
Strategies for Antimicrobial Peptides Immobilization on Surfaces to Prevent Biofilm Growth on Biomedical Devices

 , and
, and
Abstract
:1. Introduction
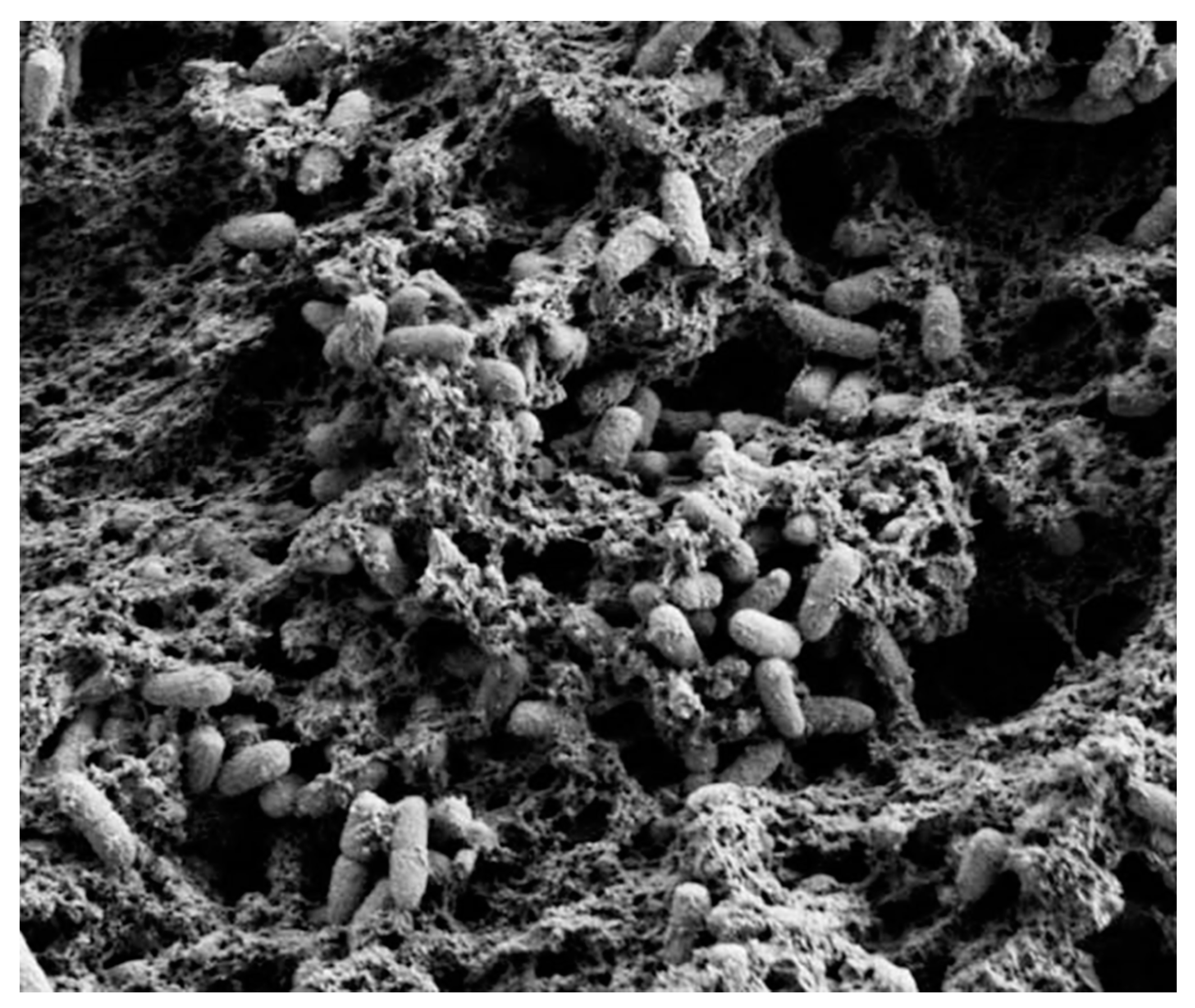
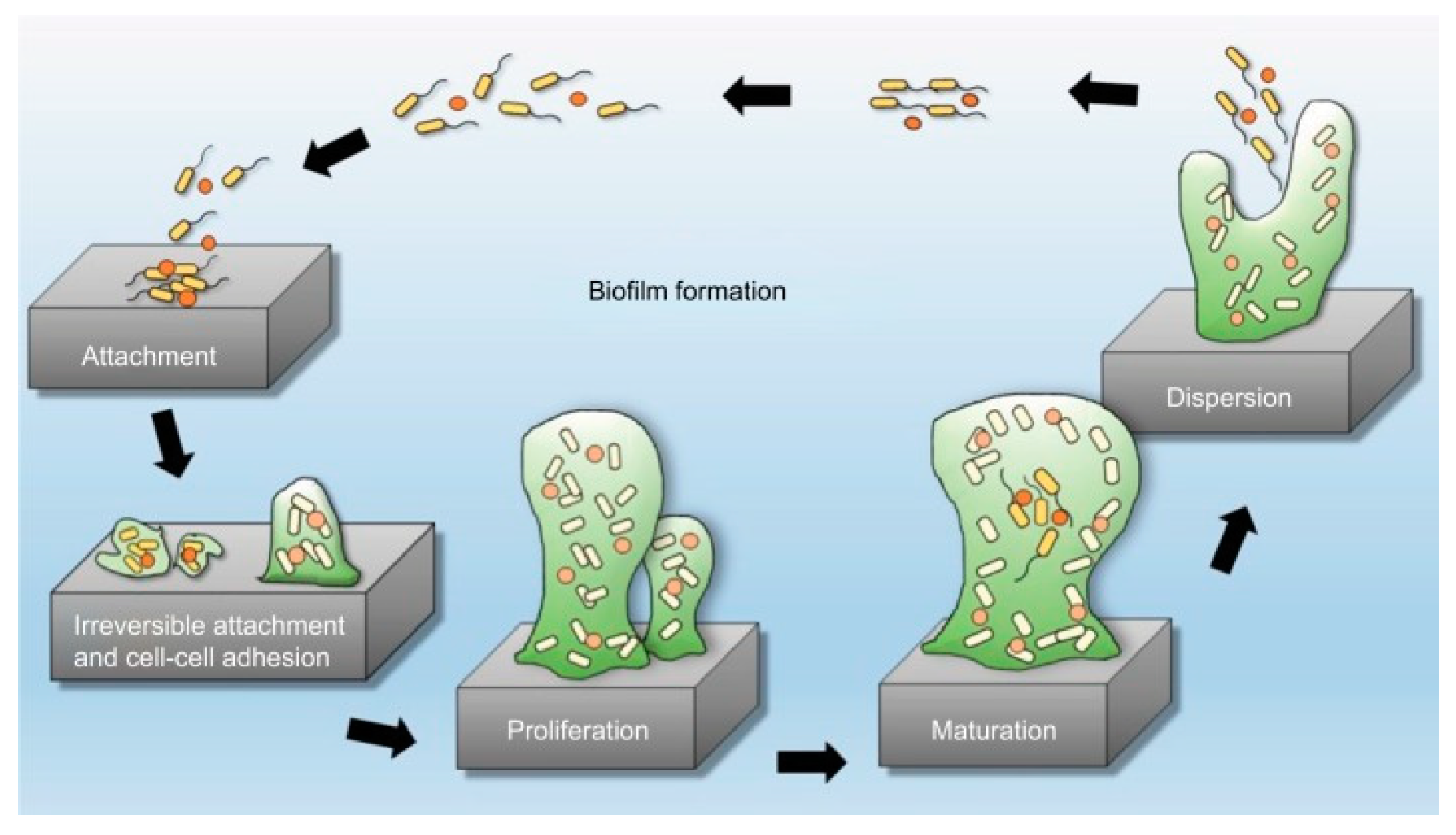
2. Biofilms Formation and Resistance
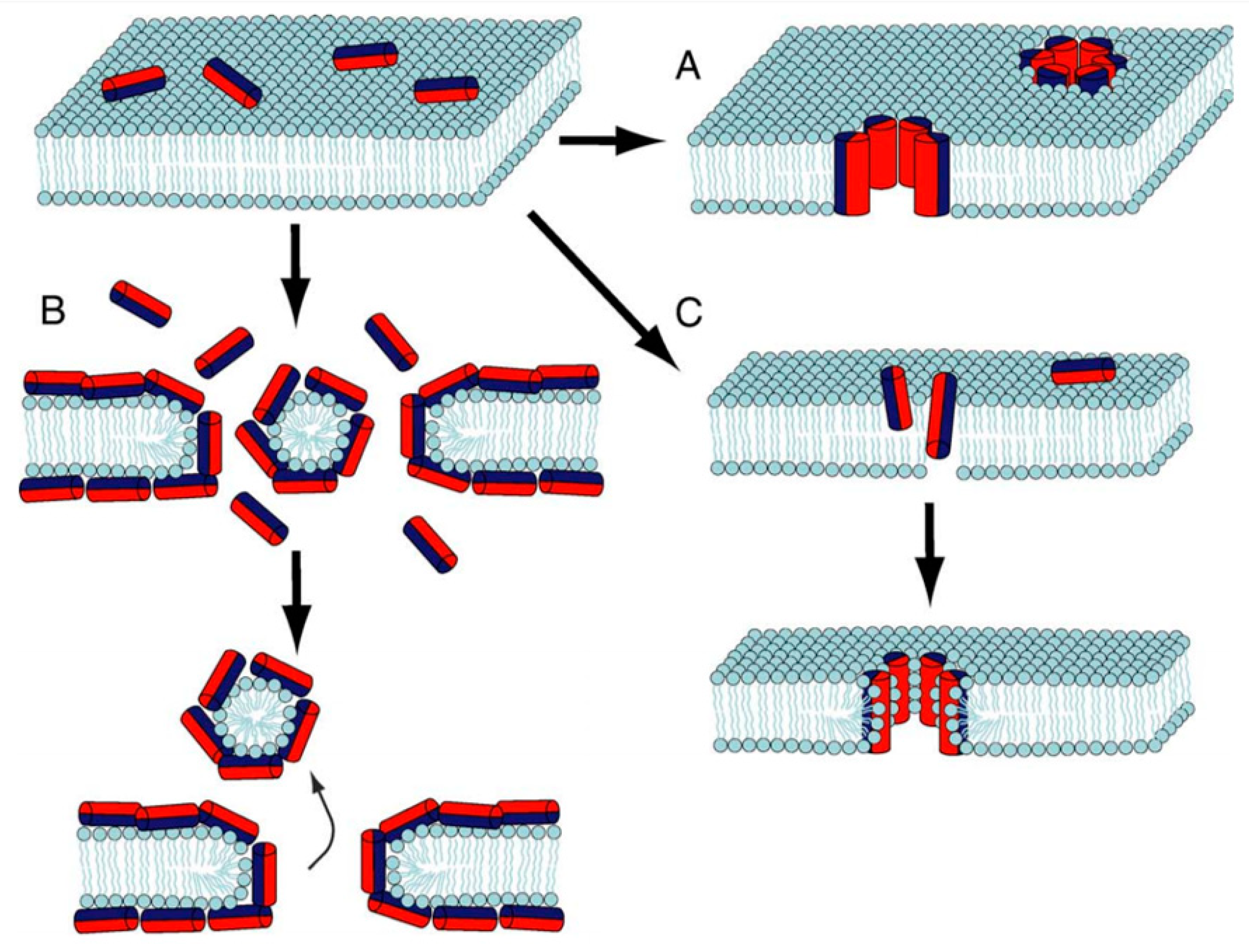
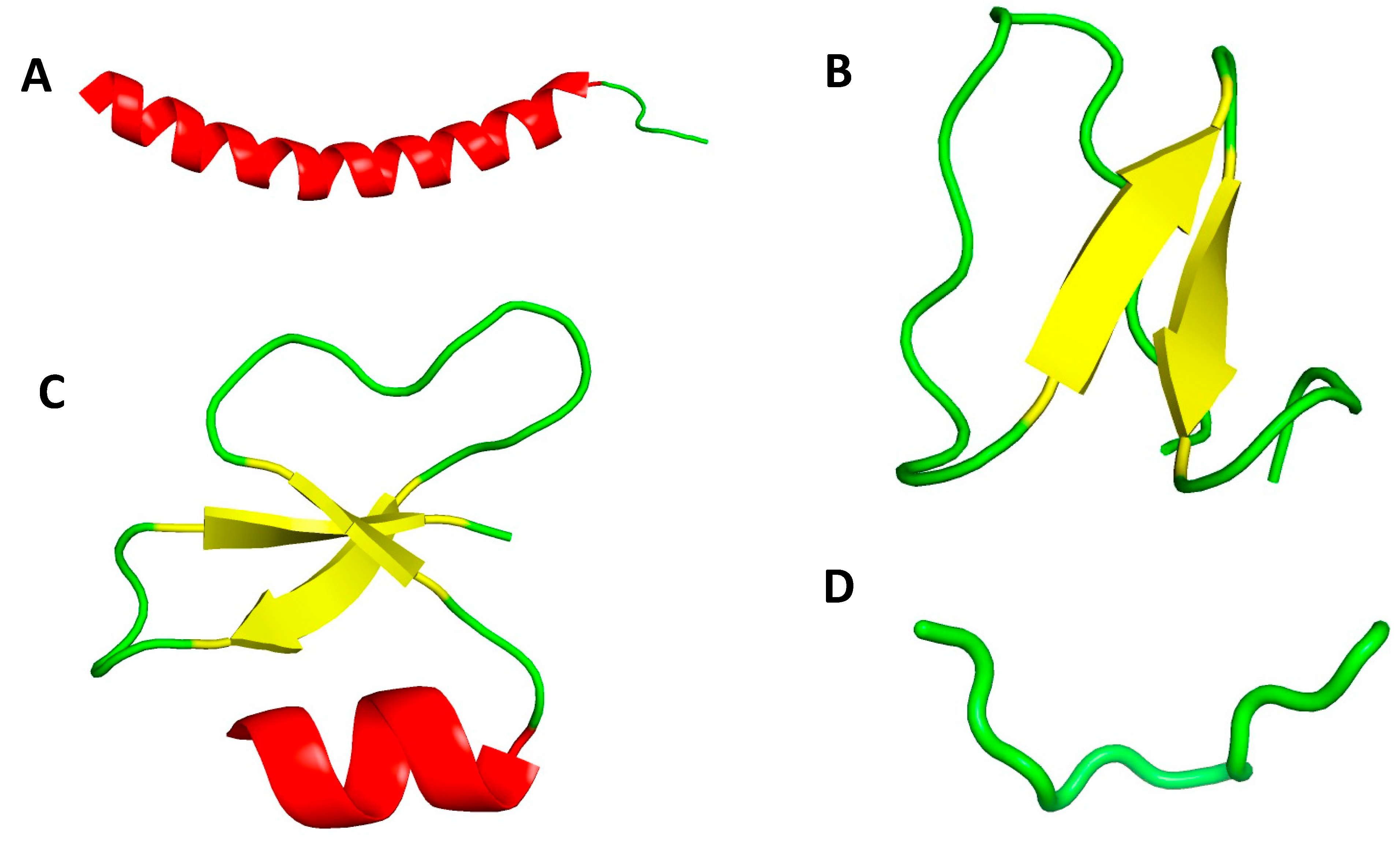
3. Antimicrobial Peptides (AMPs) Structure and Mechanism of Action
4. Strategies for AMP Immobilization on Surfaces

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AMP | Substratum | Immobilization Strategy | Studied Microorganisms | Reference |
|---|---|---|---|---|
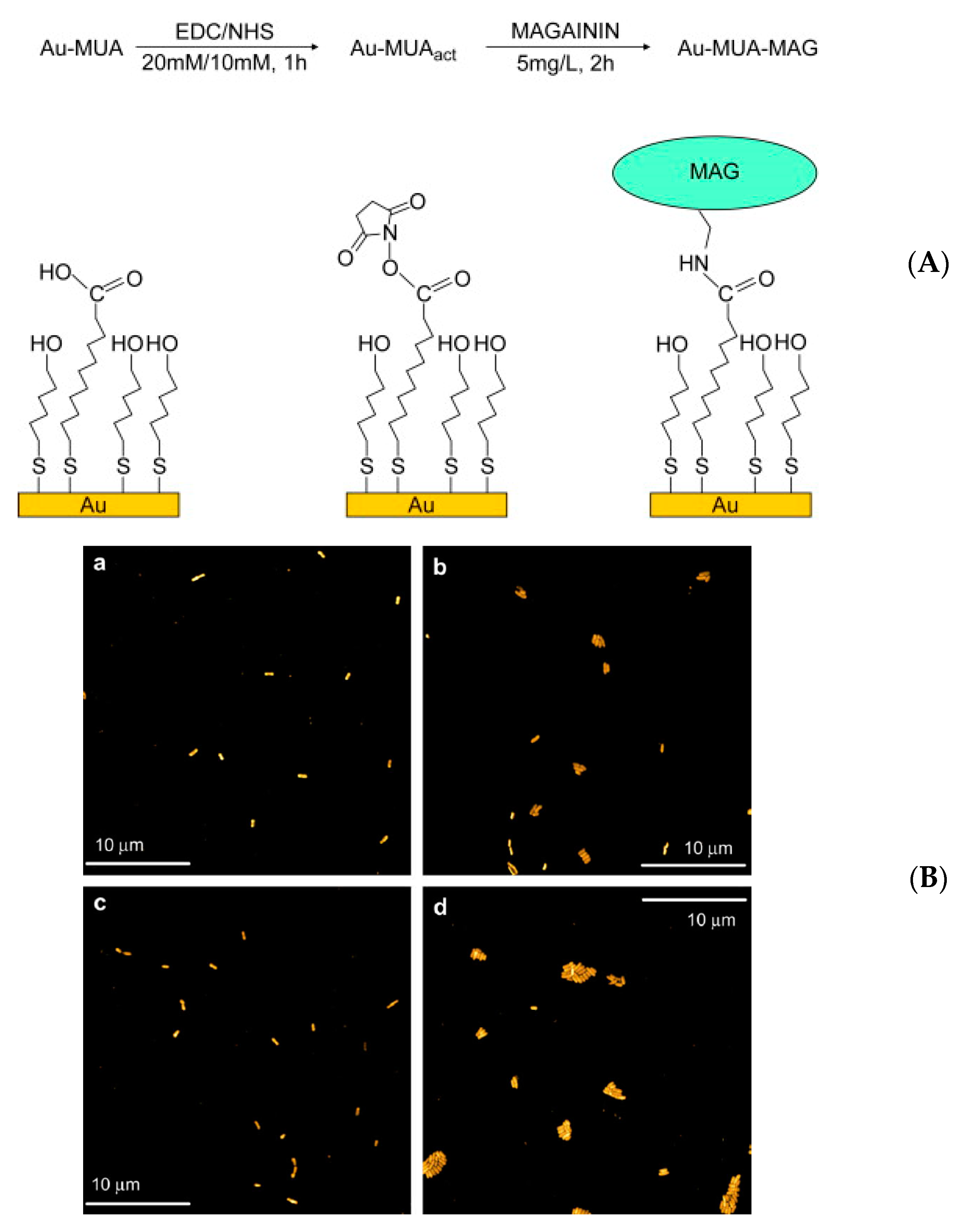
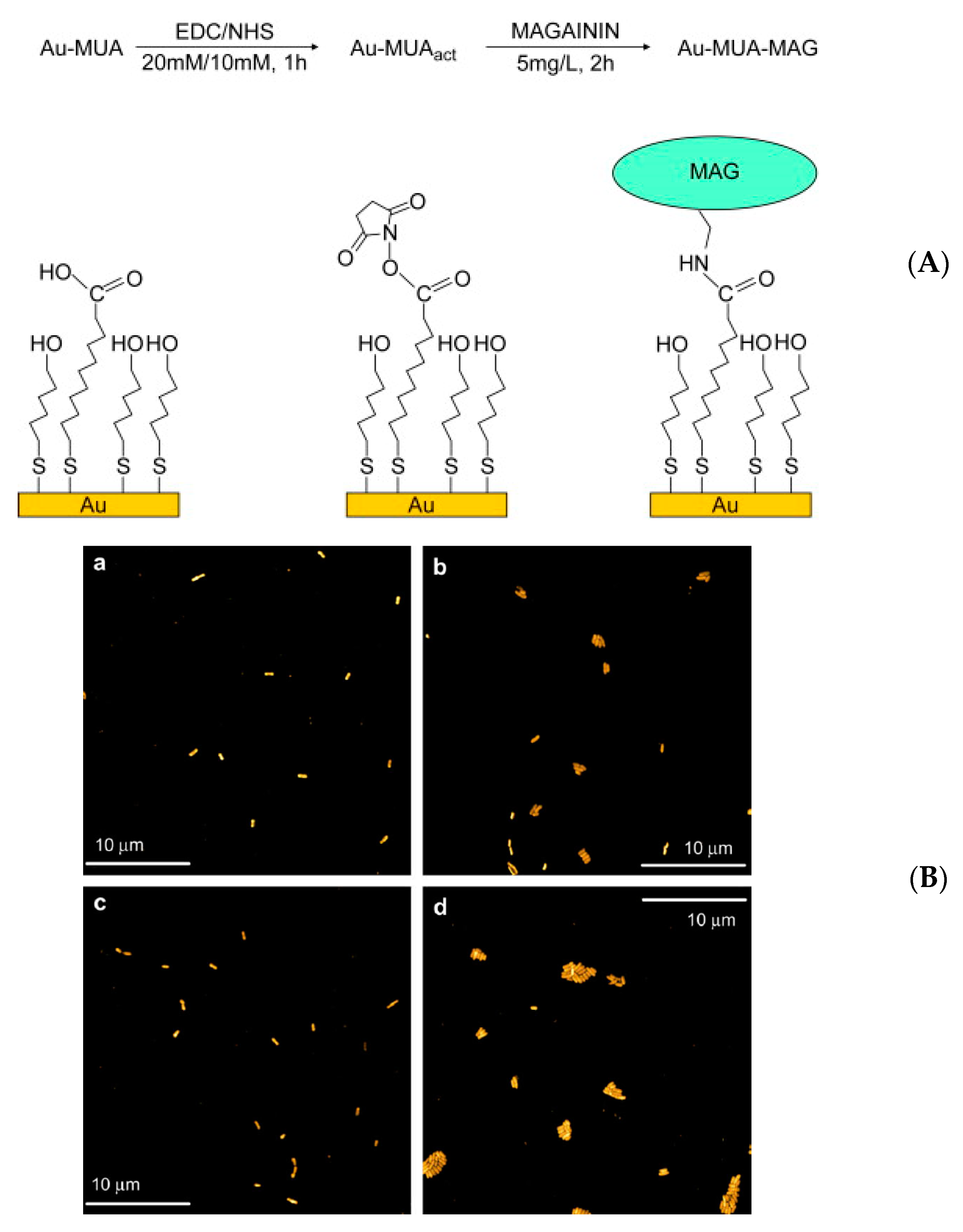
| Magainin I | Gold surface | Self-assembled monolayer | L. ivanovii, E. faecalis, S. aureus | Humblot et al., (2009) [71] |
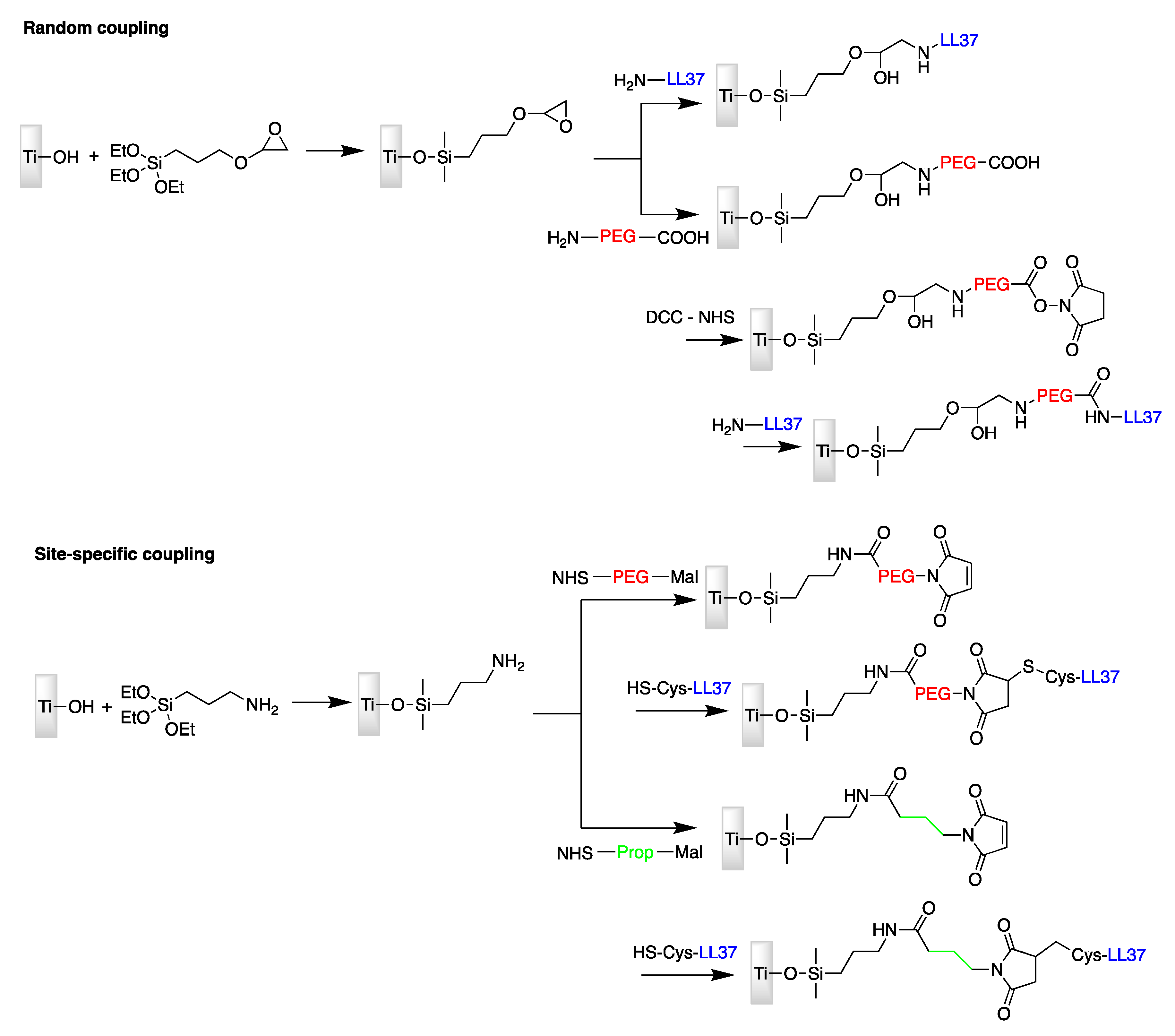
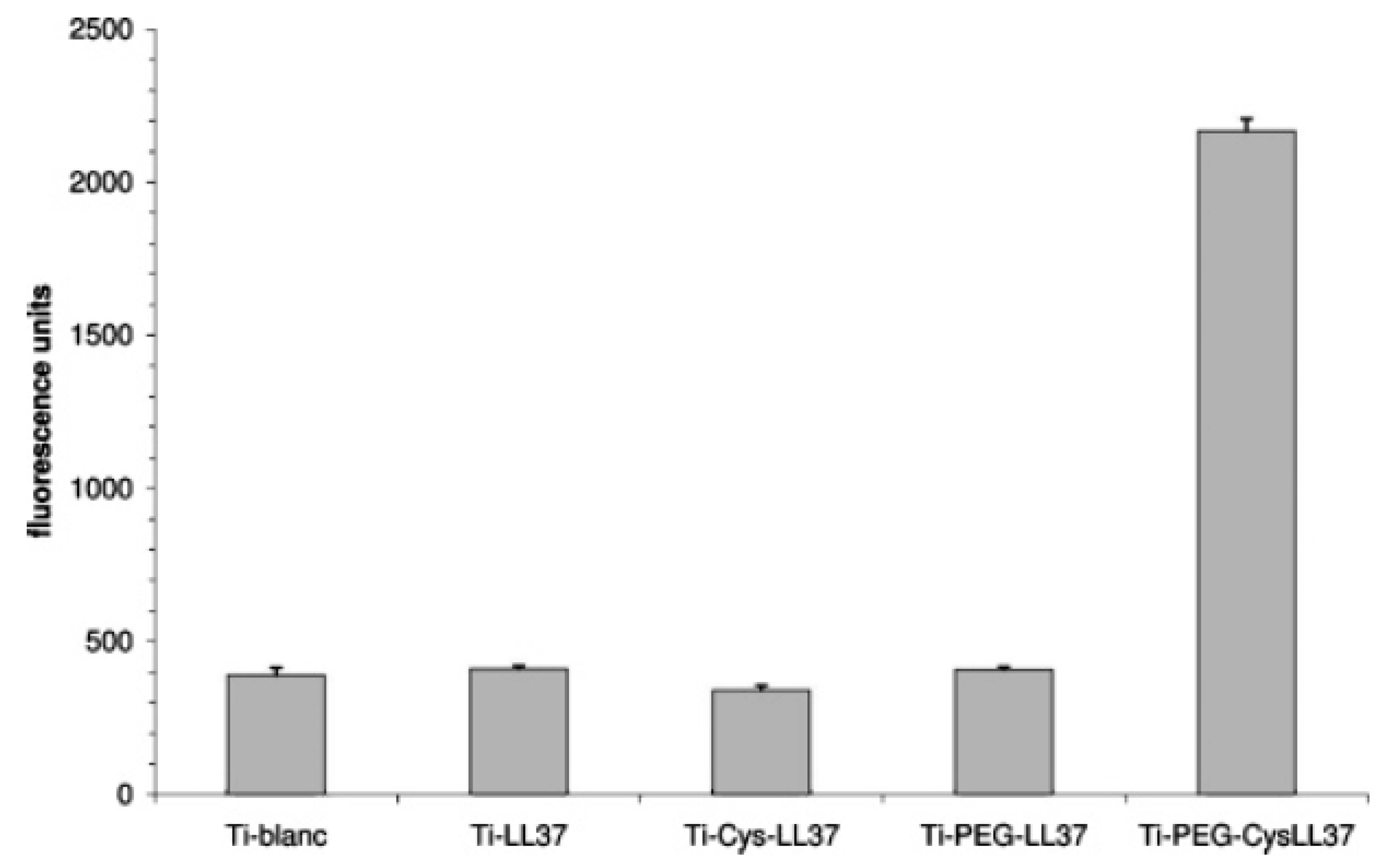
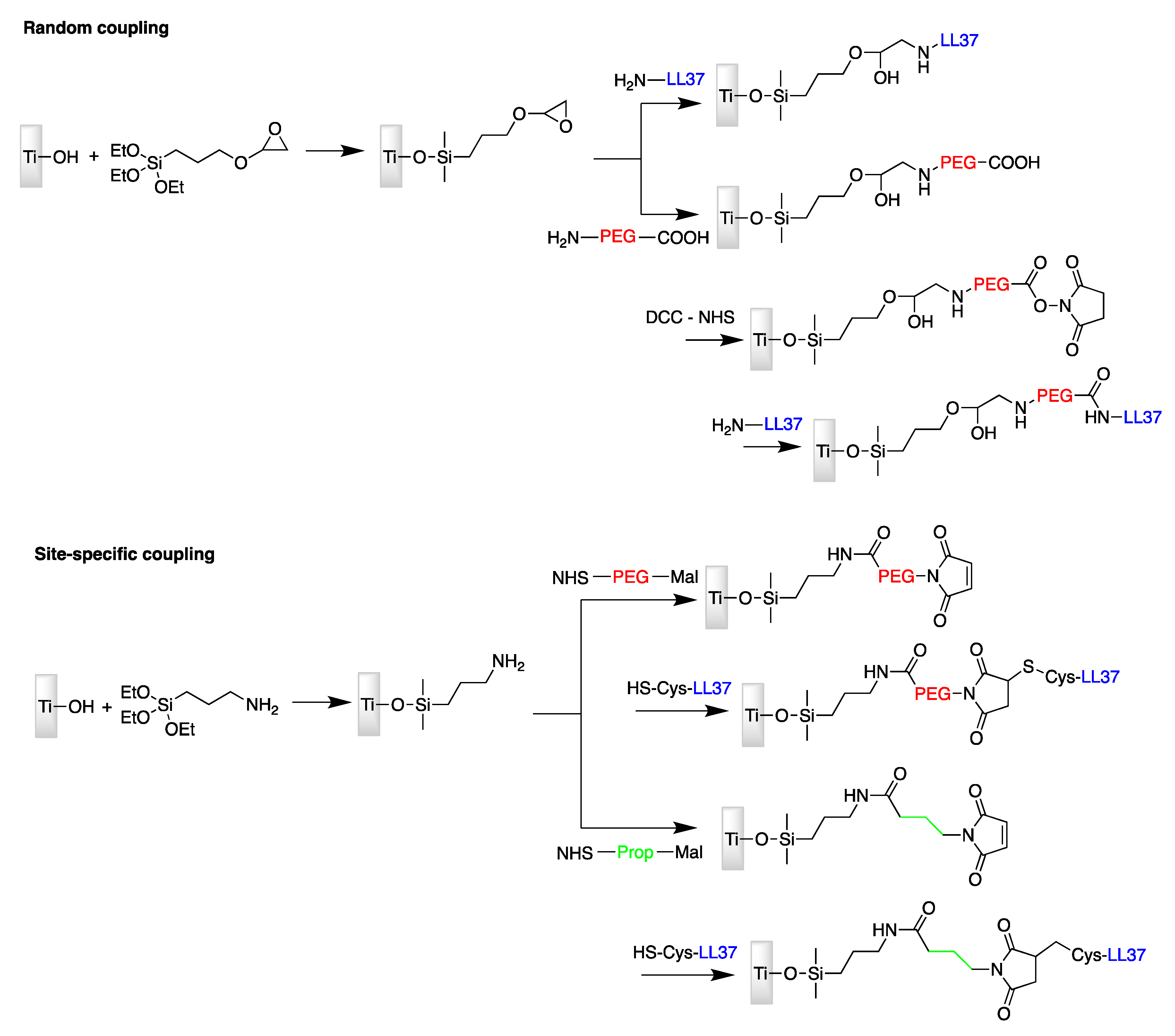
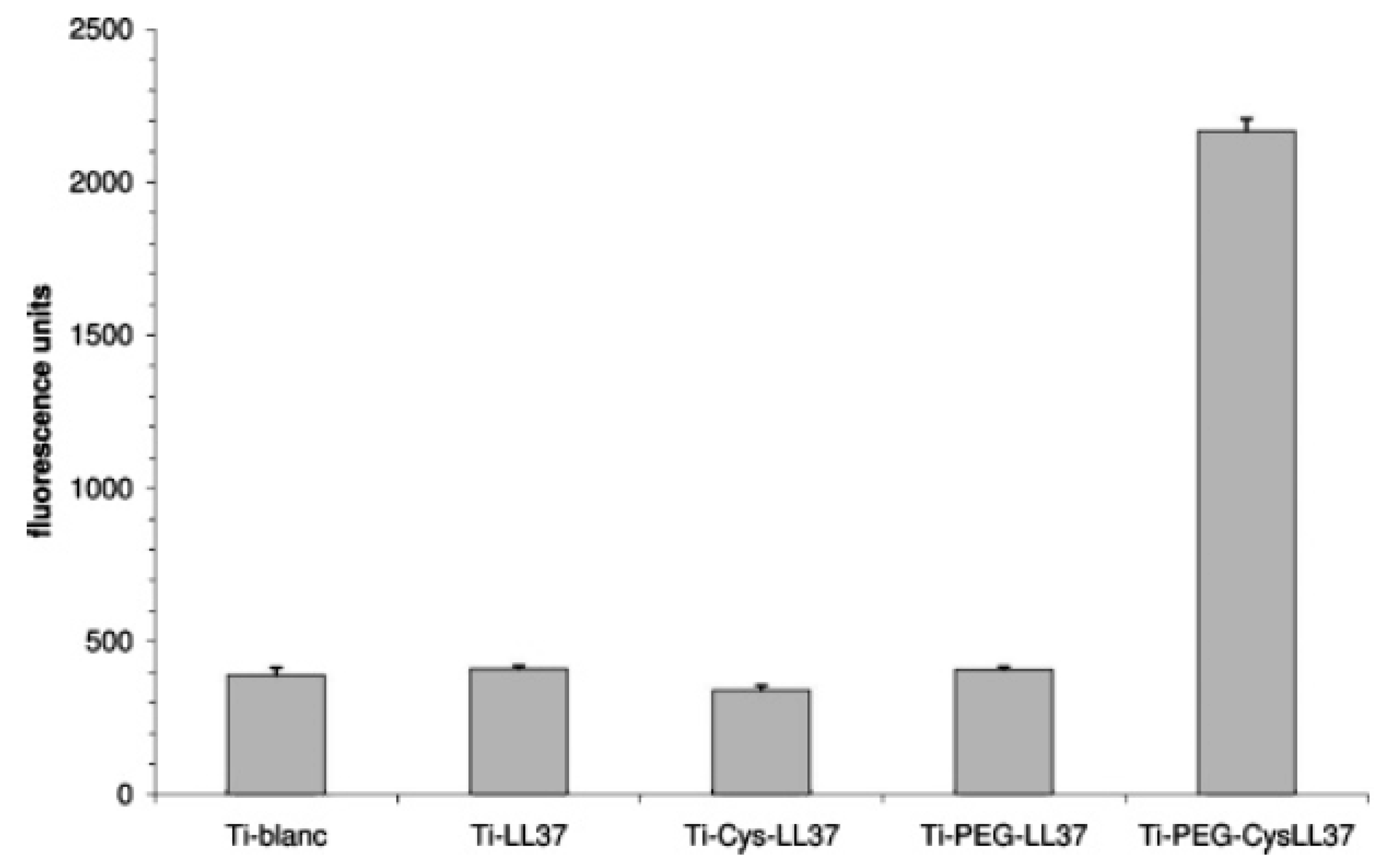
| LL37 | Titanium surface | Site-specific conjugation via amine-reactive NHS and thiol-reactive maleimide moieties | E. coli | Gabriel et al., (2006) [72] |
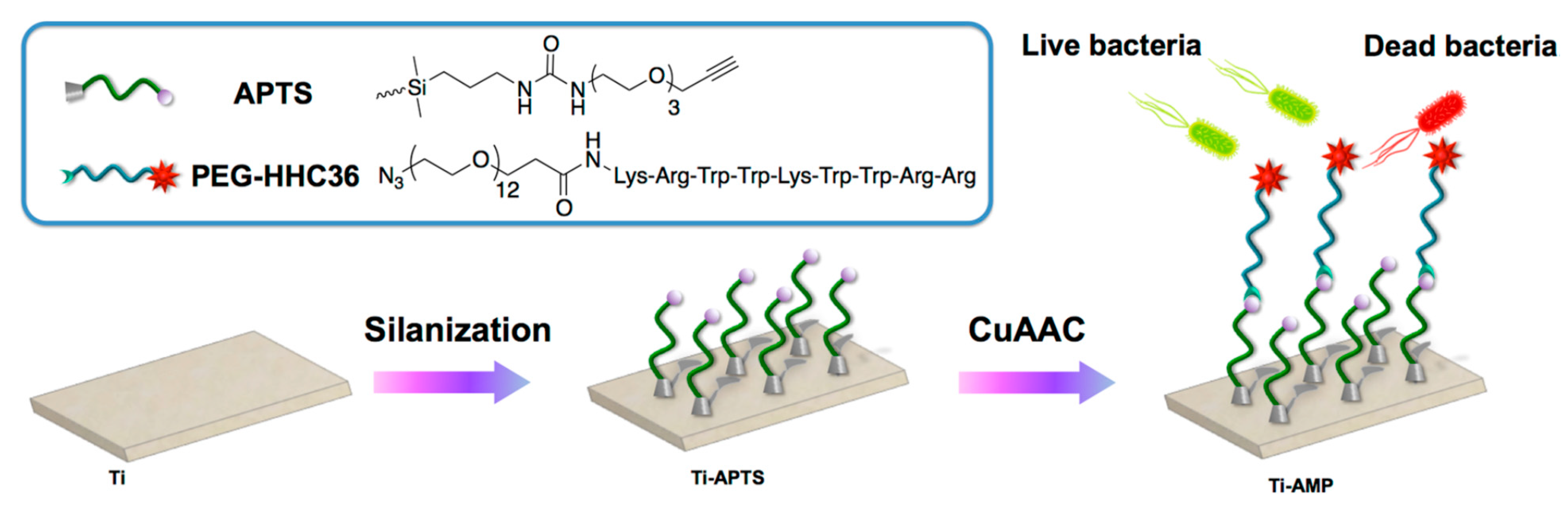
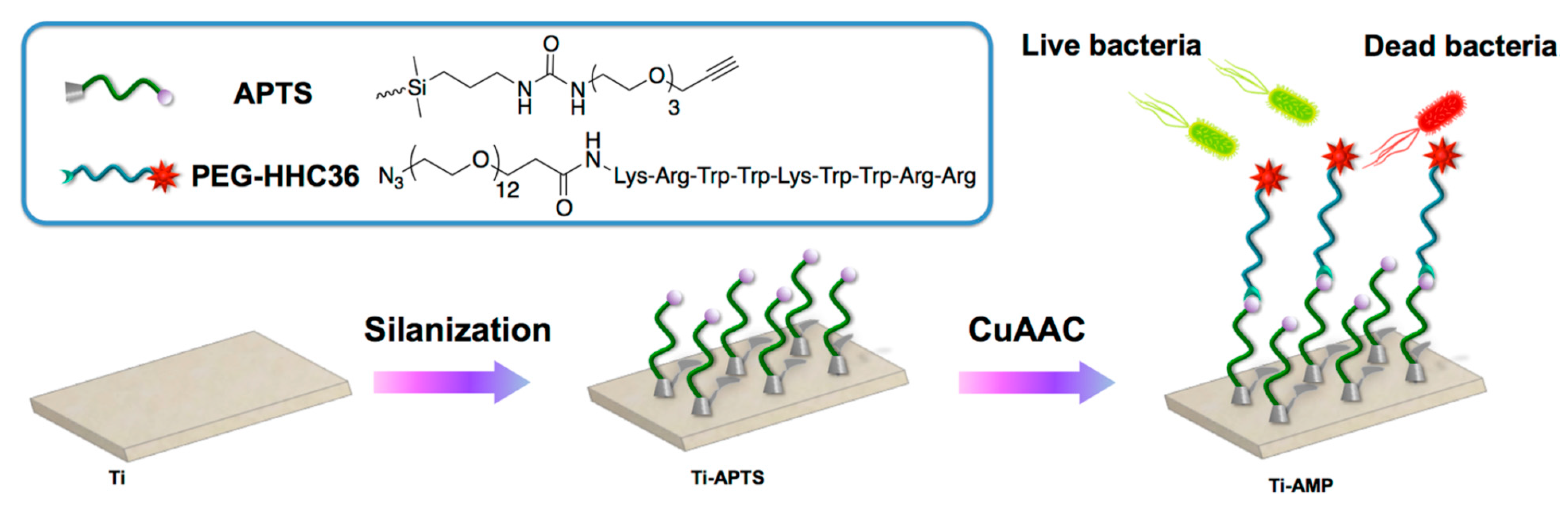
| HHC36 | Titanium surface | CuAAC click chemistry: Titanium silanized with APTS and peptide modified with PEG | S. aureus, E. coli | Chen et al., (2019) [73] |
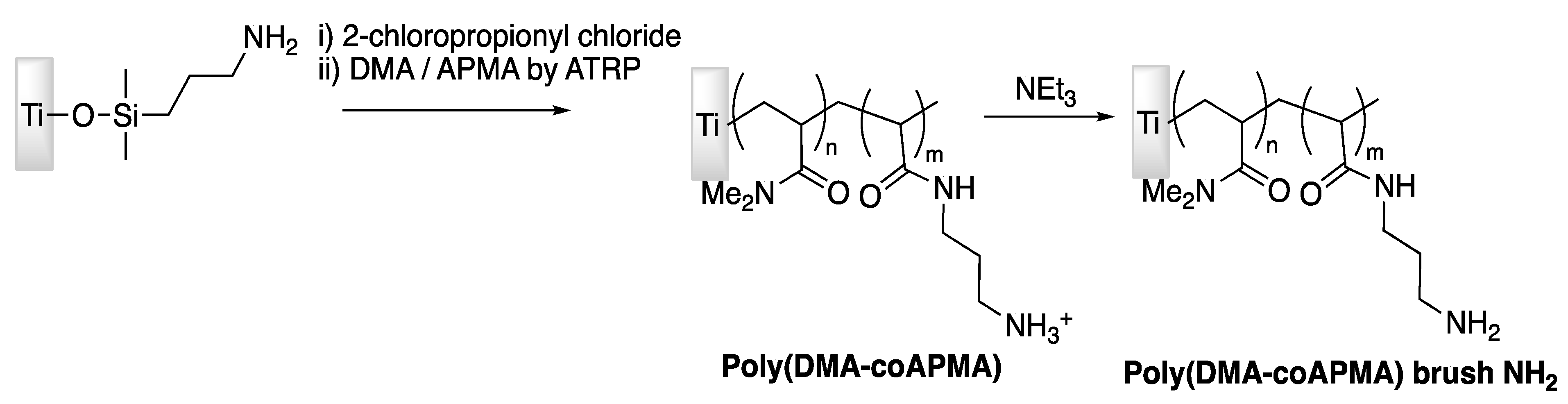
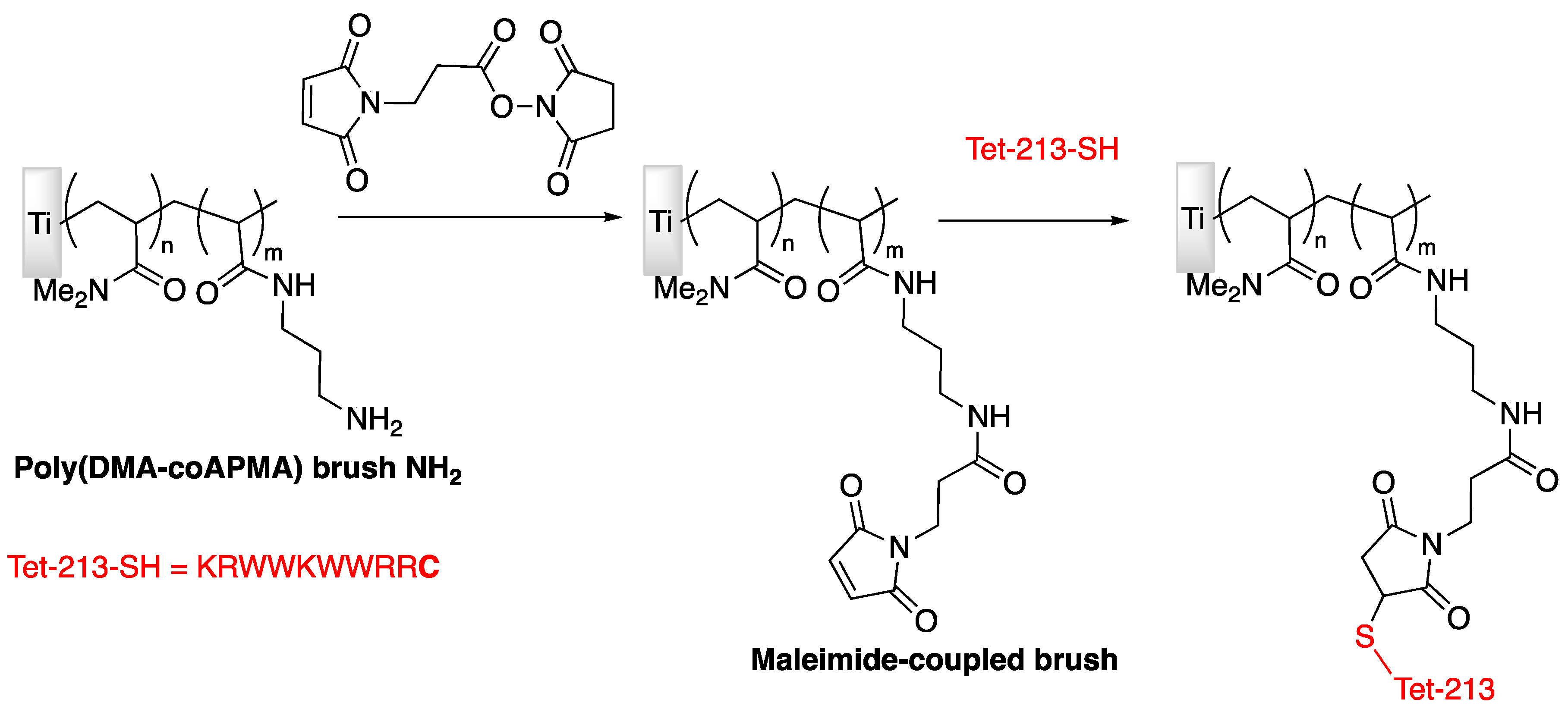
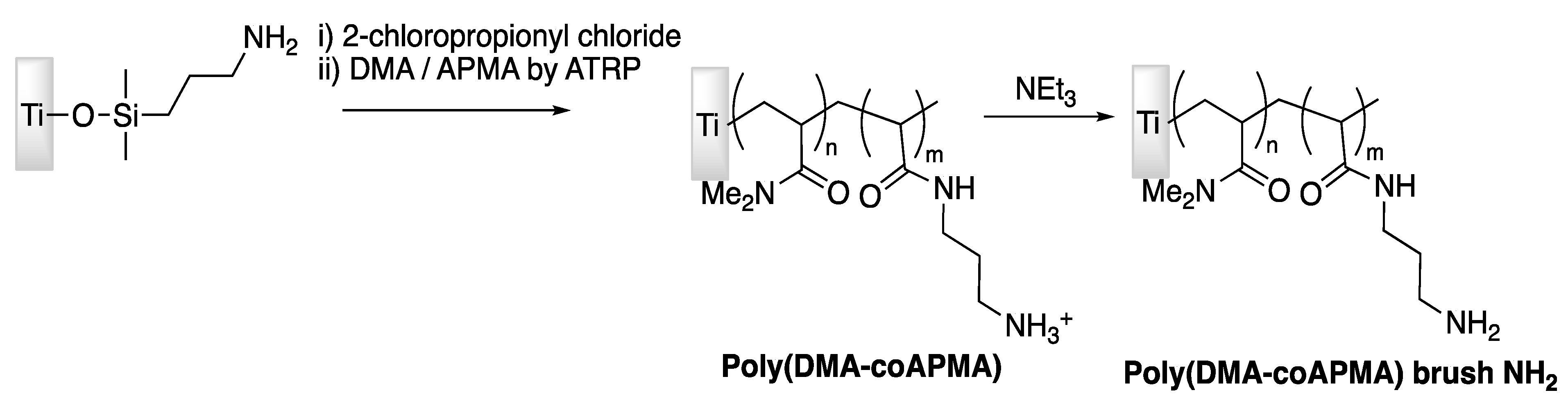
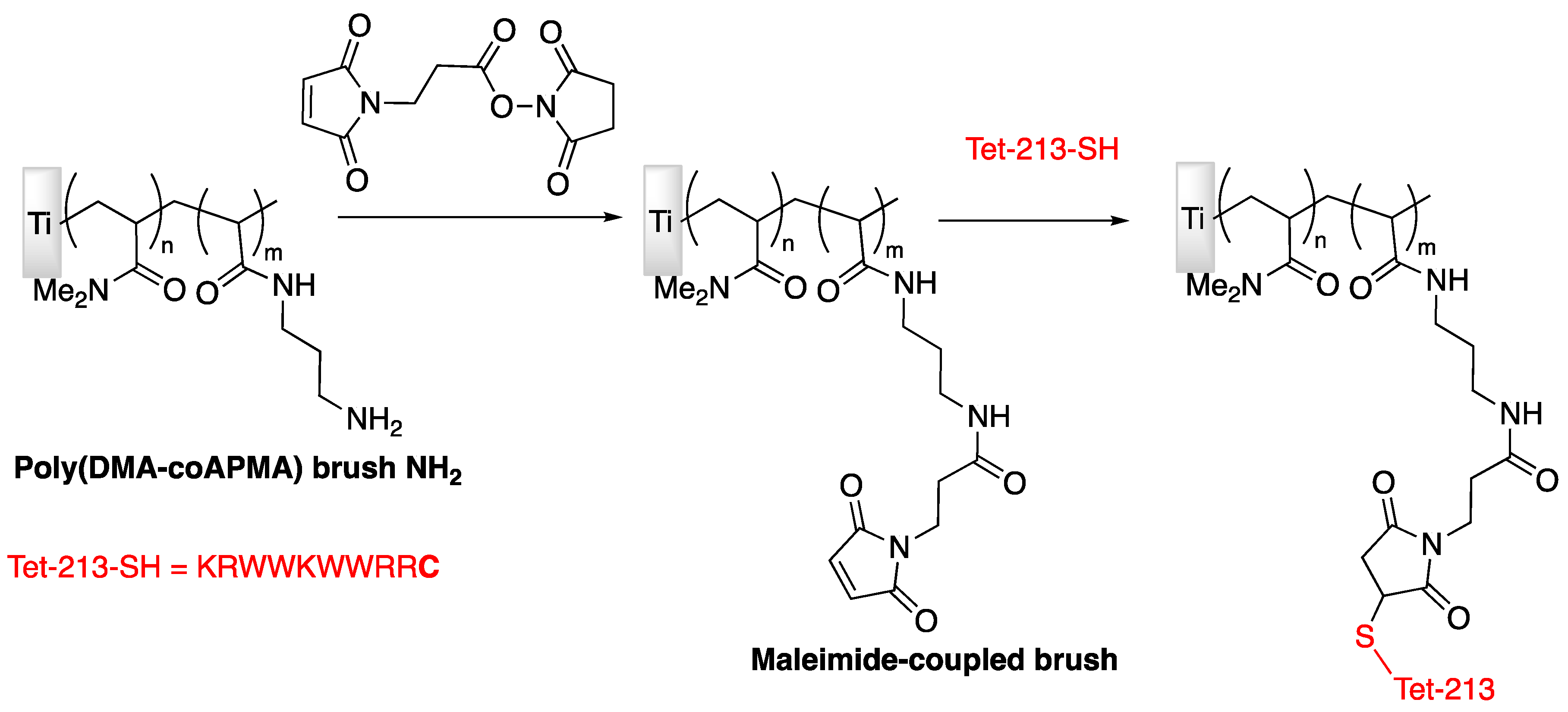
| Tet-213 | Titanium slide | Tethering on copolymer brushes of N,N-dimethylacrylamide/N-(3-aminopropyl)-methacrylamide hydrochloride | P. aeruginosa | Gao et al., (2011) [74] |
| Tet-213, 1010cys, Tet-20, Tet-21, Tet-26, HH2, MXX226 | Titanium slide | Covalently grafted hydrophilic polymer brushes conjugated with peptides | S. aureus, P. aeruginosa | Gao et al., (2011) [75] |
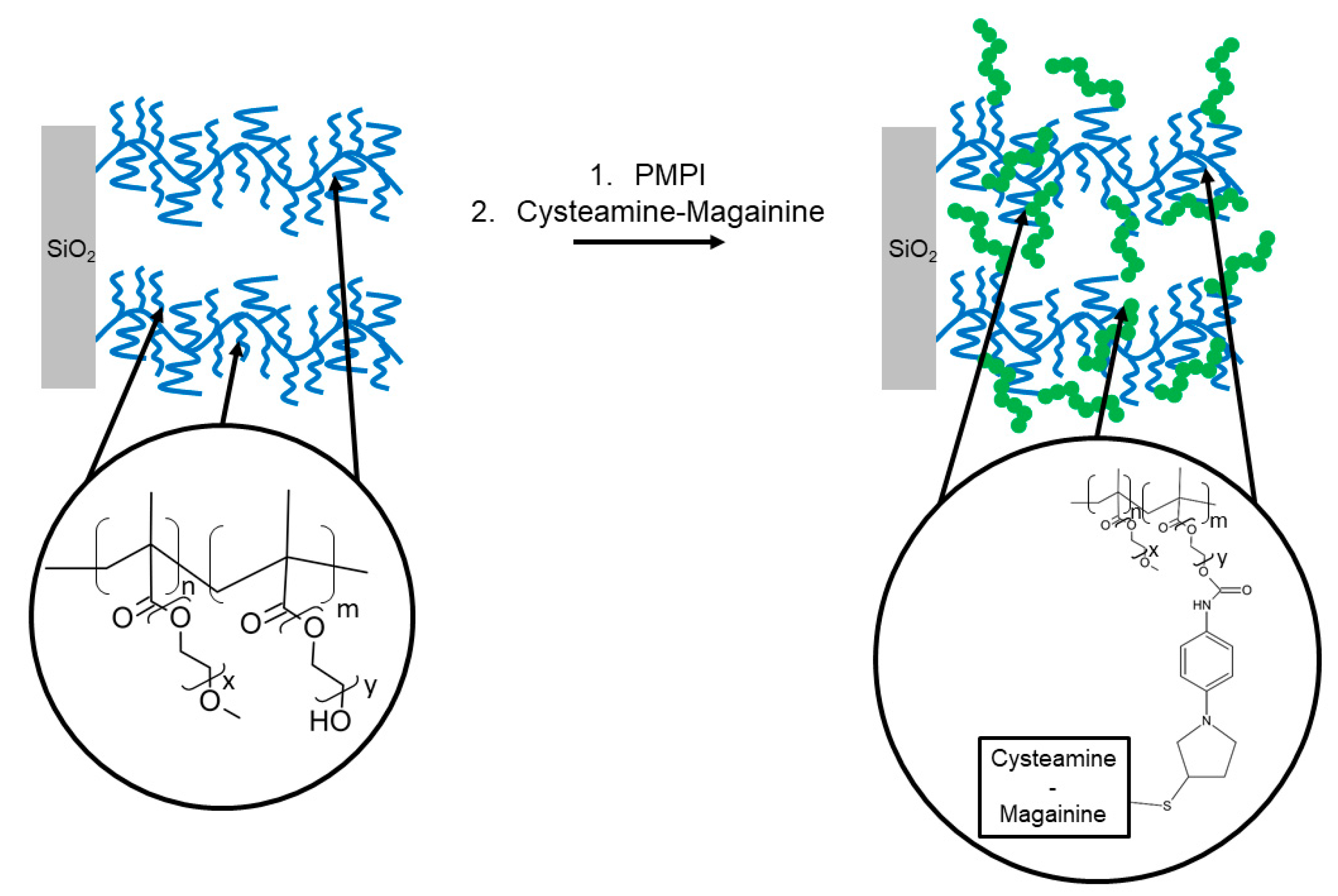
| Magainin I | Silicon wafer | Tethering on copolymer brushes of 2-(2 methoxyethoxy) ethylmethacrylate/hydroxyl-terminated oligo (ethylene glycol) Poly(MOE2MA-co-HOEGMA) | L. ivanovii, B. cereus | Glinel et al., (2009) [76] |
| CW11 | Polydimethylsiloxane (PDMS) | Cross-linking of peptides to allylglycidyl ether modified PDMS surface (PDMS-AGE-PEG) via Sulfhydryl Chemistry | E. Coli, S. aureus, P. aeruginosa | Lim et al., (2013) [77] |
| RK1 and RK2 | Silicone urinary catheter and Polydimethylsiloxane (PDMS) | Cross-linking of peptides to allylglycidyl ether modified PDMS surface (PDMS-AGE) | E. Coli, S. aureus, C. albicans | Li et al., (2014) [78] |
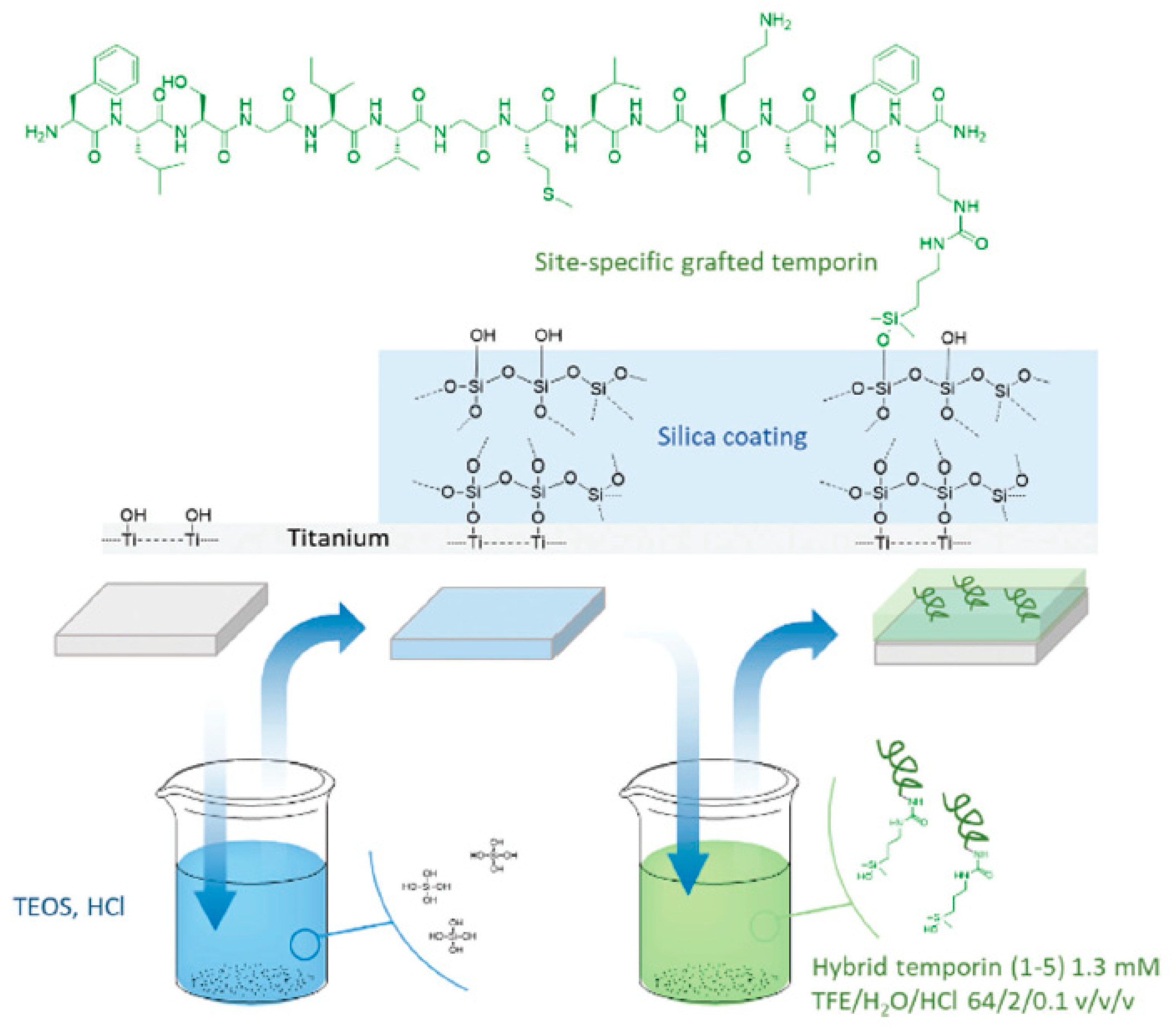
| Temporin-SHa | Titanium surface | Covalent immobilization on Si-O-Ti groups | S. epidermidis, E. coli | Masurier et al., (2018) [79] |
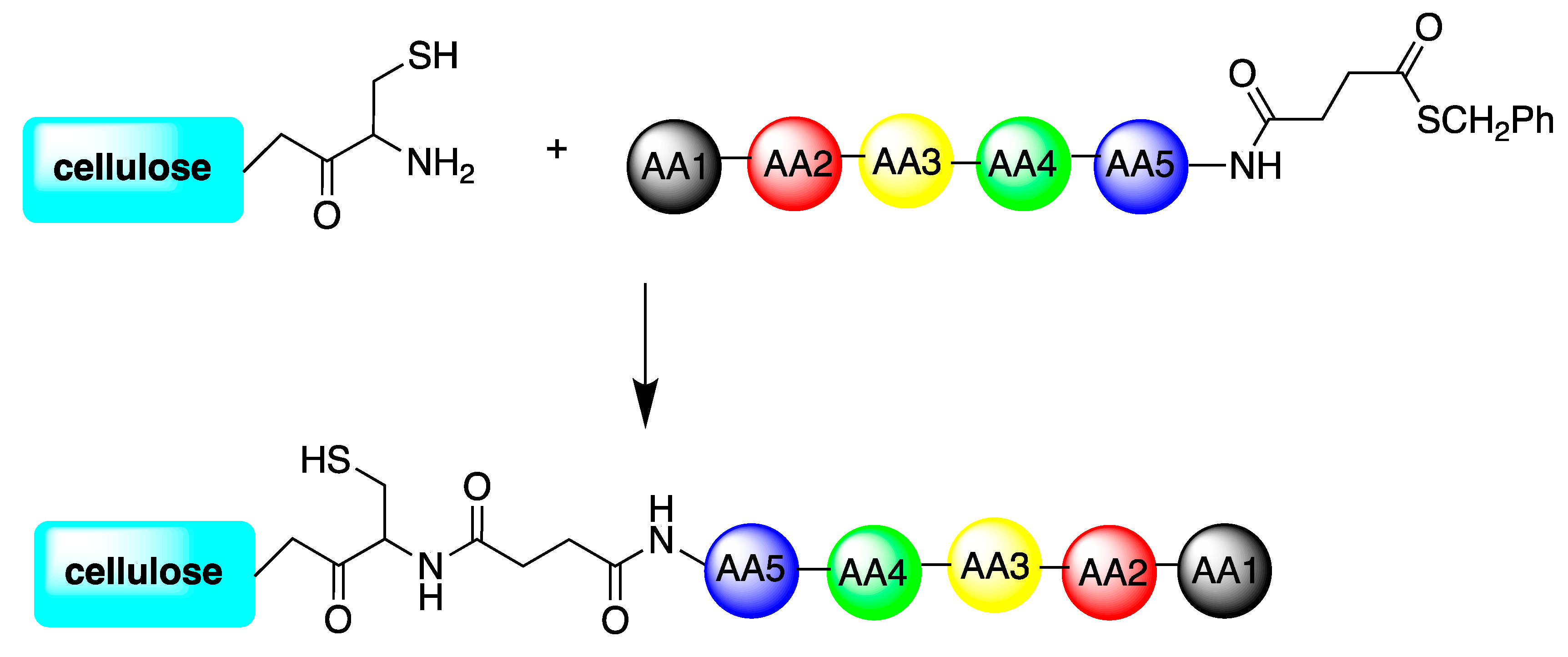
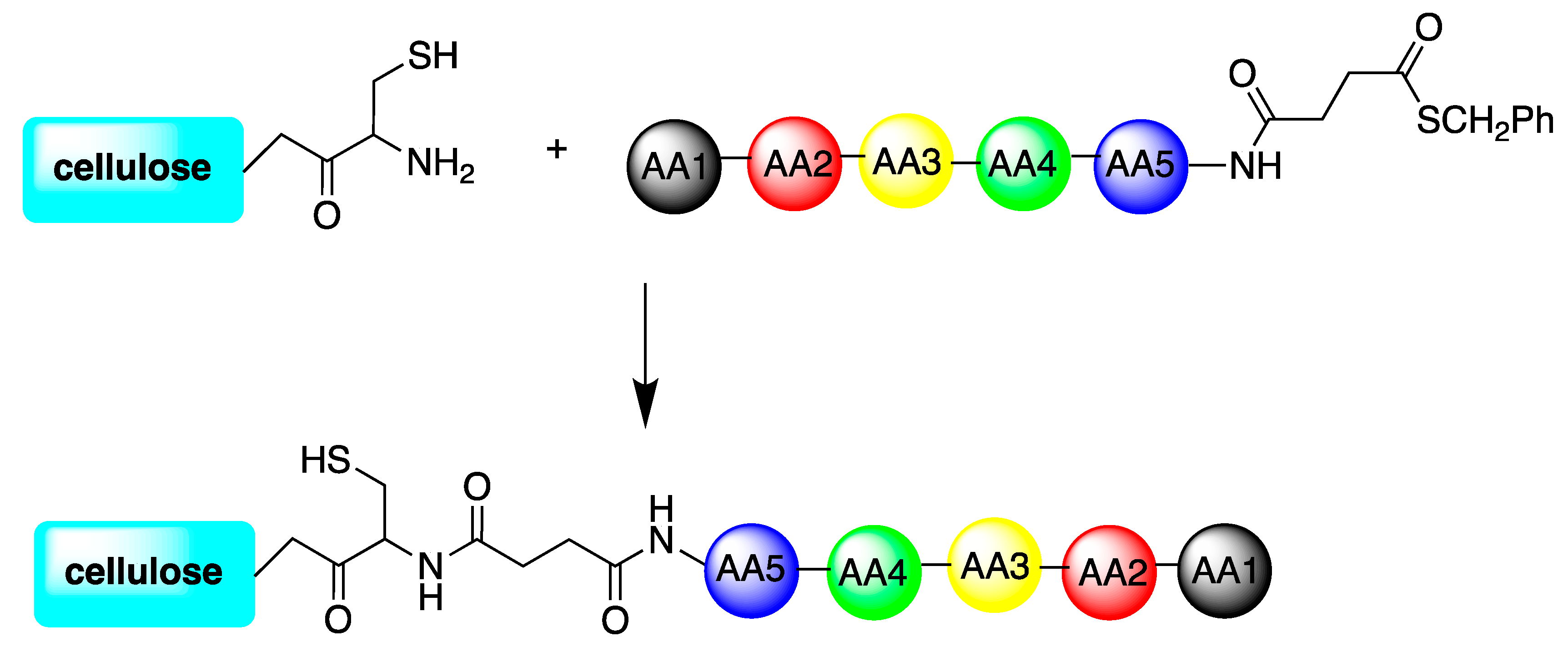
| 122 variant peptides of 2 starting sequences: Bac2A and Indolicidin | Cellulose support | Cellulose-amino-hydroxypropyl ether (CAPE) linker chemistry; or directly synthetized onto a bifunctional resin; or directly bound to the microtiter plate via biotin-streptavidin interaction | S. aureus, P. aeruginosa, C. albicans | Hilpert et al., (2009) [80] |
| TBKKG6A and lasioglossin-III | Cellulose support | cysteine-cellulose conjugate coupled to obtaining thioester peptides either on the C-terminal or the N-terminal part of the molecules | E. coli. | Sperandeo et al., (2020) [81] |
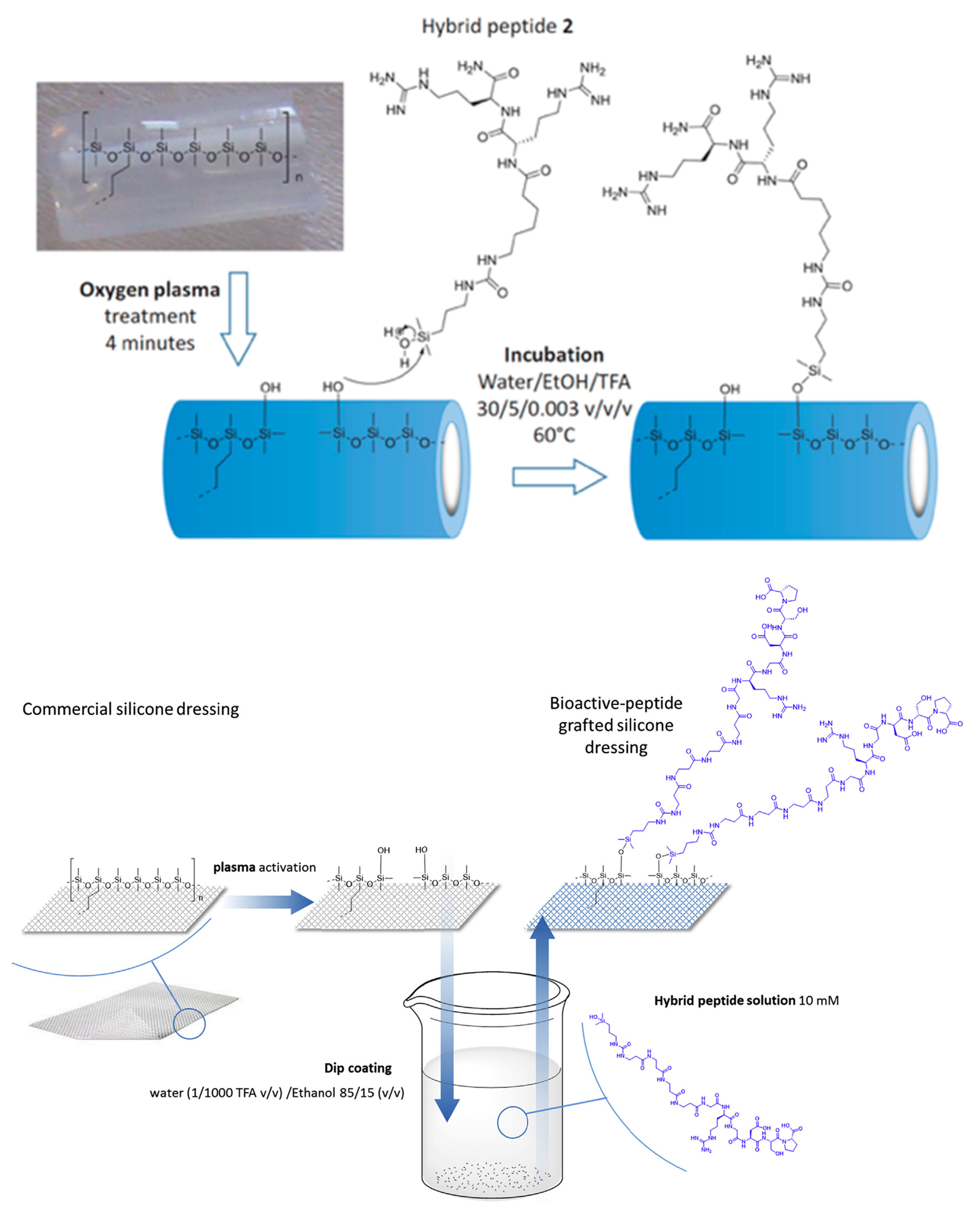
| Antibacterial hybrid peptide | Silicone catheter | Covalent immobilization on silanol groups | E. Coli, P. aeruginosa, S. aureus | Pinese et al., (2016) [82] |
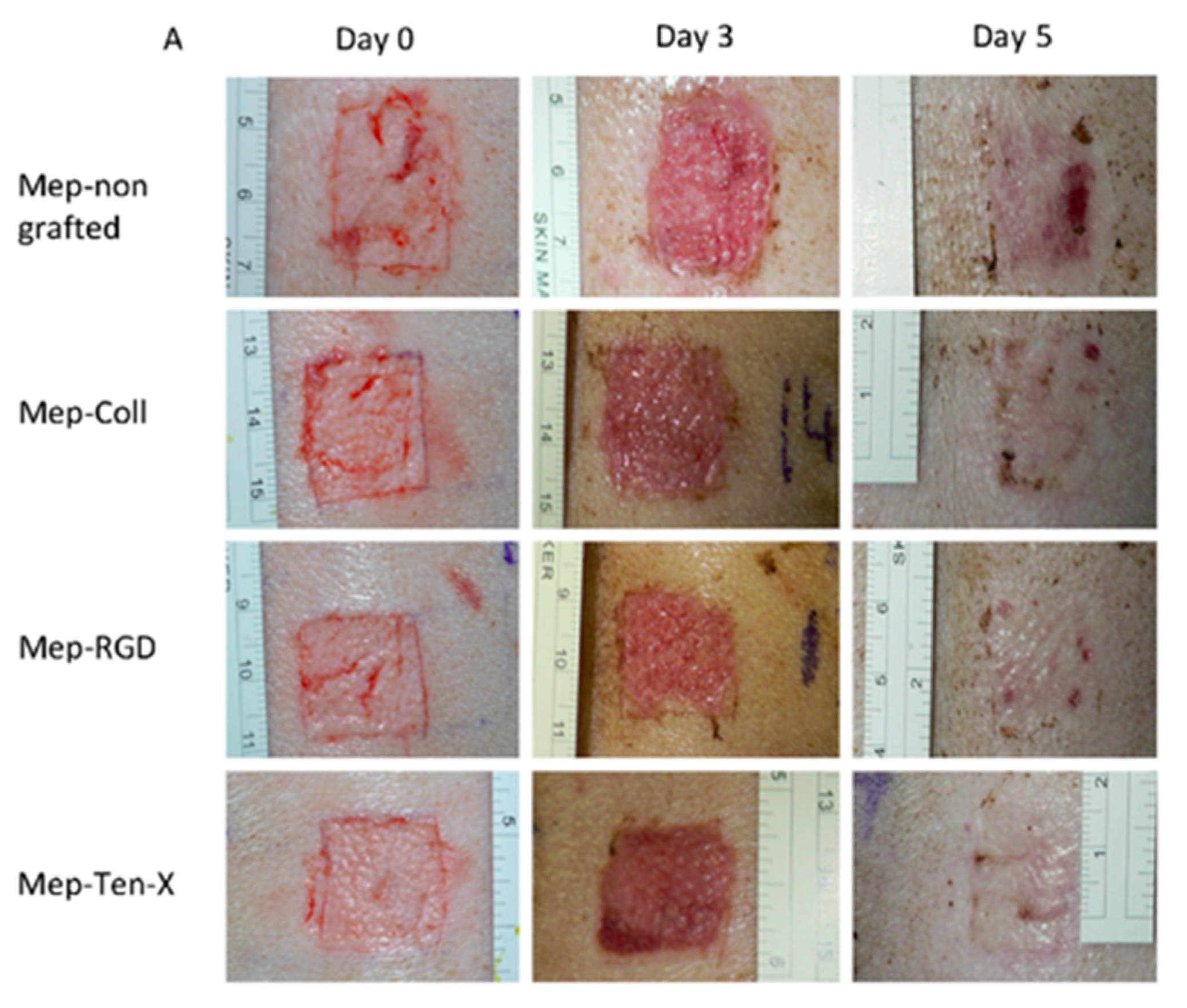
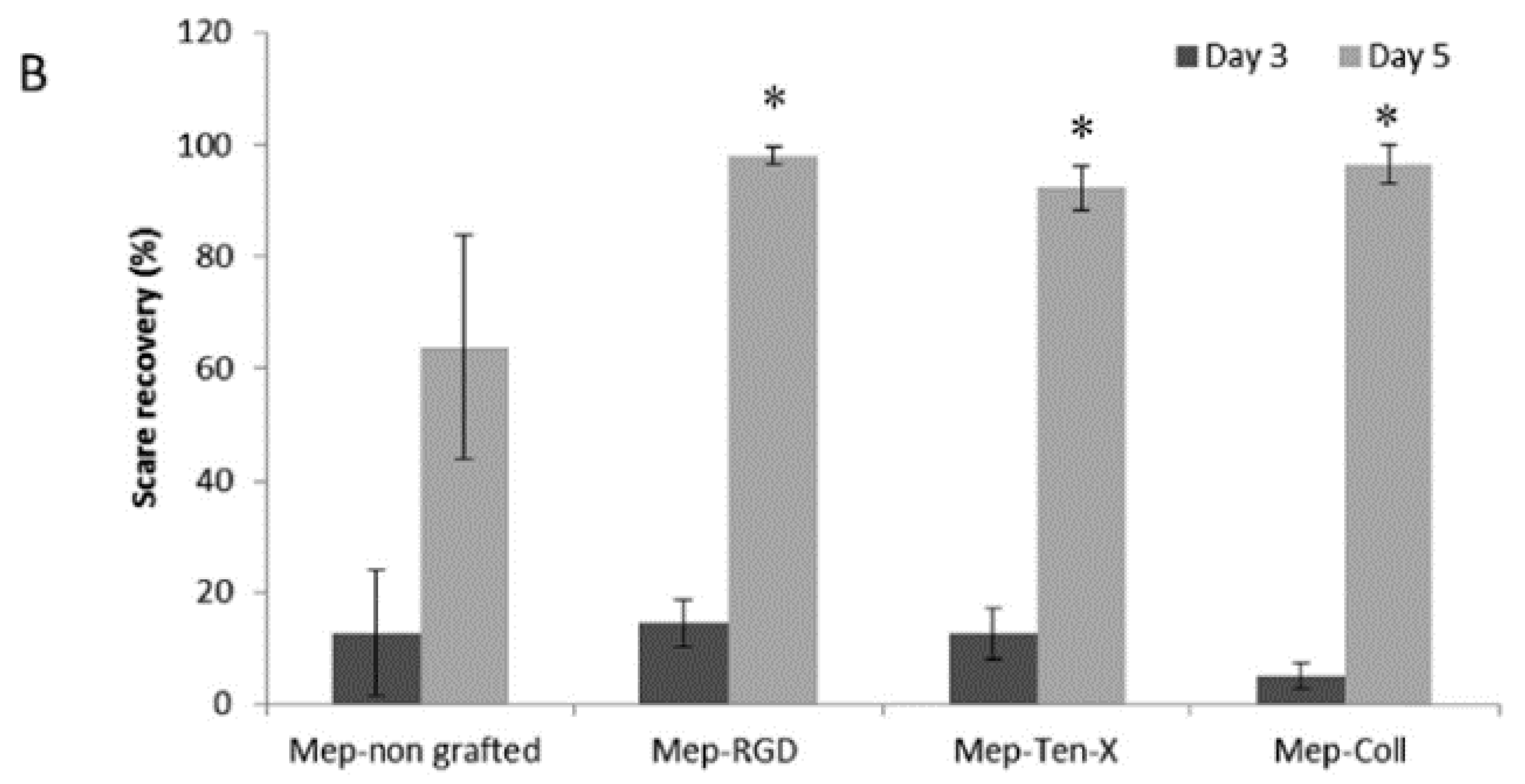
| Bioactive peptides | Silicone dressing | Covalent immobilization on silanol groups | Studies of peptides bioactivity | Pinese et al., (2017) [83] |
| Protamine, a mixture of Protamine and Melittin and Melimine | Commercial contact lens (Etafilcon-A) | Physical adsorption and covalent attachment via EDC | P. aeruginosa, S. aureus | Willcox et al., (2008) [84] |
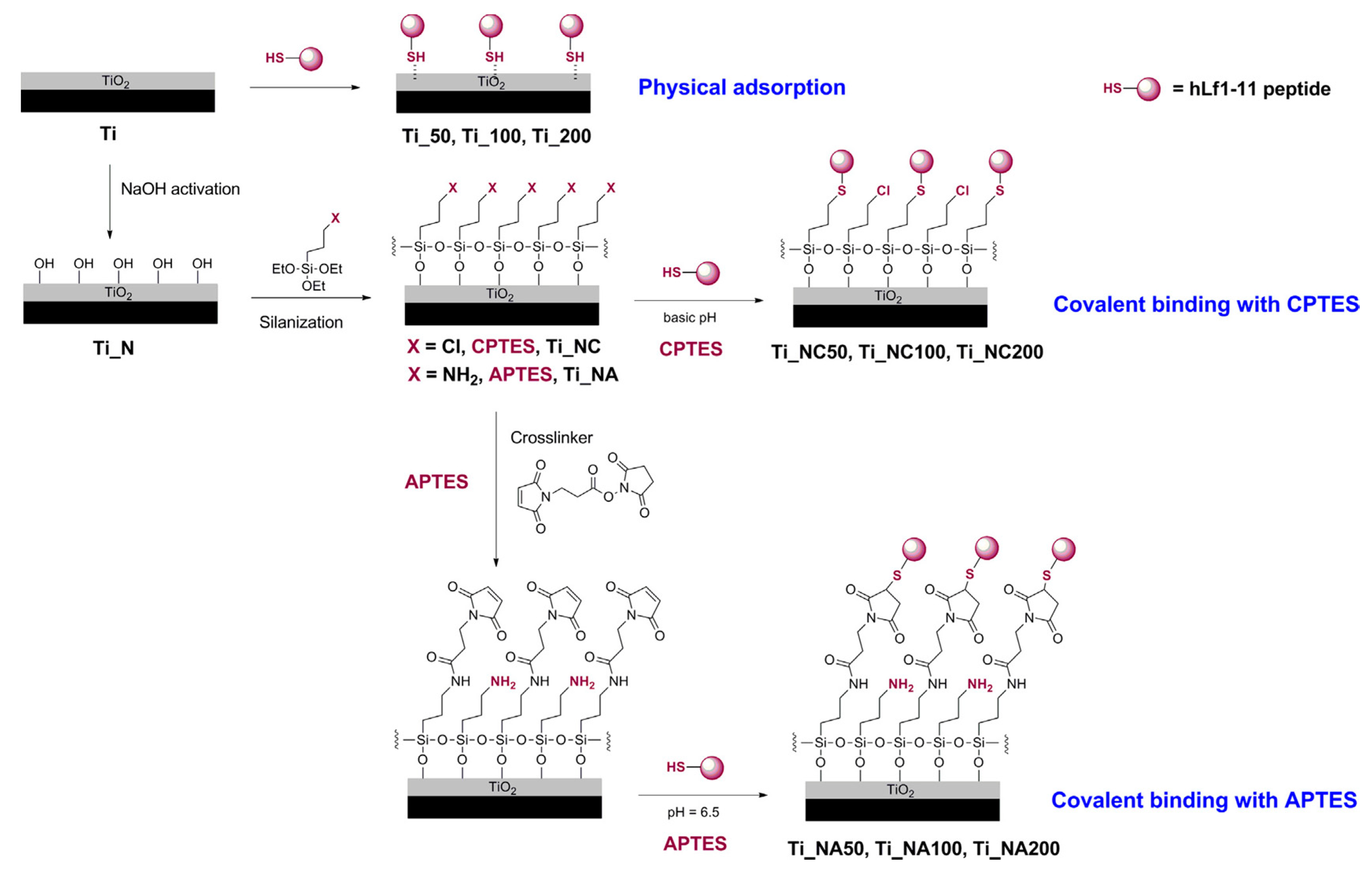
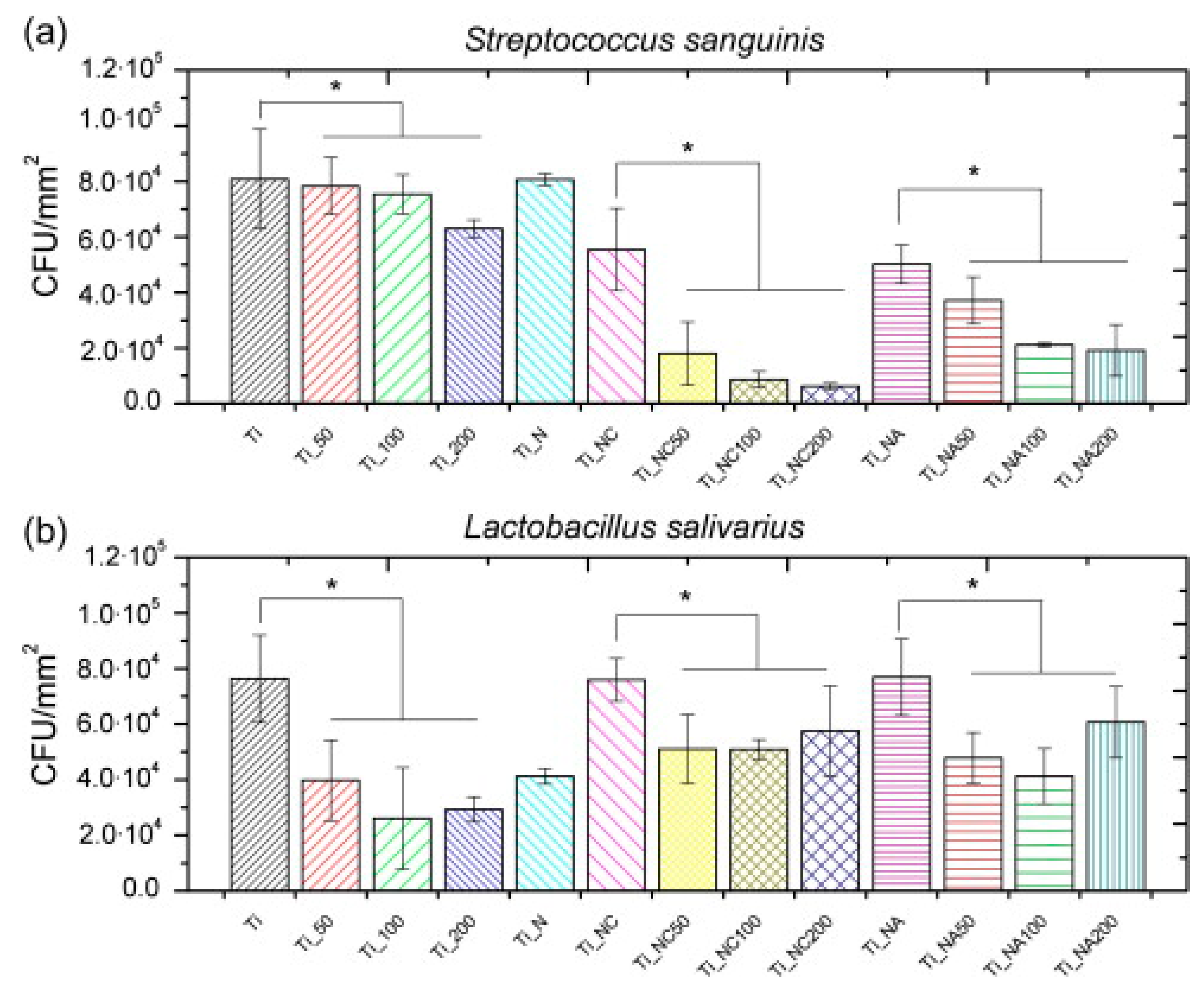
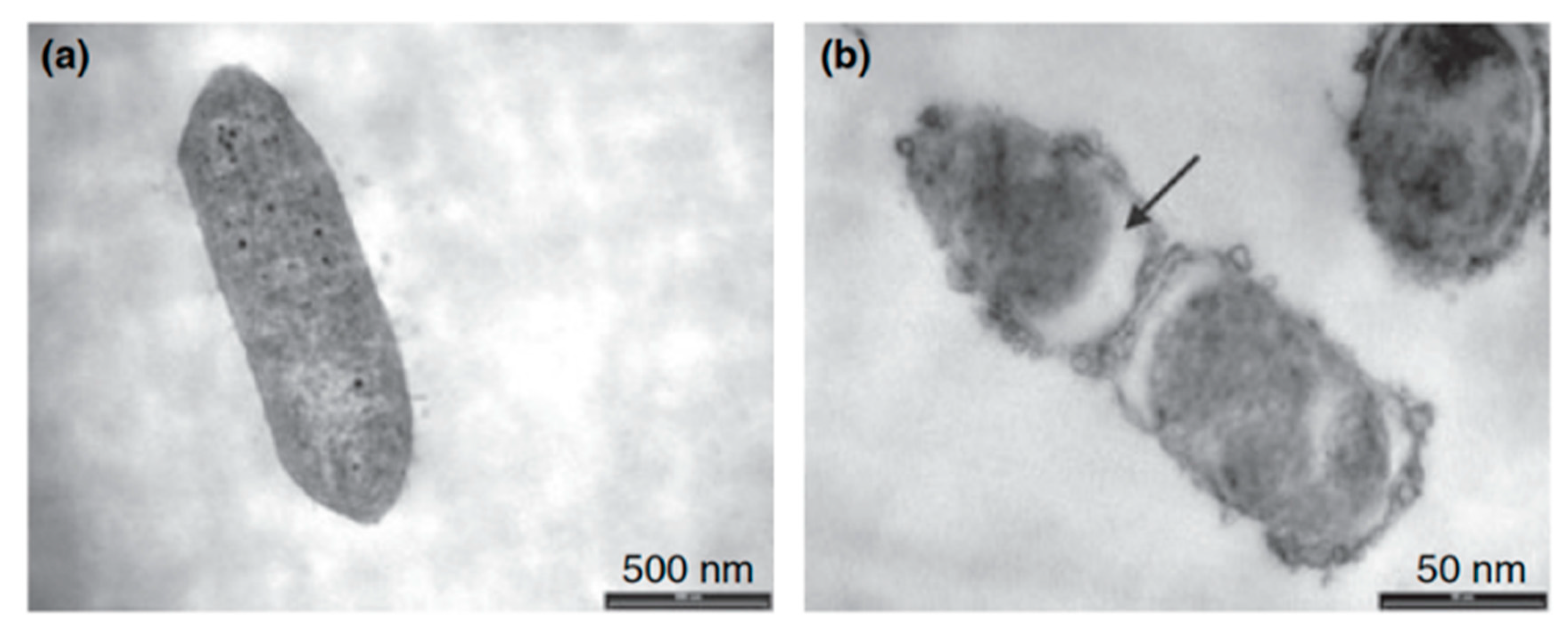
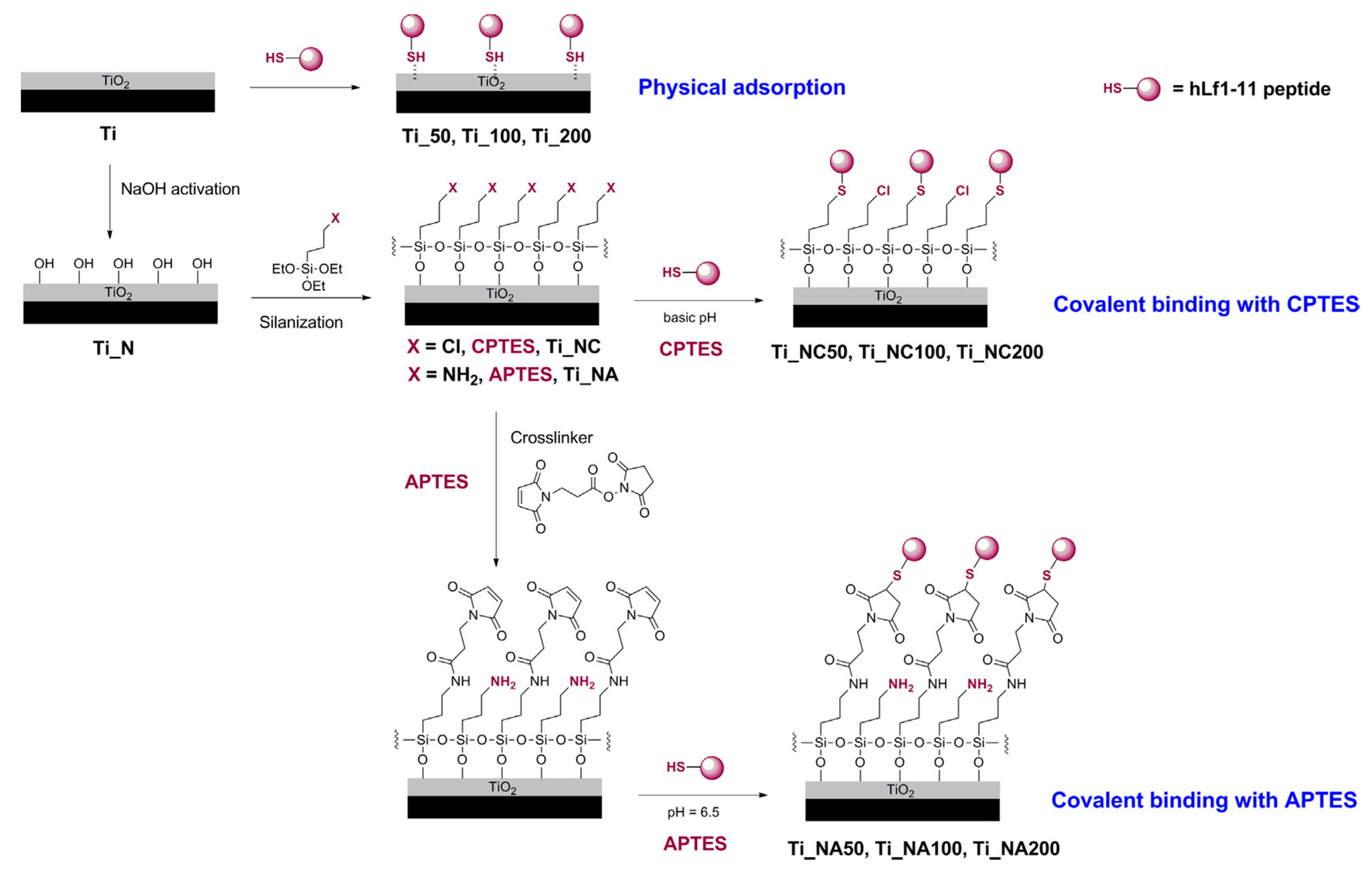
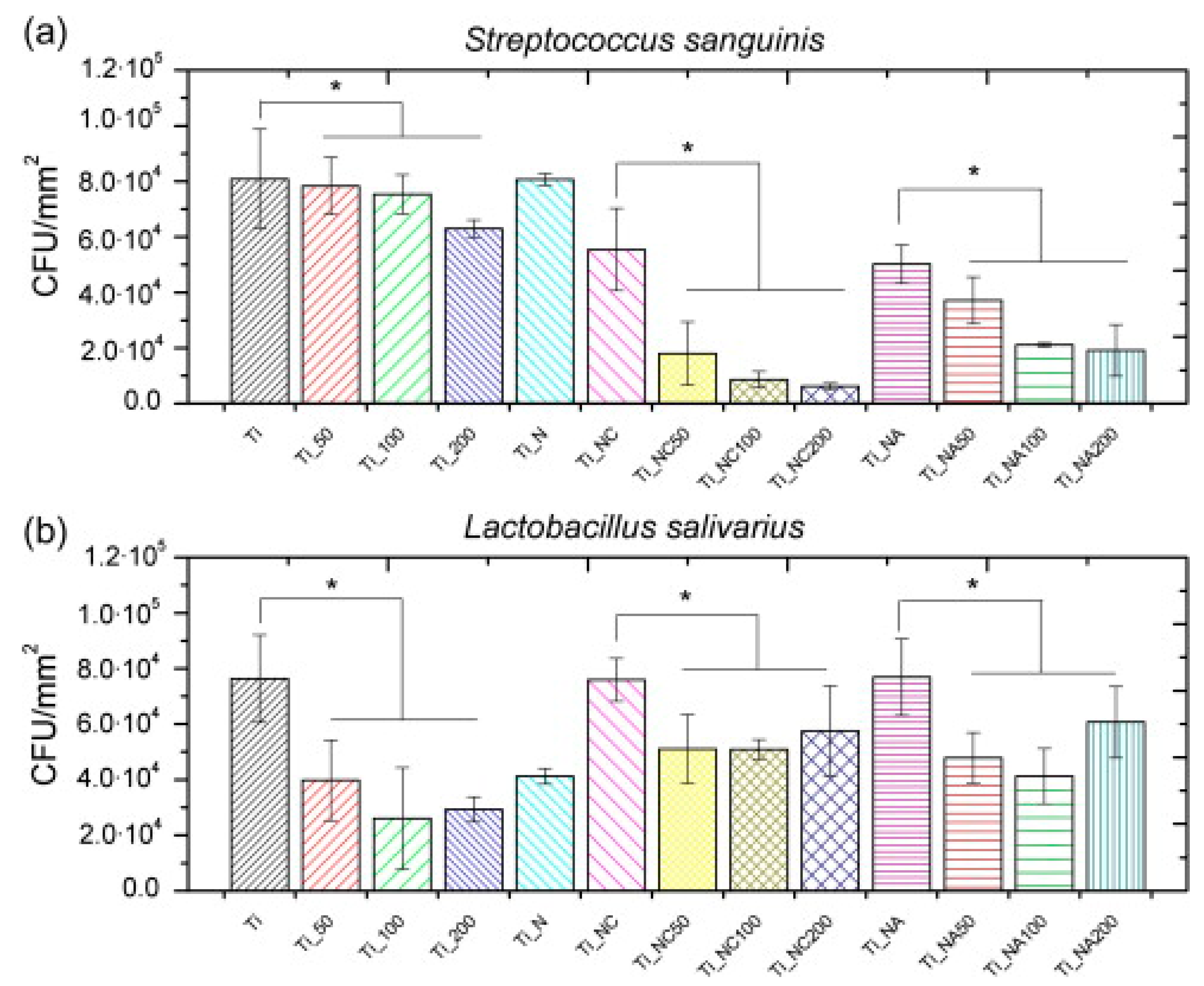
| hLF1-11 | Titanium surface silanized with CPTES or APTES | Peptide physical adsorption and covalent binding with CPTES or APTES | S. sanguinis, L. salivarius | Godoy-Gallardo et al., (2014) [85] |
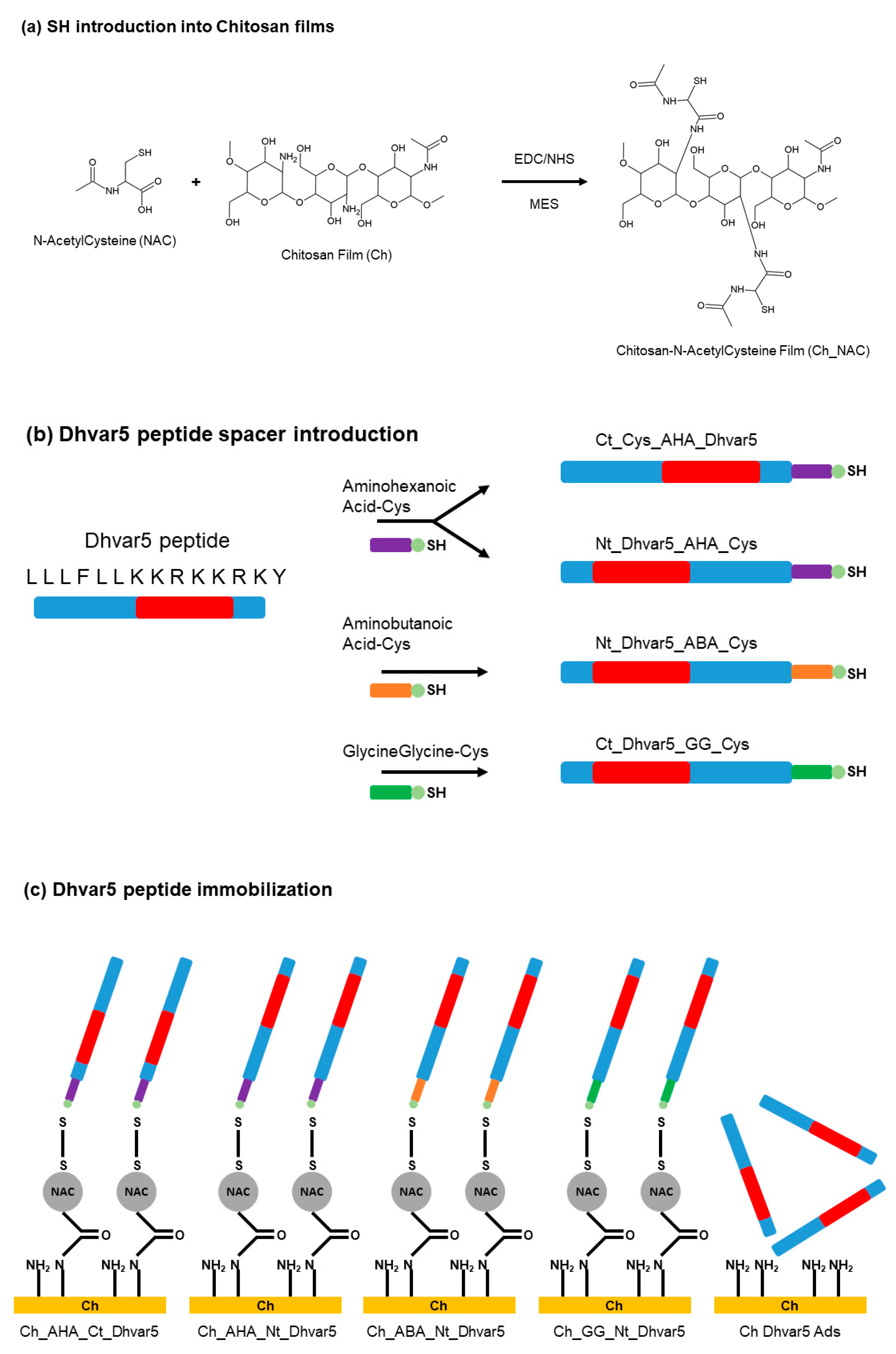
| Dhvar5 | Titanium and gold substrates | Physical adsorption and covalent binding with chitosan films, EDC-NHS chemistry and introduction of amino acids spacers | S. aureus | Costa et al., (2015) [66] |
| Defensin from Anopheles gambiae mosquitoes | Multilayer polyelectrolyte films of PEI-(PSS-PAH)2-(PGA-PLL)n | Layer-by-layer | M. luteus, E. coli | Etienne et al., (2004) [86] |
| Gramicidin A complexed with a non-denaturing anionic amphiphilic polysaccharide | Multilayer polyelectrolyte films of PEI and PLL | Layer-by-layer | E. faecalis | Guyomard et al., (2008) [87] |
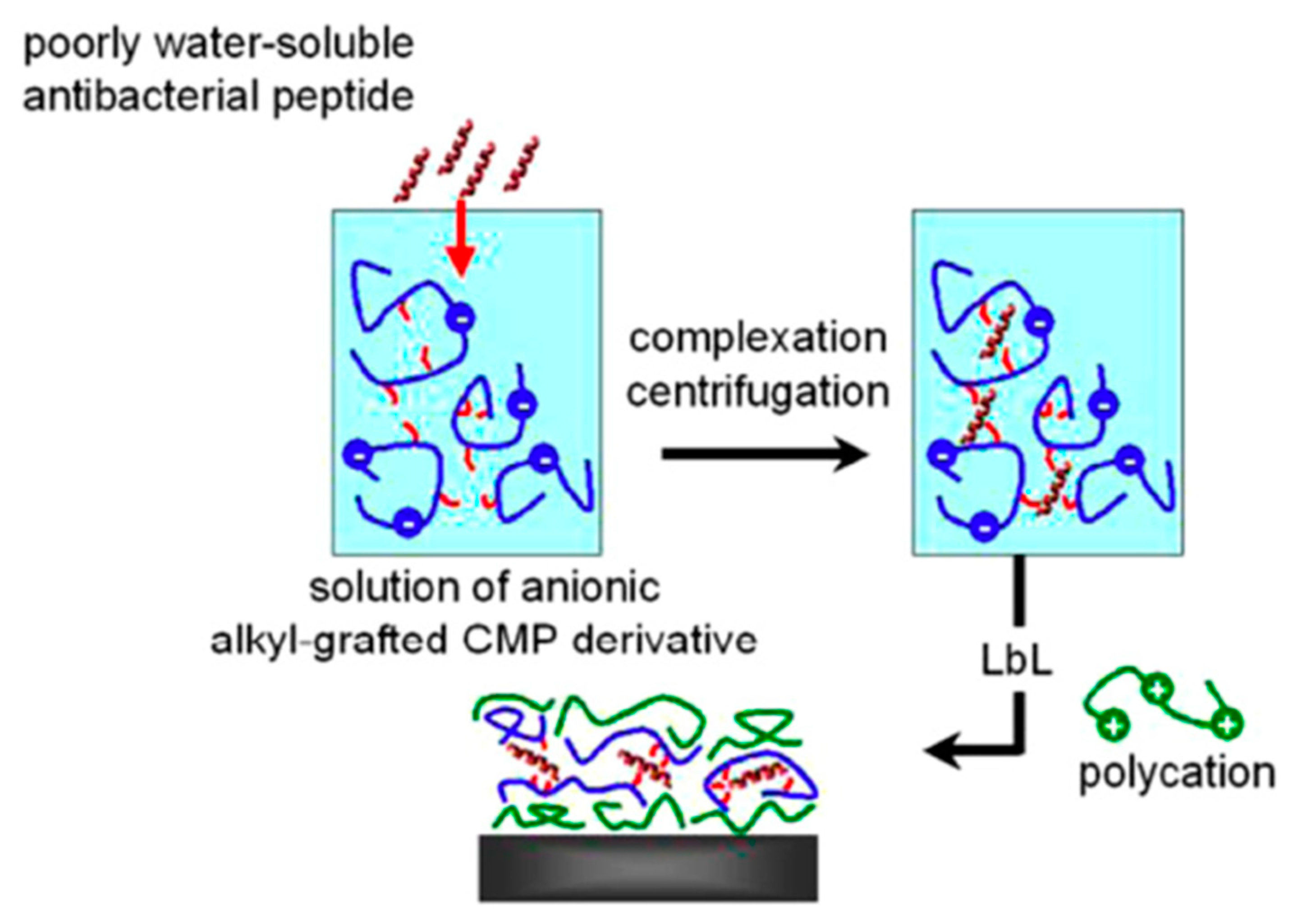
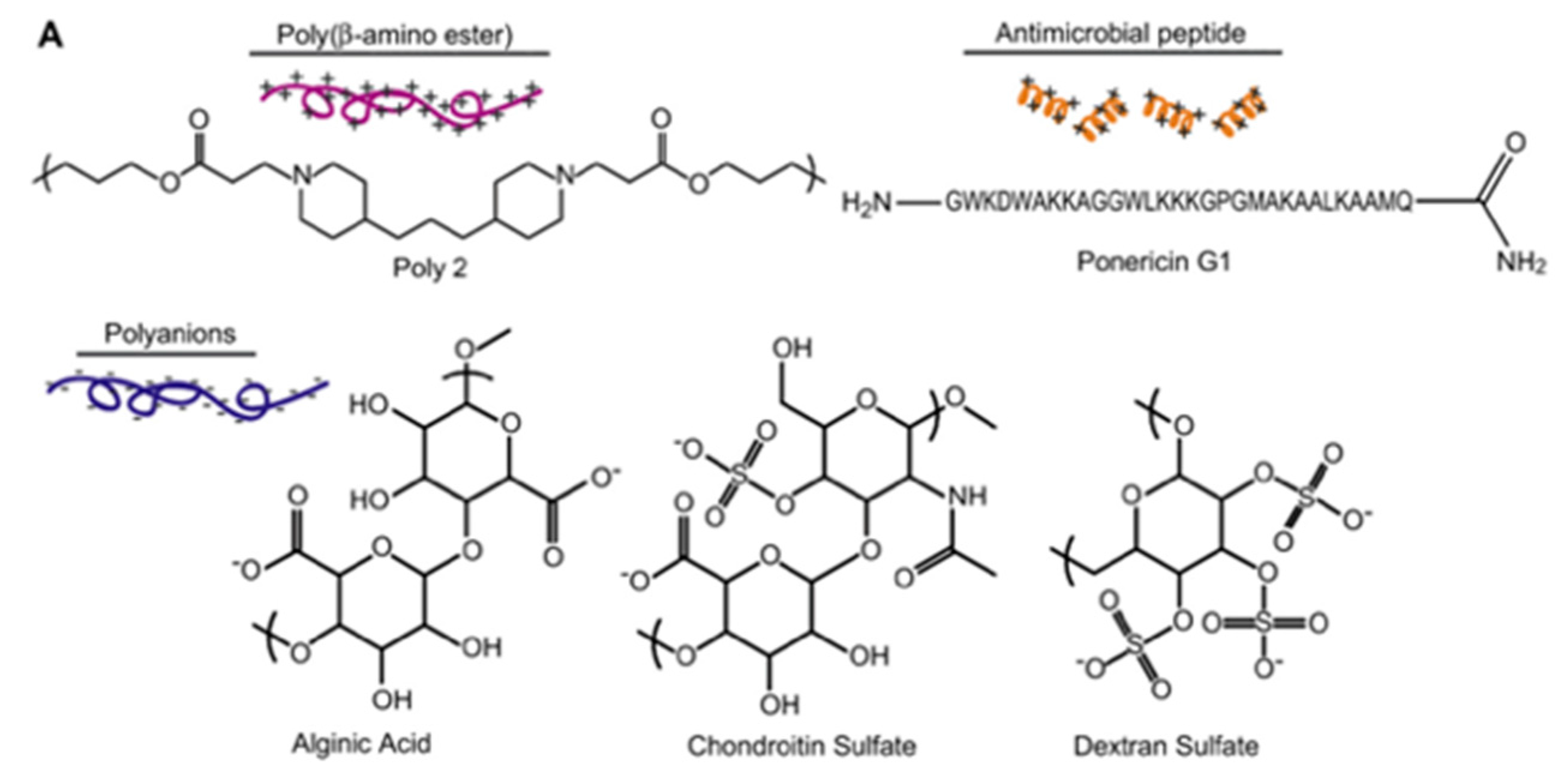
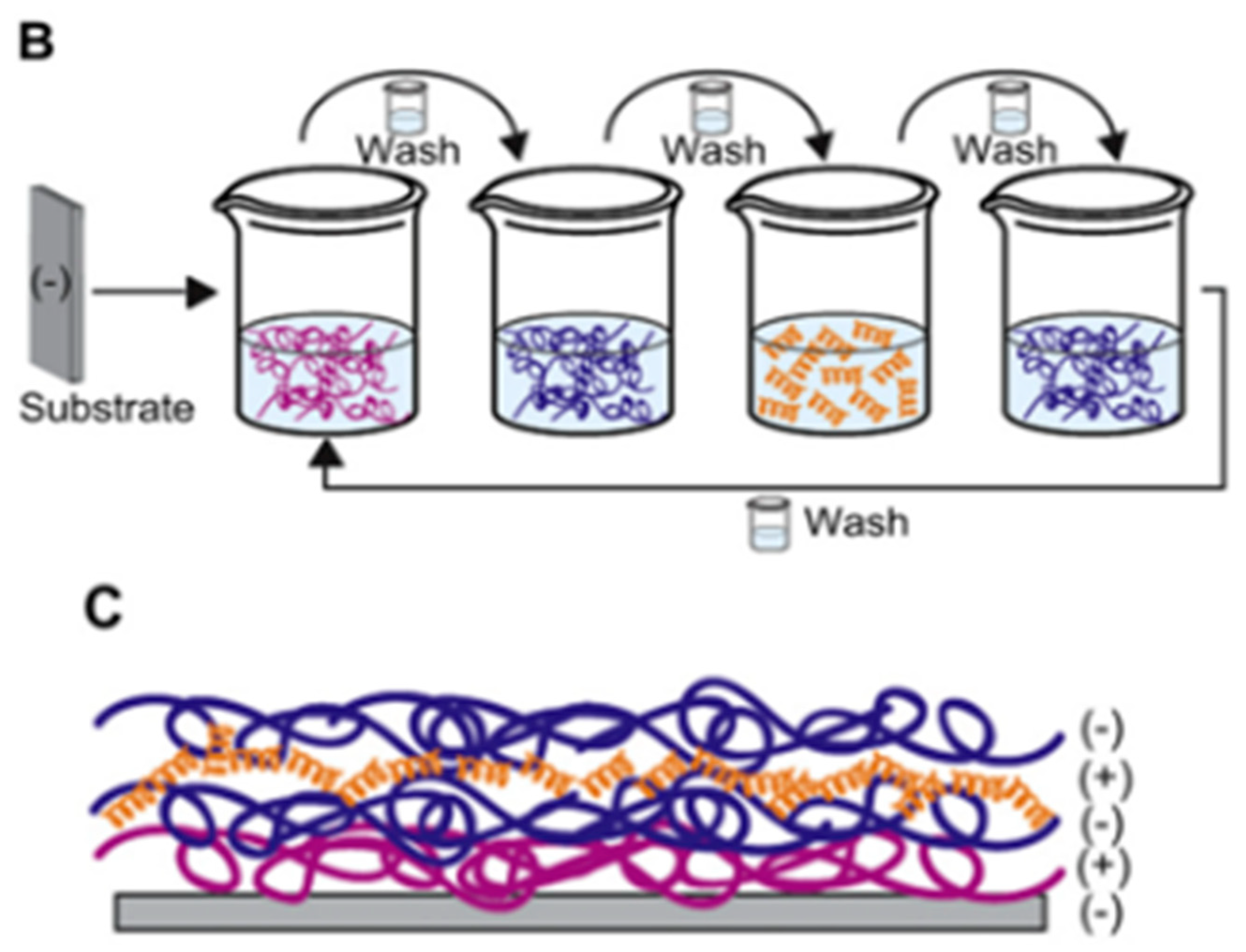
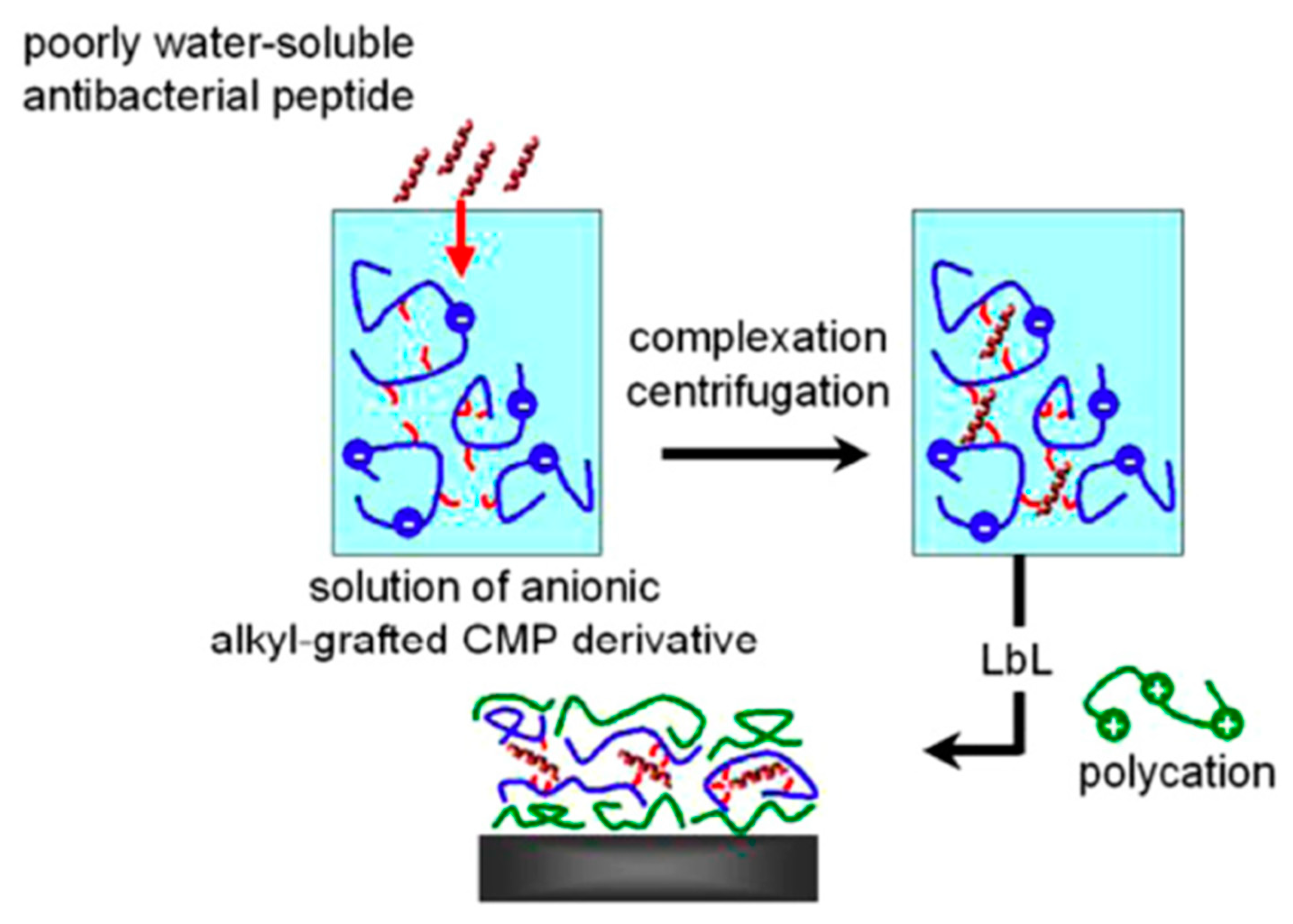
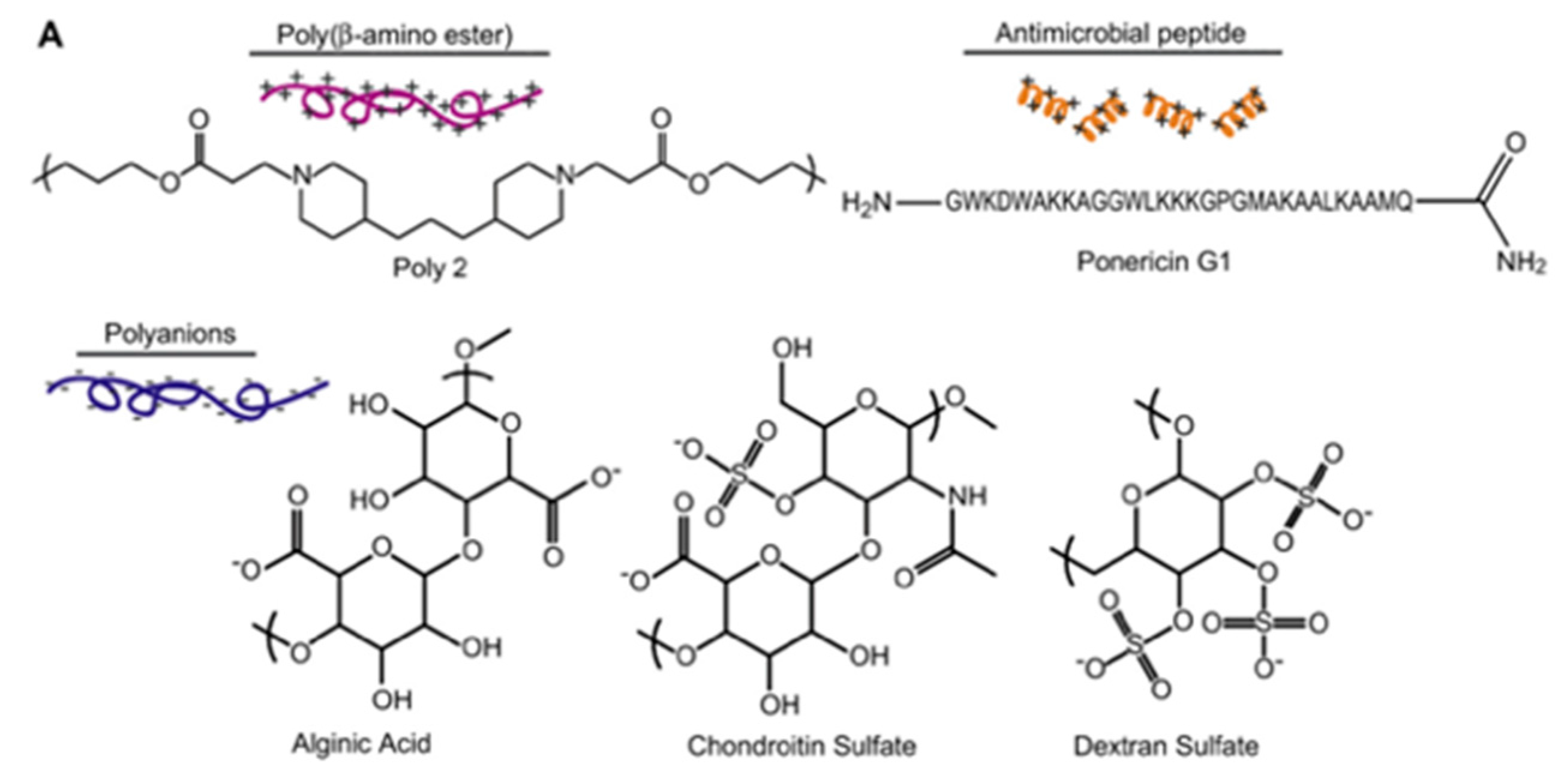
| Ponericin G1 | Silicone substrate | Layer-by-layer | S. aureus | Shukla et al., (2010) [88] |
| Magainin II | Stainless Steel | Modification of peptide with dopamine, direct grafting via catechol groups | C. farmer, V. natriegens | Cao et al., (2020) [89] |
| HHC-36 | PU substrates | Via APTES-Br SAM | E. coli, S. aureus | Zhang et al. 2019 [90] |
| MSI-78A | Au | SAMS EG4 thiols-biotin-neutravidin-maleimide | H. pilori | Parreira et al. 2019 [91] |
| MSI-78 | CaFé | Silica layer + maleimide | E. coli | Xiao et al. 2018 [92] |
4.1. Cross-Linking of Peptides to Surfaces
4.2. Direct Grafting of Modified Peptides to the Surface
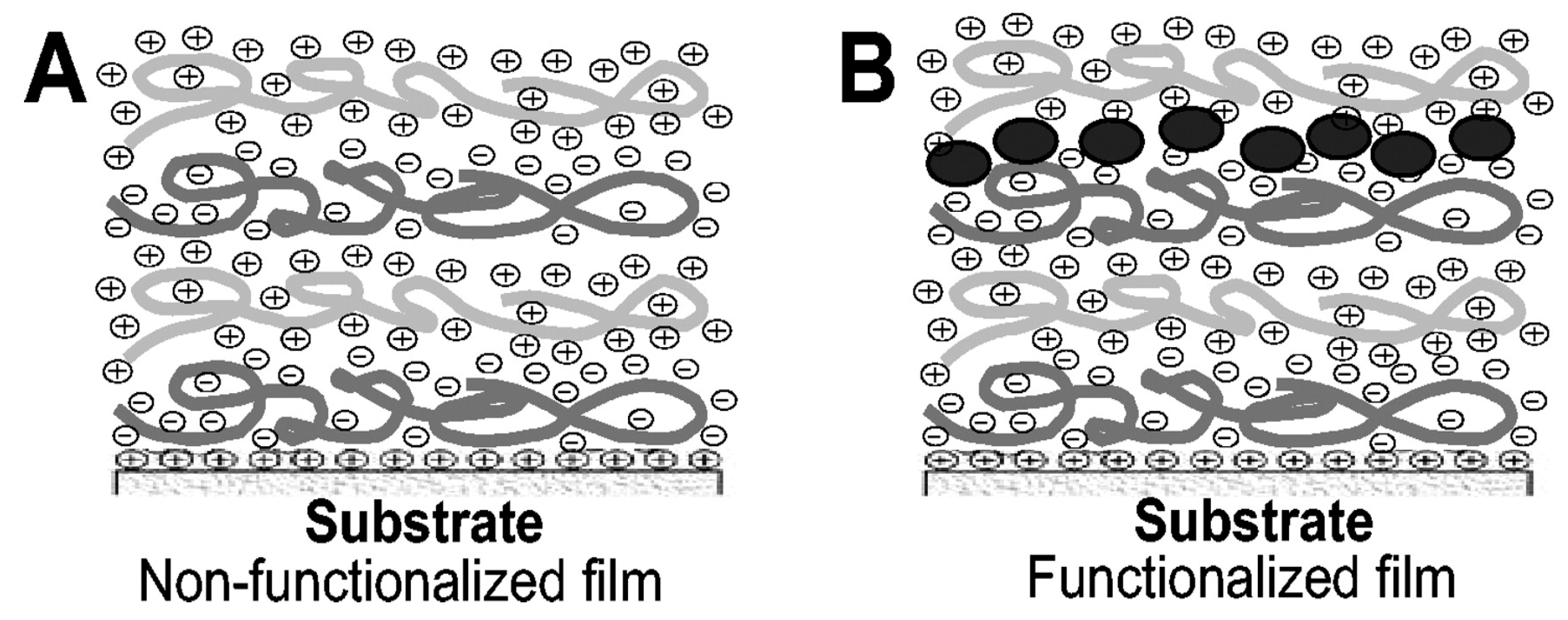
4.3. Comparison between Covalent Attachment and Physical Adsorption
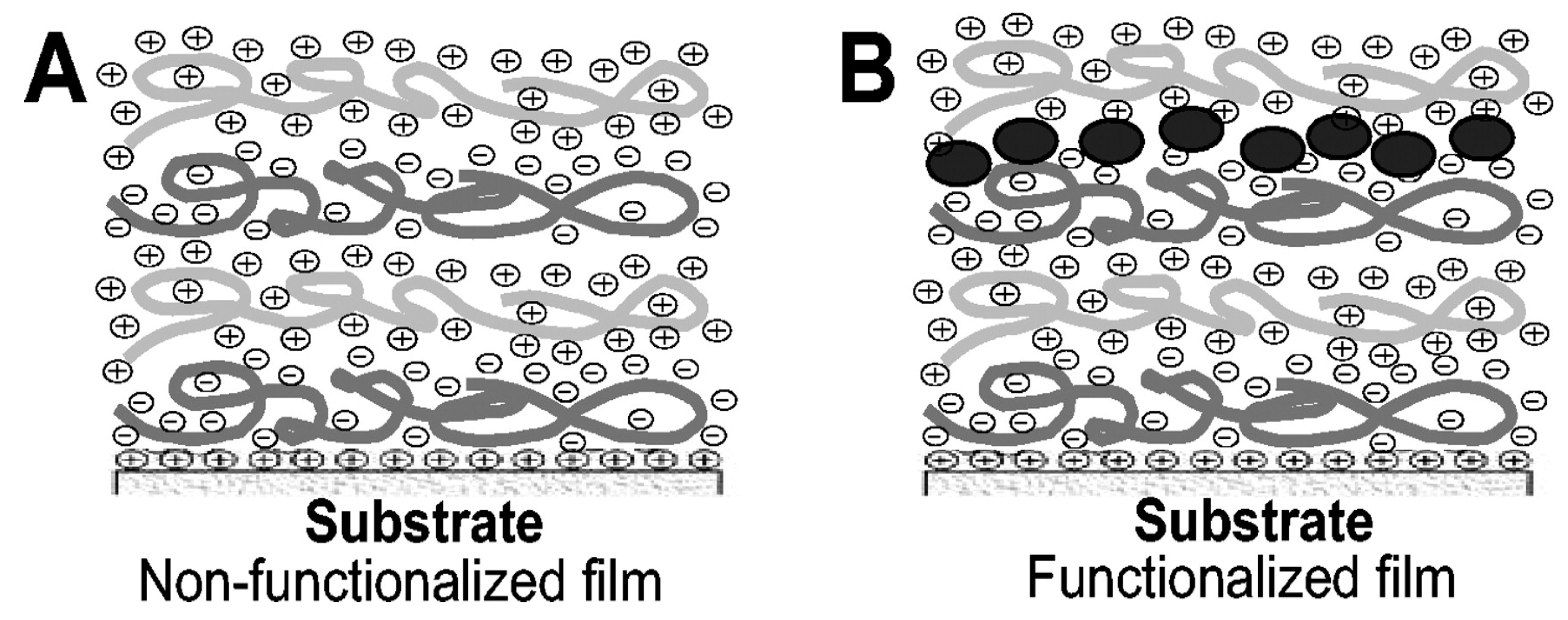
4.4. Layer-by-Layer Approach
5. Conclusions and Future Prospects
Author Contributions
Funding
Conflicts of Interest
List of Abbreviations
| AA | Amino Acid |
| AFM | Atomic Force Microscopy |
| AMP | AntiMicrobial Peptide |
| APD | Antimicrobial Peptide Database |
| APTES | 3-Aminopropyltriethoxysilane |
| ATP | Adenosine triphosphate |
| ATR-IR | Attenuated Total Reflection-Infra Red |
| CAPE | Cellulose-amino-hydroxypropyl ether |
| CFU | Colony-Forming Unit |
| CMP | Carboxymethylpullalans |
| CPTES | 3-Chloropropyltriethoxysilane |
| EDC | 1-Ethyl-3-(3-dimethylaminopropyl) carbodiimide |
| EPS | Extra Cellular Polymeric substance |
| FDA | Food and Drug Administration |
| HEMA | hydroxyethyl methacrylate |
| LbL | Layer-by-Layer |
| IL | Inhibition of Luminescence |
| MAA | Methacrylate Acid |
| MAG | Magainin |
| MIC | Minimal Inhibition Concentration |
| MUA | 11-mercaptoundecanoic acid |
| NHS | N-Hydroxysuccinimide |
| NMR | Nuclear Magnetic resonance |
| PDMA | Polydimethylacrylamide |
| PDMS | Polydimethylsiloxane |
| PEG | Poly(Ethylene Glycol) |
| PEM | Polyelectrolyte multilayers |
| PLL | Poly-L-ysine |
| PM-RAIRS | Polarisation Modulation Reflection Absorption Infrared Spectroscopy |
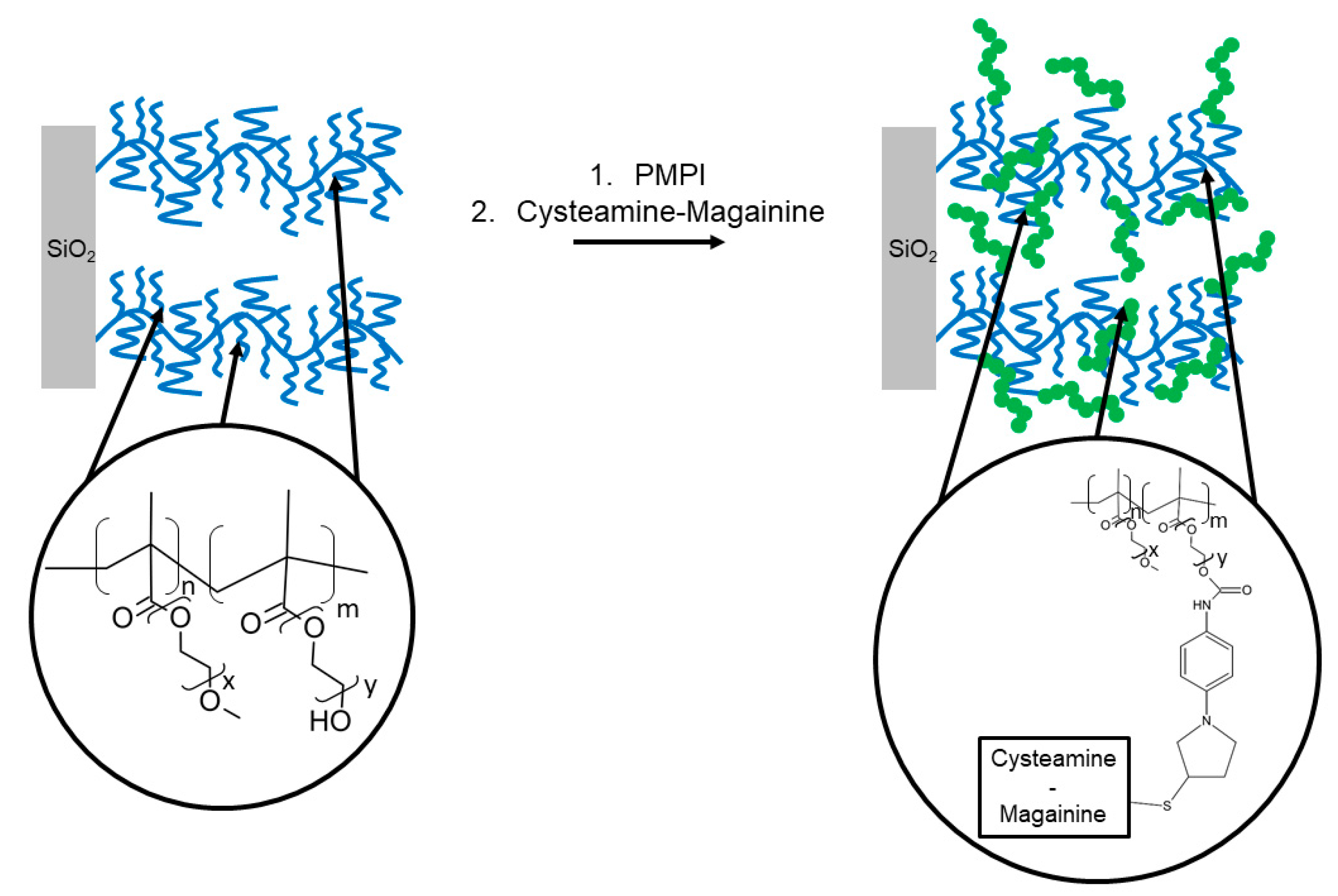
| PMPI | p-maleimidophenyl isocyanate |
| QS | Quorum Sensing |
| SAM | Self-Assembled Monolayer |
| SEM | Scanning Electron Microscopy |
| SI-ATRP | Surface-Initiated Atom Transfer Radical Polymerization |
| XPS | X-Ray Photoemission Spectroscopy |
References
- Sharma, D.; Misba, L.; Khan, A.U. Antibiotics versus biofilm: An emerging battleground in microbial communities. Antimicrob. Resist. Infect. Control 2019, 8, 76. [Google Scholar] [CrossRef] [PubMed]
- Glinel, K.; Thebault, P.; Humblot, V.; Pradier, C.M.; Jouenne, T. Antibacterial surfaces developed from bio-inspired approaches. Acta Biomater. 2012, 8, 1670–1684. [Google Scholar] [CrossRef] [PubMed]
- Bridier, A.; Briandet, R.; Thomas, V.; Dubois-Brissonnet, F. Resistance of bacterial biofilms to disinfectants: A review. Biofouling 2011, 27, 1017–1032. [Google Scholar] [CrossRef]
- Chen, M.; Yu, Q.; Sun, H. Novel Strategies for the Prevention and Treatment of Biofilm Related Infections. Int. J. Mol. Sci. 2013, 14, 18488–18501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvalho, I.; Henriques, M.; Carvalho, S. Microbial pathogens and strategies for combating them: Science, technology and education. New strategies to fight bacterial adhesion. FORMATEX 2013, 170–178. [Google Scholar]
- Jakobsen, T.H.; Tolker-Nielsen, T.; Givskov, M. Bacterial Biofilm Control by Perturbation of Bacterial Signaling Processes. Int. J. Mol. Sci. 2017, 18, 1970. [Google Scholar] [CrossRef] [Green Version]
- Graham, M.V.; Cady, N.C. Nano and Microscale Topographies for the Prevention of Bacterial Surface Fouling. Coatings 2014, 4, 37–59. [Google Scholar] [CrossRef] [Green Version]
- Jennings, M.C.; Ator, L.E.; Paniak, T.J.; Minbiole, K.P.C.; Wuest, W.M. Biofilm-Eradicating Properties of Quaternary Ammonium Amphiphiles: Simple Mimics of Antimicrobial Peptides. Chembiochem 2014, 15, 2211–2215. [Google Scholar] [CrossRef]
- Chandki, R.; Banthia, P.; Banthia, R. Biofilms: A microbial home. J. Indian Soc. Periodontol. 2011, 15, 111–114. [Google Scholar]
- Costerton, J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial biofilms: A common cause of persistent infections. Science 1999, 284, 1318–1322. [Google Scholar] [CrossRef] [Green Version]
- Jefferson, K.K. What drives bacteria to produce a biofilm? Fems Microbiol. Lett. 2004, 236, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Nwodo, U.U.; Green, E.; Okoh, A.I. Bacterial Exopolysaccharides: Functionality and Prospects. Int. J. Mol. Sci. 2012, 13, 14002–14015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutherland, I.W. Biofilm exopolysaccharides: A strong and sticky framework. Microbiology 2001, 147, 3–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donlan, R.M. Biofilm formation: A clinically relevant microbiological process. Clin. Infect. Dis. 2001, 33, 1387–1392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, M.R.; McGillicuddy, E.; Kaplan, L.J. Biofilm: Basic Principles, Pathophysiology, and Implications for Clinicians. Surg. Infect. 2014, 15, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Johnson, P.A. Novel understandings of host cell mechanisms involved in chronic lung infection: Pseudomonas aeruginosa in the cystic fibrotic lung. J. Infect. Public Health 2019, 12, 242–246. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, J.; Liu, F.; Yan, H.; Li, J.; Zhang, X.; Jha, A.K. Specific quorum sensing signal molecules inducing the social behaviors of microbial populations in anaerobic digestion. Bioresour. Technol. 2019, 273, 185–195. [Google Scholar] [CrossRef] [PubMed]
- James, G.A.; Swogger, E.; Wolcott, R.; Pulcini, E.D.; Secor, P.; Sestrich, J.; Costerton, J.W.; Stewart, P.S. Biofilms in chronic wounds. Wound Repair Regen. 2008, 16, 37–44. [Google Scholar] [CrossRef]
- Bryers, J.D. Medical Biofilms. Biotechnol. Bioeng. 2008, 100, 1–18. [Google Scholar] [CrossRef]
- Khardori, N.; Yassien, M. Biofilms in Devices-Related Infections. J. Ind. Microbiol. 1995, 15, 141–147. [Google Scholar] [CrossRef]
- Block, J.C. Biofilms in Drinking water distribution systems. In Biofilms—Science and Technology; NATO ASI Series (Series E: Applied Sciences); Melo, L., Bott, T.R., Fletcher, M., Capdeville, B., Eds.; Springer: Dordrecht, The Netherlands, 1992; pp. 469–485. [Google Scholar]
- Bott, T.R. Introduction to the problem of biofouling in industrial equipment. In Biofilms—Science and Technology; NATO ASI Series (Series E: Applied Sciences); Melo, L., Bott, T.R., Fletcher, M., Capdeville, B., Eds.; Springer: Dordrecht, The Netherlands, 1992; pp. 3–11. [Google Scholar]
- Mattila-Sandholm, T.; Wirtanen, G. Biofilm Formation in the Industry—A Review. Food Rev. Int. 1992, 8, 573–603. [Google Scholar] [CrossRef]
- Franklin, M.; Nivens, D.; Weadge, J.; Howell, P. Biosynthesis of the Pseudomonas aeruginosa Extracellular Polysaccharides, Alginate, Pel, and Psl. Front. Microbiol. 2011, 2, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, D. Understanding biofilm resistance to antibacterial agents. Nat. Rev. Drug Discov. 2003, 2, 114–122. [Google Scholar] [CrossRef]
- Palmer, J.; Flint, S.; Brooks, J. Bacterial cell attachment, the beginning of a biofilm. J. Ind. Microbiol. Biotechnol. 2007, 34, 577–588. [Google Scholar] [CrossRef] [PubMed]
- Klausen, M.; Heydorn, A.; Ragas, P.; Lambertsen, L.; Aaes-Jorgensen, A.; Molin, S.; Tolker-Nielsen, T. Biofilm formation by Pseudomonas aeruginosa wild type, flagella and type IV pili mutants. Mol. Microbiol. 2003, 48, 1511–1524. [Google Scholar] [CrossRef]
- Rabin, N.; Zheng, Y.; Opoku-Temeng, C.; Du, Y.; Bonsu, E.; Sintim, H.O. Biofilm formation mechanisms and targets for developing antibiofilm agents. Future Med. Chem. 2015, 7, 493–512. [Google Scholar] [CrossRef]
- Abu Bakar, M.; McKimm, J.; Haque, S.Z.; Majumder, M.A.A.; Haque, M. Chronic tonsillitis and biofilms: A brief overview of treatment modalities. J. Inflamm. Res. 2018, 11, 329–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, D.I.; Prenner, E.J.; Vogel, H.J. Tryptophan- and arginine-rich antimicrobial peptides: Structures and mechanisms of action. Biochim. Biophys. Acta Biomembr. 2006, 1758, 1184–1202. [Google Scholar] [CrossRef] [Green Version]
- Sitaram, N.; Nagaraj, R. Interaction of antimicrobial peptides with biological and model membranes: Structural and charge requirements for activity. Biochim. Biophys. Acta Biomembr. 1999, 1462, 29–54. [Google Scholar] [CrossRef] [Green Version]
- Hwang, P.M.; Vogel, H.J. Structure-function relationships of antimicrobial peptides. Biochem. Cell Biol. Biochim. Biol. Cell. 1998, 76, 235–246. [Google Scholar] [CrossRef]
- Zasloff, M. Magainins, a class of antimicrobial peptides from Xenopus skin—Isolation, characterization of 2 active forms, and partial CDNA sequence of a precursor. Proc. Natl. Acad. Sci. USA 1987, 84, 5449–5453. [Google Scholar] [CrossRef] [Green Version]
- Salditt, T.; Li, C.; Spaar, A. Structure of antimicrobial peptides and lipid membranes probed by interface-sensitive X-ray scattering. Biochim. Biophys. Acta Biomembr. 2006, 1758, 1483–1498. [Google Scholar] [CrossRef]
- Steinstraesser, L.; Kraneburg, U.M.; Hirsch, T.; Kesting, M.; Steinau, H.-U.; Jacobsen, F.; Al-Benna, S. Host Defense Peptides as Effector Molecules of the Innate Immune Response: A Sledgehammer for Drug Resistance? Int. J. Mol. Sci. 2009, 10, 3951–3970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, B.; Zhu, S. Mesobuthus Venom-Derived Antimicrobial Peptides Possess Intrinsic Multifunctionality and Differential Potential as Drugs. Front. Microbiol. 2018, 9, 320. [Google Scholar] [CrossRef] [Green Version]
- Holthausen, D.J.; Lee, S.H.; Kumar, V.T.V.; Bouvier, N.M.; Krammer, F.; Ellebedy, A.H.; Wrammert, J.; Lowen, A.C.; George, S.; Pillai, M.R.; et al. An Amphibian Host Defense Peptide Is Virucidal for Human H1 Hemagglutinin-Bearing Influenza Viruses. Immunity 2017, 46, 587–595. [Google Scholar] [CrossRef] [Green Version]
- Shen, B.; Song, J.; Zhao, Y.; Zhang, Y.; Liu, G.; Li, X.; Guo, X.; Li, W.; Cao, Z.; Wu, Y. Triintsin, a human pathogenic fungus-derived defensin with broad-spectrum antimicrobial activity. Peptides 2018, 107, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Terkawi, M.A.; Takano, R.; Furukawa, A.; Murakoshi, F.; Kato, K. Involvement of beta-defensin 130 (DEFB130) in the macrophage microbicidal mechanisms for killing Plasmodium falciparum. Sci. Rep. 2017, 7, 41772. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Chen, D.; Wang, L.; Lin, C.; Ma, C.; Xi, X.; Chen, T.; Shaw, C.; Zhou, M. Dermaseptin-PH: A Novel Peptide with Antimicrobial and Anticancer Activities from the Skin Secretion of the South American Orange-Legged Leaf Frog, Pithecopus (Phyllomedusa) hypochondrialis. Molecules 2017, 22, 1805. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ren, S.; Guo, C.; Zhang, W.; Zhang, X.; Zhang, B.; Li, S.; Ren, J.; Hu, Y.; Wang, H. Identification and functional analyses of novel antioxidant peptides and antimicrobial peptides from skin secretions of four East Asian frog species. Acta Biochim. Biophys. Sin. 2017, 49, 550–559. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.-H.; Zhang, J. CsCCL17, a CC chemokine of Cynoglossus semilaevis, induces leukocyte trafficking and promotes immune defense against viral infection. Fish Shellfish. Immunol. 2015, 45, 771–779. [Google Scholar] [CrossRef]
- Liu, S.; Long, Q.; Xu, Y.; Wang, J.; Xu, Z.; Wang, L.; Zhou, M.; Wu, Y.; Chen, T.; Shaw, C. Assessment of antimicrobial and wound healing effects of Brevinin-2Ta against the bacterium Klebsiella pneumoniae in dermally-wounded rats. Oncotarget 2017, 8, 111369–111385. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.; Bhatia, G.; Sharma, A.; Saxena, S. Host defense peptides: An insight into the antimicrobial world. J. Oral Maxillofac. Pathol. 2018, 22, 239–244. [Google Scholar] [CrossRef]
- Jenssen, H.; Hamill, P.; Hancock, R.E.W. Peptide antimicrobial agents. Clin. Microbiol. Rev. 2006, 19, 491–511. [Google Scholar] [CrossRef] [Green Version]
- Brogden, K.A. Antimicrobial peptides: Pore formers or metabolic inhibitors in bacteria? Nat. Rev. Microbiol. 2005, 3, 238–250. [Google Scholar] [CrossRef]
- Shai, Y. Mode of action of membrane active antimicrobial peptides. Biopolymers 2002, 66, 236–248. [Google Scholar] [CrossRef]
- Teixeira, V.; Feio, M.J.; Bastos, M. Role of lipids in the interaction of antimicrobial peptides with membranes. Prog. Lipid Res. 2012, 51, 149–177. [Google Scholar] [CrossRef] [PubMed]
- Bechinger, B. The structure, dynamics and orientation of antimicrobial peptides in membranes by multidimensional solid-state NMR spectroscopy. Biochim. Biophys. Acta Biomembr. 1999, 1462, 157–183. [Google Scholar] [CrossRef] [Green Version]
- Bin Hafeez, A.; Jiang, X.; Bergen, P.J.; Zhu, Y. Antimicrobial Peptides: An Update on Classifications and Databases. Int. J. Mol. Sci. 2021, 22, 11691. [Google Scholar] [CrossRef] [PubMed]
- Onaizi, S.A.; Leong, S.S.J. Tethering antimicrobial peptides: Current status and potential challenges. Biotechnol. Adv. 2011, 29, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef]
- Gordon, Y.J.; Romanowski, E.G.; McDermott, A.M. A review of antimicrobial peptides and their therapeutic potential as anti-infective drugs. Curr. Eye Res. 2005, 30, 505–515. [Google Scholar] [CrossRef]
- Giacometti, A.; Cirioni, O.; Kamysz, W.; Silvestri, C.; Del Prete, M.S.; Licci, A.; D’Amato, G.; Lukasiak, J.; Scalise, G. In vitro activity of citropin 1.1 alone and in combination with clinically used antimicrobial agents against Rhodococcus equi. J. Antimicrob. Chemother. 2005, 56, 410–412. [Google Scholar] [CrossRef]
- Koo, H.B.; Seo, J. Antimicrobial peptides under clinical investigation. Pept. Sci. 2019, 111, e24122. [Google Scholar] [CrossRef]
- Aoki, W.; Ueda, M. Characterization of Antimicrobial Peptides toward the Development of Novel Antibiotics. Pharmaceuticals 2013, 6, 1055–1081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, M.-D.; Won, H.-S.; Kim, J.-H.; Mishig-Ochir, T.; Lee, B.-J. Antimicrobial Peptides for Therapeutic Applications: A Review. Molecules 2012, 17, 12276–12286. [Google Scholar] [CrossRef] [Green Version]
- Koczulla, A.R.; Bals, R. Antimicrobial peptides—Current status and therapeutic potential. Drugs 2003, 63, 389–406. [Google Scholar] [CrossRef]
- Mahlapuu, M.; Hakansson, J.; Ringstad, L.; Bjorn, C. Antimicrobial Peptides: An Emerging Category of Therapeutic Agents. Front. Cell. Infect. Microbiol. 2016, 6, 194. [Google Scholar] [CrossRef] [Green Version]
- González Villa, T.; Veiga-Crespo, P. Antimicrobial Compounds: Current Strategies and New Alternatives; Springer: Berlin/Heidelberg, Germany, 2014; p. 268. [Google Scholar]
- Méndez-Samperio, P. Recent advances in the field of antimicrobial peptides in inflammatory diseases. Adv. Biomed. Res. 2013, 2, 50. [Google Scholar] [CrossRef] [PubMed]
- Giuliani, A.; Pirri, G.; Nicoletto, S.F. Antimicrobial peptides: An overview of a promising class of therapeutics. Cent. Eur. J. Biol. 2007, 2, 1–33. [Google Scholar] [CrossRef]
- Rapsch, K.; Bier, F.F.; Tadros, M.; von Nickisch-Rosenegk, M. Identification of Antimicrobial Peptides and Immobilization Strategy Suitable for a Covalent Surface Coating with Biocompatible Properties. Bioconjugate Chem. 2014, 25, 308–319. [Google Scholar] [CrossRef] [PubMed]
- Nie, B.E.; Long, T.; Ao, H.; Zhou, J.; Tang, T.; Yue, B. Covalent Immobilization of Enoxacin onto Titanium Implant Surfaces for Inhibiting Multiple Bacterial Species Infection and In Vivo Methicillin-Resistant Staphylococcus aureus Infection Prophylaxis. Antimicrob. Agents Chemother. 2017, 61, e01766. [Google Scholar] [CrossRef] [Green Version]
- Silva, R.R.; Avelino, K.Y.P.S.; Ribeiro, K.L.; Franco, O.L.; Oliveira, M.D.L.; Andrade, C.A.S. Chemical immobilization of antimicrobial peptides on biomaterial surfaces. Front. Biosci. 2016, 8, 129–142. [Google Scholar]
- Costa, F.; Carvalho, I.F.; Montelaro, R.C.; Gomes, P.; Martins, M.C.L. Covalent immobilization of antimicrobial peptides (AMPs) onto biomaterial surfaces. Acta Biomater. 2011, 7, 1431–1440. [Google Scholar] [CrossRef] [Green Version]
- Yu, K.; Lo, J.C.Y.; Mei, Y.; Haney, E.F.; Siren, E.; Kalathottukaren, M.T.; Hancock, R.E.W.; Lange, D.; Kizhakkedathu, J.N. Toward Infection-Resistant Surfaces: Achieving High Antimicrobial Peptide Potency by Modulating the Functionality of Polymer Brush and Peptide. Acs Appl. Mater. Interfaces 2015, 7, 28591–28605. [Google Scholar] [CrossRef]
- Bagheri, M.; Beyermann, M.; Dathe, M. Immobilization Reduces the Activity of Surface-Bound Cationic Antimicrobial Peptides with No Influence upon the Activity Spectrum. Antimicrob. Agents Chemother. 2009, 53, 1132–1141. [Google Scholar] [CrossRef] [Green Version]
- Alves, D.; Pereira, M.O. Mini-review: Antimicrobial peptides and enzymes as promising candidates to functionalize biomaterial surfaces. Biofouling 2014, 30, 483–499. [Google Scholar] [CrossRef]
- Atefyekta, S.; Blomstrand, E.; Rajasekharan, A.K.; Svensson, S.; Trobos, M.; Hong, J.; Webster, T.J.; Thomsen, P.; Andersson, M. Antimicrobial Peptide-Functionalized Mesoporous Hydrogels. Acs Biomater. Sci. Eng. 2021, 7, 1693–1702. [Google Scholar] [CrossRef]
- Humblot, V.; Yala, J.-F.; Thebault, P.; Boukerma, K.; Hequet, A.; Berjeaud, J.-M.; Pradier, C.-M. The antibacterial activity of Magainin I immobilized onto mixed thiols Self-Assembled Monolayers. Biomaterials 2009, 30, 3503–3512. [Google Scholar] [CrossRef]
- Gabriel, M.; Nazmi, K.; Veerman, E.C.; Amerongen, A.V.N.; Zentner, A. Preparation of LL-37-grafted titanium surfaces with bactericidal activity. Bioconjugate Chem. 2006, 17, 548–550. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.J.; Zhu, Y.C.; Xiong, M.H.; Hu, G.S.; Zhan, J.Z.; Li, T.J.; Wang, L.; Wang, Y.J. Antimicrobial Titanium Surface via Click-Immobilization of Peptide and Its in Vitro/Vivo Activity. Acs Biomater. Sci. Eng. 2019, 5, 1034–1044. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.; Yu, K.; Kindrachuk, J.; Brooks, D.E.; Hancock, R.E.W.; Kizhakkedathu, J.N. Antibacterial Surfaces Based on Polymer Brushes: Investigation on the Influence of Brush Properties on Antimicrobial Peptide Immobilization and Antimicrobial Activity. Biomacromolecules 2011, 12, 3715–3727. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.; Lange, D.; Hilpert, K.; Kindrachuk, J.; Zou, Y.; Cheng, J.T.J.; Kazemzadeh-Narbat, M.; Yu, K.; Wang, R.; Straus, S.K.; et al. The biocompatibility and biofilm resistance of implant coatings based on hydrophilic polymer brushes conjugated with antimicrobial peptides. Biomaterials 2011, 32, 3899–3909. [Google Scholar] [CrossRef]
- Glinel, K.; Jonas, A.M.; Jouenne, T.; Leprince, J.; Galas, L.; Huck, W.T.S. Antibacterial and Antifouling Polymer Brushes Incorporating Antimicrobial Peptide. Bioconjugate Chem. 2009, 20, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Lim, K.; Chua, R.R.Y.; Saravanan, R.; Basu, A.; Mishra, B.; Tarnbyah, P.A.; Ho, B.; Leong, S.S.J. Immobilization Studies of an Engineered Arginine-Tryptophan-Rich Peptide on a Silicone Surface with Antimicrobial and Antibiofilm Activity. Acs Appl. Mater. Interfaces 2013, 5, 6412–6422. [Google Scholar] [CrossRef]
- Li, X.; Li, P.; Saravanan, R.; Basu, A.; Mishra, B.; Lim, S.H.; Su, X.; Tambyah, P.A.; Leong, S.S.J. Antimicrobial functionalization of silicone surfaces with engineered short peptides having broad spectrum antimicrobial and salt-resistant properties. Acta Biomater. 2014, 10, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Masurier, N.; Tissot, J.-B.; Boukhriss, D.; Jebors, S.; Pinese, C.; Verdie, P.; Amblard, M.; Mehdi, A.; Martinez, J.; Humblot, V.; et al. Site-specific grafting on titanium surfaces with hybrid temporin antibacterial peptides. J. Mater. Chem. B 2018, 6, 1782–1790. [Google Scholar] [CrossRef]
- Hilpert, K.; Elliott, M.; Jenssen, H.; Kindrachuk, J.; Fjell, C.D.; Koerner, J.; Winkler, D.F.H.; Weaver, L.L.; Henklein, P.; Ulrich, A.S.; et al. Screening and Characterization of Surface-Tethered Cationic Peptides for Antimicrobial Activity. Chem. Biol. 2009, 16, 58–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sperandeo, P.; Bosco, F.; Clerici, F.; Polissi, A.; Gelmi, M.L.; Romanelli, A. Covalent Grafting of Antimicrobial Peptides onto Microcrystalline Cellulose. Acs Appl. Bio Mater. 2020, 3, 4895–4901. [Google Scholar] [CrossRef]
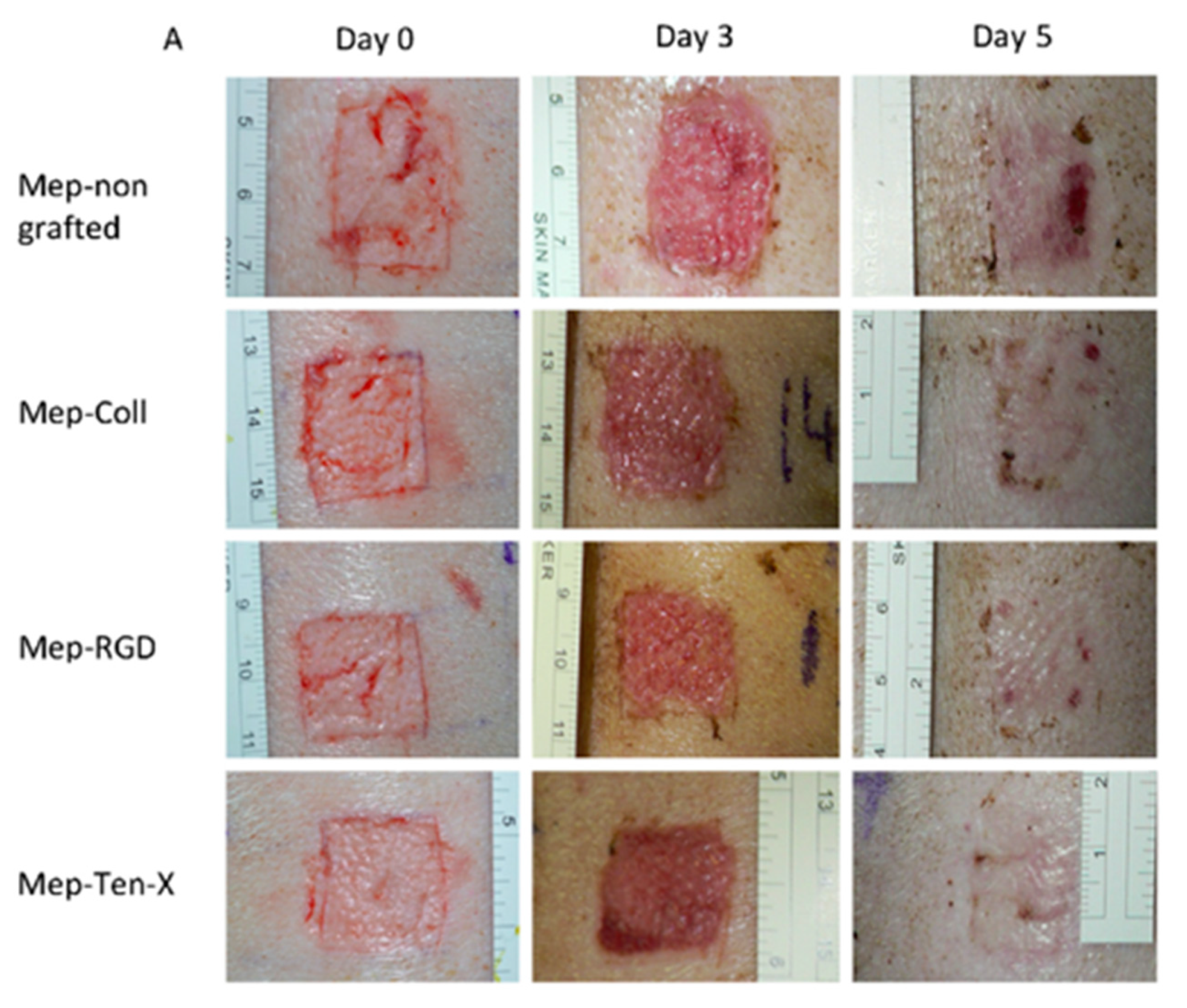
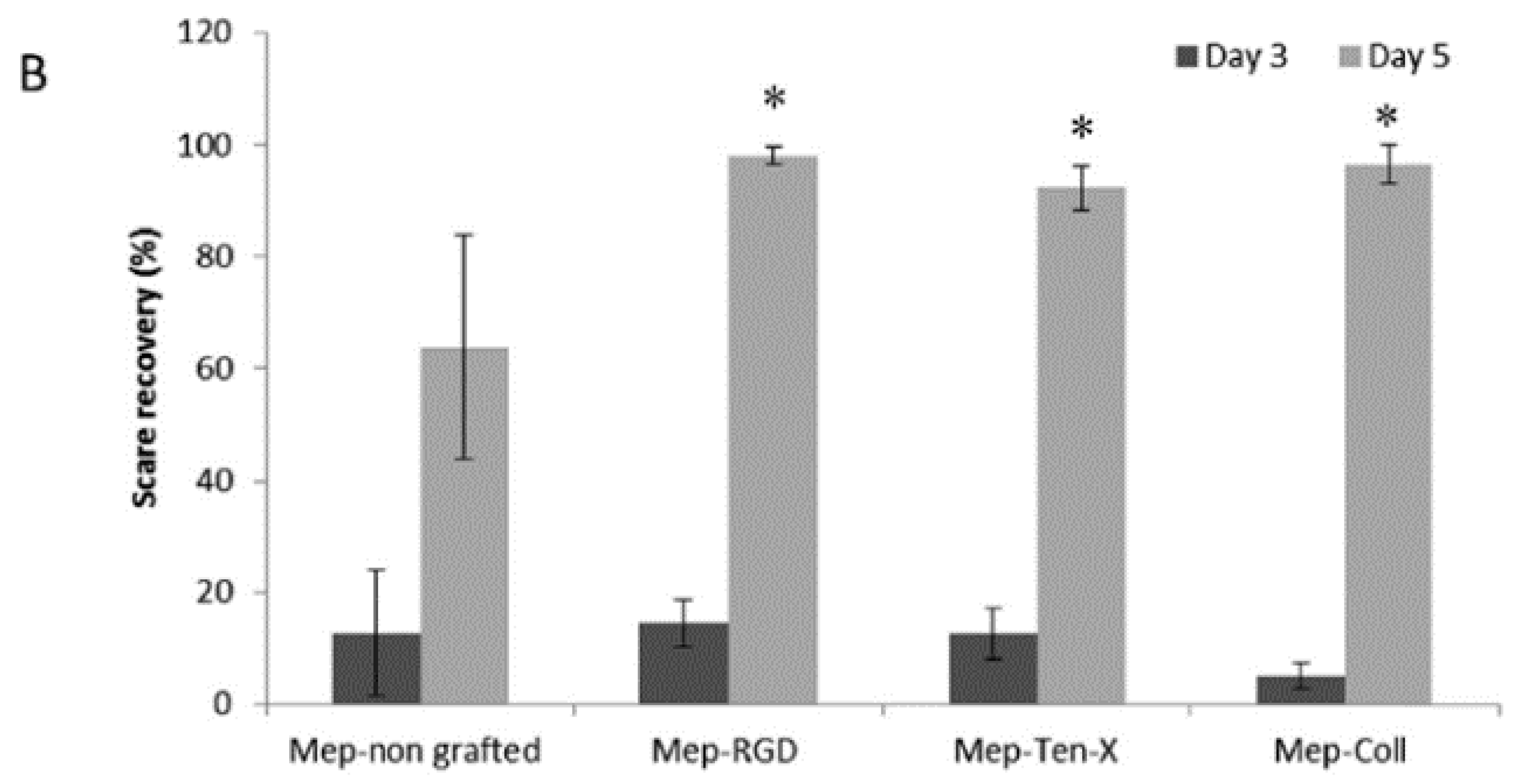
- Pinese, C.; Jebors, S.; Echalier, C.; Licznar-Fajardo, P.; Garric, X.; Humblot, V.; Calers, C.; Martinez, J.; Mehdi, A.; Subra, G. Simple and Specific Grafting of Antibacterial Peptides on Silicone Catheters. Adv. Healthc. Mater. 2016, 5, 3067–3073. [Google Scholar] [CrossRef]
- Pinese, C.; Jebors, S.; Stoebner, P.E.; Humblot, V.; Verdie, P.; Causse, L.; Garric, X.; Taillades, H.; Martinez, J.; Mehdi, A.; et al. Bioactive peptides grafted silicone dressings: A simple and specific method. Mater. Today Chem. 2017, 4, 73–83. [Google Scholar] [CrossRef]
- Willcox, M.D.P.; Hume, E.B.H.; Aliwarga, Y.; Kumar, N.; Cole, N. A novel cationic-peptide coating for the prevention of microbial colonization on contact lenses. J. Appl. Microbiol. 2008, 105, 1817–1825. [Google Scholar] [CrossRef]
- Godoy-Gallardo, M.; Mas-Moruno, C.; Fernandez-Calderon, M.C.; Perez-Giraldo, C.; Manero, J.M.; Albericio, F.; Gil, F.J.; Rodriguez, D. Covalent immobilization of hLf1-11 peptide on a titanium surface reduces bacterial adhesion and biofilm formation. Acta Biomater. 2014, 10, 3522–3534. [Google Scholar] [CrossRef]
- Etienne, O.; Picart, C.; Taddei, C.; Haikel, Y.; Dimarcq, J.L.; Schaaf, P.; Voegel, J.C.; Ogier, J.A.; Egles, C. Multilayer polyelectrolyte films functionalized by insertion of defensin: A new approach to protection of implants from bacterial colonization. Antimicrob. Agents Chemother. 2004, 48, 3662–3669. [Google Scholar] [CrossRef] [Green Version]
- Guyomard, A.; De, E.; Jouenne, T.; Malandain, J.-J.; Muller, G.; Glinel, K. Incorporation of a hydrophobic antibacterial peptide into amphiphilic polyelectrolyte multilayers: A bioinspired approach to prepare biocidal thin coatings. Adv. Funct. Mater. 2008, 18, 758–765. [Google Scholar] [CrossRef]
- Shukla, A.; Fleming, K.E.; Chuang, H.F.; Chau, T.M.; Loose, C.R.; Stephanopoulos, G.N.; Hammond, P.T. Controlling the release of peptide antimicrobial agents from surfaces. Biomaterials 2010, 31, 2348–2357. [Google Scholar] [CrossRef]
- Cao, P.; Du, C.W.; He, X.Y.; Zhang, C.; Yuan, C.Q. Modification of a derived antimicrobial peptide on steel surface for marine bacterial resistance. Appl. Surf. Sci. 2020, 510, 145512. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Zhao, Y.Q.; Zhang, Y.D.; Wang, A.Z.; Ding, X.K.; Li, Y.; Duan, S.; Ding, X.J.; Xu, F.J. Antimicrobial Peptide-Conjugated Hierarchical Antifouling Polymer Brushes for Functionalized Catheter Surfaces. Biomacromolecules 2019, 20, 4171–4179. [Google Scholar] [CrossRef]
- Parreira, P.; Monteiro, C.; Graca, V.; Gomes, J.; Maia, S.; Gomes, P.; Goncalves, I.C.; Martins, M.C.L. Surface Grafted MSI-78A Antimicrobial Peptide has High Potential for Gastric Infection Management. Sci. Rep. 2019, 9, 18212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, M.Y.; Jasensky, J.; Gerszberg, J.; Chen, J.J.; Tian, J.Y.; Lin, T.; Lu, T.Y.; Lahann, J.; Chen, Z. Chemically Immobilized Antimicrobial Peptide on Polymer and Self-Assembled Monolayer Substrates. Langmuir 2018, 34, 12889–12896. [Google Scholar] [CrossRef]
- Hilpert, K.; Volkmer-Engert, R.; Walter, T.; Hancock, R.E.W. High-throughput generation of small antibacterial peptides with improved activity. Nat. Biotechnol. 2005, 23, 1008–1012. [Google Scholar] [CrossRef]
- Schlusselhuber, M.; Humblot, V.; Casale, S.; Methivier, C.; Verdon, J.; Leippe, M.; Berjeaud, J.-M. Potent antimicrobial peptides against Legionella pneumophila and its environmental host, Acanthamoeba castellanii. Appl. Microbiol. Biotechnol. 2015, 99, 4879–4891. [Google Scholar] [CrossRef] [PubMed]
- Mishra, B.; Basu, A.; Chua, R.R.Y.; Saravanan, R.; Tambyah, P.A.; Ho, B.; Chang, M.W.; Leong, S.S.J. Site specific immobilization of a potent antimicrobial peptide onto silicone catheters: Evaluation against urinary tract infection pathogens. J. Mater. Chem. B 2014, 2, 1706–1716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mijnendonckx, K.; Leys, N.; Mahillon, J.; Silver, S.; Van Houdt, R. Antimicrobial silver: Uses, toxicity and potential for resistance. Biometals 2013, 26, 609–621. [Google Scholar] [CrossRef]
- Percival, S.L.; Bowler, P.G.; Russell, D. Bacterial resistance to silver in wound care. J. Hosp. Infect. 2005, 60, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Costa, F.; Maia, S.R.; Gomes, P.A.C.; Martins, M.C.L. Dhvar5 antimicrobial peptide (AMP) chemoselective covalent immobilization results on higher antiadherence effect than simple physical adsorption. Biomaterials 2015, 52, 531–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hancock, R.E.W.; Haney, E.F.; Gill, E.E. The immunology of host defence peptides: Beyond antimicrobial activity. Nat. Rev. Immunol. 2016, 16, 321–334. [Google Scholar] [CrossRef]
- Biswaro, L.S.; da Costa Sousa, M.G.; Rezende, T.M.B.; Dias, S.C.; Franco, O.L. Antimicrobial Peptides and Nanotechnology, Recent Advances and Challenges. Front. Microbiol. 2018, 9, 855. [Google Scholar] [CrossRef] [Green Version]
- Molchanova, N.; Hansen, P.R.; Franzyk, H. Advances in Development of Antimicrobial Peptidomimetics as Potential Drugs. Molecules 2017, 22, 1430. [Google Scholar] [CrossRef] [Green Version]
- Pfalzgraff, A.; Brandenburg, K.; Weindl, G. Antimicrobial Peptides and Their Therapeutic Potential for Bacterial Skin Infections and Wounds. Front. Pharmacol. 2018, 9, 281. [Google Scholar] [CrossRef]
- Porto, W.F.; Irazazabal, L.N.; Humblot, V.; Haney, E.F.; Ribeiro, S.M.; Hancock, R.E.W.; Ladram, A.; Franco, O.L. EcDBS1R6: A novel cationic antimicrobial peptide derived from a signal peptide sequence. Biochim. Biophys. Acta Gen. Subj. 2020, 1864, 129633. [Google Scholar] [CrossRef]
- Porto, W.F.; Irazazabal, L.; Alves, E.S.F.; Ribeiro, S.M.; Matos, C.O.; Pires, A.S.; Fensterseifer, I.C.M.; Miranda, V.J.; Haney, E.F.; Humblot, V.; et al. In silico optimization of a guava antimicrobial peptide enables combinatorial exploration for peptide design. Nat. Commun. 2018, 9, 129633. [Google Scholar] [CrossRef] [PubMed]







| AMP | AMP Source | Target a | Phase | Company |
|---|---|---|---|---|
| EA-230 | hCG derivative | Sepsis and renal failure | II | Exponential Biotherapies |
| protection | ||||
| CZEN-002 | α-MSH derivative | Anti-fungal | II | Zengen |
| XMP-629 | BPI derivative | Impetigo and acne rosacea | III | Xoma Ltd. |
| Neuprex(rBPI21) | BPR derivative | Pediatric meningococcemia | III | Xoma Ltd. |
| Delmitide(RDP58) | HLA class I derivative | Inflammatory bowel disease | II | Genzyme |
| Ghrelin | Endogenous HDP | Chronic respiratory failure | II | University of Miyazaki; Papworth Hospital |
| NVB-302 | Lantibiotic | C. difficile | I | Novacta |
| hLF1-11 | Lactoferricin derivative | MRSA, K. pneumoniae, L. monocytogenes | I/II | AM-Pharma |
| Wap-8294A2 (Lotilibcin) | Lysobactor spp. | G(+) bacteria(VRE, MRSA) | I/II | aRigen |
| DPK-060 | Kininogen derivative | Acute external otitis | II | ProMore Pharma |
| PXL01 | Lactoferrin analog | Postsurgical adhesions | III | ProMore Pharma |
| PAC113 | Histatin 5 analog | Oral candidiasis | II | Pacgen Biopharmaceuticals |
| POL7080 | Protegrin analog | P. aeruginosa K. pneumoniae | III | Polyphor Ltd. |
| OP-145 | LL-37 derivative | Chronic middle ear infection | II | Dr. Reddy’s Research |
| LL-37 | Human cathelicidin | Leg ulcer | II | ProMore Pharma |
| Novexatin (NP213) | Cyclic cationic peptide | Fungal nail infection | II | Novabiotics |
| Iseganan (IB-367) | Protegrin analog | Pneumonia, stomatitis | III | IntraBiotics Pharmaceuticals |
| Pexiganan (MSI-78) | Magainin analog | Diabetic foot ulcers | III | Dipexium Pharmaceuticals |
| Omiganan (CLS001) | Indolicidin derivative | Rosacea | III | Cutanea Life Sciences |
| Surotomycin | Cyclic lipopeptide | C. difficile (diarrhea) | III | Cubist Pharmaceuticals/Merck |
| Ramoplanin (NTI-851) | Actinoplanes spp. | G(+) (VRE, C. difficile) | III | Nano-therapeutics |
| Friulimicin B | Cyclic lipopeptide | Pneumonia, MRSA | I | MerLion Pharmaceuticals |
| MU1140 | Lantibiotic | G(+) bacteria (MRSA, C. difficile) | P | Oragenics |
| HB1275 | Lipopeptide | Fungal skin infections | P | Helix Biomedix |
| HB1345 | Lipopeptide | Skin infections, acne | P | Helix Biomedix |
| Arenicin (AP139) | Arenicola marina | G(−) bacteria, UTI | P | Adenium Biotech |
| AP114 | Arenicin analog | C. difficile | P | Adenium Biotech |
| AP138 | Arenicin analog | MRSA | P | Adenium Biotech |
| Novamycin (NP339) | Poly-arginine cationic peptide | Fungal infections | P | Novabiotics |
| Avidocin and Purocin | Modified bacteriocin | G(−) bacteria | P | Pylum Biosciences |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nicolas, M.; Beito, B.; Oliveira, M.; Tudela Martins, M.; Gallas, B.; Salmain, M.; Boujday, S.; Humblot, V. Strategies for Antimicrobial Peptides Immobilization on Surfaces to Prevent Biofilm Growth on Biomedical Devices. Antibiotics 2022, 11, 13. https://doi.org/10.3390/antibiotics11010013
Nicolas M, Beito B, Oliveira M, Tudela Martins M, Gallas B, Salmain M, Boujday S, Humblot V. Strategies for Antimicrobial Peptides Immobilization on Surfaces to Prevent Biofilm Growth on Biomedical Devices. Antibiotics. 2022; 11(1):13. https://doi.org/10.3390/antibiotics11010013
Chicago/Turabian StyleNicolas, Mathieu, Bruno Beito, Marta Oliveira, Maria Tudela Martins, Bruno Gallas, Michèle Salmain, Souhir Boujday, and Vincent Humblot. 2022. "Strategies for Antimicrobial Peptides Immobilization on Surfaces to Prevent Biofilm Growth on Biomedical Devices" Antibiotics 11, no. 1: 13. https://doi.org/10.3390/antibiotics11010013
APA StyleNicolas, M., Beito, B., Oliveira, M., Tudela Martins, M., Gallas, B., Salmain, M., Boujday, S., & Humblot, V. (2022). Strategies for Antimicrobial Peptides Immobilization on Surfaces to Prevent Biofilm Growth on Biomedical Devices. Antibiotics, 11(1), 13. https://doi.org/10.3390/antibiotics11010013