
The Antimicrobial Resistance Characteristics of Imipenem-Non-Susceptible, Imipenemase-6-Producing Escherichia coli

 ,
,  , , ,
, , ,
Abstract
1. Introduction
2. Results
2.1. Antimicrobial Susceptibilities
2.2. Phenotypic and Genotypic Detection of Carbapenemase and ESBL
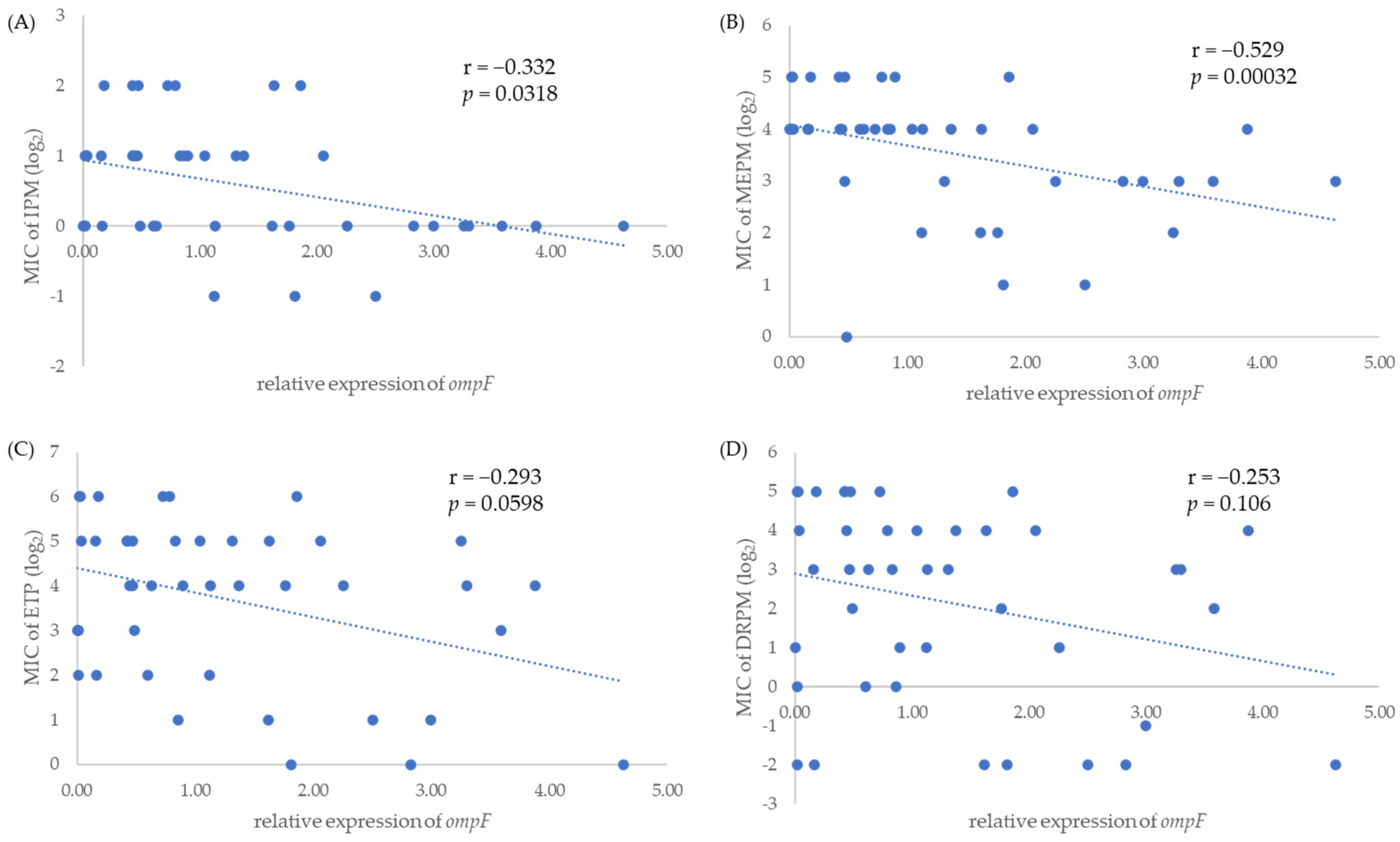
2.3. Relative Expression Level of ompC and ompF
2.4. Efflux Pump Activity and Carbapenem Resistance
2.5. Plasmid Replicon Typing
3. Discussion
4. Materials and Methods
4.1. Bacterial Collection
4.2. Antimicrobial Susceptibility Tests
4.3. Phenotypic Detections of ESBL Production in E. coli Isolates
4.4. Detection of Carbapenemase Genes and ESBL Genes
4.5. q-RT-PCR for Porin Coding Genes ompC and ompF
4.6. Efflux Pump Inhibitory Assay
4.7. Plasmid Replicon Typing
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yamamoto, N.; Asada, R.; Kawahara, R.; Hagiya, H.; Akeda, Y.; Shanmugakani, R.K.; Yoshida, H.; Yukawa, S.; Yamamoto, K.; Takayama, Y.; et al. Prevalence of, and risk factors for, carriage of carbapenem-resistant Enterobacteriaceae among hospitalized patients in Japan. J. Hosp. Infect. 2017, 97, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Yano, H.; Kuga, A.; Okamoto, R.; Kitasato, H.; Kobayashi, T.; Inoue, M. Plasmid-Encoded Metallo-β-Lactamase (IMP-6) Conferring Resistance to Carbapenems, Especially Meropenem. Antimicrob. Agents Chemother. 2001, 45, 1343–1348. [Google Scholar] [CrossRef] [PubMed]
- Kayama, S.; Shigemoto, N.; Kuwahara, R.; Oshima, K.; Hirakawa, H.; Hisatsune, J.; Jové, T.; Nishio, H.; Yamasaki, K.; Wada, Y.; et al. Complete Nucleotide Sequence of the IncN Plasmid Encoding IMP-6 and CTX-M-2 from Emerging Carbapenem-Resistant Enterobacteriaceae in Japan. Antimicrob. Agents Chemother. 2015, 59, 1356–1359. [Google Scholar] [CrossRef] [PubMed]
- Ambler, R.P. The structure of beta-lactamases. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1980, 289, 321–331. [Google Scholar] [CrossRef] [PubMed]
- Doumith, M.; Ellington, M.J.; Livermore, D.M.; Woodford, N. Molecular mechanisms disrupting porin expression in ertapenem-resistant Klebsiella and Enterobacter spp. clinical isolates from the UK. J. Antimicrob. Chemother. 2009, 63, 659–667. [Google Scholar] [CrossRef]
- Schwaber, M.J.; Navon-Venezia, S.; Kaye, K.S.; Ben-Ami, R.; Schwartz, D.; Carmeli, Y. Clinical and economic impact of bacteremia with extended- spectrum-beta-lactamase-producing Enterobacteriaceae. Antimicrob. Agents Chemother. 2006, 50, 1257–1262. [Google Scholar] [CrossRef] [PubMed]
- Yonekawa, S.; Mizuno, T.; Nakano, R.; Nakano, A.; Suzuki, Y.; Asada, T.; Ishii, A.; Kakuta, N.; Tsubaki, K.; Mizuno, S.; et al. Molecular and Epidemiological Characteristics of Carbapenemase-Producing Klebsiella pneumoniae Clinical Isolates in Japan. mSphere 2020, 5, e00490-20. [Google Scholar] [CrossRef] [PubMed]
- Haidar, G.; Clancy, C.J.; Chen, L.; Samanta, P.; Shields, R.K.; Kreiswirth, B.N.; Nguyen, M.H. Identifying Spectra of Activity and Therapeutic Niches for Ceftazidime-Avibactam and Imipenem-Relebactam against Carbapenem-Resistant Enterobacteriaceae. Antimicrob. Agents Chemother. 2017, 61, e00642-17. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.P.; Woodford, N. Global spread of antibiotic resistance: The example of New Delhi metallo-β-lactamase (NDM)-mediated carbapenem resistance. J. Med. Microbiol. 2013, 62, 499–513. [Google Scholar] [CrossRef]
- Hamzaoui, Z.; Ocampo-Sosa, A.; Fernandez Martinez, M.; Landolsi, S.; Ferjani, S.; Maamar, E.; Saidani, M.; Slim, A.; Martinez-Martinez, L.; Boutiba-Ben Boubaker, I. Role of association of OmpK35 and OmpK36 alteration and blaESBL and/or blaAmpC genes in conferring carbapenem resistance among non-carbapenemase-producing Klebsiella pneumoniae. Int. J. Antimicrob. Agents 2018, 52, 898–905. [Google Scholar] [CrossRef]
- Senchyna, F.; Gaur, R.L.; Sandlund, J.; Truong, C.; Tremintin, G.; Kültz, D.; Gomez, C.A.; Tamburini, F.B.; Andermann, T.; Bhatt, A.; et al. Diversity of resistance mechanisms in carbapenem-resistant Enterobacteriaceae at a health care system in Northern California, from 2013 to 2016. Diagn. Microbiol. Infect. Dis. 2019, 93, 250–257. [Google Scholar] [CrossRef]
- Lin, X.; Wang, C.; Guo, C.; Tian, Y.; Li, H.; Peng, X. Differential regulation of OmpC and OmpF by AtpB in Escherichia coli exposed to nalidixic acid and chlortetracycline. J. Proteom. 2012, 75, 5898–5910. [Google Scholar] [CrossRef] [PubMed]
- Weston, N.; Sharma, P.; Ricci, V.; Piddock, L.J.V. Regulation of the AcrAB-TolC efflux pump in Enterobacteriaceae. Res. Microbiol. 2018, 169, 425–431. [Google Scholar] [CrossRef]
- Van Dyk, T.K.; Templeton, L.J.; Cantera, K.A.; Sharpe, P.L.; Sariaslani, F.S. Characterization of the Escherichia coli AaeAB efflux pump: A metabolic relief valve? J. Bacteriol. 2004, 186, 7196–7204. [Google Scholar] [CrossRef] [PubMed]
- Lamers, R.P.; Cavallari, J.F.; Burrows, L.L. The Efflux Inhibitor Phenylalanine-Arginine Beta-Naphthylamide (PAβN) Permeabilizes the Outer Membrane of Gram-Negative Bacteria. PLoS ONE 2013, 8, e60666. [Google Scholar] [CrossRef] [PubMed]
- Tikhonova, E.B.; Yamada, Y.; Zgurskaya, H.I. Sequential mechanism of assembly of multidrug efflux pump AcrAB-TolC. Chem. Biol. 2011, 18, 454–463. [Google Scholar] [CrossRef] [PubMed]
- Lomovskaya, O.; Warren, M.S.; Lee, A.; Galazzo, J.; Fronko, R.; Lee, M.; Blais, J.; Cho, D.; Chamberland, S.; Renau, T.; et al. Identification and characterization of inhibitors of multidrug resistance efflux pumps in Pseudomonas aeruginosa: Novel agents for combination therapy. Antimicrob. Agents Chemother. 2001, 45, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Dupont, H.; Gaillot, O.; Goetgheluck, A.-S.; Plassart, C.; Emond, J.-P.; Lecuru, M.; Gaillard, N.; Derdouri, S.; Lemaire, B.; Girard de Courtilles, M.; et al. Molecular Characterization of Carbapenem-Nonsusceptible Enterobacterial Isolates Collected during a Prospective Interregional Survey in France and Susceptibility to the Novel Ceftazidime-Avibactam and Aztreonam-Avibactam Combinations. Antimicrob. Agents Chemother. 2015, 60, 215–221. [Google Scholar] [CrossRef]
- Vera-Leiva, A.; Carrasco-Anabalón, S.; Lima, C.A.; Villagra, N.; Domínguez, M.; Bello-Toledo, H.; González-Rocha, G. The efflux pump inhibitor phenylalanine-arginine β-naphthylamide (PAβN) increases resistance to carbapenems in Chilean clinical isolates of KPC-producing Klebsiella pneumoniae. J. Glob. Antimicrob. Resist. 2018, 12, 73–76. [Google Scholar] [CrossRef]
- Carattoli, A.; Bertini, A.; Villa, L.; Falbo, V.; Hopkins, K.L.; Threlfall, E.J. Identification of plasmids by PCR-based replicon typing. J. Microbiol. Methods 2005, 63, 219–228. [Google Scholar] [CrossRef]
- Lee, W.; Chung, H.-S.; Lee, Y.; Yong, D.; Jeong, S.H.; Lee, K.; Chong, Y. Comparison of matrix-assisted laser desorption ionization-time-of-flight mass spectrometry assay with conventional methods for detection of IMP-6, VIM-2, NDM-1, SIM-1, KPC-1, OXA-23, and OXA-51 carbapenemase-producing Acinetobacter spp., Pseudomonas aeruginosa, and Klebsiella pneumoniae. Diagn. Microbiol. Infect. Dis. 2013, 77, 227–230. [Google Scholar] [CrossRef]
- Okanda, T.; Matsumoto, T. In vitro effect of an antimicrobial combination therapy without colistin and tigecycline for CPE and non-CPE. J. Infect. Chemother. 2020, 26, 322–330. [Google Scholar] [CrossRef]
- Ohno, Y.; Nakamura, A.; Hashimoto, E.; Matsutani, H.; Abe, N.; Fukuda, S.; Hisashi, K.; Komatsu, M.; Nakamura, F. Molecular epidemiology of carbapenemase-producing Enterobacteriaceae in a primary care hospital in Japan, 2010–2013. J. Infect. Chemother. 2017, 23, 224–229. [Google Scholar] [CrossRef]
- Liu, S.; Huang, N.; Zhou, C.; Lin, Y.; Zhang, Y.; Wang, L.; Zheng, X.; Zhou, T.; Wang, Z. Molecular Mechanisms and Epidemiology of Carbapenem-Resistant Enterobacter cloacae Complex Isolated from Chinese Patients During 2004-2018. Infect. Drug Resist. 2021, 14, 3647–3658. [Google Scholar] [CrossRef] [PubMed]
- Ellappan, K.; Belgode Narasimha, H.; Kumar, S. Coexistence of multidrug resistance mechanisms and virulence genes in carbapenem-resistant Pseudomonas aeruginosa strains from a tertiary care hospital in South India. J. Glob. Antimicrob. Resist. 2018, 12, 37–43. [Google Scholar] [CrossRef]
- Melvin, P.W.; James, S.L.; April, M.B.; Shelley, C.; Sharon, K.C.; Marcelo, F.G.; Howard, G.; Romney, M.H.; Thomas, J.K.; Brandi, L.; et al. M100Ed31|Performance Standards for Antimicrobial Susceptibility Testing, 31st ed. Available online: https://clsi.org/standards/products/microbiology/documents/m100/ (accessed on 9 October 2021).
- Hattori, T.; Kawamura, K.; Arakawa, Y. Comparison of test methods for detecting metallo-β-lactamase-producing Gram-negative bacteria. Jpn. J. Infect. Dis. 2013, 66, 512–518. [Google Scholar] [CrossRef][Green Version]
- Tada, T.; Miyoshi-Akiyama, T.; Shimada, K.; Shimojima, M.; Kirikae, T. IMP-43 and IMP-44 metallo-β-lactamases with increased carbapenemase activities in multidrug-resistant Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2013, 57, 4427–4432. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, S.; Shigemura, K.; Osawa, K.; Kitagawa, K.; Ishii, A.; Kuntaman, K.; Shirakawa, T.; Miyara, T.; Fujisawa, M. Genetic analysis of ESBL-producing Klebsiella pneumoniae isolated from UTI patients in Indonesia. J. Infect. Chemother. 2021, 27, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Shibata, N.; Doi, Y.; Yamane, K.; Yagi, T.; Kurokawa, H.; Shibayama, K.; Kato, H.; Kai, K.; Arakawa, Y. PCR typing of genetic determinants for metallo-beta-lactamases and integrases carried by gram-negative bacteria isolated in Japan, with focus on the class 3 integron. J. Clin. Microbiol. 2003, 41, 5407–5413. [Google Scholar] [CrossRef]
- Mushi, M.F.; Mshana, S.E.; Imirzalioglu, C.; Bwanga, F. Carbapenemase Genes among Multidrug Resistant Gram Negative Clinical Isolates from a Tertiary Hospital in Mwanza, Tanzania. BioMed Res. Int. 2014, 2014, 303104. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.S.; Kim, K.; Huh, J.Y.; Jung, B.; Kang, M.S.; Hong, S.G. Multiplex PCR for Rapid Detection of Genes Encoding Class A Carbapenemases. Ann. Lab. Med. 2012, 32, 359–361. [Google Scholar] [CrossRef] [PubMed]
- Farajzadeh Sheikh, A.; Rostami, S.; Jolodar, A.; Tabatabaiefar, M.A.; Khorvash, F.; Saki, A.; Shoja, S.; Sheikhi, R. Detection of Metallo-Beta Lactamases Among Carbapenem-Resistant Pseudomonas Aeruginosa. Jundishapur J. Microbiol. 2014, 7, e12289. [Google Scholar] [CrossRef] [PubMed]
- Marchaim, D.; Navon-Venezia, S.; Schwaber, M.J.; Carmeli, Y. Isolation of imipenem-resistant Enterobacter species: Emergence of KPC-2 carbapenemase, molecular characterization, epidemiology, and outcomes. Antimicrob. Agents Chemother. 2008, 52, 1413–1418. [Google Scholar] [CrossRef] [PubMed]
- Shahada, F.; Chuma, T.; Kosugi, G.; Kusumoto, M.; Iwata, T.; Akiba, M. Distribution of extended-spectrum cephalosporin resistance determinants in Salmonella enterica and Escherichia coli isolated from broilers in southern Japan. Poult. Sci. 2013, 92, 1641–1649. [Google Scholar] [CrossRef]
- Peleg, A.Y.; Adams, J.; Paterson, D.L. Tigecycline Efflux as a Mechanism for Nonsusceptibility in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2007, 51, 2065–2069. [Google Scholar] [CrossRef]
- Kanda, Y. Investigation of the freely available easy-to-use software “EZR” for medical statistics. Bone Marrow Transplant. 2013, 48, 452–458. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Susceptible | Intermediate | Resistant | MIC50 (μg/mL) | MIC90 (μg/mL) | |
|---|---|---|---|---|---|
| IPM | 21 (50.0%) | 14 (33.3%) | 7 (16.7%) | 1.5 | 4 |
| MEPM | 1 (2.4%) | 2 (4.8%) | 39 (92.9%) | 16 | 32 |
| ETP | 0 (0%) | 3 (7.1%) | 39 (92.9%) | 16 | 64 |
| DRPM | 11 (26.2%) | 4 (9.5%) | 27 (64.3%) | 8 | 32 |
| PIPC | 1 (2.4%) | 0 (0%) | 41 (97.6%) | >64 | >64 |
| CAZ | 0 (0%) | 1 (2.4%) | 41 (97.6%) | >16 | >16 |
| CFPM | 1 (2.4%) | 1 (2.4%) | 40 (95.2%) | >16 | >16 |
| CPFX | 1 (2.4%) | 0 (0%) | 41 (97.6%) | >2 | >2 |
| LVFX | 1 (2.4%) | 0 (%) | 41 (97.6%) | >4 | >4 |
| AMK | 41 (97.6%) | 1 (2.4%) | 0 (0%) | <4 | 16 |
| GM | 11 (26.2%) | 20 (47.6%) | 11 (26.2%) | 8 | >8 |
| Carbapenemase Producing | Carbapenemase Genes | ESBL Producing | ESBL Genes | |||
|---|---|---|---|---|---|---|
| blaIMP-6 | blaCTX-M-2 | blaCTX-M-14 | blaCTX-M-15 | |||
| Number of isolates (n = 42) | 42 | 42 | 29 | 17 | 1 | 2 |
| (%) | 100.0 | 100.0 | 69.0 | 40.5 | 2.4 | 4.8 |
| Total (n = 42) | IMP-6 + CTX-M-2 (n = 17) | IMP-6 (n = 25) | p-Value | |
|---|---|---|---|---|
| FIA | 37 (88.1%) | 15 (88.2%) | 22 (88.0%) | 1 |
| FIB | 28 (66.7%) | 9 (52.9%) | 19 (76.0%) | 0.184 |
| FIC | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) | N.A. |
| F | 40 (95.2%) | 17 (100.0%) | 23 (92.0%) | 0.506 |
| FII | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) | N.A. |
| HI-1 | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) | N.A. |
| HI-2 | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) | N.A. |
| I1 | 18 (42.9%) | 1 (5.9%) | 17 (68.0%) | <0.001 * |
| L/M | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) | N.A. |
| N | 36 (85.7%) | 16 (94.1%) | 20 (80.0) | 0.374 |
| P | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) | N.A. |
| W | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) | N.A. |
| T | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) | N.A. |
| A/C | 2 (4.8%) | 0 (0.0%) | 2 (8.0) | 0.506 |
| K | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) | N.A. |
| B/O | 4 (9.5%) | 2 (11.8%) | 2 (8.0) | 1 |
| X | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) | N.A. |
| Y | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) | N.A. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Onishi, R.; Shigemura, K.; Osawa, K.; Yang, Y.-M.; Maeda, K.; Fang, S.-B.; Sung, S.-Y.; Onuma, K.; Uda, A.; Miyara, T.; et al. The Antimicrobial Resistance Characteristics of Imipenem-Non-Susceptible, Imipenemase-6-Producing Escherichia coli. Antibiotics 2022, 11, 32. https://doi.org/10.3390/antibiotics11010032
Onishi R, Shigemura K, Osawa K, Yang Y-M, Maeda K, Fang S-B, Sung S-Y, Onuma K, Uda A, Miyara T, et al. The Antimicrobial Resistance Characteristics of Imipenem-Non-Susceptible, Imipenemase-6-Producing Escherichia coli. Antibiotics. 2022; 11(1):32. https://doi.org/10.3390/antibiotics11010032
Chicago/Turabian StyleOnishi, Reo, Katsumi Shigemura, Kayo Osawa, Young-Min Yang, Koki Maeda, Shiuh-Bin Fang, Shian-Ying Sung, Kenichiro Onuma, Atsushi Uda, Takayuki Miyara, and et al. 2022. "The Antimicrobial Resistance Characteristics of Imipenem-Non-Susceptible, Imipenemase-6-Producing Escherichia coli" Antibiotics 11, no. 1: 32. https://doi.org/10.3390/antibiotics11010032
APA StyleOnishi, R., Shigemura, K., Osawa, K., Yang, Y.-M., Maeda, K., Fang, S.-B., Sung, S.-Y., Onuma, K., Uda, A., Miyara, T., & Fujisawa, M. (2022). The Antimicrobial Resistance Characteristics of Imipenem-Non-Susceptible, Imipenemase-6-Producing Escherichia coli. Antibiotics, 11(1), 32. https://doi.org/10.3390/antibiotics11010032