
Antibacterial Fractions from Erodium cicutarium Exposed—Clinical Strains of Staphylococcus aureus in Focus

 ,
,  , , ,
, , , 

Abstract
:1. Introduction
2. Results and Discussion
2.1. Serial Microdilution Broth Assay and Agar Well Diffusion Assay
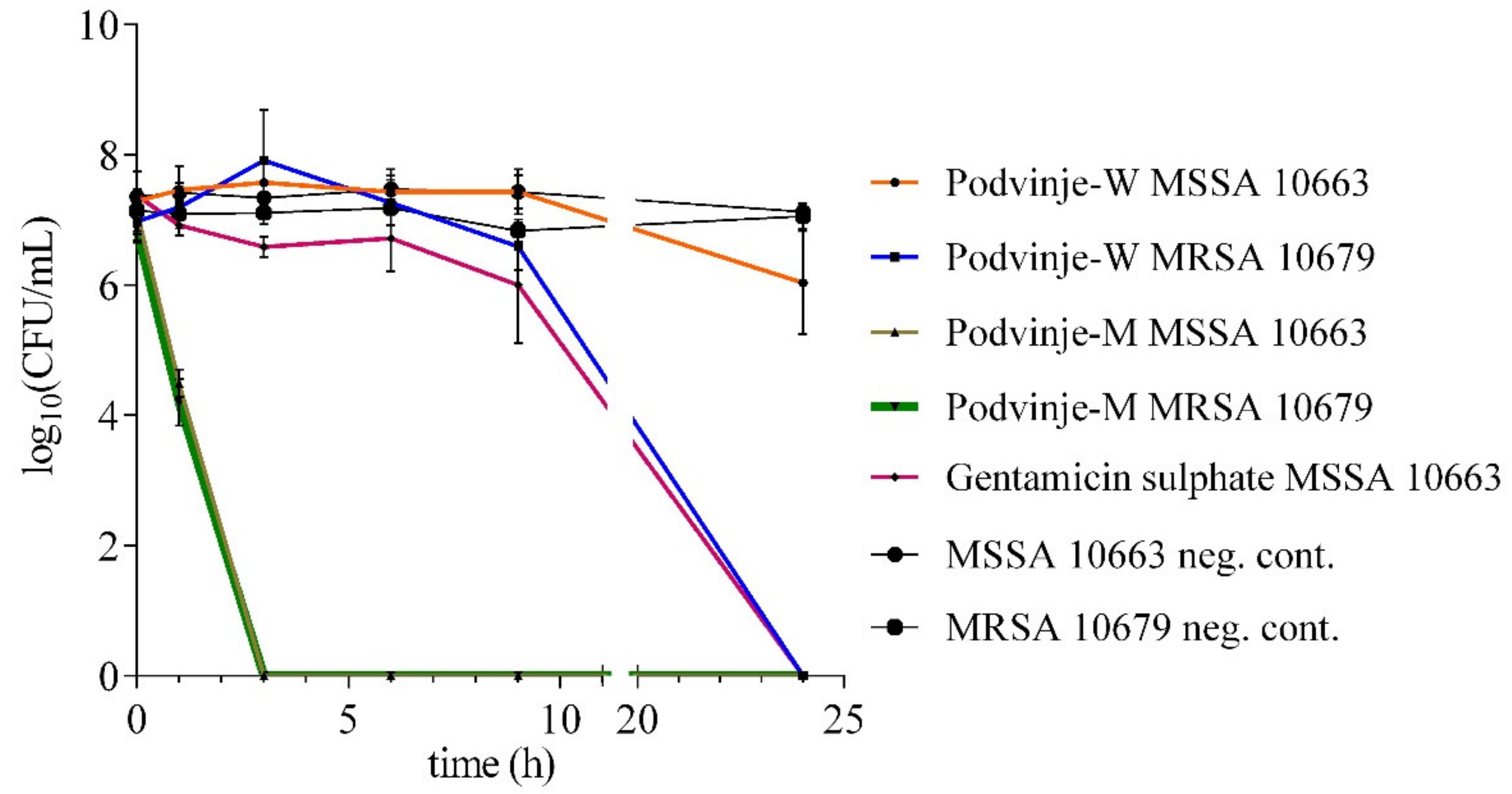
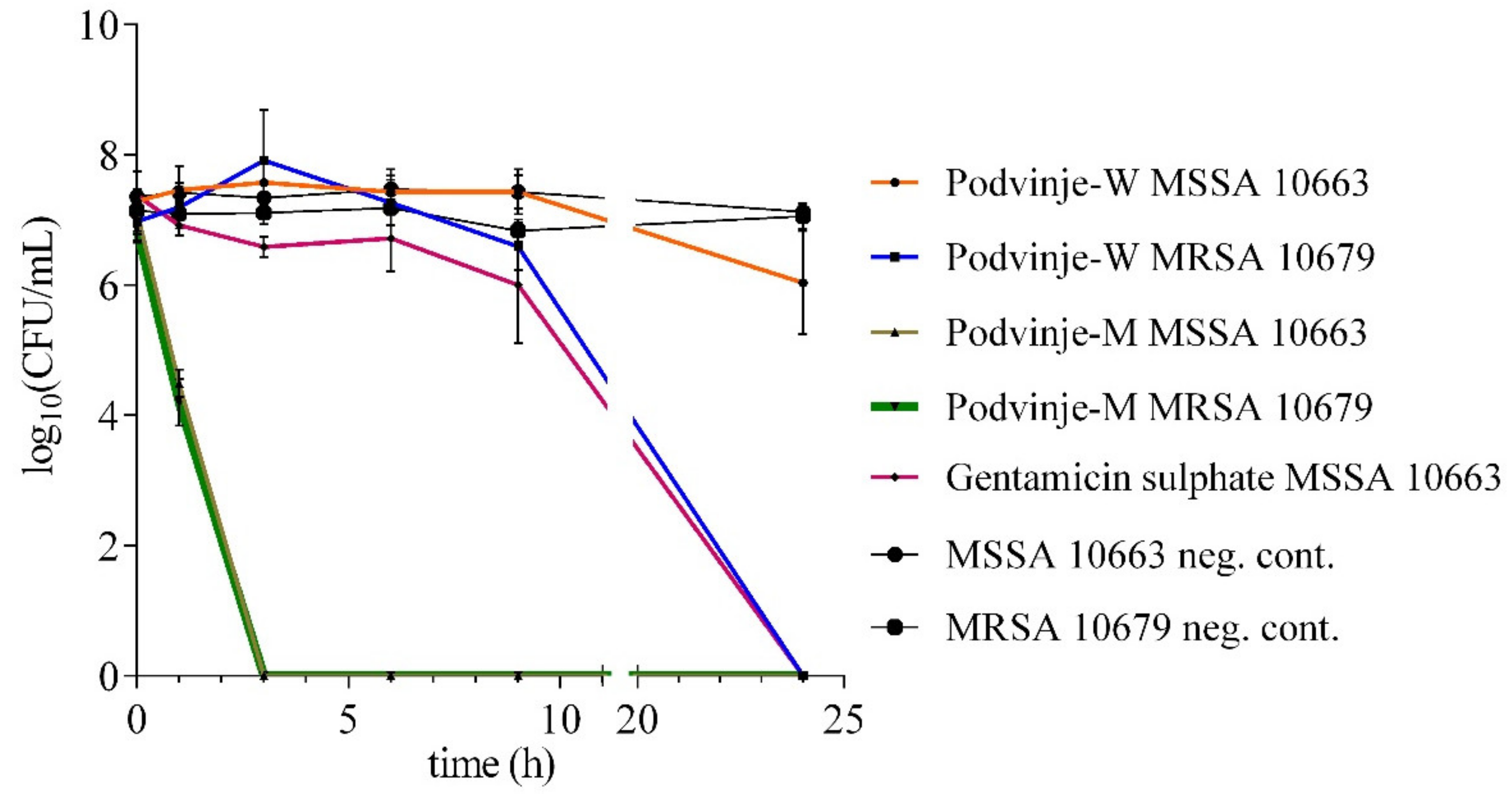
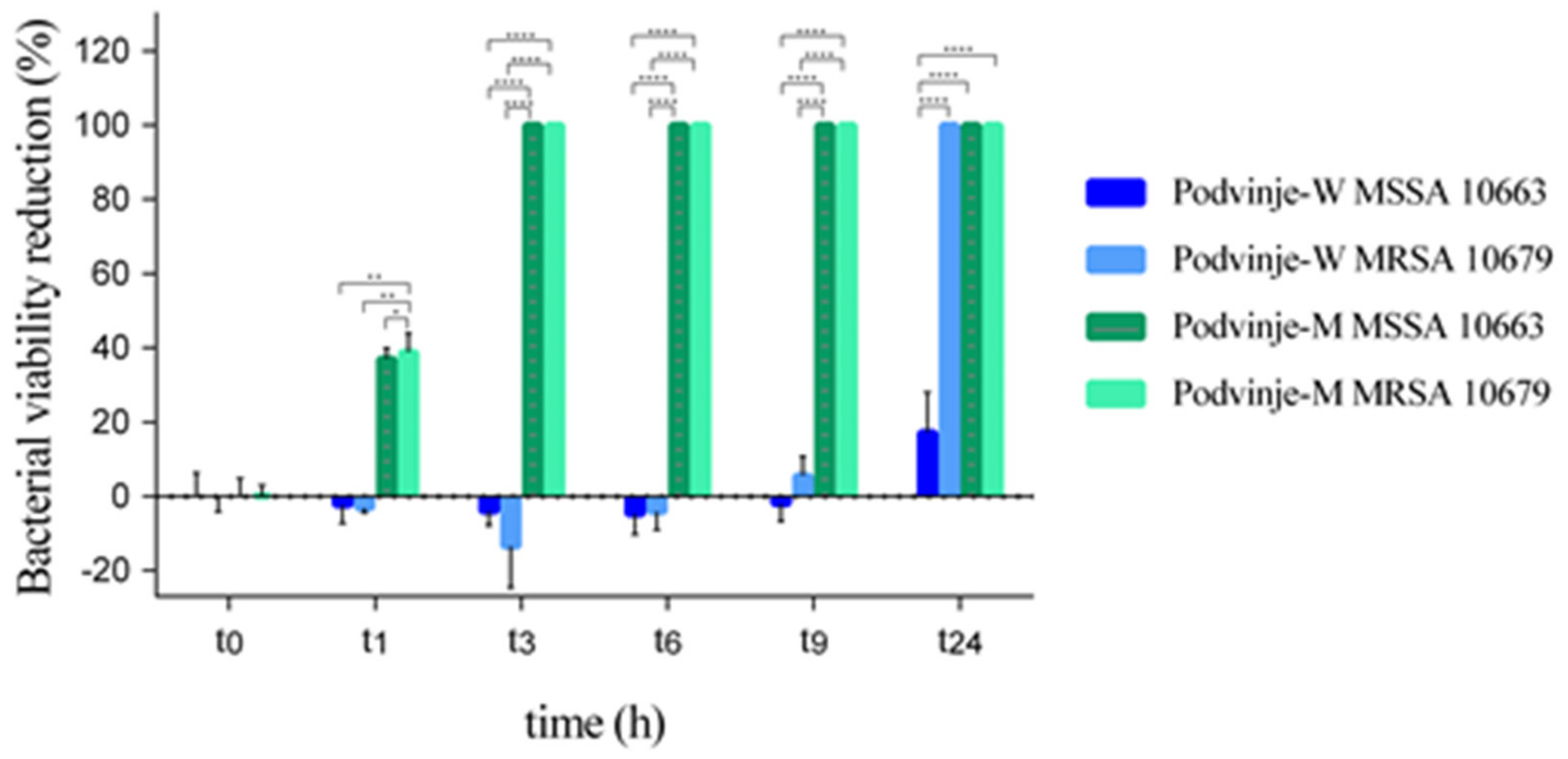
2.2. “Time-Kill” Assay
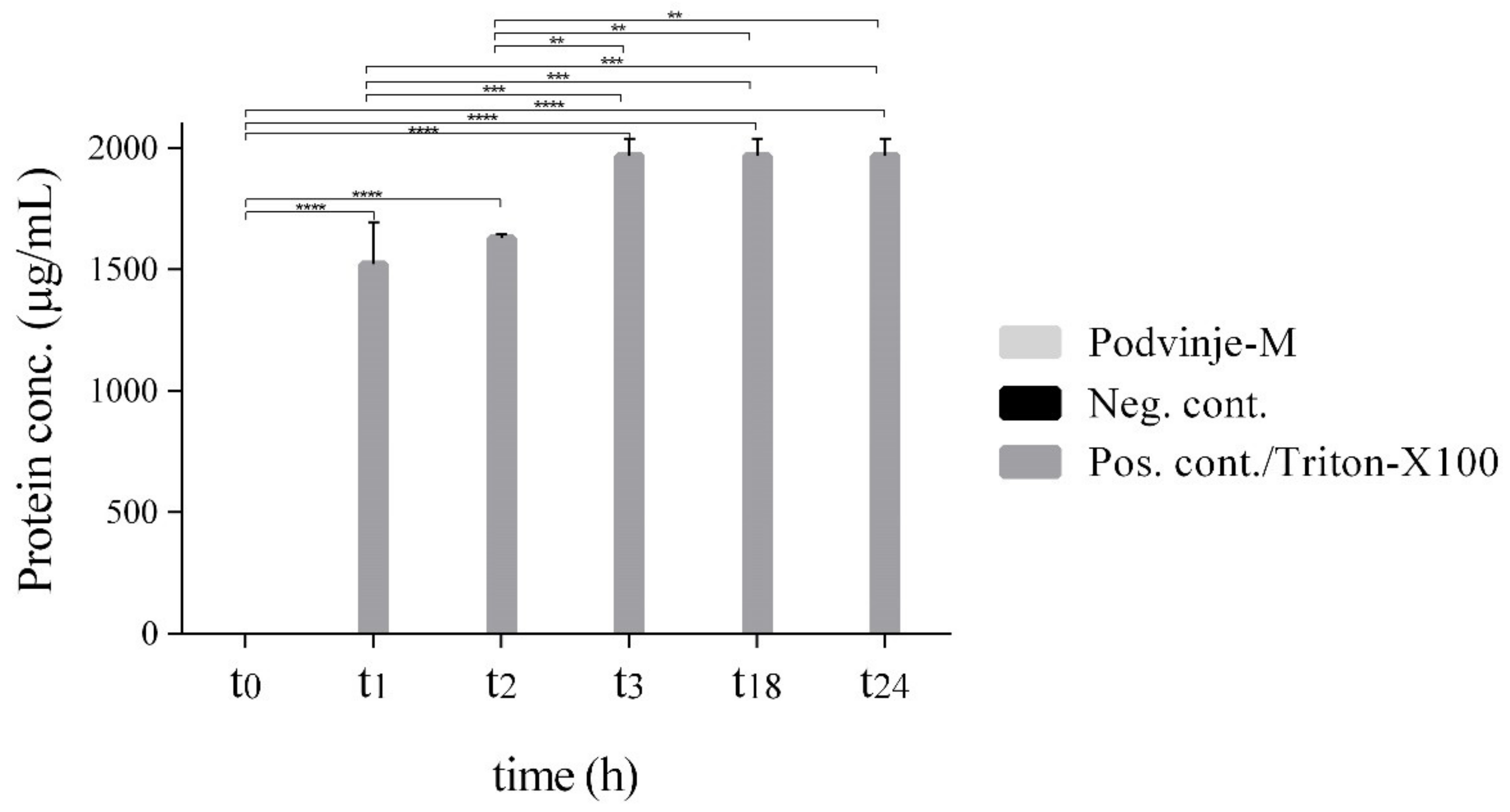
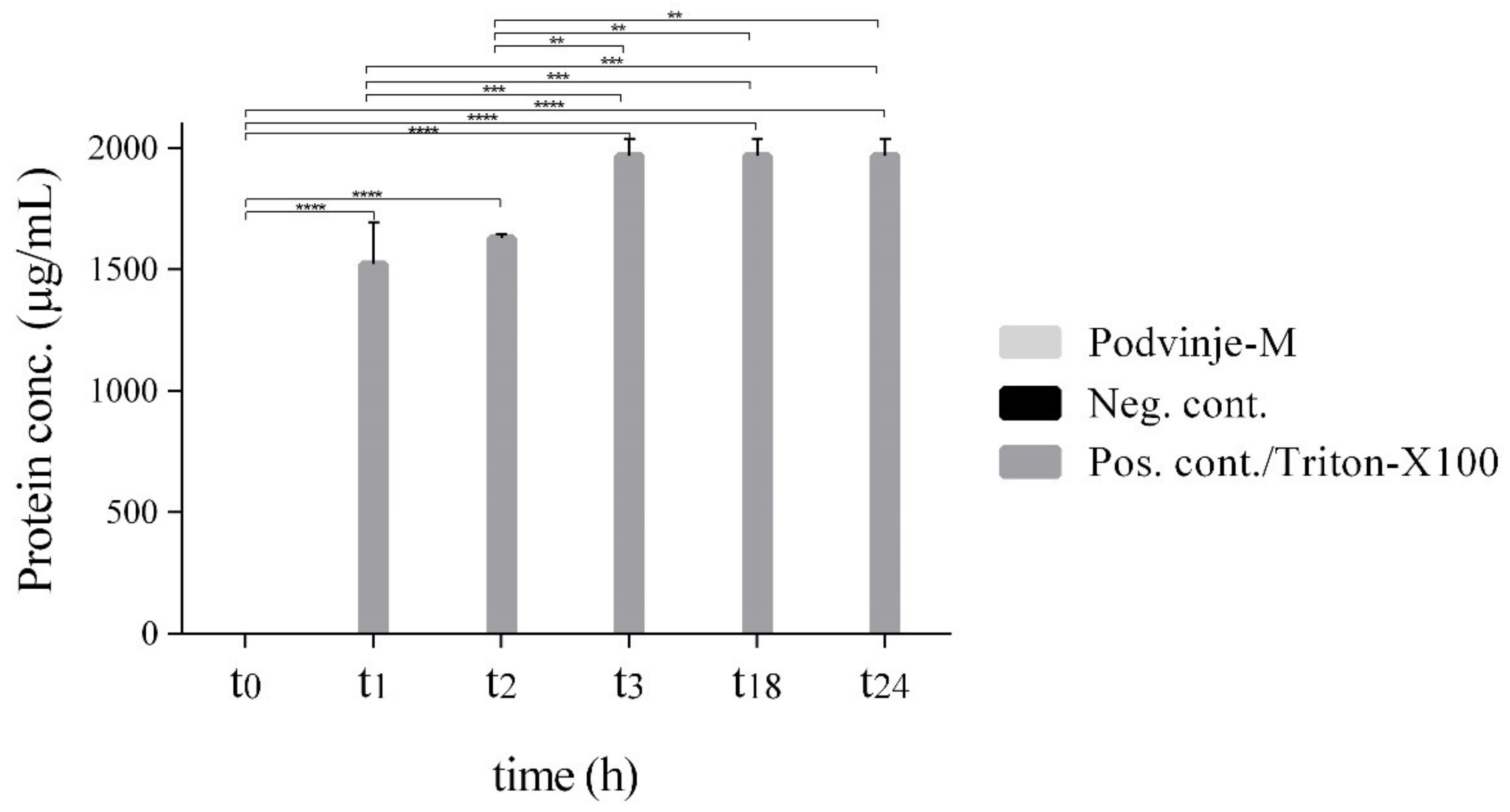
2.3. Modulation of Cell Wall Integrity
2.4. Evaluation of Anti–Biofilm Activity
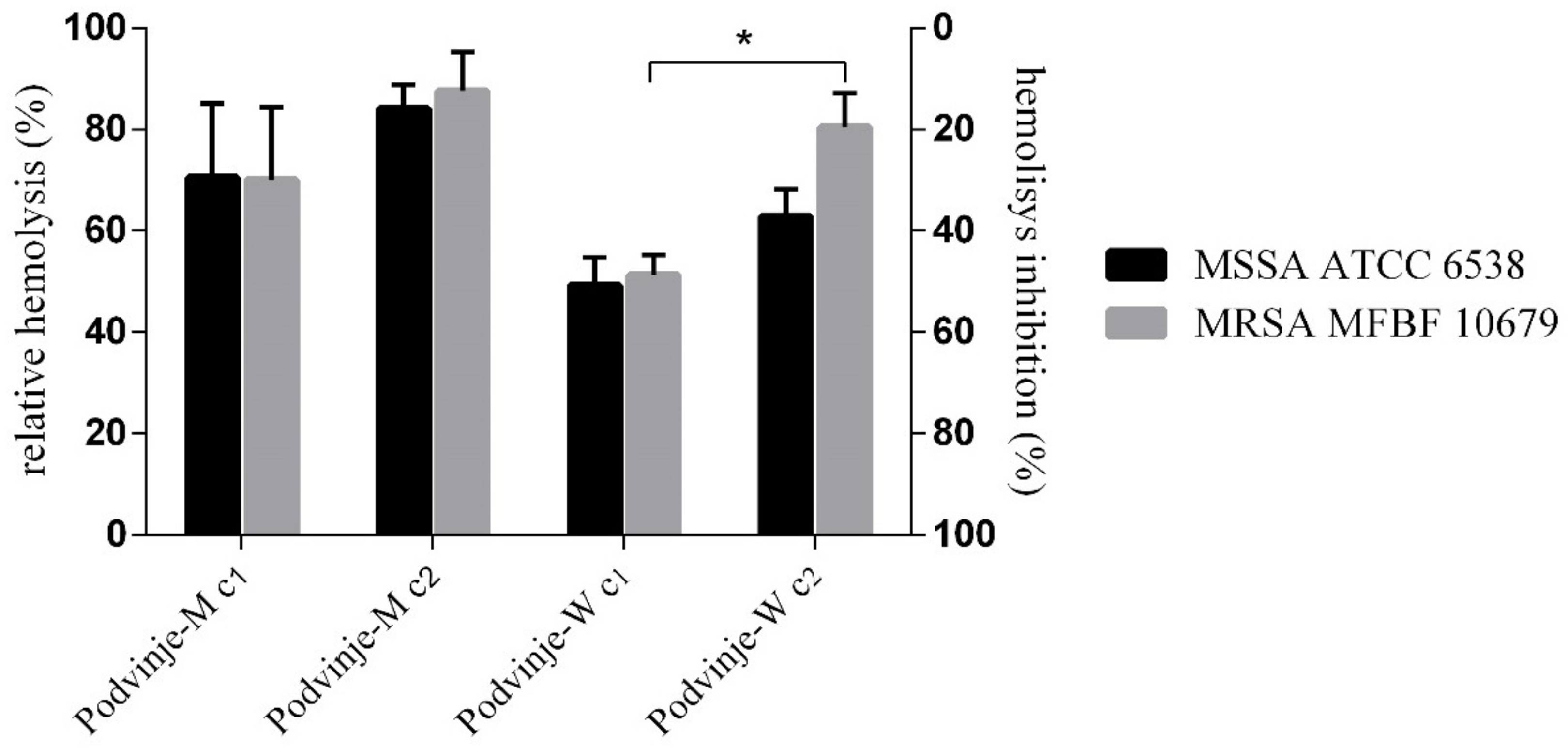
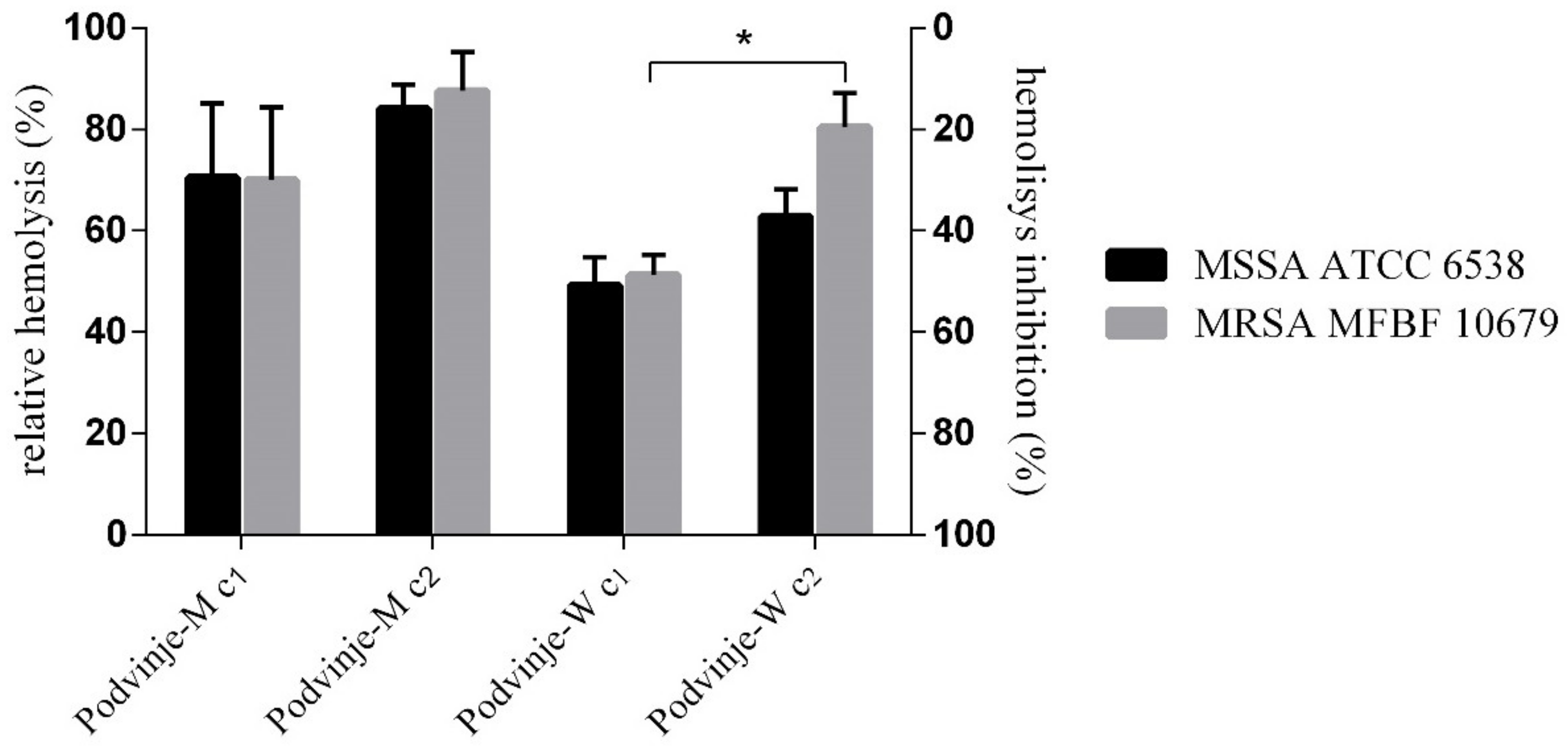
2.5. Modulation of Bacterial Hemolysis
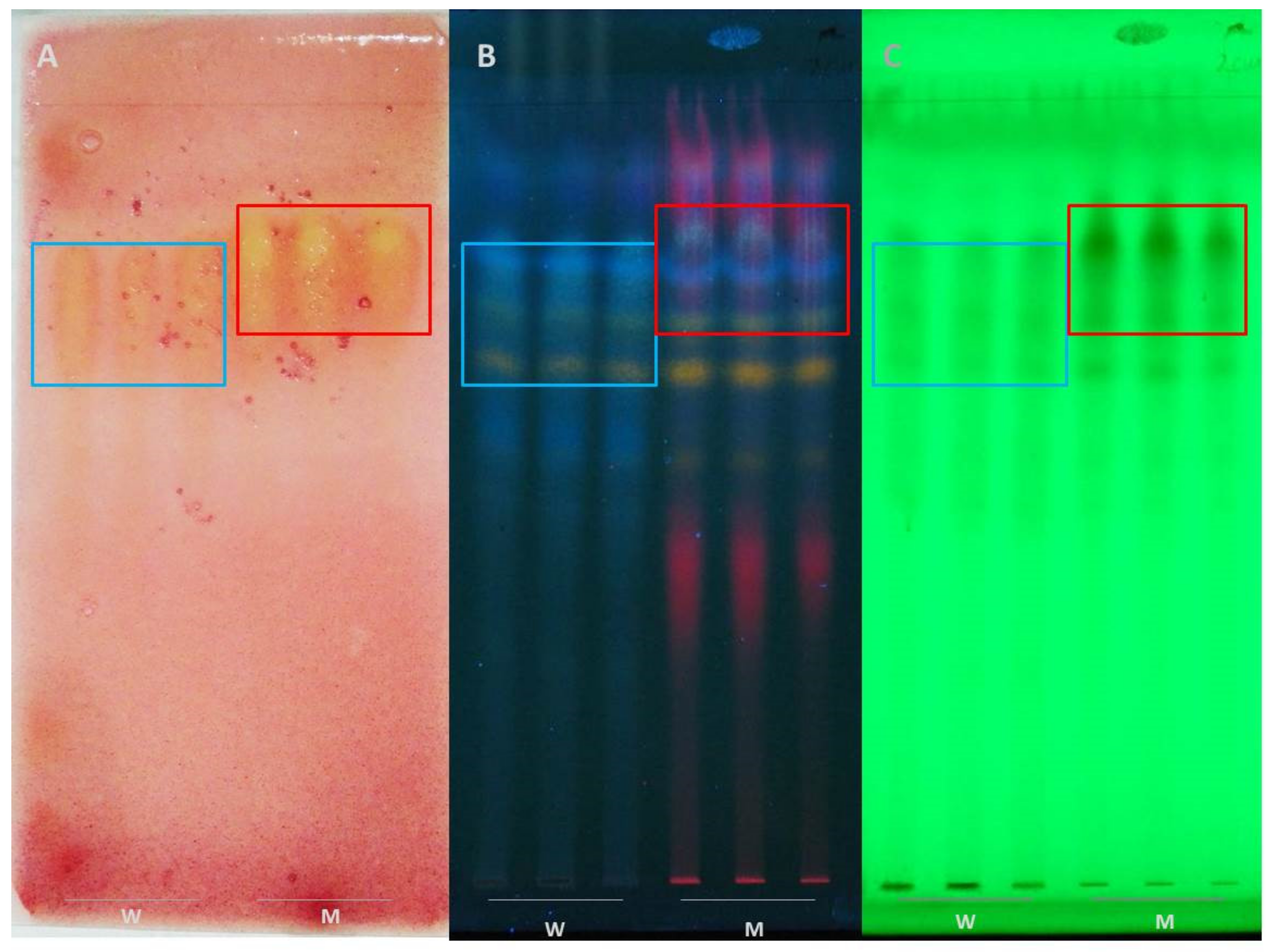
2.6. TLC and Bioautography In Situ
3. Materials and Methods
3.1. Plant Material and Sample Preparation
3.2. Microbial Species, Media and Positive Controls
3.3. Antimicrobial Susceptibility Testing of Clinical S. aureus Strains
3.4. Agar Well Diffusion Assay
3.5. Serial Microdilution Broth Assay
3.6. “Time-Kill” Assay
3.7. Modulation of Cell Wall Integrity
3.8. Evaluation of Anti–Biofilm Activity
3.9. Anti-Hemolytic Activity of Extracts
- S. aureus ATCC 6538 → Podvinje-M: c1 (MIC/2) = 1.88 mg/mL and c2 (MIC/4) = 0.94 mg/mL; Podvinje-W c1 (MIC/2) = 4.17 mg/mL and c2 (MIC/4) = 2.08 mg/mL
- MRSA MFBF 10679 → Podvinje-M: c1 (MIC/2) = 1.25 mg/mL and c2 (MIC/4) = 0.63 mg/mL; Podvinje-W c1 (MIC/2) = 2.50 mg/mL and c2 (MIC/4) = 1.25 mg/mL.
3.10. TLC and Bioautography
3.10.1. TLC
3.10.2. TLC-Bioautography
3.10.3. Semi-Preparative TLC and UHPLC-LTQ OrbiTrap MS4
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Abreu, A.C.; McBain, A.J.; Simões, M. Plants as sources of new antimicrobials and resistance-modifying agents. Nat. Prod. Rep. 2012, 29, 1007–1021. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, A.; Upadhyaya, I.; Kollanoor-Johny, A.; Venkitanarayanan, K. Combating pathogenic microorganisms using plant-derived antimicrobials: A minireview of the mechanistic basis. Biomed. Res. Int. 2014, 2014, 761741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lis-Balchin, M.; Hart, S.L. A pharmacological appraisal of the folk medicinal usage of Pelargonium grossularioides and Erodium cicutarium. J. Herbs Spices Med. Plants 1994, 2, 41–48. [Google Scholar] [CrossRef]
- Fiz-Palacios, O.; Vargas, P.; Vila, R.; Papadopulos, A.S.T.; Aldasoro, J.J. The uneven phylogeny and biogeography of Erodium (Geraniaceae): Radiations in the Mediterranean and recent recurrent intercontinental colonization. Ann. Bot. 2010, 106, 871–884. [Google Scholar] [CrossRef] [Green Version]
- Evangelista, D.; Hotton, S.; Dumais, J. The mechanics of explosive dispersal and self-burial in the seeds of the filaree, Erodium cicutarium (Geraniaceae). J. Exp. Biol. 2011, 214, 521–529. [Google Scholar] [CrossRef] [Green Version]
- Ljoljić Bilić, V.; Stabentheiner, E.; Kremer, D.; Dunkić, V.; Jurišić Grubešić, R.; Vuković Rodríguez, J. Phytochemical and micromorphological characterization of Croatian populations of Erodium cicutarium. Nat. Prod. Commun. 2019, 14, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Fecka, I.; Kowalczyk, A.; Cisowski, W. Phenolic acids and depsides from some species of the Erodium genera. Z. Naturforsch. Sect. C J. Biosci. 2001, 56, 943–950. [Google Scholar] [CrossRef]
- Radulović, N.; Dekić, M.; Stojanović-Radić, Z.; Palić, R. Volatile constituents of Erodium cicutarium (L.) L’ Hérit. (Geraniaceae). Cent. Eur. J. Biol. 2009, 4, 404–410. [Google Scholar] [CrossRef]
- Stojanović-Radić, Z.; Čomić, L.; Radulović, N.; Dekić, M.; Ranđelović, V.; Stefanović, O. Chemical composition and antimicrobial activity of Erodium species: E. ciconium L., E. cicutarium L., and E. absinthoides Willd. (Geraniaceae). Chem. Pap. 2010, 64, 368–377. [Google Scholar] [CrossRef]
- Nikitina, V.S.; Kuz’mina, L.Y.; Melent’ev, A.I.; Shendel’, G. V Antibacterial activity of polyphenolic compounds isolated from plants of Geraniaceae and Rosaceae families. Prikl. Biokhim. Mikrobiol. 2007, 43, 705–712. [Google Scholar] [CrossRef]
- Fecka, I.; Cisowski, W. TLC determination of tannins and flavonoids in extracts from some Erodium species using chemically modified stationary phases. Jpc J. Planar Chromatogr. Mod. Tlc 2002, 15, 429–432. [Google Scholar] [CrossRef]
- Munekata, P.E.S.; Alcántara, C.; Collado, M.C.; Garcia-Perez, J.V.; Saraiva, J.A.; Lopes, R.P.; Barba, F.J.; do Prado Silva, L.; Sant’Ana, A.S.; Fierro, E.M.; et al. Ethnopharmacology, phytochemistry and biological activity of Erodium species: A review. Food Res. Int. 2019, 126, 108659. [Google Scholar] [CrossRef]
- Fecka, I.; Cisowski, W. Tannins and flavonoids from the Erodium cicutarium herb. Z. Naturforsch. Sect. B J. Chem. Sci. 2005, 60, 555–560. [Google Scholar] [CrossRef]
- Ljoljić Bilić, V.; Gašić, U.; Milojković-Opsenica, D.; Nemet, I.; Rončević, S.; Kosalec, I.; Vuković Rodriguez, J. First extensive polyphenolic profile of Erodium cicutarium with novel insights to elemental composition and antioxidant activity. Chem. Biodivers. 2020, 17, e2000280. [Google Scholar] [CrossRef]
- Sroka, Z.; Rzadkowska-Bodalska, H.; Mazol, I. Antioxidative effect of extracts from Erodium cicutarium. Z. Naturforsch. Sect. C J. Biosci. 1994, 49, 881–884. [Google Scholar] [CrossRef]
- Sarikurkcu, C.; Targan, S.; Ozer, M.S.; Tepe, B. Fatty acid composition, enzyme inhibitory, and antioxidant activities of the ethanol extracts of selected wild edible plants consumed as vegetables in the Aegean region of Turkey. Int. J. Food Prop. 2017, 20, 560–572. [Google Scholar] [CrossRef] [Green Version]
- Bussmann, R.W.; Malca-García, G.; Glenn, A.; Sharon, D.; Chait, G.; Díaz, D.; Pourmand, K.; Jonat, B.; Somogy, S.; Guardado, G.; et al. Minimum inhibitory concentrations of medicinal plants used in Northern Peru as antibacterial remedies. J. Ethnopharmacol. 2010, 132, 101–108. [Google Scholar] [CrossRef] [Green Version]
- Gohar, A.A.; Lahloub, M.F.; Niwa, M. Antibacterial polyphenol from Erodium glaucophyllum. Z. Naturforsch. Sect. C J. Biosci. 2003, 58, 670–674. [Google Scholar] [CrossRef]
- Lis-Balchin, M. The essential oils of Pelargonium grossularioides and Erodium cicutarium (Geraniaceae). J. Essent. Oil Res. 1993, 5, 317–318. [Google Scholar] [CrossRef]
- Choubey, S.; Goyal, S.; Varughese, L.R.; Kumar, V.; Sharma, A.K.; Beniwal, V. Probing gallic acid for its broad spectrum applications. Mini-Rev. Med. Chem. 2018, 18, 1283–1293. [Google Scholar] [CrossRef]
- Borges, A.; Ferreira, C.; Saavedra, M.J.; Simões, M. Antibacterial activity and mode of action of ferulic and gallic acids against pathogenic bacteria. Microb. Drug Resist. 2013, 19, 256–265. [Google Scholar] [CrossRef]
- Kang, M.S.; Oh, J.S.; Kang, I.C.; Hong, S.J.; Choi, C.H. Inhibitory effect of methyl gallate and gallic acid on oral bacteria. J. Microbiol. 2008, 46, 744–750. [Google Scholar] [CrossRef]
- Beoletto, V.G.; de las Mercedes Oliva, M.; Marioli, J.M.; Carezzano, M.E.; Demo, M.S. Antimicrobial Natural Products against Bacterial Biofilms. In Antibiotic Resistance: Mechanisms and New Antimicrobial Approaches; Kon, K., Rai, M., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 291–308. ISBN 9780128036426. [Google Scholar]
- Lee, D.S.; Je, J.Y. Gallic acid-grafted-chitosan inhibits foodborne pathogens by a membrane damage mechanism. J. Agric. Food Chem. 2013, 61, 6574–6579. [Google Scholar] [CrossRef]
- Shibata, H.; Kondo, K.; Katsuyama, R.; Kawazoe, K.; Sato, Y.; Murakami, K.; Takaishi, Y.; Arakaki, N.; Higuti, T. Alkyl gallates, intensifiers of β-lactam susceptibility in methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2005, 49, 549–555. [Google Scholar] [CrossRef] [Green Version]
- De, R.; Sarkar, A.; Ghosh, P.; Ganguly, M.; Karmakar, B.C.; Saha, D.R.; Halder, A.; Chowdhury, A.; Mukhopadhyay, A.K. Antimicrobial activity of ellagic acid against Helicobacter pylori isolates from India and during infections in mice. J. Antimicrob. Chemother. 2018, 73, 1595–1603. [Google Scholar] [CrossRef]
- Lipińska, L.; Klewicka, E.; Sójka, M. Structure, occurrence and biological activity of ellagitannins: A general review. Acta Sci. Pol. Technol. Aliment. 2014, 13, 289–299. [Google Scholar] [CrossRef] [Green Version]
- Puljula, E.; Walton, G.; Woodward, M.J.; Karonen, M. Antimicrobial activities of ellagitannins against Clostridiales perfringens, Escherichia coli, Lactobacillus plantarum and Staphylococcus aureus. Molecules 2020, 25, 3714. [Google Scholar] [CrossRef]
- Cowan, M.M. Plant products as antimicrobial agents. Clin. Microbiol. Rev. 1999, 12, 564–582. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Luo, M.; Fu, Y.J.; Zu, Y.G.; Wang, W.; Zhang, L.; Yao, L.P.; Zhao, C.J.; Sun, Y. Effect of corilagin on membrane permeability of Escherichia coli, Staphylococcus aureus and Candida albicans. Phytother. Res. 2013, 27, 1517–1523. [Google Scholar] [CrossRef]
- Junkins, A.D.; Lockhart, S.R.; Heilmann, K.P.; Dohrn, C.L.; Von Stein, D.L.; Winokur, P.L.; Doern, G.V.; Richter, S.S. BD Phoenix and Vitek 2 detection of mecA-mediated resistance in Staphylococcus aureus with cefoxitin. J. Clin. Microbiol. 2009, 47, 2879–2882. [Google Scholar] [CrossRef] [Green Version]
- European Pharmacopoeia Commission. European Pharmacopoeia, 5th ed.; Council Of Europe: Strasbourg, France, 2005; pp. 188–191. [Google Scholar]
- European Committee for Antimicrobial Susceptibility Testing (EUCAST) of the European Society for Clinical Microbiology and Infectious Diseases (ESCMID). EUCAST Determination of minimum inhibitory concentrations (MICs) of antibacterial agents by broth dilution. Clin. Microbiol. Infect. 2003, 9, 9–15. [Google Scholar] [CrossRef] [Green Version]
- Arendrup, M.C.; Meletiadis, J.; Mouton, J.W.; Lagrou, K.; Hamal, P.; Guinea, J.; Subcommittee on Antifungal Susceptibility Testing (AFST) of the ESCMID European Committee for Antimicrobial Susceptibility Testing (EUCAST). Definitive Document E. Def 7.3.1. January 2017—Method for the Determination of Broth Dilution Minimum Inhibitory Concentrations of Antifungal AGENTS for yeasts. Available online: http://www.eucast.org/ast_of_fungi/methodsinantifungalsusceptibilitytesting/susceptibility_testing_of_yeasts/ (accessed on 16 July 2021).
- Jakas, A.; Vlahoviček-Kahlina, K.; Ljolić-Bilić, V.; Horvat, L.; Kosalec, I. Design and synthesis of novel antimicrobial peptide scaffolds. Bioorg. Chem. 2020, 103, 104178. [Google Scholar] [CrossRef] [PubMed]
- Zorić, N.; Kosalec, I.; Tomić, S.; Bobnjarić, I.; Jug, M.; Vlainić, T.; Vlainić, J. Membrane of Candida albicans as a target of berberine. BMC Complement. Altern. Med. 2017, 17, 268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vlainić, J.; Kosalec, I.; Pavić, K.; Hadjipavlou-Litina, D.; Pontiki, E.; Zorc, B. Insights into biological activity of ureidoamides with primaquine and amino acid moieties. J. Enzyme Inhib. Med. Chem. 2018, 33, 376–382. [Google Scholar] [CrossRef] [Green Version]
- Ferro, T.A.F.; Araújo, J.M.M.; dos Santos Pinto, B.L.; dos Santos, J.S.; Souza, E.B.; da Silva, B.L.R.; Colares, V.L.P.; Novais, T.M.G.; Filho, C.M.B.; Struve, C.; et al. Cinnamaldehyde inhibits Staphylococcus aureus virulence factors and protects against infection in a Galleria mellonella model. Front. Microbiol. 2016, 7, 2052. [Google Scholar] [CrossRef] [Green Version]
- Pepeljnjak, S.; Kosalec, I. Galangin expresses bactericidal activity against multiple-resistant bacteria: MRSA, Enterococcus spp. and Pseudomonas aeruginosa. FEMS Microbiol. Lett. 2004, 240, 111–116. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Antibiotic/MIC a (μg/mL) | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cefoxitin-Screen Test | Oxacillin | Gentamicin | Tobramycin | Levofloxacin | Moxyfloxacin | Erythromycin | Clindamycin | Linezolid | Teicoplanin | Vancomycin | Tetracycline | Tigecycline | Fosfomycin | Fusidic Acid | Trimethoprim/ Sulfamethox Azole | |
| S. aureus MFBF b 10663 (MSSA c) | - | 2 | ≤0.5 | ≤1 | 1 | 0.5 | ≤0.25 | ≤0.25 | 2 | 2 | 2 | ≤1 | ≤0.12 | 16 | ≤0.5 | ≤10 |
| S. aureus MFBF b 10679 (MRSA d) | + | ≥4 | 1 | 4 | 4 | 2 | ≥8 | 1 | ≥8 | 8 | 8 | 2 | 0.25 | ≥8 | 1 | 160 |
| Sample | S. aureus ATCC 6538 | MRSA f MFBF 10679 | MSSA g MFBF 10663 | P. aeruginosa ATCC 27853 | C. albicans ATCC 90028 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean ± S.D. | |||||||||||
| ZI a (mm) | MIC b (mg/mL) | ZI a (mm) | MIC b (mg/mL) | ZI a (mm) | MIC b (mg/mL) | ZI a (mm) | MIC b (mg/mL) | ZI a (mm) | MIC b (mg/mL) | ||
| N = 5 | N = 3 | N = 5 | N = 3 | N = 5 | N = 3 | N = 5 | N = 3 | N = 5 | N = 3 | ||
| Podvinje | W d | 21 ± 1 1,2 | 8.33 ± 2.9 2,3 | 16 ± 1 2 | 5.00 ± 0.0 1,2,3 | 22 ± 2 3 | 3.33 ± 1.4 1 | 16 ± 1 1,2 | >20 | 6 ± 0 | >20 |
| M e | 19 ± 2 | 3.75 ± 2.2 | 19 ± 2 2,3 | 2.50 ± 0.0 | 22 ± 1 2,3 | 3.33 ± 1.4 | 15 ± 2 | 20.00 ± 0.0 3 | 15 ± 2 1 | >20 | |
| Plitvice | W d | 16 ± 2 2 | 10.00 ± 0.0 2,3 | 14 ± 2 *,2 | 10.00 ± 0.0 *,3 | 20 ± 2 3 | 10.00 ± 0.0 * | 13 ± 2 | >20 | 6 ± 0 | >20 |
| M e | 18 ± 1 | 10.00 ± 0.0 | 19 ± 1 3 | 4.17 ± 1.4 | 22 ± 1 2,3 | 3.33 ± 1.4 | 15 ± 1 3 | 20.00 ± 0.0 3 | 13 ± 1 2,3 | >20 | |
| Trešnjevka | W d | 12 ± 2 *,3 | 20.00 ± 0.0 * | 6 ± 0 *,3 | 10.00 ± 0.0 *,3 | 16 ± 1 3 | 6.67 ± 2.9 | 12 ± 1 *,3 | >20 | 6 ± 0 | >20 |
| M e | 17 ± 1 | 3.75 ± 2.2 | 16 ± 2 | 2.5 ± 0.0 | 20 ± 1 | 3.33 ± 1.4 | 15 ± 2 | 20.00 ± 0.0 3 | 11 ± 1 | >20 | |
| Buzin | W d | 20 ± 2 | 20.00 ± 0.0 * | 16 ± 2 | 20.00 ± 0.0 * | 20 ± 3 * | 6.67 ± 2.9 | 15 ± 1 | >20 | 6 ± 0 | >20 |
| M e | 19 ± 2 | 6.67 ± 2.9 | 13 ± 2 | 6.67 ± 2.9 | 19 ± 1 | 3.33 ± 1.4 | 12 ± 2 | 10.00 ± 0.0 | 6 ± 0 | >20 | |
| Gentamicin sulphate | 17±1 | 0.001 ± 0.0 | 13 ± 2 | 0.001 ± 0.000 | 12 ± 1 | 0.001 ± 0.000 | 12 ± 1 | 0.003 ± 0.000 | NT c | NT c | |
| Nystatin | NT c | 25 ± 1 | 0.03 ± 0.00 | ||||||||
| Sample or Control | MSSA ATCC 6538 | MRSA MFBF 10679 | ||
|---|---|---|---|---|
| MBFIC50 a | MBFIC90 a | MBFIC50 a | MBFIC90 a | |
| µg/mL | ||||
| Podvinje-W | 1.05 ± 0.97 | 5095.37 ± 1143.88 | 61.84 ± 56.00 | 8509.34 ± 1268.83 |
| Podvinje-M | 0.41 ± 0.27 | 3083.98 ± 549.69 | 3.17 ± 5.30 | 8091.00 ± 477.24 |
| Gentamicin sulphate | 0.01 ± 0.01 | 0.99 ± 0.03 | 0.05 ± 0.03 | 1.66 ± 0.49 |
| Sample or Control | Hemoglobin Equivalents (mg/mL) | Relative Hemolysis (%) | ||
|---|---|---|---|---|
| MSSA ATCC 6538 | MRSA MFBF 10679 | MSSA ATCC 6538 | MRSA MFBF 10679 | |
| Podvinje-M c1 | 5.36 | 5.33 | 70.50 ± 14.71 | 70.14 ± 14.22 |
| Podvinje-M c2 | 6.56 | 7.41 | 84.15 ± 4.74 | 87.61 ± 7.57 |
| Podvinje-W c1 | 3.47 | 3.90 | 49.30 ± 5.51 | 51.28 ± 4.00 |
| Podvinje-W c2 | 4.55 | 6.60 | 62.90 ± 5.23 | 80.43 ± 6.66 |
| NC (M) | 7.53 | 7.97 | 100.00 ± 0.00 | 100.00 ± 0.00 |
| PC (V) | 7.73 | 8.42 | 100.00 ± 0.00 | 100.00 ± 0.00 |
| PC/Triton X-100 (2%) | 8.39 | 103.50 | ||
| No | Compound Name | tR, min | Molecular Formula, [M–H]- | Calculated Mass, [M–H]- | Exact Mass, [M–H]- | Δ mDa | MS2 Fragments, (% Base Peak) | MS3 Fragments, (% Base Peak) | MS4 Fragments, (% Base Peak) | Podvinje-M | Podvinje-W |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Gallic acid derivatives | mg/kg GAE * | ||||||||||
| 1 | Galloyl hexoside isomer 1 | 2.84 | C13H15O10- | 331.06707 | 331.06312 | 3.95 | 125(12), 169(100), 170(8), 193(12), 211(28), 271(65), 272(9) | 125(100) | 81(33), 97(15), 107(100), 133(5) | 35.43 | 15.41 |
| 2 | Gallic acid | 3.88 | C7H5O5- | 169.01425 | 169.01234 | 1.91 | 124(4), 125(100) | 53(6), 79(17), 81(100), 83(3), 97(69), 107(16) | NA | 140.51 | 54.64 |
| 3 | Galloyl pentoside isomer 1 | 5.54 | C12H13O9- | 301.05651 | 301.05327 | 3.24 | 125(5), 149(55), 169(100), 170(7), 255(6), 256(5), 257(4) | 125(100) | 81(55), 97(20), 107(100) | 3.87 | 9.69 |
| 4 | Galloyl hexoside isomer 2 | 5.57 | C13H15O10- | 331.06707 | 331.06263 | 4.44 | 125(5), 169(100), 170(6), 223(3), 234(4), 285(3) | 125(100) | 81(61), 83(6), 97(100), 107(27), 239(16) | 9.02 | ND |
| 5 | Galloyl-shikimic acid | 5.85 | C14H13O9- | 325.05651 | 325.05217 | 4.34 | 125(9), 169(100), 170(4) | 125(100) | 53(5), 79(3), 81(47), 97(53), 107(100) | 375.42 | 531.45 |
| 6 | Galloyl pentoside isomer 2 | 5.95 | C12H13O9- | 301.05651 | 301.05302 | 3.49 | 125(4), 149(58), 169(100), 170(4) | 125(100) | 81(32), 84(3), 97(34), 107(100), 109(3) | 60.54 | 137.42 |
| 7 | Protocatechuic acid | 6.19 | C7H5O4- | 153.01933 | 153.01803 | 1.30 | 108(3), 109(100), 110(5) | 65(100), 81(62) | NA | 22.60 | 3.82 |
| 8 | Methylgalloyl hexoside isomer 1 | 6.44 | C14H17O10- | 345.08272 | 345.07854 | 4.18 | 183(100), 184(4) | 124(85), 168(100), 183(3) | 124(100) | 101.03 | 76.57 |
| 9 | Digalloyl hexoside | 6.50 | C20H19O14- | 483.07803 | 483.07317 | 4.86 | 169(12), 193(10), 211(14), 271(100), 272(11), 313(25), 331(22) | 169(12), 211(100) | 124(24), 165(10), 167(28), 168(100), 183(9) | 11.46 | 7.53 |
| 10 | Methylgalloyl hexoside isomer 2 | 6.81 | C14H17O10- | 345.08272 | 345.08100 | 1.72 | 183(100), 184(8), 299(12), 300(3), 323(3) | 124(100), 168(93) | 78(100), 96(25), 106(59) | 15.05 | ND |
| 11 | Corilagin | 7.13 | C27H21O18- | 633.07334 | 633.06856 | 4.78 | 275(17), 301(100), 302(13), 419(5), 463(20), 613(10), 614(7) | 185(34), 201(13), 229(61), 257(100), 284(24), 301(15) | 185(100), 201(15), 213(6), 229(83), 230(6) | 94.00 | 61.15 |
| 12 | Methylgallate | 7.29 | C8H7O5- | 183.02990 | 183.02832 | 1.58 | 124(100), 137(12), 153(12), 167(14), 168(100), 169(7), 183(9) | 124(100) | 78(100), 79(4), 106(45), 140(27) | 24.67 | 4.10 |
| 13 | Digalloyl-shikimic acid | 7.31 | C21H17O13- | 477.06746 | 477.06271 | 4.75 | 169(25), 263(76), 289(100), 290(13), 307(31), 325(47), 453(12) | 93(4), 137(100), 151(5), 245(9) | 93(100) | 14.97 | 12.28 |
| 14 | Methylgalloyl-galloyl hexoside | 7.38 | C21H21O14- | 497.09368 | 497.08902 | 4.66 | 183(6), 313(3), 345(100), 346(11), 465(11), 466(3) | 183(100) | 124(81), 168(100), 183(3) | 16.34 | 19.42 |
| 15 | Methylgalloyl-caffeoyl hexoside | 8.59 | C23H23O13- | 507.11441 | 507.10987 | 4.54 | 179(5), 183(6), 323(18), 345(100), 346(13), 916(13), 917(6) | 183(100) | 124(90), 168(100) | 5.63 | 2.33 |
| 16 | Methylgalloyl-coumaroyl hexoside | 9.21 | C23H23O12- | 491.11950 | 491.11539 | 4.11 | 183(14), 329(51), 330(10), 345(100), 346(13), 409(7), 457(9) | 183(100) | 124(71), 168(100), 183(3) | 5.09 | 2.37 |
| 17 | Trimethylellagic acid isomer 1 | 9.62 | C17H11O4- | 343.04594 | 343.04253 | 3.41 | 171(3), 297(4), 299(4), 315(3), 325(5), 328(100), 329(15) | 313(100), 314(9) | 285(41), 286(3), 298(100), 299(4) | 78.57 | 61.98 |
| 18 | Ellagic acid | 9.65 | C14H5O8- | 300.99899 | 300.99735 | 1.64 | 185(41), 229(83), 255(48), 257(100), 271(61), 284(38), 301(37) | 185(100), 186(12), 201(10), 213(18), 228(5), 229(70) | 141(100), 157(46) | 11.77 | ND |
| 19 | Trimethylellagic acid isomer 2 | 10.74 | C17H11O4- | 343.04594 | 343.04226 | 3.68 | 295(4), 297(3), 325(3), 328(100), 329(17), 330(3) | 313(100), 314(10) | 285(40), 298(100), 299(8) | 34.25 | 16.44 |
| Flavonol glycosides | mg/kg RE * | ||||||||||
| 20 | Quercetin 3-O-(2″-hexosyl)hexoside | 7.50 | C27H29O17- | 625.14102 | 625.13645 | 4.57 | 271(18), 300(15), 300(100), 301(50), 445(26), 463(10), 505(11) | 151(21), 179(25), 254(10), 255(53), 271(100), 272(21) | 199(32), 215(28), 227(79), 243(100), 271(14) | 2.57 | 112.39 |
| 21 | Kaempferol 3-O-(2″-hexosyl)hexoside | 7.81 | C27H29O16- | 609.14611 | 609.14166 | 4.45 | 255(11), 257(9), 284(47), 285(100), 286(11), 429(46), 430(8) | 151(47), 213(31), 229(42), 241(50), 256(47), 257(100) | 163(48), 187(13), 213(18), 229(100), 239(25) | 2.77 | 11.46 |
| 22 | Quercetin 3-O-(6″-rhamnosyl)glucoside (Rutin) | 7.99 | C27H29O16- | 609.14611 | 609.14278 | 3.33 | 255(5), 271(7), 299(5), 300(42), 301(100), 302(13), 343(8) | 151(78), 179(100), 256(10), 257(13), 272(13), 273(17) | 151(100) | 13.95 | 33.20 |
| 23 | Quercetin 3-O-galactoside (Hyperoside) | 8.31 | C21H19O12- | 463.08820 | 463.08349 | 4.71 | 300(12), 301(100), 302(11), 381(3), 445(4) | 151(77), 179(100), 255(46), 257(12), 271(72), 272(23) | 151(100) | 18.32 | 13.40 |
| 24 | Quercetin 3-O-hexuronide | 8.31 | C21H17O13- | 477.06692 | 477.06196 | 4.96 | 301(100), 302(13), 315(7), 429(6), 431(3), 453(9) | 107(5), 151(80), 179(100), 193(5), 257(13), 273(20) | 151(100) | 5.05 | 6.75 |
| 25 | Kaempferol 7-O-(6″-rhamosyl)glucoside | 8.41 | C27H29O15- | 593.15119 | 593.14656 | 4.63 | 257(3), 284(6), 285(100), 286(12) | 197(20), 213(25), 229(49), 241(33), 257(100), 267(47) | 163(75), 187(17), 213(32), 229(100), 239(29) | 35.26 | 28.72 |
| 26 | Isorhamnetin 3-O-(6″-rhamnosyl)glucoside (Narcissin) | 8.48 | C28H31O16- | 623.16176 | 623.15692 | 4.84 | 255(3), 271(5), 300(14), 315(100), 316(13), 317(3) | 272(6), 287(5), 300(100) | 255(65), 271(100), 272(39) | 97.90 | 107.05 |
| 27 | Isorhamnetin 3-O-glucoside | 8.82 | C22H21O12- | 477.10385 | 477.09994 | 3.91 | 271(8), 285(9), 314(100), 315(45), 316(6), 357(17), 453(7) | 243(24), 271(74), 285(100), 286(50), 299(25), 300(44) | 270(100) | 11.10 | 6.40 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ljoljić Bilić, V.; Gašić, U.M.; Milojković-Opsenica, D.; Rimac, H.; Vuković Rodriguez, J.; Vlainić, J.; Brlek-Gorski, D.; Kosalec, I. Antibacterial Fractions from Erodium cicutarium Exposed—Clinical Strains of Staphylococcus aureus in Focus. Antibiotics 2022, 11, 492. https://doi.org/10.3390/antibiotics11040492
Ljoljić Bilić V, Gašić UM, Milojković-Opsenica D, Rimac H, Vuković Rodriguez J, Vlainić J, Brlek-Gorski D, Kosalec I. Antibacterial Fractions from Erodium cicutarium Exposed—Clinical Strains of Staphylococcus aureus in Focus. Antibiotics. 2022; 11(4):492. https://doi.org/10.3390/antibiotics11040492
Chicago/Turabian StyleLjoljić Bilić, Vanja, Uroš M. Gašić, Dušanka Milojković-Opsenica, Hrvoje Rimac, Jadranka Vuković Rodriguez, Josipa Vlainić, Diana Brlek-Gorski, and Ivan Kosalec. 2022. "Antibacterial Fractions from Erodium cicutarium Exposed—Clinical Strains of Staphylococcus aureus in Focus" Antibiotics 11, no. 4: 492. https://doi.org/10.3390/antibiotics11040492
APA StyleLjoljić Bilić, V., Gašić, U. M., Milojković-Opsenica, D., Rimac, H., Vuković Rodriguez, J., Vlainić, J., Brlek-Gorski, D., & Kosalec, I. (2022). Antibacterial Fractions from Erodium cicutarium Exposed—Clinical Strains of Staphylococcus aureus in Focus. Antibiotics, 11(4), 492. https://doi.org/10.3390/antibiotics11040492