2893–2902. [Google Scholar] [CrossRef]
- Brain, R.A.; Hanson, M.L.; Solomon, K.R.; Brooks, B.W. Aquatic Plants Exposed to Pharmaceuticals: Effects and Risks. Rev. Environ. Contam. Toxicol. 2008, 192, 67–115. [Google Scholar] [PubMed]
- Jechalke, S.; Heuer, H.; Siemens, J.; Amelung, W.; Smalla, K. Fate and Effects of Veterinary Antibiotics in Soil. Trends Microbiol. 2014, 22, 536–545. [Google Scholar] [CrossRef] [PubMed]
- Boxall, A.B.A.; Rudd, M.A.; Brooks, B.W.; Caldwell, D.J.; Choi, K.; Hickmann, S.; Innes, E.; Ostapyk, K.; Staveley, J.P.; Verslycke, T.; et al. Pharmaceuticals and Personal Care Products in the Environment: What Are the Big Questions? Environ. Health Perspect. 2012, 120, 1221–1229. [Google Scholar] [CrossRef] [PubMed]
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Robles Aguilar, G.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global Burden of Bacterial Antimicrobial Resistance in 2019: A Systematic Analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- Gentilini, F.; Turba, M.E.; Pasquali, F.; Mion, D.; Romagnoli, N.; Zambon, E.; Terni, D.; Peirano, G.; Pitout, J.D.D.; Parisi, A.; et al. Hospitalized Pets as a Source of Carbapenem-Resistance. Front. Microbiol. 2018, 9, 2872. [Google Scholar] [CrossRef] [PubMed]
- Köck, R.; Daniels-Haardt, I.; Becker, K.; Mellmann, A.; Friedrich, A.W.; Mevius, D.; Schwarz, S.; Jurke, A. Carbapenem-Resistant Enterobacteriaceae in Wildlife, Food-Producing, and Companion Animals: A Systematic Review. Clin. Microbiol. Infect. 2018, 24, 1241–1250. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Berçot, B.; Millemann, Y.; Bonnin, R.A.; Pannaux, G.; Nordmann, P. Carbapenemaseproducing Acinetobacter Spp. in Cattle, France. Emerg. Infect. Dis. 2012, 18, 523–525. [Google Scholar] [CrossRef]
- Jansen, K.U.; Knirsch, C.; Anderson, A.S. The Role of Vaccines in Preventing Bacterial Antimicrobial Resistance. Nat. Med. 2018, 24, 10–19. [Google Scholar] [CrossRef]
- Van Boeckel, T.; Pires, J.; Silvester, R.; Zhao, C.; Song, J.; Criscuolo, N.; Gilbert, M.; Bonhoeffer, S.; Laxminarayan, R. Global Trends in Antimicrobial Resistance in Animals in Low-and Middle-Income Countries. Int. J. Infect. Dis. 2020, 101, 19. [Google Scholar] [CrossRef]
- Marshall, B.M.; Levy, S.B. Food Animals and Antimicrobials: Impacts on Human Health. Clin. Microbiol. Rev. 2011, 24, 718–733. [Google Scholar] [CrossRef]
- Renault, V.; Humblet, M.F.; Saegerman, C. Biosecurity Concept: Origins, Evolution and Perspectives. Animals 2022, 12, 63. [Google Scholar] [CrossRef] [PubMed]
- Callaway, T.R.; Lillehoj, H.; Chuanchuen, R.; Gay, C.G. Erratum: Callaway et Al. Alternatives to Antibiotics: A Symposium on the Challenges and Solutions for Animal Health and Production. Antibiotics 2021, 10, 1024. [Google Scholar] [CrossRef] [PubMed]
- Holmes, A.H.; Moore, L.S.P.; Sundsfjord, A.; Steinbakk, M.; Regmi, S.; Karkey, A.; Guerin, P.J.; Piddock, L.J. V Understanding the Mechanisms and Drivers of Antimicrobial Resistance. Lancet 2016, 387, 176–187. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Antibacterial Agents in Clinical Development: An Analysis of the Antibacterial Clinical Development Pipeline; WHO: Geneva, Switzerland, 2019. [Google Scholar]



| Year of Discovery | Microorganisms | Synthetic Antimicrobials | ||
|---|---|---|---|---|
| Actinomycetes | Bacteria | Fungi | ||
| 1910–1940 | Salvarsan Sulfonamides sufapyridine | |||
| 1940 | Streptomycin Aminoglycosides Tetrecyclines Amphenicols | Polypeptides Bacitracin | Penicillins | Sulfones Salicylates |
| 1950 | Macrolides Glycopeptides Tuberactinomycins | Polymyxins | Nitrofurans Pyridinamides | |
| 1960 | Ansamycins Lincosamides Streptogramins Cycloserine | Fusidic acid Cephalosporins Enniatins | Quinolones Azoles Thioamides Ethambutol Phenazines Diaminopyrimidines | |
| 1970 | Phosphonates Fosfomycin | |||
| 1980 | Carbapenems | Mupirocin Monobactams | ||
| 2000 | Lipopetides | Pleuromutilins | Oxazolidinones linezolid | |
| 2010 | Liparmycins | Diarylquinolines | ||
| Antimicrobial Agents | Mechanism of Action | Examples | References |
|---|---|---|---|
| Penicillins and cephalosporins | Enzymatic inactivation by β-lactamase; enzymatic modification by acylase and esterase; outer membrane protein deletion; alteration of penicillin-binding proteins | β-lactamase containing gram-negative rods | [117,118,119] |
| Monobactams | Enzymatic inactivation by β-lactamase | Haemophilus influenza; Pseudomonas aeruginosa | [120,121] |
| Carbapenems | Enzymatic inactivation by β-lactamase; outer-membrane protein deletion | Neisseria gonorrhoea; A. baumannii; Citrobacter portucalensis; K. pneumoniae; Escherichia coli | [122,123] |
| Carbacephems | Cell wall synthesis inhibition | ESBL-producing Enterobacteria | [124] |
| Imipenem | Decreased Cell membrane permeability | Pseudomonas sp.; K. | [125,126] |
| Vancomycin | Inhibition of glycopeptides access | S. aureus; Enterococcus sp. | [127,128] |
| Trimethoprim | Increased production of dihydrofolate reductase; production of trimethoprim-insensitive dihydrofolate reductase | Streptococcus agalactiae; E. coli; Burkholderia pseudomallei | [129,130,131] |
| Sulfonamides | Increased production of p-aminobenzoic acid; increased production of pteridine; increased production of sulfonamide-insensitive dihydropteroate synthetase | Haemophilus influenza; S. pneumoniae; S. pyogenes; Neisseria meningitidis | [132] |
| Aminoglycosides | Enzymatic modification by acetylation, phosphorylation, and nucleotidylation; ribosomal alteration; diminished drug uptake | Clostridium perfringens; Bacteroides fragilis; S. aureus; Bacillus cereus | [133,134,135] |
| Chloramphenicol | Enzymatic inactivation by acetylation; decreased drug permeability | Streptomyces venezuelae; Pseudomonas putida; Pneumococcus sp.; E. coli | [136,137,138,139] |
| Macrolides | Enzymatic modification by esterase; alteration of 23S ribosomal RNA | S. pneumoniae, S. aureus | [137,140] |
| Lincosamides | Enzymatic modification by nucleotidylation or phosphorylation; alteration of 23S ribosomal RNA | S. pneumoniae; S. agalactiae; Acinetobacter baumannii | [140,141,142] |
| Tetracyclines | Active efflux preceded by chemical modification; ribosomal alterations | E. coli, Shigella sp., S. pneumoniae, S.s aureus, Clostridium perfringens, Helicobacter pylori | [143,144] |
| Quinolones | Alteration of subunit A of DNA gyrase; decreased drug permeability | Stenotrophomonas maltophilia; Pseudomonas species; Enterobacteriaceae | [145,146,147] |
| Threat Status | Organism | Estimated Clinical Cases Per Year | Estimated Healthcare Cost (US Dollars) | Descriptions |
|---|---|---|---|---|
| Urgent | Carbapenem-resistant Acinetobacter | 8500 (700) | 281 million | Carbapenem-resistant Acinetobacter causes pneumonia and wound, bloodstream, and urinary tract infections. Nearly all these infections happen in patients who recently received care in a healthcare facility |
| Clostridioides difficile (C. difficile) | 223,900 (12,800) | 1 billion | C. difficile causes life-threatening diarrhoea and colitis (inflammation of the colon), mostly in people who have had both recent medical care and antibiotics. | |
| Carbapenem-resistant Enterobacterales (CRE) | 13,100 (1100) | 130 million | CRE are a major concern for patients in healthcare facilities. Some Enterobacterales are resistant to nearly all antibiotics, leaving more toxic or less effective treatment options. | |
| Drug-resistant N. gonorrhoeae (N. gonorrhoeae) | 550,000 | 133.4 million | N. gonorrhoeae causes the sexually transmitted disease Gonorrhoea that can result in life-threatening ectopic pregnancy and infertility and can increase the risk of getting and giving HIV. | |
| Serious | Drug-resistant Campylobacter | 448,400 (70) | 270 million | Campylobacter usually causes diarrhoea (often bloody), fever, and abdominal cramps and can spread from animals to people through contaminated food, especially raw or undercooked chicken |
| ESBL-producing Enterobacterales | 197,400 (9100) | 1.2 billion | ESBL-producing Enterobacterales are a concern in healthcare settings and the community. They can spread rapidly and cause or complicate infections in healthy people. ESBLs are enzymes that break down commonly used antibiotics, such as penicillins and cephalosporins, making them ineffective. | |
| Vancomycin-resistant Enterococcus (VRE) | 54,500 (5400) | 539 million | Enterococci can cause severe infections for patients in healthcare settings, including bloodstream, surgical site, and urinary tract infections. | |
| Multidrug-resistant Pseudomonas aeruginosa (P. aeruginosa) | 32,600 (2700) | 767 million | P. aeruginosa infections usually occur in people with weakened immune systems and can be particularly dangerous for patients with chronic lung diseases. | |
| Drug-resistant non-typhoidal Salmonella | 212,500 (70) | 400 million | Non-typhoidal Salmonella can spread from animals to people through food and usually causes diarrhoea, fever, and abdominal cramps. Some infections spread to the blood and can have life-threatening complications. | |
| Drug-resistant Salmonella serotype Typhi | 4100 (<5) | 11 to 21 million | Salmonella Typhi causes severe typhoid fever, which can be life-threatening. Most people in the U.S. become infected while traveling to countries where the disease is common. | |
| Drug-resistant Shigella | 77,000 (<5) | 93 million | Shigella spreads in feces through direct contact or contaminated surfaces, food, or water. Most people with Shigella infections develop diarrhoea, fever, and stomach cramps. | |
| Methicillin-resistant S. aureus (MRSA) | 323,700 (10,600) | 1.7 billion | S. aureus are common bacteria that spread in healthcare facilities and the community. In addition, MRSA can cause difficult-to-treat staph infections because of resistance to some antibiotics. | |
| Drug-resistant S. pneumoniae | 900,000 (3600) | 4 billion | S. pneumoniae causes pneumococcal disease, ranging from ear and sinus infections to pneumonia and bloodstream infections | |
| Drug-resistant Tuberculosis | 847 (62) | 1.6 million | TB is caused by M. tuberculosis. It is among the most common infectious diseases and a frequent cause of death worldwide. | |
| Concerning | Erythromycin-resistant Group A Streptococcus (GAS) | 5400 (450) | 2.6 million | GAS can cause many infections ranging from minor illnesses to severe and deadly diseases, including strep throat, pneumonia, flesh-eating infections, and sepsis. |
| Clindamycin-resistant Group B Streptococcus (GBS) | 13,000 (720) | NA | GBS can cause severe illness in people of all ages. | |
| Watch list | Azole-resistant Aspergillus fumigatus | NA | NA | Aspergillus fumigatus, a ubiquitously distributed opportunistic pathogen, is the leading agent of aspergillosis, ranking first among fungal killers. |
| Drug-resistant Mycoplasma genitalium | Mycoplasma genitalium is one of the important causes of non-gonococcal urethritis. | |||
| Drug-resistant Bordetella perstussis | Pertussis (whooping cough), a highly contagious respiratory illness caused by Bordetella pertussis |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Selvarajan, R.; Obize, C.; Sibanda, T.; Abia, A.L.K.; Long, H. Evolution and Emergence of Antibiotic Resistance in Given Ecosystems: Possible Strategies for Addressing the Challenge of Antibiotic Resistance. Antibiotics 2023, 12, 28. https://doi.org/10.3390/antibiotics12010028
Selvarajan R, Obize C, Sibanda T, Abia ALK, Long H. Evolution and Emergence of Antibiotic Resistance in Given Ecosystems: Possible Strategies for Addressing the Challenge of Antibiotic Resistance. Antibiotics. 2023; 12(1):28. https://doi.org/10.3390/antibiotics12010028
Chicago/Turabian StyleSelvarajan, Ramganesh, Chinedu Obize, Timothy Sibanda, Akebe Luther King Abia, and Haijun Long. 2023. "Evolution and Emergence of Antibiotic Resistance in Given Ecosystems: Possible Strategies for Addressing the Challenge of Antibiotic Resistance" Antibiotics 12, no. 1: 28. https://doi.org/10.3390/antibiotics12010028
APA StyleSelvarajan, R., Obize, C., Sibanda, T., Abia, A. L. K., & Long, H. (2023). Evolution and Emergence of Antibiotic Resistance in Given Ecosystems: Possible Strategies for Addressing the Challenge of Antibiotic Resistance. Antibiotics, 12(1), 28. https://doi.org/10.3390/antibiotics12010028