
Lacticaseicin 30 and Colistin as a Promising Antibiotic Formulation against Gram-Negative β-Lactamase-Producing Strains and Colistin-Resistant Strains

 , ,
, ,  and
and
Abstract
1. Introduction
2. Results
2.1. Multifaceted Resistance of Clinical Gram-Negative Bacteria Used in This Work
2.2. Amplification and Sequencing of the mcr Gene Involved in the Resistance to Colistin
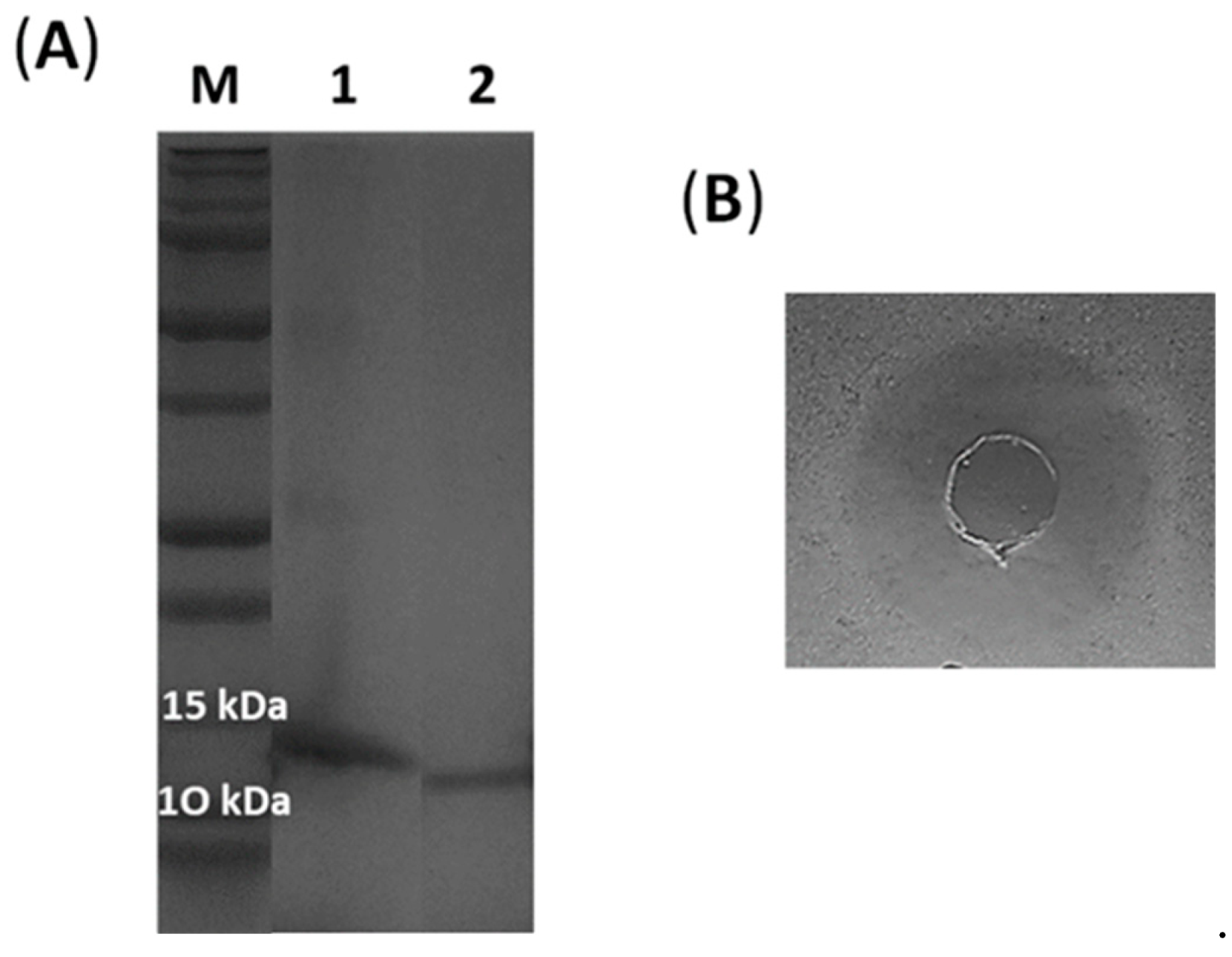
2.3. Expression and Characterization of the Recombinant Lacticaseicin 30
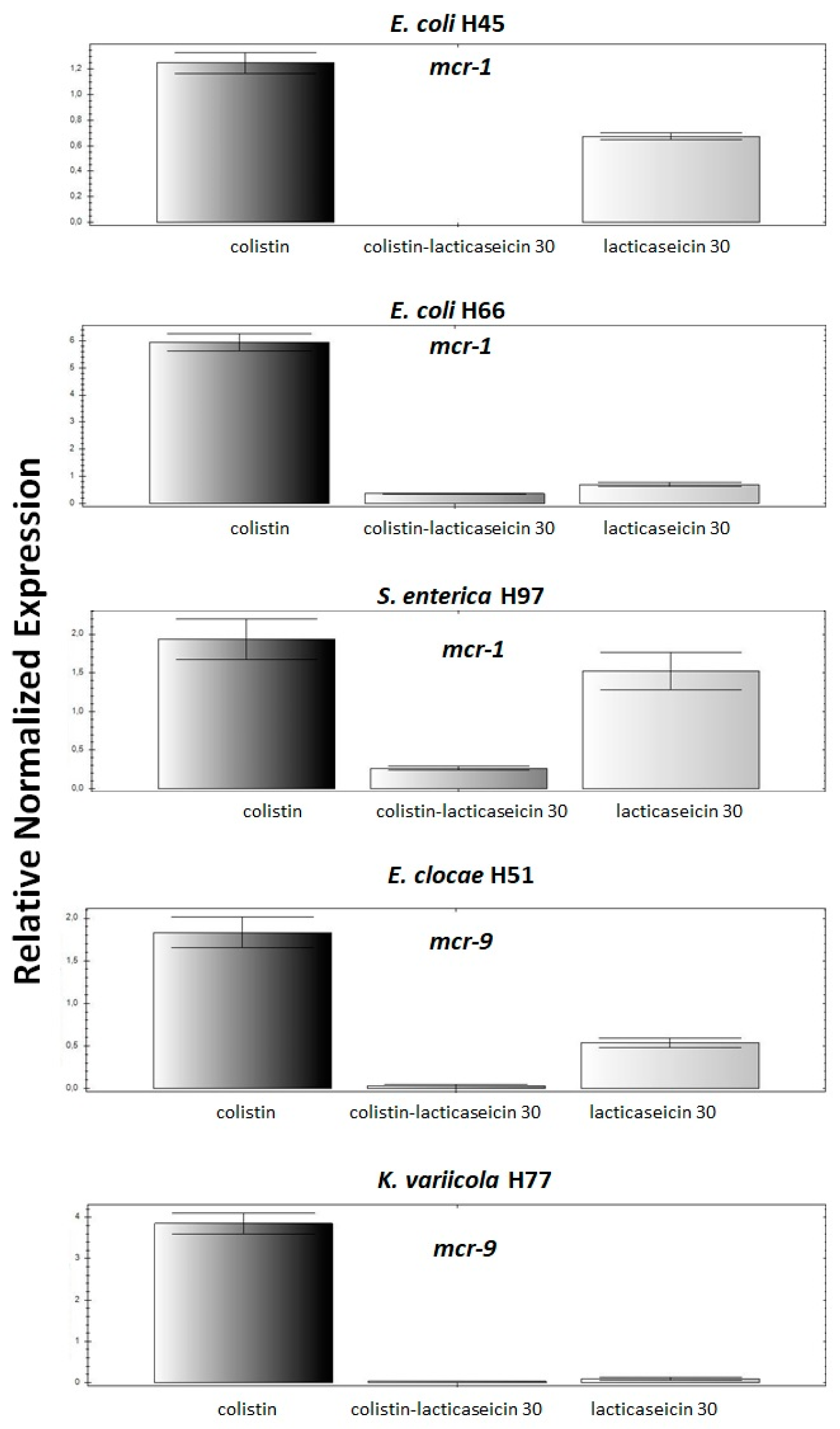
2.4. Lacticaseicin 30 Potentiates the Activity of Colistin and Attenuates the Expression of mcr Genes in the Clinical Strains
2.5. The Synergetic Interaction between Lacticaseicin 30-Colistin Downregulated Expression of mcr Gene
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Culture Conditions
4.2. Antibiotic Susceptibility of Clinical Strains
4.3. Genomic DNA Extraction
4.4. Detection of mcr Gene in the Clinical Strains Exhibiting a Colistin Resistant Phenotype
4.5. Random Amplification of Polymorphic DNA PCR (RAPD PCR)
4.6. Production and Purification of Recombinant Lacticaseicin 30
4.7. Determination of Minimal Inhibitory Concentrations (MICs) in Checkerboard Assays
4.8. Total RNAs Extraction and cDNA Synthesis
4.9. Quantitative PCR (qPCR) and Analysis of mcr Genes Expression
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Jim O’Neill. Nat. Rev. Drug Discov. 2016, 15, 526. [CrossRef]
- Goel, N.; Fatima, S.W.; Kumar, S.; Sinha, R.; Khare, S.K. Antimicrobial Resistance in Biofilms: Exploring Marine Actinobacteria as a Potential Source of Antibiotics and Biofilm Inhibitors. Biotechnol. Rep. 2021, 30, e00613. [Google Scholar] [CrossRef]
- Brinkac, L.; Voorhies, A.; Gomez, A.; Nelson, K.E. The Threat of Antimicrobial Resistance on the Human Microbiome. Microb. Ecol. 2017, 74, 1001–1008. [Google Scholar] [CrossRef]
- Ventola, C.L. The Antibiotic Resistance Crisis: Part 1: Causes and Threats. Pharm. Ther. 2015, 40, 277–283. [Google Scholar]
- Sun, Y.; Kang, C.; Liu, F.; Zhou, Y.; Luo, L.; Qiao, H. RGD Peptide-Based Target Drug Delivery of Doxorubicin Nanomedicine. Drug Dev. Res. 2017, 78, 283–291. [Google Scholar] [CrossRef]
- Aslam, B.; Wang, W.; Arshad, M.I.; Khurshid, M.; Muzammil, S.; Rasool, M.H.; Nisar, M.A.; Alvi, R.F.; Aslam, M.A.; Qamar, M.U.; et al. Antibiotic Resistance: A Rundown of a Global Crisis. Infect. Drug Resist. 2018, 11, 1645–1658. [Google Scholar] [CrossRef]
- Naghmouchi, K.; Drider, D.; Fliss, I. Action of Divergicin M35, a Class IIa Bacteriocin, on Liposomes and Listeria. J. Appl. Microbiol. 2007, 102, 1508–1517. [Google Scholar] [CrossRef]
- Drider, D.; Rebuffat, S. Prokaryotic Antimicrobial Peptides: From Genes to Applications; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2011; ISBN 978-1-4419-7692-5. [Google Scholar]
- Hammami, R.; Fernandez, B.; Lacroix, C.; Fliss, I. Anti-Infective Properties of Bacteriocins: An Update. Cell Mol. Life Sci. 2013, 70, 2947–2967. [Google Scholar] [CrossRef]
- Flaherty, R.A.; Freed, S.D.; Lee, S.W. The Wide World of Ribosomally Encoded Bacterial Peptides. PLoS Pathog. 2014, 10, e1004221. [Google Scholar] [CrossRef]
- Drider, D. Gut Microbiota Is an Important Source of Bacteriocins and Their In Situ Expression Can Be Explored for Treatment of Bacterial Infections. Probiotics Antimicrob. Proteins 2021, 13, 1759–1765. [Google Scholar] [CrossRef]
- Cotter, P.D.; Ross, R.P.; Hill, C. Bacteriocins—A Viable Alternative to Antibiotics? Nat. Rev. Microbiol. 2013, 11, 95–105. [Google Scholar] [CrossRef]
- Kumariya, R.; Garsa, A.K.; Rajput, Y.S.; Sood, S.K.; Akhtar, N.; Patel, S. Bacteriocins: Classification, Synthesis, Mechanism of Action and Resistance Development in Food Spoilage Causing Bacteria. Microb. Pathog. 2019, 128, 171–177. [Google Scholar] [CrossRef]
- Drider, D.; Fimland, G.; Hechard, Y.; McMullen, L.M.; Prevost, H. The Continuing Story of Class IIa Bacteriocins. Microbiol. Mol. Biol. Rev. 2006, 70, 564–582. [Google Scholar] [CrossRef]
- Soltani, S.; Hammami, R.; Cotter, P.D.; Rebuffat, S.; Said, L.B.; Gaudreau, H.; Bédard, F.; Biron, E.; Drider, D.; Fliss, I. Bacteriocins as a New Generation of Antimicrobials: Toxicity Aspects and Regulations. FEMS Microbiol. Rev. 2021, 45, fuaa039. [Google Scholar] [CrossRef]
- Alvarez-Sieiro, P.; Montalbán-López, M.; Mu, D.; Kuipers, O.P. Bacteriocins of Lactic Acid Bacteria: Extending the Family. Appl. Microbiol. Biotechnol. 2016, 100, 2939–2951. [Google Scholar] [CrossRef]
- Pérez-Ramos, A.; Madi-Moussa, D.; Coucheney, F.; Drider, D. Current Knowledge of the Mode of Action and Immunity Mechanisms of LAB-Bacteriocins. Microorganisms 2021, 9, 2107. [Google Scholar] [CrossRef]
- Stern, N.J.; Svetoch, E.A.; Eruslanov, B.V.; Perelygin, V.V.; Mitsevich, E.V.; Mitsevich, I.P.; Pokhilenko, V.D.; Levchuk, V.P.; Svetoch, O.E.; Seal, B.S. Isolation of a Lactobacillus Salivarius Strain and Purification of Its Bacteriocin, Which Is Inhibitory to Campylobacter Jejuni in the Chicken Gastrointestinal System. Antimicrob. Agents Chemother. 2006, 50, 3111–3116. [Google Scholar] [CrossRef]
- Messaoudi, S.; Kergourlay, G.; Dalgalarrondo, M.; Choiset, Y.; Ferchichi, M.; Prévost, H.; Pilet, M.-F.; Chobert, J.-M.; Manai, M.; Dousset, X. Purification and Characterization of a New Bacteriocin Active against Campylobacter Produced by Lactobacillus Salivarius SMXD51. Food Microbiol. 2012, 32, 129–134. [Google Scholar] [CrossRef]
- Belguesmia, Y.; Hazime, N.; Kempf, I.; Boukherroub, R.; Drider, D. New Bacteriocins from Lacticaseibacillus Paracasei CNCM I-5369 Adsorbed on Alginate Nanoparticles Are Very Active against Escherichia Coli. Int. J. Mol. Sci. 2020, 21, 8654. [Google Scholar] [CrossRef]
- Raetz, C.R.H.; Whitfield, C. Lipopolysaccharide Endotoxins. Annu. Rev. Biochem. 2002, 71, 635–700. [Google Scholar] [CrossRef]
- Vaara, M. Agents That Increase the Permeability of the Outer Membrane. Microbiol. Rev. 1992, 56, 395–411. [Google Scholar] [CrossRef]
- Ayoub Moubareck, C. Polymyxins and Bacterial Membranes: A Review of Antibacterial Activity and Mechanisms of Resistance. Membranes 2020, 10, 181. [Google Scholar] [CrossRef] [PubMed]
- Bialvaei, A.Z.; Samadi Kafil, H. Colistin, Mechanisms and Prevalence of Resistance. Curr. Med. Res. Opin. 2015, 31, 707–721. [Google Scholar] [CrossRef] [PubMed]
- Madi-Moussa, D.; Coucheney, F.; Drider, D. Expression of Five Class II Bacteriocins with Activity against Escherichia Coli in Lacticaseibacillus Paracasei CNCM I-5369, and in a Heterologous Host. Biotechnol. Rep. 2021, 30, e00632. [Google Scholar] [CrossRef] [PubMed]
- Weldhagen, G.F.; Poirel, L.; Nordmann, P. Ambler Class A Extended-Spectrum β-Lactamases in Pseudomonas Aeruginosa: Novel Developments and Clinical Impact. Antimicrob. Agents Chemother. 2003, 47, 2385–2392. [Google Scholar] [CrossRef]
- Meini, S.; Tascini, C.; Cei, M.; Sozio, E.; Rossolini, G.M. AmpC β-Lactamase-Producing Enterobacterales: What a Clinician Should Know. Infection 2019, 47, 363–375. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- De Kraker, M.E.A.; Stewardson, A.J.; Harbarth, S. Will 10 Million People Die a Year Due to Antimicrobial Resistance by 2050? PLoS Med. 2016, 13, e1002184. [Google Scholar] [CrossRef]
- O’Neill, J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations. Rev. Antimicrob. Resist. 2016, 1–84. [Google Scholar]
- Friedman, N.D.; Temkin, E.; Carmeli, Y. The Negative Impact of Antibiotic Resistance. Clin. Microbiol. Infect. 2016, 22, 416–422. [Google Scholar] [CrossRef]
- World Health Organization. Global Action Plan on Antimicrobial Resistance; World Health Organization: Geneva, Switzerland, 2015; ISBN 978-92-4-150976-3. [Google Scholar]
- European Union EU. Action on Antimicrobial Resistance. Available online: https://ec.europa.eu/health/antimicrobial-resistance/eu-action-on-antimicrobial-resistance_en (accessed on 19 October 2021).
- Hamel, M.; Rolain, J.-M.; Baron, S.A. The History of Colistin Resistance Mechanisms in Bacteria: Progress and Challenges. Microorganisms 2021, 9, 442. [Google Scholar] [CrossRef]
- Jafari, F.; Elyasi, S. Prevention of Colistin Induced Nephrotoxicity: A Review of Preclinical and Clinical Data. Expert Rev. Clin. Pharmacol. 2021, 14, 1113–1131. [Google Scholar] [CrossRef] [PubMed]
- Lhermie, G.; la Ragione, R.M.; Weese, J.S.; Olsen, J.E.; Christensen, J.P.; Guardabassi, L. Indications for the Use of Highest Priority Critically Important Antimicrobials in the Veterinary Sector. J. Antimicrob. Chemother. 2020, 75, 1671–1680. [Google Scholar] [CrossRef]
- Olaitan, A.O.; Morand, S.; Rolain, J.-M. Mechanisms of Polymyxin Resistance: Acquired and Intrinsic Resistance in Bacteria. Front. Microbiol. 2014, 5, 643. [Google Scholar] [CrossRef]
- Liu, Y.-Y.; Wang, Y.; Walsh, T.R.; Yi, L.-X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of Plasmid-Mediated Colistin Resistance Mechanism MCR-1 in Animals and Human Beings in China: A Microbiological and Molecular Biological Study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Gao, R.; Hu, Y.; Li, Z.; Sun, J.; Wang, Q.; Lin, J.; Ye, H.; Liu, F.; Srinivas, S.; Li, D.; et al. Dissemination and Mechanism for the MCR-1 Colistin Resistance. PLoS Pathog. 2016, 12, e1005957. [Google Scholar] [CrossRef]
- Gharaibeh, M.H.; Shatnawi, S.Q. An Overview of Colistin Resistance, Mobilized Colistin Resistance Genes Dissemination, Global Responses, and the Alternatives to Colistin: A Review. Vet. World 2019, 12, 1735. [Google Scholar] [CrossRef] [PubMed]
- Lei, C.-W.; Zhang, Y.; Wang, Y.-T.; Wang, H.-N. Detection of Mobile Colistin Resistance Gene Mcr-10.1 in a Conjugative Plasmid from Enterobacter Roggenkampii of Chicken Origin in China. Antimicrob. Agents Chemother. 2020, 64, e01191-20. [Google Scholar] [CrossRef]
- Chikindas, M.L.; Weeks, R.; Drider, D.; Chistyakov, V.A.; Dicks, L.M. Functions and Emerging Applications of Bacteriocins. Curr. Opin. Biotechnol. 2018, 49, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Zgheib, H.; Drider, D.; Belguesmia, Y. Broadening and Enhancing Bacteriocins Activities by Association with Bioactive Substances. Int. J. Environ. Res. Public Health 2020, 17, 7835. [Google Scholar] [CrossRef]
- Cantón, R.; Novais, A.; Valverde, A.; Machado, E.; Peixe, L.; Baquero, F.; Coque, T.M. Prevalence and Spread of Extended-Spectrum β-Lactamase-Producing Enterobacteriaceae in Europe. Clin. Microbiol. Infect. 2008, 14, 144–153. [Google Scholar] [CrossRef]
- Bradford, P.A. Extended-Spectrum β-Lactamases in the 21st Century: Characterization, Epidemiology, and Detection of This Important Resistance Threat. Clin. Microbiol. Rev. 2001, 14, 933–951. [Google Scholar] [CrossRef] [PubMed]
- Castanheira, M.; Simner, P.J.; Bradford, P.A. Extended-Spectrum β-Lactamases: An Update on Their Characteristics, Epidemiology and Detection. JAC Antimicrob. Resist. 2021, 3, dlab092. [Google Scholar] [CrossRef]
- Paterson, D.L.; Bonomo, R.A. Extended-Spectrum Beta-Lactamases: A Clinical Update. Clin. Microbiol. Rev. 2005, 18, 657–686. [Google Scholar] [CrossRef]
- Ghafourian, S.; Sadeghifard, N.; Soheili, S.; Sekawi, Z. Extended Spectrum Beta-Lactamases: Definition, Classification and Epidemiology. Curr. Issues Mol. Biol. 2015, 17, 11–21. [Google Scholar] [PubMed]
- Field, D.; Seisling, N.; Cotter, P.D.; Ross, R.P.; Hill, C. Synergistic Nisin-Polymyxin Combinations for the Control of Pseudomonas Biofilm Formation. Front. Microbiol. 2016, 7, 1713. [Google Scholar] [CrossRef]
- Xu, X.; Xu, L.; Yuan, G.; Wang, Y.; Qu, Y.; Zhou, M. Synergistic Combination of Two Antimicrobial Agents Closing Each Other’s Mutant Selection Windows to Prevent Antimicrobial Resistance. Sci. Rep. 2018, 8, 7237. [Google Scholar] [CrossRef]
- Al Atya, A.K.; Belguesmia, Y.; Chataigne, G.; Ravallec, R.; Vachée, A.; Szunerits, S.; Boukherroub, R.; Drider, D. Anti-MRSA Activities of Enterocins DD28 and DD93 and Evidences on Their Role in the Inhibition of Biofilm Formation. Front. Microbiol. 2016, 7, 817. [Google Scholar] [CrossRef]
- Naghmouchi, K.; Belguesmia, Y.; Baah, J.; Teather, R.; Drider, D. Antibacterial Activity of Class I and IIa Bacteriocins Combined with Polymyxin E against Resistant Variants of Listeria Monocytogenes and Escherichia Coli. Res. Microbiol. 2011, 162, 99–107. [Google Scholar] [CrossRef]
- Naghmouchi, K.; Baah, J.; Hober, D.; Jouy, E.; Rubrecht, C.; Sané, F.; Drider, D. Synergistic Effect between Colistin and Bacteriocins in Controlling Gram-Negative Pathogens and Their Potential to Reduce Antibiotic Toxicity in Mammalian Epithelial Cells. Antimicrob. Agents Chemother. 2013, 57, 2719–2725. [Google Scholar] [CrossRef] [PubMed]
- Jorge, P.; Pérez-Pérez, M.; Pérez Rodríguez, G.; Pereira, M.O.; Lourenço, A. A Network Perspective on Antimicrobial Peptide Combination Therapies: The Potential of Colistin, Polymyxin B and Nisin. Int. J. Antimicrob. Agents 2017, 49, 668–676. [Google Scholar] [CrossRef] [PubMed]
- Balto, H.A.; Shakoor, Z.A.; Kanfar, M.A. Combined Effect of a Mixture of Tetracycline, Acid, and Detergent, and Nisin against Enterococcus Faecalis and Actinomyces Viscosus Biofilms. Saudi Med. J. 2015, 36, 211–215. [Google Scholar] [CrossRef] [PubMed]
- Mathur, H.; Field, D.; Rea, M.C.; Cotter, P.D.; Hill, C.; Ross, R.P. Bacteriocin-Antimicrobial Synergy: A Medical and Food Perspective. Front. Microbiol. 2017, 8, 1205. [Google Scholar] [CrossRef]
- Ellis, J.-C.; Ross, R.P.; Hill, C. Nisin Z and Lacticin 3147 Improve Efficacy of Antibiotics against Clinically Significant Bacteria. Future Microbiol. 2019, 14, 1573–1587. [Google Scholar] [CrossRef]
- Angelopoulou, A.; Field, D.; Pérez-Ibarreche, M.; Warda, A.K.; Hill, C.; Ross, R.P. Vancomycin and Nisin A Are Effective against Biofilms of Multi-Drug Resistant Staphylococcus Aureus Isolates from Human Milk. PLoS ONE 2020, 15, e0233284. [Google Scholar] [CrossRef]
- Belguesmia, Y.; Spano, G.; Drider, D. Potentiating Effects of Leaderless Enterocin DD14 in Combination with Methicillin on Clinical Methicillin-Resistant Staphylococcus Aureus S1 Strain. Microbiol. Res. 2021, 252, 126864. [Google Scholar] [CrossRef]
- Bendjeddou, K.; Hamma-Faradji, S.; Meddour, A.A.; Belguesmia, Y.; Cudennec, B.; Bendali, F.; Daube, G.; Taminiau, B.; Drider, D. Gut Microbiota, Body Weight and Histopathological Examinations in Experimental Infection by Methicillin-Resistant Staphylococcus Aureus: Antibiotic versus Bacteriocin. Benef. Microbes 2021, 12, 295–305. [Google Scholar] [CrossRef]
- Zgheib, H.; Belguesmia, Y.; Boukherroub, R.; Drider, D. Alginate Nanoparticles Enhance Anti-Clostridium Perfringens Activity of the Leaderless Two-Peptide Enterocin DD14 and Affect Expression of Some Virulence Factors. Probiotics Antimicrob. Proteins 2021, 4, 1213–1227. [Google Scholar] [CrossRef]
- Ling, Z.; Yin, W.; Shen, Z.; Wang, Y.; Shen, J.; Walsh, T.R. Epidemiology of Mobile Colistin Resistance Genes Mcr-1 to Mcr-9. J. Antimicrob. Chemother. 2020, 75, 3087–3095. [Google Scholar] [CrossRef]
- Carroll, L.M.; Gaballa, A.; Guldimann, C.; Sullivan, G.; Henderson, L.O.; Wiedmann, M. Identification of Novel Mobilized Colistin Resistance Gene Mcr-9 in a Multidrug-Resistant, Colistin-Susceptible Salmonella Enterica Serotype Typhimurium Isolate. mBio 2019, 10, e00853-19. [Google Scholar] [CrossRef] [PubMed]
- Zhou, K.; Luo, Q.; Wang, Q.; Huang, C.; Lu, H.; Rossen, J.W.A.; Xiao, Y.; Li, L. Silent Transmission of an IS1294b-Deactivated Mcr-1 Gene with Inducible Colistin Resistance. Int. J. Antimicrob. Agents 2018, 51, 822–828. [Google Scholar] [CrossRef] [PubMed]
- Kieffer, N.; Royer, G.; Decousser, J.-W.; Bourrel, A.-S.; Palmieri, M.; Ortiz de la Rosa, J.-M.; Jacquier, H.; Denamur, E.; Nordmann, P.; Poirel, L. Mcr-9, an Inducible Gene Encoding an Acquired Phosphoethanolamine Transferase in Escherichia Coli, and Its Origin. Antimicrob. Agents Chemother. 2019, 63, e00965-19. [Google Scholar] [CrossRef]
- Kieffer, N.; Nordmann, P.; Poirel, L. Moraxella Species as Potential Sources of MCR-like Polymyxin Resistance Determinants. Antimicrob. Agents Chemother. 2017, 61, e00129-17. [Google Scholar] [CrossRef] [PubMed]
- Bardet, L.; Okdah, L.; le Page, S.; Baron, S.A.; Rolain, J.-M. Comparative Evaluation of the UMIC Colistine Kit to Assess MIC of Colistin of Gram-Negative Rods. BMC Microbiol. 2019, 19, 60. [Google Scholar] [CrossRef]
- Sambrook, J.; Russell, D.W. Agarose Gel Electrophoresis. Cold Spring Harb. Protoc. 2006, 2006, pdb.prot4020. [Google Scholar] [CrossRef]
- Li, J.; Koh, J.-J.; Liu, S.; Lakshminarayanan, R.; Verma, C.S.; Beuerman, R.W. Membrane Active Antimicrobial Peptides: Translating Mechanistic Insights to Design. Front. Neurosci. 2017, 11, 73. [Google Scholar] [CrossRef] [PubMed]
- Borowiak, M.; Baumann, B.; Fischer, J.; Thomas, K.; Deneke, C.; Hammerl, J.A.; Szabo, I.; Malorny, B. Development of a Novel Mcr-6 to Mcr-9 Multiplex PCR and Assessment of Mcr-1 to Mcr-9 Occurrence in Colistin-Resistant Salmonella Enterica Isolates from Environment, Feed, Animals and Food (2011–2018) in Germany. Front. Microbiol. 2020, 11, 80. [Google Scholar] [CrossRef]
- Nielsen, K.L.; Godfrey, P.A.; Stegger, M.; Andersen, P.S.; Feldgarden, M.; Frimodt-Møller, N. Selection of Unique Escherichia Coli Clones by Random Amplified Polymorphic DNA (RAPD): Evaluation by Whole Genome Sequencing. J. Microbiol. Methods 2014, 103, 101–103. [Google Scholar] [CrossRef]
- Deschaght, P.; van Simaey, L.; Decat, E.; van Mechelen, E.; Brisse, S.; Vaneechoutte, M. Rapid Genotyping of Achromobacter Xylosoxidans, Acinetobacter Baumannii, Klebsiella Pneumoniae, Pseudomonas Aeruginosa and Stenotrophomonas Maltophilia Isolates Using Melting Curve Analysis of RAPD-Generated DNA Fragments (McRAPD). Res. Microbiol. 2011, 162, 386–392. [Google Scholar] [CrossRef]
- Albufera, U.; Bhugaloo-Vial, P.; Issack, M.I.; Jaufeerally-Fakim, Y. Molecular Characterization of Salmonella Isolates by REP-PCR and RAPD Analysis. Infect. Genet. Evol. 2009, 9, 322–327. [Google Scholar] [CrossRef]
- Schägger, H. Tricine-SDS-PAGE. Nat. Protoc. 2006, 1, 16–22. [Google Scholar] [CrossRef]
- Tagg, J.R.; McGiven, A.R. Assay System for Bacteriocins. Appl. Microbiol. 1971, 21, 943. [Google Scholar] [CrossRef]
- Ahmad, V.; Khan, M.S.; Jamal, Q.M.S.; Alzohairy, M.A.; Al Karaawi, M.A.; Siddiqui, M.U. Antimicrobial Potential of Bacteriocins: In Therapy, Agriculture and Food Preservation. Int. J. Antimicrob. Agents 2017, 49, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wiegand, I.; Hilpert, K.; Hancock, R.E.W. Agar and Broth Dilution Methods to Determine the Minimal Inhibitory Concentration (MIC) of Antimicrobial Substances. Nat. Protoc. 2008, 3, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Petersen, P.J.; Labthavikul, P.; Jones, C.H.; Bradford, P.A. In Vitro Antibacterial Activities of Tigecycline in Combination with Other Antimicrobial Agents Determined by Chequerboard and Time-Kill Kinetic Analysis. J. Antimicrob. Chemother. 2006, 57, 573–576. [Google Scholar] [CrossRef] [PubMed]
- Postollec, F.; Falentin, H.; Pavan, S.; Combrisson, J.; Sohier, D. Recent Advances in Quantitative PCR (QPCR) Applications in Food Microbiology. Food Microbiol. 2011, 28, 848–861. [Google Scholar] [CrossRef]
- Hurtado, A.; Reguant, C.; Bordons, A.; Rozès, N. Expression of Lactobacillus Pentosus B96 Bacteriocin Genes under Saline Stress. Food Microbiol. 2011, 28, 1339–1344. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Strains | D72A | D72B | D72C | D72D | D72E | Conclusion |
|---|---|---|---|---|---|---|
| Enterobacter cloacae H51 | R | S | R | S | R | EsβL + AmpC Inducible |
| E. coli H52 | R | R | S | S | I | AmpC |
| E. coli H66 | R | R | S | S | S | AmpC |
| Klebsiella oxytoca H40 | R | S | R | S | S | EsβL |
| K. pneumoniae H71 | R | R | R | R | R | EsβL + AmpC + Carbapenemase |
| K. variicola H77 | R | S | I | S | S | EsβL + AmpC |
| K. pneumoniae H79 | R | S | R | S | S | EsβL |
| Strains | mcr Gene | Lacticaseicin 30 (µg/mL) | Colistin (µg/mL) | FIC Lacticaseicin 30 | FIC Colistin | FIC I | Conclusions |
|---|---|---|---|---|---|---|---|
| E. coli 184 | mcr-1 | 40 | 8 | 0.25 | 0.125 | 0.375 | Synergism |
| Enterobacter cloacae H51 | mcr-9 | 40 | 16 | 0.25 | 0.25 | 0.5 | Synergism |
| E. coli H45 | mcr-1 | 40 | 8 | 0.25 | 0.25 | 0.5 | Synergism |
| E. coli H52 | - | 40 | 16 | 0.125 | 0.25 | 0.375 | Synergism |
| E. coli H66 | mcr-1 | 40 | 16 | 0.125 | 0.25 | 0.375 | Synergism |
| Klebsiella oxytoca H40 | mcr-1 | 40 | 64 | 1 | 1 | 2 | Indifference |
| K. pneumoniae H71 | mcr-1 | 40 | 8 | 0.5 | 1 | 1.5 | Indifference |
| K. variicola H77 | mcr-9 | 40 | 8 | 0.25 | 0.25 | 0.5 | Synergism |
| K. pneumoniae H79 | mcr-1 | 40 | 4 | 0.25 | 0.5 | 0.75 | Indifference |
| K. pneumoniae H87 | mcr-1 | 40 | 32 | 1 | 1µ | 2 | Indifference |
| Salmonella enterica H10 | mcr-1 | 40 | 4 | 1 | 1 | 2 | Indifference |
| S. enterica Typhimurium H97 | mcr-1 | 40 | 8 | 0.125 | 0.125 | 0.25 | Synergism |
| Strains | Relevant Characteristics |
|---|---|
| Producing strain | |
| Escherichia coli Rosetta-T7-6His-030 | Strain harboring the plasmid carrying the orf030 gene, encoding the lacticaseicin 30 [25] |
| Control target strains | |
| Escherichia coli ATCC8739 | [20] |
| Escherichia coli 184 | Strain harboring the plasmid carrying the mcr-1 gene, responsible for colistin resistance [20] |
| Gram-negative clinical target strains | |
| Enterobacter cloacae H51 | This work |
| Escherichia coli H45 | This work |
| Escherichia coli H52 | This work |
| Escherichia coli H66 | This work |
| Klebsiella oxytoca H40 | This work |
| Klebsiella pneumoniae β-lactamase H71 | This work |
| Klebsiella variicola β-lactamase H77 | This work |
| Klebisella pneumoniae β-lactamase H79 | This work |
| Klebsiella pneumoniae β-lactamase H87 | This work |
| Salmonella enterica Enteritidis H10 | This work |
| Salmonella enterica Typhimurium H97 | This work |
| Target Gene | Name | Sequence 5′ → 3′ | Tm (°C) | Size (bp) | References |
|---|---|---|---|---|---|
| Primers used for mcr gene detection | |||||
| mcr-1 | F-mcr-1 | AGTCCGTTTGTTCTTGTGGC | 58 | 320 | [69] |
| R-mcr-1 | AGATCCTTGGTCTCGGCTTG | ||||
| mcr-2 | F-mcr-2 | CAAGTGTGTTGGTCGCAGTT | 58 | 715 | [69] |
| R-mcr-2 | TCTAGCCCGACAAGCATACC | ||||
| mcr-3 | F-mcr-3 | AAATAAAAATTGTTCCGCTTATG | 58 | 929 | [69] |
| R-mcr-3 | AATGGAGATCCCCGTTTTT | ||||
| mcr-4 | F-mcr-4 | TCACTTTCATCACTGCGTTG | 58 | 1116 | [69] |
| R-mcr-4 | TTGGTCCATGACTACCAATG | ||||
| mcr-5 | F-mcr-5 | ATGCGGTTGTCTGCATTTATC | 58 | 1644 | [69] |
| R-mcr-5 | TCATTGTGGTTGTCCTTTTCTG | ||||
| mcr-6 | F-mcr-6 | AGCTATGTCAATCCCGTGAT | 52 | 252 | [70] |
| R-mcr-6 | ATTGGCTAGGTTGTCAATC | ||||
| mcr-7 | F-mcr-7 | GCCCTTCTTTTCGTTGTT | 50 | 551 | [70] |
| R-mcr-7 | GGTTGGTCTCTTTCTCGT | ||||
| mcr-8 | F-mcr-8 | TCAACAATTCTACAAAGCGTG | 53 | 856 | [70] |
| R-mcr-8 | AATGCTGCGCGAATGAAG | ||||
| mcr-9 | F-mcr-9 | TTCCCTTTGTTCTGGTTG | 55 | 1011 | [70] |
| R-mcr-9 | GCAGGTAATAAGTCGGTC | ||||
| mcr-10 | F-mcr-10 | GGACCGACCTATTACCAGCG | 64 | [41] | |
| R-mcr-10 | GGCATTATGCTGCAGACACG | ||||
| Primers used for qPCR analyses | |||||
| mcr-1 | F-mcr-1-qPCR | CGCGATGCTACTGATCACCA | 58 | 100 | In this study |
| R-mcr-1-qPCR | AAAATAACTGGTCACCGCGC | ||||
| mcr-9 | F-mcr-9-qPCR | ATCCGTTCCGTGCATGTTCT | 58 | 100 | In this study |
| R-mcr-9-qPCR | CACCGGTTTTCTGCACGATG | ||||
| 16sRNA | F-16sRNA-qPCR | GTAGGTGGCAAGCGTTATCC | 58 | 101 | In this study |
| R-16sRNA-qPCR | GATGCGCTTCCTCGGTTAAG | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Madi-Moussa, D.; Belguesmia, Y.; Charlet, A.; Drider, D.; Coucheney, F. Lacticaseicin 30 and Colistin as a Promising Antibiotic Formulation against Gram-Negative β-Lactamase-Producing Strains and Colistin-Resistant Strains. Antibiotics 2022, 11, 20. https://doi.org/10.3390/antibiotics11010020
Madi-Moussa D, Belguesmia Y, Charlet A, Drider D, Coucheney F. Lacticaseicin 30 and Colistin as a Promising Antibiotic Formulation against Gram-Negative β-Lactamase-Producing Strains and Colistin-Resistant Strains. Antibiotics. 2022; 11(1):20. https://doi.org/10.3390/antibiotics11010020
Chicago/Turabian StyleMadi-Moussa, Désiré, Yanath Belguesmia, Audrey Charlet, Djamel Drider, and Françoise Coucheney. 2022. "Lacticaseicin 30 and Colistin as a Promising Antibiotic Formulation against Gram-Negative β-Lactamase-Producing Strains and Colistin-Resistant Strains" Antibiotics 11, no. 1: 20. https://doi.org/10.3390/antibiotics11010020
APA StyleMadi-Moussa, D., Belguesmia, Y., Charlet, A., Drider, D., & Coucheney, F. (2022). Lacticaseicin 30 and Colistin as a Promising Antibiotic Formulation against Gram-Negative β-Lactamase-Producing Strains and Colistin-Resistant Strains. Antibiotics, 11(1), 20. https://doi.org/10.3390/antibiotics11010020