
Strain Improvement and Strain Maintenance Revisited. The Use of Actinoplanes teichomyceticus ATCC 31121 Protoplasts in the Identification of Candidates for Enhanced Teicoplanin Production


 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Actinoplanes teichomyceticus Protoplast Production and Regeneration as the Base for Genome Separation
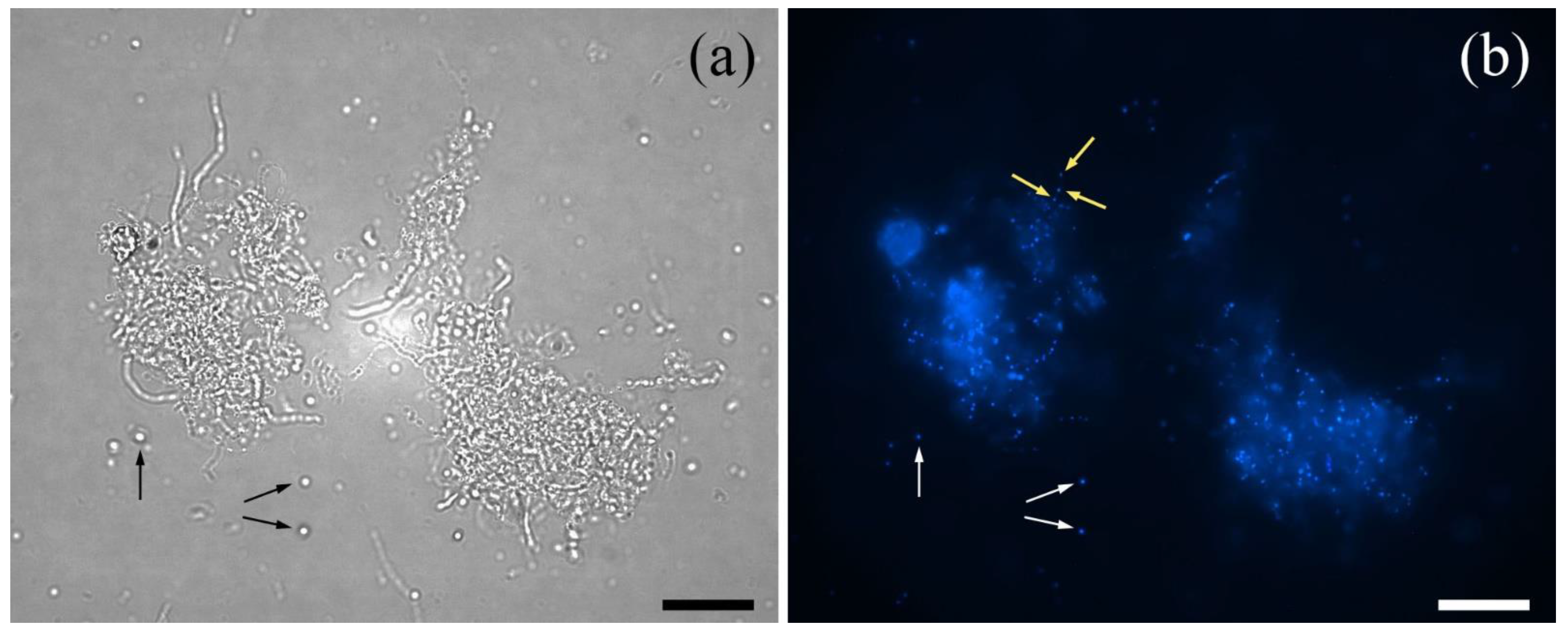
2.2. Macroscopic and Microscopic Analysis of Hyphae-Derived and Protoplast-Derived Clones
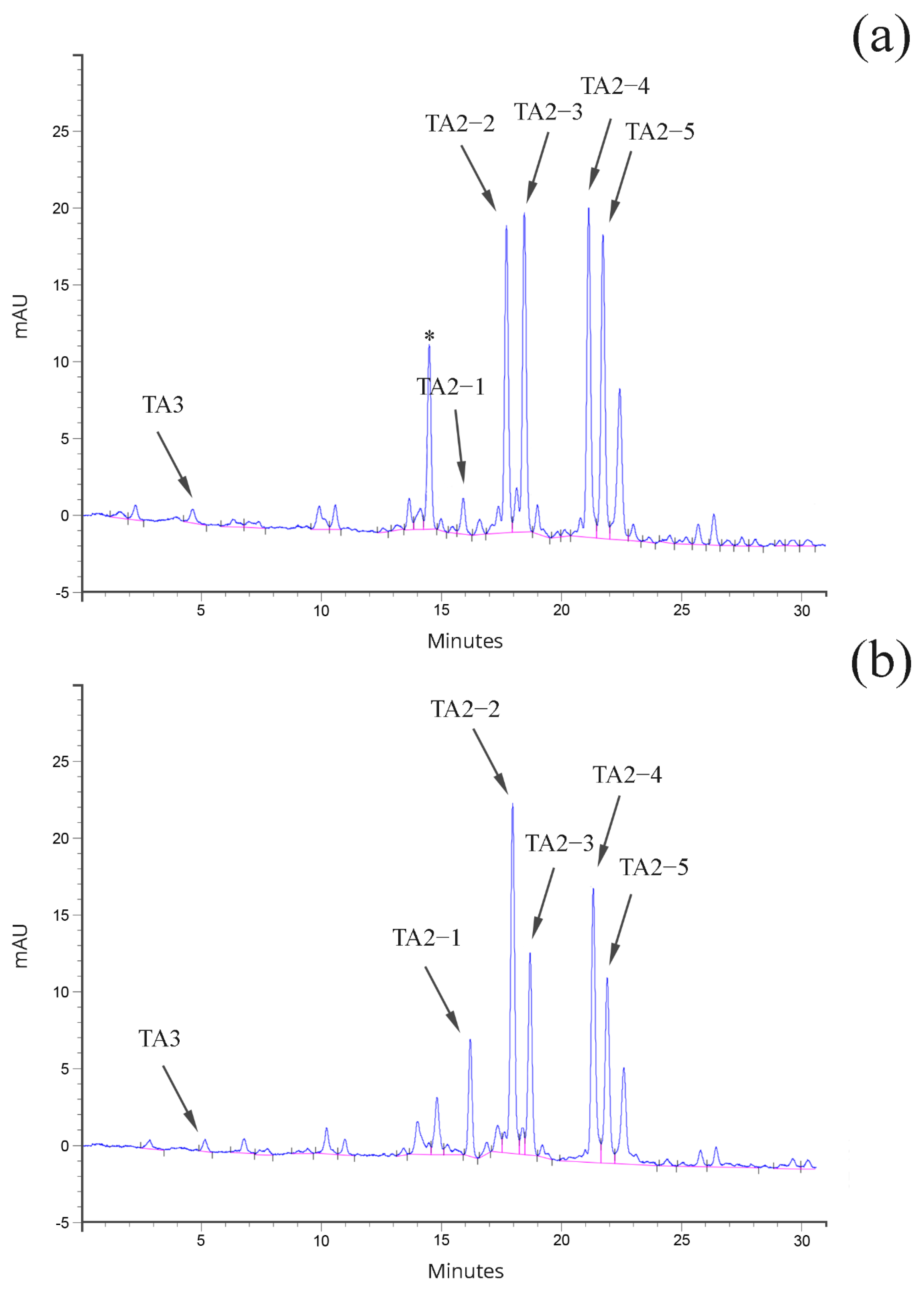
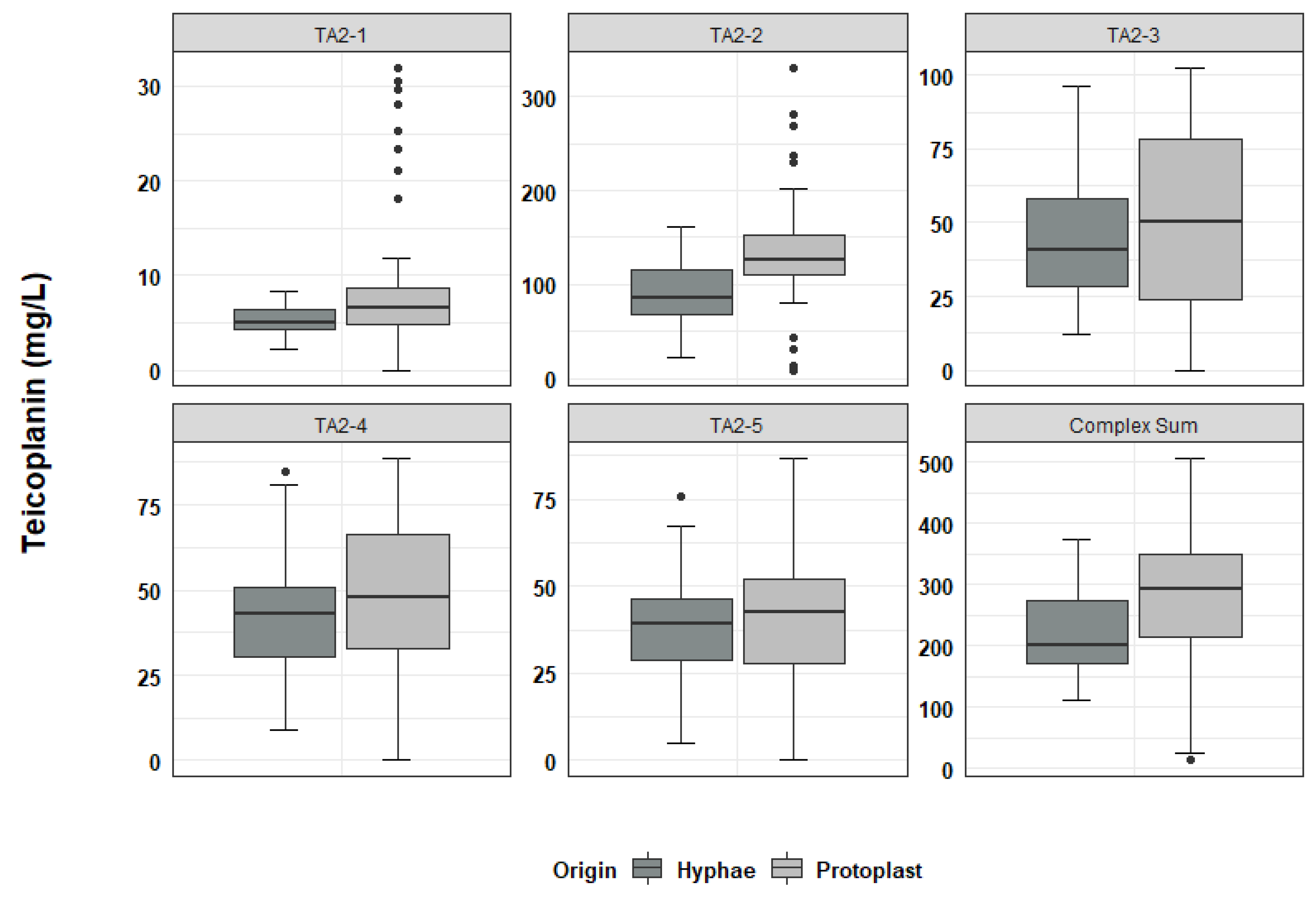
2.3. Analysis of Teicoplanin Production and Complex Composition
3. Discussion
4. Materials and Methods
4.1. Strains and Cultural Conditions
4.2. Protoplast Preparation
4.3. Regeneration of Mycelium from Protoplasts
4.4. DAPI Staining of Mycelium and Protoplasts
4.5. Teicoplanin Production and Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Gestel, J.; Vlamakis, H.; Kolter, R. From Cell Differentiation to Cell Collectives: Bacillus subtilis Uses Division of Labor to Migrate. PLoS Biol. 2015, 13, e1002141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- West, S.A.; Cooper, G.A. Division of Labour in Microorganisms: An Evolutionary Perspective. Nat. Rev. Microbiol. 2016, 14, 716–723. [Google Scholar] [CrossRef] [PubMed]
- Wahl, L.M. The Division of Labor: Genotypic versus Phenotypic Specialization. Am. Nat. 2002, 160, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Hopwood, D.A. Streptomyces in Nature and Medicine: The Antibiotic Makers; Oxford University Press: Oxford, UK, 2007. [Google Scholar]
- Barka, E.A.; Vatsa, P.; Sanchez, L.; Gaveau-Vaillant, N.; Jacquard, C.; Klenk, H.-P.; Clément, C.; Ouhdouch, Y.; van Wezel, G.P. Taxonomy, Physiology, and Natural Products of Actinobacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 1–43. [Google Scholar] [CrossRef] [Green Version]
- Flärdh, K.; Buttner, M.J. Streptomyces morphogenetics: Dissecting differentiation in a filamentous bacterium. Nat. Rev. Microbiol. 2009, 7, 36–49. [Google Scholar] [CrossRef]
- Zhang, Z.; Claessen, D.; Rozen, D.E. Understanding Microbial Divisions of Labor. Front. Microbiol. 2016, 7, 2070. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Du, C.; de Barsy, F.; Liem, M.; Liakopoulos, A.; van Wezel, G.P.; Choi, Y.H.; Claessen, D.; Rozen, D.E. Antibiotic production in Streptomyces is organized by a division of labor through terminal genomic differentiation. Sci. Adv. 2020, 6, eaay5781. [Google Scholar] [CrossRef] [Green Version]
- Lazzarini, A.; Cavaletti, L.; Toppo, G.; Marinelli, F. Rare genera of actinomycetes as potential producers of new antibiotics. Antonie van Leeuwenhoek 2000, 78, 399–405. [Google Scholar] [CrossRef]
- Marcone, G.L.; Binda, E.; Berini, F.; Marinelli, F. Old and new glycopeptide antibiotics: From product to gene and back in the post-genomic era. Biotechnol. Adv. 2018, 36, 534–554. [Google Scholar] [CrossRef]
- Ceccarelli, G.; Alessandri, F.; Oliva, A.; Borrazzo, C.; Dell’Isola, S.; Ialungo, A.M.; Rastrelli, E.; Pelli, M.; Raponi, G.; Turriziani, O.; et al. The role of teicoplanin in the treatment of SARS-CoV-2 infection: A retrospective study in critically ill COVID-19 patients (Tei-COVID study). J. Med Virol. 2021, 93, 4319–4325. [Google Scholar] [CrossRef] [PubMed]
- Azam, F.; Eid, E.E.M.; Almutairi, A. Targeting SARS-CoV-2 main protease by teicoplanin: A mechanistic insight by docking, MM/GBSA and molecular dynamics simulation. J. Mol. Struct. 2021, 1246, 131124. [Google Scholar] [CrossRef] [PubMed]
- EDQM. Monografia della Farmacopea Europea. In Teicoplanin Monograph, 10th ed.; 2019; p. 3967. Available online: https://www.edqm.eu/en/european-pharmacopoeia-ph-eur-10th-edition (accessed on 14 October 2021).
- Vobis, G. The genus Actinoplanes and related genera. In The Prokaryotes; Springer: New York, NY, USA, 2006. [Google Scholar]
- Marcone, G.L.; Carrano, L.; Marinelli, F.; Beltrametti, F. Protoplast preparation and reversion to the normal filamentous growth in antibiotic-producing uncommon actinomycetes. J. Antibiot. 2010, 63, 83–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flärdh, K. Growth polarity and cell division in Streptomyces. Curr. Opin. Microbiol. 2003, 6, 564–571. [Google Scholar] [CrossRef]
- Flärdh, K. Cell polarity and the control of apical growth in Streptomyces. Curr. Opin. Microbiol. 2010, 13, 758–765. [Google Scholar] [CrossRef]
- Donczew, M.; Mackiewicz, P.; Wróbel, A.; Flärdh, K.; Zakrzewska-Czerwińska, J.; Jakimowicz, D. ParA and ParB coordinate chromosome segregation with cell elongation and division during Streptomyces sporulation. Open Biol. 2016, 6, 150263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borghi, A.; Coronelli, C.; Faniuolo, L.; Allievi, G.; Pallanza, R.; Gallo, G.G. Teichomycins, new antibiotics from Actinoplanes teichomyceticus nov. sp. IV. Separation and characterization of the components of teichomycin (teicoplanin). J. Antibiot. 1984, 37, 615–620. [Google Scholar] [CrossRef]
- The Japanese Pharmacopoeia, 17th ed.; The Ministry of Health, Labour and Welfare: Tokyo, Japan, 1982. Available online: https://jpdb.nihs.go.jp/jp/DetailList_en.aspx?submit=Detail(en)&keyword=Teicoplanin (accessed on 14 October 2021).
- Borghi, A.; Edwards, D.; Zerilli, L.F.; Lancini, G.C. Factors affecting the normal and branched-chain acyl moieties of teicoplanin components produced by Actinoplanes teichomyceticus. J. Gen. Microbiol. 1991, 137, 587–592. [Google Scholar] [CrossRef] [Green Version]
- Baltz, R.H. Genetic methods and strategies for secondary metabolite yield improvement in actinomycetes. Antonie van Leeuwenhoek 2001, 79, 251–259. [Google Scholar] [CrossRef]
- Zhang, Y.-X.; Perry, K.; Vinci, V.A.; A Powell, K.; Stemmer, W.P.C.; Del Cardayré, S.B. Genome shuffling leads to rapid phenotypic improvement in bacteria. Nat. Cell Biol. 2002, 415, 644–646. [Google Scholar] [CrossRef]
- Yushchuk, O.; Homoniuk, V.; Datsiuk, Y.; Ostash, B.; Marinelli, F.; Fedorenko, V. Development of a gene expression system for the uncommon actinomycete Actinoplanes rectilineatus NRRL B-16090. J. Appl. Genet. 2020, 61, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Yushchuk, O.; Andreo-Vidal, A.; Marcone, G.L.; Bibb, M.; Marinelli, F.; Binda, E. New Molecular Tools for Regulation and Improvement of A40926 Glycopeptide Antibiotic Production in Nonomuraea gerenzanensis ATCC 39727. Front. Microbiol. 2020, 11, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horbal, L.; Kobylyanskyy, A.; Truman, A.W.; Zaburranyi, N.; Ostash, B.; Luzhetskyy, A.; Marinelli, F.; Fedorenko, V. The pathway-specific regulatory genes, tei15* and tei16*, are the master switches of teicoplanin production in Actinoplanes teichomyceticus. Appl. Microbiol. Biotechnol. 2014, 98, 9295–9309. [Google Scholar] [CrossRef] [PubMed]
- Binda, E.; Berini, F.; Marinelli, F.; Bava, A.; Beltrametti, F. Heterogeneous A40926 Self-Resistance Profile in Nonomuraea gerenzanensis Population Informs Strain Improvement. Fermentation 2021, 7, 140. [Google Scholar] [CrossRef]
- Komatsu, M.; Uchiyama, T.; Ōmura, S.; Cane, D.E.; Ikeda, H. Genome-minimized Streptomyces host for the heterologous expression of secondary metabolism. Proc. Natl. Acad. Sci. USA 2010, 107, 2646–2651. [Google Scholar] [CrossRef] [Green Version]
- Zacharia, V.M.; Ra, Y.; Sue, C.; Alcala, E.; Reaso, J.N.; Ruzin, S.E.; Traxler, M.F. Genetic Network Architecture and Environmental Cues Drive Spatial Organization of Phenotypic Division of Labor in Streptomyces coelicolor. mBio 2021, 12. [Google Scholar] [CrossRef]
- Birch, A.; Häusler, A.; Hütter, R. Genome rearrangement and genetic instability in Streptomyces spp. J. Bacteriol. 1990, 172, 4138–4142. [Google Scholar] [CrossRef] [Green Version]
- Volff, J.-N.; Altenbuchner, J. Genetic instability of the Streptomyces chromosome. Mol. Microbiol. 1998, 27, 239–246. [Google Scholar] [CrossRef]
- Leblond, P.; Demuyter, P.; Moutier, L.; Laakel, M.; Decaris, B.; Simonet, J.M. Hypervariability, a new phenomenon of genetic instability, related to DNA amplification in Streptomyces ambofaciens. J. Bacteriol. 1989, 171, 419–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramijan, K.; Zhang, Z.; van Wezel, G.P.; Claessen, D. Genome rearrangements and megaplasmid loss in the filamentous bacterium Kitasatospora viridifaciens are associated with protoplast formation and regeneration. Antonie van Leeuwenhoek 2020, 113, 825–837. [Google Scholar] [CrossRef] [Green Version]
- Taurino, C.; Frattini, L.; Marcone, G.L.; Gastaldo, L.; Marinelli, F. Actinoplanes teichomyceticus ATCC 31121 as a cell factory for producing teicoplanin. Microb. Cell Factories 2011, 10, 82. [Google Scholar] [CrossRef] [Green Version]
- Parenti, F.; Beretta, G.; Berti, M.; Arioli, V. Teichomycins, new antibiotics from Actinoplanes teichomyceticus nov. sp. I. Description of the producer strain, fermentation studies and biological properties. J. Antibiot. 1978, 31, 276–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beltrametti, F.; Lazzarini, A.; Brunati, C.; Selva, E.; Marinelli, F. Production of demannosyl-A40926 by a Nonomuraea sp. ATCC 39727 mutant strain. J. Antibiot. 2003, 56, 310–313. [Google Scholar] [CrossRef] [Green Version]
- Hobbs, G.; Frazer, C.M.; Gardner, D.C.J.; Cullum, J.A.; Oliver, S.G. Dispersed growth of Streptomyces in liquid culture. Appl. Microbiol. Biotechnol. 1989, 31, 272–277. [Google Scholar] [CrossRef]
- Beltrametti, F.; Consolandi, A.; Carrano, L.; Bagatin, F.; Rossi, R.; Leoni, L.; Zennaro, E.; Selva, E.; Marinelli, F. Resistance to Glycopeptide Antibiotics in the Teicoplanin Producer Is Mediated by van Gene Homologue Expression Directing the Synthesis of a Modified Cell Wall Peptidoglycan. Antimicrob. Agents Chemother. 2007, 51, 1135–1141. [Google Scholar] [CrossRef] [Green Version]
- Beltrametti, F.; Barucco, D.; Rossi, R.; Selva, E.; Marinelli, F. Protoplast Fusion and Gene Recombination in the Uncommon Actinomycete Planobispora rosea Producing GE2270. J. Antibiot. 2007, 60, 447–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirahama, T.; Furumai, T.; Okanishi, M. A Modified Regeneration Method for Streptomycete Protoplasts. Agric. Biol. Chem. 1981, 45, 1271–1273. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: http://www.r-project.org/index.html (accessed on 14 October 2021).

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mellere, L.; Bava, A.; Capozzoli, C.; Branduardi, P.; Berini, F.; Beltrametti, F. Strain Improvement and Strain Maintenance Revisited. The Use of Actinoplanes teichomyceticus ATCC 31121 Protoplasts in the Identification of Candidates for Enhanced Teicoplanin Production. Antibiotics 2022, 11, 24. https://doi.org/10.3390/antibiotics11010024
Mellere L, Bava A, Capozzoli C, Branduardi P, Berini F, Beltrametti F. Strain Improvement and Strain Maintenance Revisited. The Use of Actinoplanes teichomyceticus ATCC 31121 Protoplasts in the Identification of Candidates for Enhanced Teicoplanin Production. Antibiotics. 2022; 11(1):24. https://doi.org/10.3390/antibiotics11010024
Chicago/Turabian StyleMellere, Luca, Adriana Bava, Carmine Capozzoli, Paola Branduardi, Francesca Berini, and Fabrizio Beltrametti. 2022. "Strain Improvement and Strain Maintenance Revisited. The Use of Actinoplanes teichomyceticus ATCC 31121 Protoplasts in the Identification of Candidates for Enhanced Teicoplanin Production" Antibiotics 11, no. 1: 24. https://doi.org/10.3390/antibiotics11010024
APA StyleMellere, L., Bava, A., Capozzoli, C., Branduardi, P., Berini, F., & Beltrametti, F. (2022). Strain Improvement and Strain Maintenance Revisited. The Use of Actinoplanes teichomyceticus ATCC 31121 Protoplasts in the Identification of Candidates for Enhanced Teicoplanin Production. Antibiotics, 11(1), 24. https://doi.org/10.3390/antibiotics11010024