
NDM-1 Introduction in Portugal through a ST11 KL105 Klebsiella pneumoniae Widespread in Europe

 ,
, 
Abstract
:1. Introduction
2. Results
2.1. Epidemiological Context and Infection Control Measures
2.2. Strain Relatedness
2.3. Molecular Characterization
2.4. Whole-Genome Data and Phylogenetic Analysis
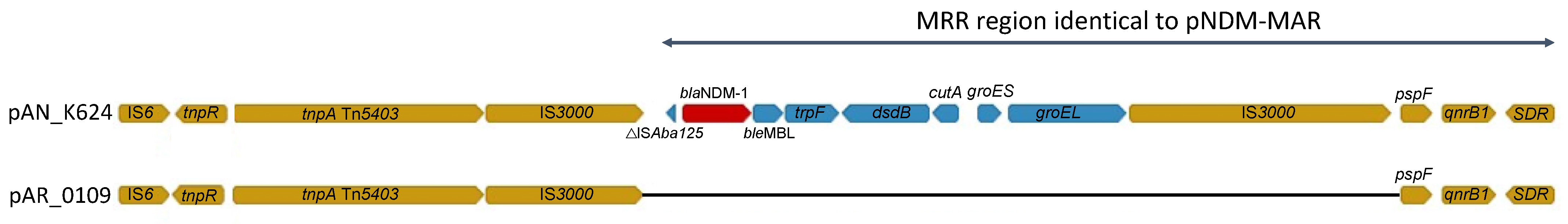
2.5. Genetic Support for blaNDM-1 Acquisition
3. Discussion
4. Materials and Methods
4.1. Setting
4.2. Index Case
4.3. Additional Cases
4.4. Bacterial Identification and Carbapenemase Detection
4.5. Antibiotic Resistance Phenotypes and Genotypes
4.6. Identification of Outbreak Strains
4.7. Whole Genome and Phylogenetic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wyres, K.L.; Lam, M.M.C.; Holt, K.E. Population genomics of Klebsiella pneumoniae. Nat. Rev. Microbiol. 2020, 18, 344–359. [Google Scholar] [CrossRef] [PubMed]
- David, S.; Reuter, S.; Harris, S.R.; Glasner, C.; Feltwell, T.; Argimon, S.; Abudahab, K.; Goater, R.; Giani, T.; Errico, G.; et al. Epidemic of carbapenem-resistant Klebsiella pneumoniae in Europe is driven by nosocomial spread. Nat. Microbiol. 2019, 4, 1919–1929. [Google Scholar] [CrossRef]
- Lee, C.R.; Lee, J.H.; Park, K.S.; Kim, Y.B.; Jeong, B.C.; Lee, S.H. Global Dissemination of Carbapenemase-Producing Klebsiella pneumoniae: Epidemiology, Genetic Context, Treatment Options, and Detection Methods. Front. Microbiol. 2016, 13, 895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerra, A.M.; Lira, A.; Lameirão, A.; Selaru, A.; Abreu, G.; Lopes, P.; Mota, M.; Novais, Â.; Peixe, L. Multiplicity of carbapenemase-producers three years after a KPC-3-producing Klebsiella pneumoniae ST147-K64 hospital outbreak. Antibiotics 2020, 13, 806. [Google Scholar] [CrossRef]
- Bonnin, R.A.; Jousset, A.B.; Chiarelli, A.; Emeraud, C.; Glaser, P.; Naas, T.; Dortet, L. Emergence of New Non-Clonal Group 258 High-Risk Clones among Klebsiella pneumoniae Carbapenemase-Producing K. pneumoniae Isolates, France. Emerg. Infect. Dis. 2020, 26, 1212–1220. [Google Scholar] [CrossRef] [PubMed]
- Hernández-García, M.; Pérez-Viso, B.; Carmen Turrientes, M.; Díaz-Agero, C.; López-Fresneña, N.; Bonten, M.; Malhotra-Kumar, S.; Ruiz-Garbajosa, P.; Cantón, R. Characterization of carbapenemase-producing Enterobacteriaceae from colonized patients in a university hospital in Madrid, Spain, during the R-GNOSIS project depicts increased clonal diversity over time with maintenance of high-risk clones. J. Antimicrob. Chemother. 2018, 73, 3039–3043. [Google Scholar] [CrossRef]
- Bonura, C.; Giuffrè, M.; Aleo, A.; Fasciana, T.; Di Bernardo, F.; Stampone, T.; Giammanco, A.; The MDR-GN Working Group; Palma, D.M.; Mammina, C. An Update of the Evolving Epidemic of blaKPC Carrying Klebsiella pneumoniae in Sicily, Italy, 2014: Emergence of Multiple Non-ST258 Clones. PLoS ONE 2015, 10, e0132936. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Mathema, B.; Pitout, J.D.; DeLeo, F.R.; Kreiswirth, B.N. Epidemic Klebsiella pneumoniae ST258 is a hybrid strain. mBio 2014, 5, e01355-14. [Google Scholar] [CrossRef] [Green Version]
- Dong, N.; Zhang, R.; Liu, L.; Li, R.; Lin, D.; Chan, E.W.; Chen, S. Genome analysis of clinical multilocus sequence Type 11 Klebsiella pneumoniae from China. Microb. Genom. 2018, 4, e000149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wyres, K.L.; Hawkey, J.; Hetland, M.A.K.; Fostervold, A.; Wick, R.R.; Judd, L.M.; Hamidian, M.; Howden, B.P.; Löhr, I.H.; Holt, K.E. Emergence and rapid global dissemination of CTX-M-15-associated Klebsiella pneumoniae strain ST307. J. Antimicrob. Chemother. 2019, 74, 577–581. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Liu, L.; Zhou, H.; Chan, E.W.; Li, J.; Fang, Y.; Li, Y.; Liao, K.; Chen, S. Nationwide surveillance of clinical carbapenem-resistant Enterobacteriaceae (CRE) strains in China. EBioMedicine 2017, 19, 98–106. [Google Scholar] [CrossRef] [Green Version]
- Zhou, K.; Xiao, T.; David, S.; Wang, Q.; Zhou, Y.; Guo, L.; Aanensen, D.; Holt, K.E.; Thomson, N.T.; Grundmann, H.; et al. Novel Subclone of Carbapenem-Resistant Klebsiella pneumoniae Sequence Type 11 with Enhanced Virulence and Transmissibility, China. Emerg. Infect. Dis. 2020, 26, 289–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, C.; Sousa, C.; Lopes, J.A.; Novais, Â.; Peixe, L. A Front Line on Klebsiella pneumoniae Capsular Polysaccharide Knowledge: Fourier Transform Infrared Spectroscopy as an Accurate and Fast Typing Tool. mSystems 2020, 5, e00386-19. [Google Scholar] [CrossRef] [Green Version]
- Silva, L.; Rodrigues, C.; Lira, A.; Leão, M.; Mota, M.; Lopes, P.; Novais, Â.; Peixe, L. Fourier transform infrared (FT-IR) spectroscopy typing: A real-time analysis of an outbreak by carbapenem-resistant Klebsiella pneumoniae. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 2471–2475. [Google Scholar] [CrossRef]
- Direção-Geral da Saúde. Infeções e Resistências aos Antimicrobianos: Relatório Anual do Programa Prioritário 2018. Available online: https://www.dgs.pt/portal-da-estatistica-da-saude/diretorio-de-informacao/diretorio-de-informacao/por-serie-1003038-pdf.aspx?v=%3d%3dDwAAAB%2bLCAAAAAAABAArySzItzVUy81MsTU1MDAFAHzFEfkPAAAA (accessed on 5 November 2019).
- Rodrigues, C.; Machado, E.; Ramos, H.; Peixe, L.; Novais, Â. Expansion of ESBL-producing Klebsiella pneumoniae in hospitalized patients: A successful story of international clones (ST15, ST147, ST336) and epidemic plasmids (IncR, IncFIIK). Int. J. Med. Microbiol. 2014, 304, 1100–11088. [Google Scholar] [CrossRef]
- Rodrigues, C.; Bavlovič, J.; Machado, E.; Amorim, J.; Peixe, L.; Novais, Â. KPC-3-Producing Klebsiella pneumoniae in Portugal Linked to Previously Circulating Non-CG258 Lineages and Uncommon Genetic Platforms (Tn4401d-IncFIA and Tn4401d-IncN). Front. Microbiol. 2016, 7, 1000. [Google Scholar] [CrossRef] [Green Version]
- Aires-de-Sousa, M.; Ortiz de la Rosa, J.M.; Gonçalves, M.L.; Pereira, A.L.; Nordmann, P.; Poirel, L. Epidemiology of Carbapenemase-Producing Klebsiella pneumoniae in a Hospital, Portugal. Emerg. Infect. Dis. 2019, 25, 1632–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perdigão, J.; Modesto, A.; Pereira, A.L.; Neto, O.; Matos, V.; Godinho, A.; Phelan, J.; Charleston, J.; Sadar, A.; de Sessions, P.F.; et al. Whole-genome sequencing resolves a polyclonal outbreak by extended-spectrum beta-lactam and carbapenem-resistant Klebsiella pneumoniae in a Portuguese tertiary-care hospital. Microb. Genom. 2019, 7, 000349. [Google Scholar] [CrossRef]
- Lopes, E.; Saavedra, M.J.; Costa, E.; de Lencastre, H.; Poirel, L.; Aires-de-Sousa, M. Epidemiology of carbapenemase-producing Klebsiella pneumoniae in northern Portugal: Predominance of KPC-2 and OXA-48. J. Glob. Antimicrob. Resist. 2020, 22, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Aires-de-Sousa, M.; Ortiz de la Rosa, J.M.; Goncalves, M.L.; Costa, A.; Nordmann, P.; Poirel, L. Occurrence of NDM-1-producing Morganella morganii and Proteus mirabilis in a single patient in Portugal: Probable in vivo transfer by conjugation. J. Antimicrob. Chemother. 2020, 75, 903–906. [Google Scholar] [CrossRef]
- Manageiro, V.; Sampaio, D.A.; Pereira, P.; Rodrigues, P.; Veira, L.; Palos, C.; Caniça, M. Draft Genome Sequence of the First NDM-1-Producing Providencia stuartii Strain Isolated in Portugal. Genome Announc. 2015, 3, e01077-15. [Google Scholar] [CrossRef] [Green Version]
- Wailan, A.M.; Paterson, D.L. The spread and acquisition of NDM-1: A multifactorial problem. Expert. Rev. Anti. Infect. Ther. 2014, 12, 91–115. [Google Scholar] [CrossRef] [PubMed]
- Politi, L.; Gartzonika, K.; Spanakis, N.; Zarkotou, O.; Poulou, A.; Skoura, L.; Vrioni, G.; Tsakris, A. Emergence of NDM-1-producing Klebsiella pneumoniae in Greece: Evidence of a widespread clonal outbreak. J. Antimicrob. Chemother. 2019, 74, 2197–2202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, T.G.; Novais, Â.; Rodrigues, C.; Nascimento, R.; Freitas, F.; Machado, E.; Peixe, L. Dynamics of clonal and plasmid backgrounds of Enterobacteriaceae producing acquired AmpC in Portuguese clinical settings over time. Int. J. Antimicrob. Agents 2019, 53, 650–656. [Google Scholar] [CrossRef]
- Izdebski, R.; Sitkiewicz, M.; Urbanowicz, P.; Krawczyk, M.; Gniadkowski, M. Genomic background of the Klebsiella pneumoniae NDM-1 outbreak in Poland, 2012-18. J. Antimicrob. Chemother. 2020, 75, 3156–3162. [Google Scholar] [CrossRef] [PubMed]
- van Dorp, L.; Wang, Q.; Shaw, L.P.; Acman, M.; Brynildsrud, O.B.; Eldholm, V.; Wang, R.; Gao, H.; Yin, Y.; Chen, H.; et al. Rapid phenotypic evolution in multidrug-resistant Klebsiella pneumoniae hospital outbreak strains. Microb. Genom. 2019, 5, e000263. [Google Scholar] [CrossRef]
- Bowers, J.R.; Kitchel, B.; Driebe, E.M.; MacCannell, D.R.; Roe, C.; Lemmer, D.; de Man, T.; Rasheed, J.K.; Engelthaler, D.M.; Keim, P.; et al. Genomic Analysis of the Emergence and Rapid Global Dissemination of the Clonal Group 258 Klebsiella pneumoniae Pandemic. PLoS ONE 2015, 10, e0133727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novais, Â.; Freitas, A.R.; Rodrigues, C.; Peixe, L. Fourier transform infrared spectroscopy: Unlocking fundamentals and prospects for bacterial strain typing. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 427–448. [Google Scholar] [CrossRef]
- Naas, T.; Oueslati, S.; Bonnin, R.A.; Dabos, M.L.; Zavala, A.; Dortet, L.; Retailleau, P.; Iorga, B.I. Beta-lactamase database (BLDB)—structure and function. J. Enzyme Inhib. Med. Chem. 2017, 32, 917–919. [Google Scholar] [CrossRef]
- Pires, J.; Novais, A.; Peixe, L. Blue-carba, an easy biochemical test for detection of diverse carbapenemase producers directly from bacterial cultures. J. Clin. Microbiol. 2013, 51, 4281–4283. [Google Scholar] [CrossRef] [Green Version]
- Bogaerts, P.; Rezende de Castro, R.; de Mendonça, R.; Huang, T.D.; Denis, O.; Glupczynski, Y. Validation of carbapebemase and extended-spectrum β-lactamase multiplex endpoint PCR assays according to ISO 15189. J. Antimicrob. Chemother. 2013, 68, 1576–1582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brisse, S.; Passet, V.; Haugaard, A.B.; Babosan, A.; Kassis-Chikhani, N.; Struve, C.; Decré, D. wzi gene sequencing, a rapid method to determine the capsular type of Klebsiella strains. J. Clin. Microbiol. 2013, 51, 4073–4078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arredondo-Alonso, S.; Rogers, M.R.C.; Braat, J.C.; Verschuuren, T.D.; Top, J.; Corander, J.; Willems, R.J.L.; Schürch, A.C. mlplasmids: A user-friendly tool to predict plasmid- and chromosome-derived sequences for single species. Microb. Genom. 2018, 4, e000224. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Patient N° | Sex/Age | Isolates | Date of Isolation | Hospital Unit | Period of Hospitalization | Patient’s Origin | Sample Type | Pathology | Colonization at Admission | Antibiotherapy (Previous 3 Months) | Previous Hospitalization (Date, Unit) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | M/71 | K607 K606 * | 27/11/18 28/11/18 | Surgery | 26–30 October 2018 | Hospital | C (bile) C (RS) | Cholangiocarcinoma | Not tested | Piperacillin-Tazobactam | None |
| 2 | M/79 | K608 | 28/11/18 | Surgery | 15 November–3 December 2018 | Hospital | C (RS) | Acute pancreatitis | Not tested | None | None |
| 3 | M/55 | K609 | 30/11/18 | Surgery | 24 October–5 December 2018 | Hospital | C (RS) | Trauma | - | Amoxacillin-clavulanic acid | 15 September–22 October 2018 (different hospital) |
| 4 | M/66 | K610 | 30/11/18 | Surgery | 23–30 November 2018 | Hospital | C (RS) | Adenocarcinoma | Not tested | None | None |
| 5 | M/66 | K611 | 14/12/19 | Emergency | 14 December 2018–24 January 2019 | Outpatient | C (RS) | Colon carcinoma | + | Ciprofloxacin | 10–28 November 2018 (Surgery) |
| 6 | M/68 | K612 | 25/12/19 | Emergency | 25 December 2018–11 January 2019 (Medicine) | Outpatient | C (RS) | Soft tissue infection | + | Tigecycline, Ceftazidime | 31 October–9 December 2018 (Surgery) |
| 7 | M/87 | K624 * K625 | 25/03/19 | Emergency | 25 March–18 April 2019 | Household | I (urine) C (RS) | Pneumonia | - | Cefuroxime | 28 January–4 March 2019 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Novais, Â.; Ferraz, R.V.; Viana, M.; da Costa, P.M.; Peixe, L. NDM-1 Introduction in Portugal through a ST11 KL105 Klebsiella pneumoniae Widespread in Europe. Antibiotics 2022, 11, 92. https://doi.org/10.3390/antibiotics11010092
Novais Â, Ferraz RV, Viana M, da Costa PM, Peixe L. NDM-1 Introduction in Portugal through a ST11 KL105 Klebsiella pneumoniae Widespread in Europe. Antibiotics. 2022; 11(1):92. https://doi.org/10.3390/antibiotics11010092
Chicago/Turabian StyleNovais, Ângela, Rita Veiga Ferraz, Mariana Viana, Paula Martins da Costa, and Luísa Peixe. 2022. "NDM-1 Introduction in Portugal through a ST11 KL105 Klebsiella pneumoniae Widespread in Europe" Antibiotics 11, no. 1: 92. https://doi.org/10.3390/antibiotics11010092
APA StyleNovais, Â., Ferraz, R. V., Viana, M., da Costa, P. M., & Peixe, L. (2022). NDM-1 Introduction in Portugal through a ST11 KL105 Klebsiella pneumoniae Widespread in Europe. Antibiotics, 11(1), 92. https://doi.org/10.3390/antibiotics11010092