1. Introduction
An important area of endothelial biology is understanding how the physical properties of an endothelial barrier are formed and regulated under normal physiological homeostasis as well as during disease states, acute infections, and by a range of other insults (e.g., drugs, physical trauma) [
1]. For in vitro-based research, this requires advanced technologies to investigate the barrier function, ideally in real-time and in a noninvasive continuous manner. In this paper, we highlight the power and utility of electrical cell-substrate impedance sensing (ECIS) biosensor technology to reveal massive differences in the capability of brain endothelial cells to form a barrier by subtly varying the ingredients of the culture medium.
ECIS biosensor technology was developed by Applied Biophysics (USA) and has a broad range of applications depending on the cell type of interest. For endothelial cells and epithelial cells, ECIS technology is capable of measuring and modelling a number of barrier-related parameters as well as changes in cellular behaviour [
2]. This is possible because ECIS is a multifrequency AC impedance biosensor. Impedance (Z; ohms) is comprised of resistance (R) and capacitance (C). Capacitance provides measurements related to the overall coverage of the well by the cell layer, whereas resistance is indicative of the barrier functions of the endothelial cells. Due to the multifrequency nature of ECIS, the impedance data can be mathematically modelled to indicate the contribution of the paracellular junctional space, the basal adhesion of the cells, and the cell membrane to the total impedance to current. These modelled values are referred to as R
b, α, and C
m, respectively [
3]. Other impedance biosensors such as the xCELLigence RTCA system from ACEA, which we have used extensively for a variety of research applications, are not capable of conducting multifrequency impedance measurements and therefore cannot derive the contributions from the various cellular barrier compartments [
4,
5,
6,
7,
8]. ECIS biosensor technology is the only commercially available system capable of measuring and modelling these parameters, and in this paper, we highlight a simple example of its power to do so.
Brain endothelial cells, like all endothelial cells, have specific intrinsic properties that control the formation and function of the barrier between the brain parenchyma and the blood [
1,
9]. In order for these properties to be realised, brain endothelial cells must be cultured under suitable culture conditions to produce a strong and durable barrier [
10,
11]. Here, we demonstrate the power of ECIS biosensor technology to reveal that subtle differences in culture conditions have substantial effects on barrier formation of an immortalised brain endothelial cell line, which we have extensively characterised previously [
6,
7,
8,
12]. Importantly, the R
b component of the barrier was the parameter affected greatest by the media modifications, and here, we show how rapidly these changes can occur. Furthermore, these differences translate into changes in expression of key paracellular junctional proteins which were assessed using immunocytochemistry. This paper exemplifies the necessity of ECIS measurements being able to separate changes occurring at the paracellular vs. basal adhesion level, the value of the measurements being conducted in real-time to provide a temporal profile of the barrier formation, and correlating the subsequent changes that occur to overall impedance measurements.
2. Materials and Methods
2.1. Cell Culture
Human cerebral microvascular endothelial cells (HCMVEC’s) (cat# T0259, ABM Good, Richmond, BC, Canada) were grown and passaged in T75 flasks coated with 1 µg/mL rat tail collagen I (Gibco, Waltham, MA, USA). Cells were maintained in M199 media (Gibco, Waltham, MA, USA) supplemented with 10% FBS, 1 µg/mL hydrocortisone (Sigma, St Louis, MO, USA), 80 µM butyryl cAMP (Sigma, St Louis, MO, USA), 10 µg/mL heparin (Sigma), 1 ng/mL human EGF (Peprotech, Rocky Hill, NJ, USA), 3 ng/mL human FGF (Peprotech, Rocky Hill, NJ, USA), and 2 mM Glutamax (Gibco, Waltham, MA, USA). This media is referred to as Enriched Media. In some experiments, different media were used as described in the figure legends. Minimal Media contained M199 media supplemented with 2% FBS, 39 ng/mL hydrocortisone, and 80 µM butyrul cAMP. Cells were maintained at 37 °C in a humidified 5% CO2 incubator.
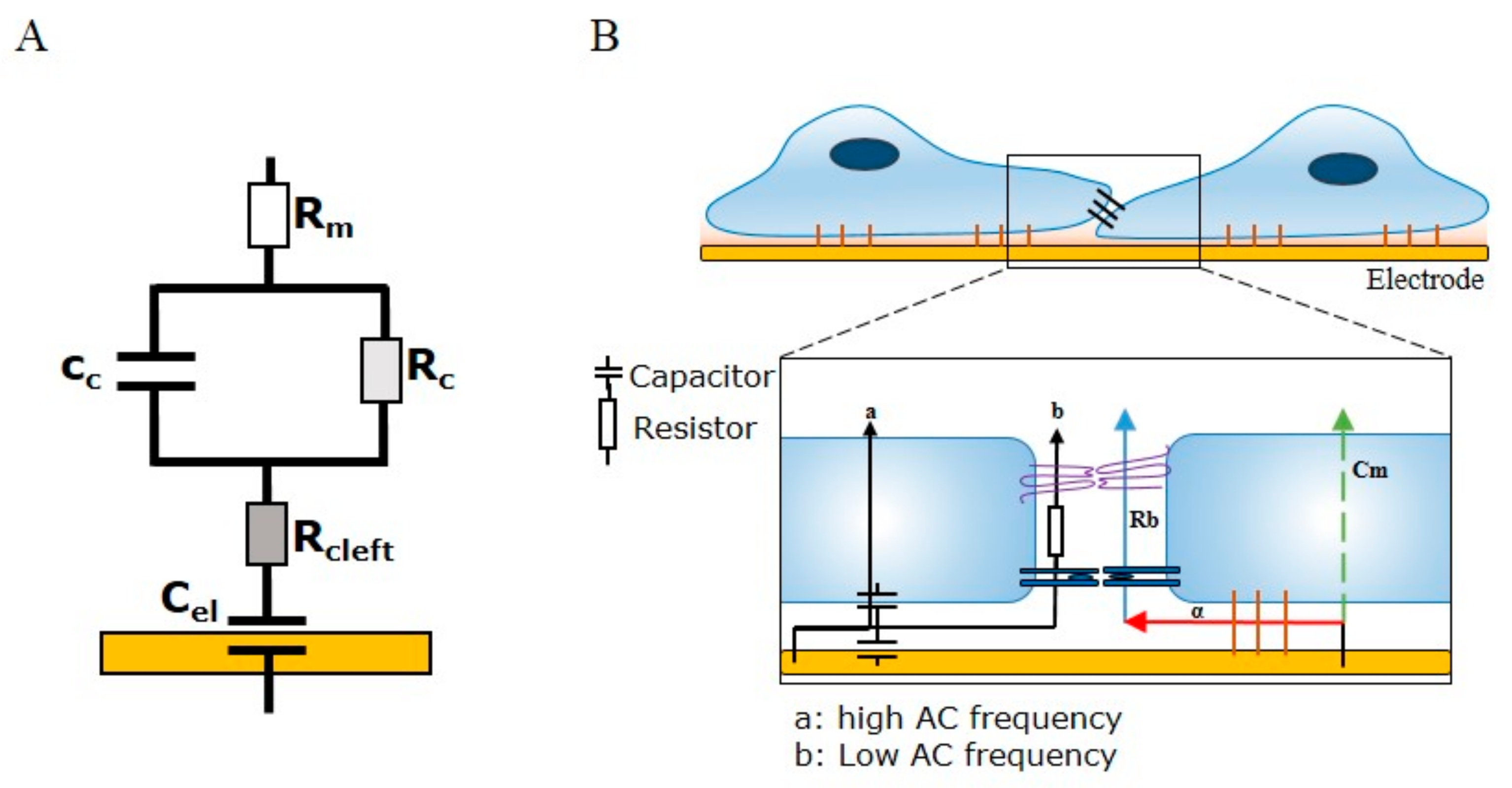
2.2. ECIS Theory and Modelling
ECIS involves the growth of cells on top of gold interdigitating electrodes. The application of a weak alternating current (10–10
5 Hz) through the electrode array provides a means of measuring the ability of cells grown in a monolayer to impede the movement of electrons through and between individual cells. The impedance measurement provides information on two indicators of cell behaviour—resistance (R) and capacitance (C). While capacitance provides measurements related to the overall coverage of the electrode by the cell layer, resistance is indicative of the barrier function of the cells. The principle behind being able to separate impedance (Z) into the two cell-behaviour indicators is dependent on the AC frequency (
f) passing through the cell layer, as adherent cells are able to alter the flow of current through a monolayer in a frequency-dependent manner. At low frequencies (10
2–10
4 Hz), the AC is less able to pass through the cell body due to the capacitive effect of the cell membrane largely resisting current flow (reactance;
), forcing the current to flow around the cell bodies and thus through the intercellular space. At low frequencies, therefore, the movement of current between the cells is mostly restricted by the presence of intercellular junctions. Conversely, at high frequencies (>10,000 Hz), the current is able to flow through the cell body due to low reactive capacitance, where the capacitive function of the cell membrane is indicative of the degree of cell coverage over the electrode. Capacitance is useful, as changes in the area (A) of cell membranes and distance (d) of cells above the electrode allow for interpretations of cell attachment, cell spreading, and cell loss changes (as per the formula: capacitance =
). The power of ECIS is further enhanced by the ability to apply mathematical modelling to derive three parameters that describe three related properties of cells: R
b (paracellular barrier), α (basal adhesion), and C
m (cell membrane capacitance). Of note, to model the parameter R
b, four assumptions of cell behaviour need to be met: (I) the electrode needs to be covered by a confluent monolayer; (II) the cells need to have a uniform radius and distance above the electrode; (III) the current needs to be flowing radially within the space between the cells and electrode; and (IV) the current density needs to remain constant [
3]. In terms of modelling endothelial barrier function in vitro, of most importance is the parameter R
b (Ω cm
2), as it describes the tightness of the intercellular space, which is highly dependent on cell–cell junctions. The two remaining parameters, α (Ω
0.5 cm) and C
m (μF/cm
2), are indicative of the current flow below cells or through cells, respectively. The parameter α is useful in describing changes in cell radius and basal adhesion, while C
m describes the changes in membrane composition as a function of capacitance. ECIS biosensor technology is the only biosensor technology currently available that can model each of these important cellular parameters.
Figure 1 summarises the principles of ECIS and the theory behind barrier integrity measurements and the modelled parameters α, R
b, and C
m.
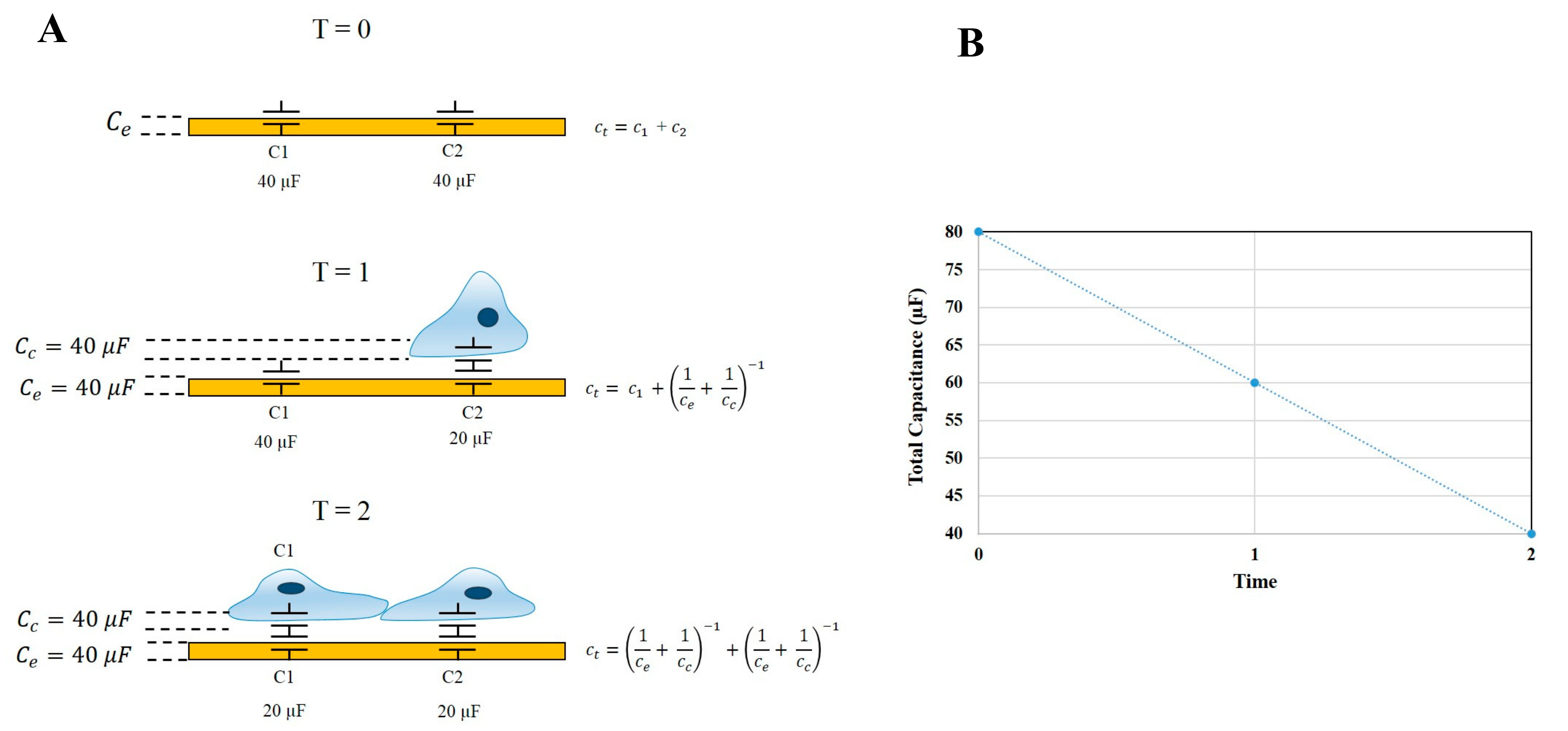
An important function of ECIS-Zθ technology is the ability to derive capacitance readings from the measured impedance. Capacitance measurements relate to the ability of the electrode-substrate and cell-substrate interfaces to accumulate charge on their respective surfaces. In a cell-free well, the total capacitance of the system is equivalent to the capacitance of the electrode. However, as cells begin to adhere and cover the electrode arrays, the total capacitance (C
T) is derived from the combined electrode capacitance (C
e) and cell capacitance (C
c). The capacitance of the electrode and cell system can be simplified and represented as being equivalent to two capacitors in series, as per the equation
. Therefore, as cells progressively cover the gold electrodes, C
T is no longer equivalent to C
e, but instead, a greater proportion of C
T is influenced by the two in-series capacitors of the cell-electrode system. Thus, by definition, as C
c increases (as a function of the cell area increasing), C
T must decrease. This is depicted in
Figure 2, where at the point of cell seeding, the capacitance of the system is high and is followed by a steady decrease while cells adhere and spread across the electrodes. Using this theory, any increase in capacitance can be attributed to either an increase in electrode exposure as cells reorganise their junctional space or changes in the physical capacitive properties of the cell membrane, such as changes in composition, area, and thickness.
2.3. Conducting ECIS Experiments
ECIS experimental procedure involved pretreating a 96w20idf ECIS plate with 10 mM L-cysteine prior to coating with 1 µg/mL collagen I (Gibco, USA). The endothelial cells were seeded at 20,000 cells per well in 100 µL of media (see figure legend for specific type) and were monitored until a barrier had formed, typically ~48 h post seeding. Upon barrier formation, media was completely removed and replaced with 100 µL of each respective media type being investigated. The endothelial barrier resistance was then monitored for at least 48 h, at which point multifrequency data was collected and modelled using ECIS software (Applied Biophysics, Troy, NY, USA). ECIS measurements were acquired from three independent biological repeats with three replicates per experiment. Shown is one independent experiment with three replicates, which is representative of the three biological repeats. The results were graphed using GraphPad Prism 7.03 software (GraphPad Software Inc., LaJolla, CA, USA) and represented as the average mean ± SD of the three replicates.
2.4. Immunocytochemistry
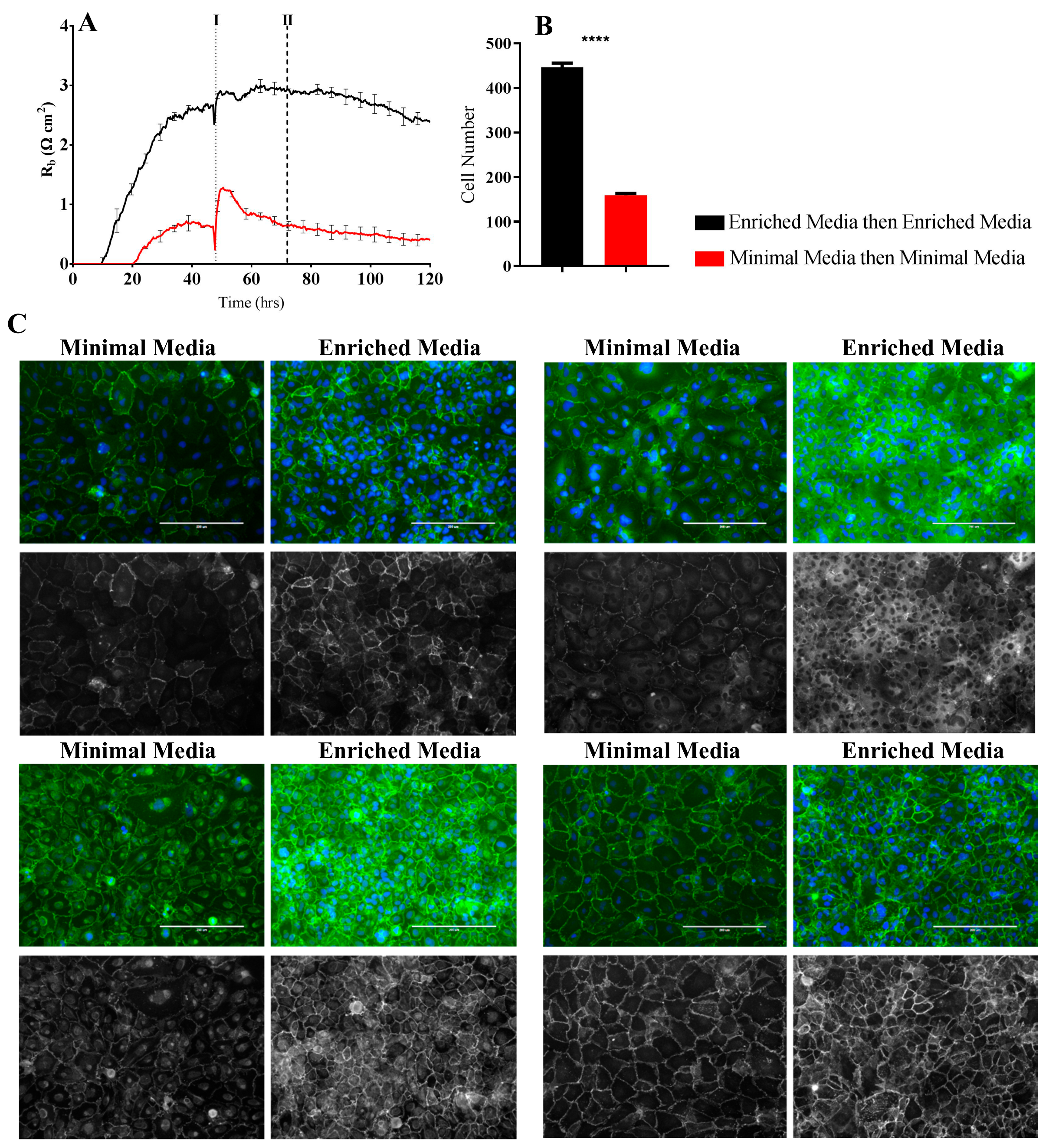
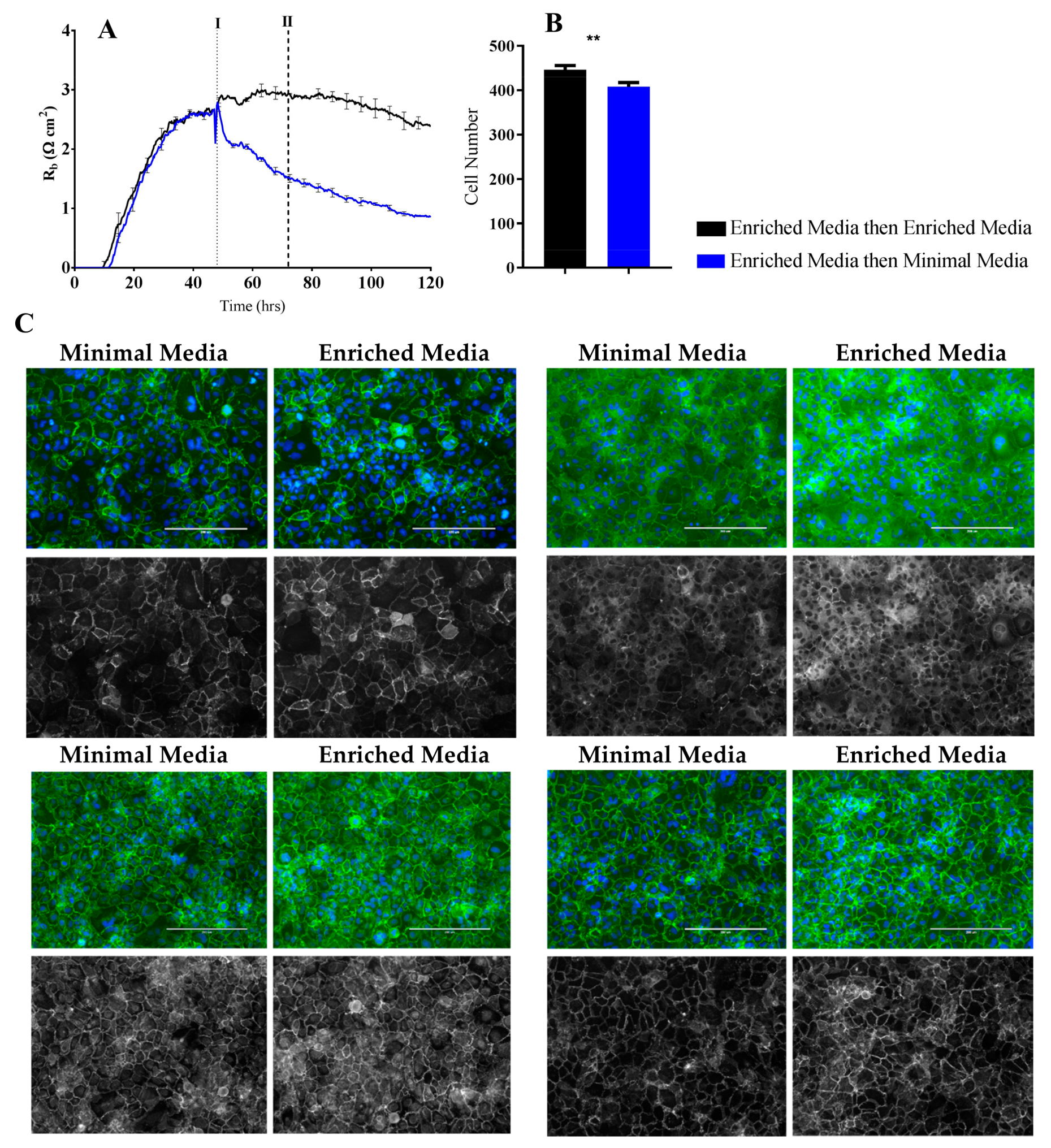
Endothelial cells were seeded onto collagen-I-coated 96-well plates at 20,000 cells per well in either 100 µL Enriched Media or Minimal Media. Immunocytochemistry (ICC) was carried out as companion experiments to ECIS and cells were grown until a barrier had formed as per ECIS recordings. At the point of barrier formation, all media was removed and replaced with each respective test media type. For CD144, Zonula occludens-1 (ZO-1), β-catenin, and α-catenin immunolabelling cells were prefixed in 2% paraformaldehyde (PFA) for 2 min and then fixed in 4% PFA for 10 min. PFA was aspirated and cells were rinsed with PBS prior to permeabilisation with 0.1% Triton-X100 in PBS (PBST) for 10 min. Cells were then washed three times for 10 min in PBS and then blocked with 1% BSA in PBS for 45 min. Following blocking, the cells were washed thrice with PBST and incubated with anti-CD144 (cat# sc-9989, Santa Cruz, Dallas, Texas, USA, 1:200), anti-ZO-1 (cat# 33-9100, ThermoFisher, Waltham, MA, USA, 1:200), anti-β-catenin (cat# 13-8400, ThermoFisher, Waltham, MA, USA, 1:250), or anti-α-catenin (cat# 13-9700, ThermoFisher, Waltham, MA, USA, 1:250) antibodies for 1 h at room temperature. Cells were washed in PBST three times and were incubated with Alexa Fluor 488 conjugated secondary antibody (Goat anti-Mouse, cat# A-11001, Invitrogen, Carlsbad, CA, USA, 1:400) and Hoechst 33342 nuclei stain (R37165, Invitrogen, Carlsbad, CA, 1:10,000) for 1 h at room temperature. Cells were washed for a final three times in PBST and imaged on EVOS FL Auto Imaging System (Invitrogen, Carlsbad, CA, USA) to acquire wide-field images of GFP and DAPI channels. Images were merged using Image J software. Cell-count analysis was carried out using Image J software of Hoechst stained nuclei.
5. Discussion
Endothelial cells line all of the blood and lymphatic structures throughout the body. They are phenotypically highly variable across tissues where they form selective barriers, the strength of which also varies across tissues [
13]. A major area of endothelial biology is understanding how these barriers are formed and regulated in both health and disease [
14]. In this paper, we highlight the power of ECIS biosensor technology to measure and model multiple aspects of brain endothelial barrier formation in a real-time autonomous manner. These aspects include determining (I) when the barrier has formed; (II) when the barrier is at its strongest; (III) temporal stability of the barrier; and (IV) as demonstrated here, the culture conditions required to produce a strong stable barrier. Importantly, we highlight the sensitivity of the technology to detect real-time changes occurring within the paracellular space (defined as R
b), which we can attribute to molecular changes in the expression of specific junctional proteins.
ECIS technology is an impedance biosensor capable of applying a very low alternating current at multiple frequencies (from 10 to 10
5 Hz) [
3]. The two direct measurements that are produced are resistance and capacitance. In terms of endothelial cells, impedance measured across multiple frequencies can be mathematically modelled using the ECIS software to derive several important parameters which collectively relate to the overall barrier formation of the cells. These are R
b (resistance of the paracellular space), α (resistance due to basal adhesion), and C
m (capacitance of the cell membrane). ECIS is the only impedance biosensor to use multifrequency AC. This is a major distinguishing factor in comparison to xCELLigence technology from ACEA, which generates the data from a single AC frequency. Therefore, although xCELLigence can measure these parameters collectively (total resistance), it cannot model which aspect of barrier function changes [
7].
Understanding which components of the barrier are contributing to the barrier strength and the resistance measured by ECIS is very important. Consider the scenario where endothelial cells have very strong basal adhesion but weak paracellular adhesion, which would be expected of permeable lymphatic endothelial cells [
15]. This will produce a high resistance, primarily derived from the basal resistance. However, endothelial cells of the blood brain barrier (BBB) have a strong paracellular barrier governed by junctional proteins, as well as strong basal adhesion. Therefore, for in vitro studies, it is highly pertinent to know: (I) when the endothelial barrier has formed, which is not simply when a monolayer is visible; (II) the stability and duration of that the barrier; and (III) the strength of both the paracellular and basal barriers.
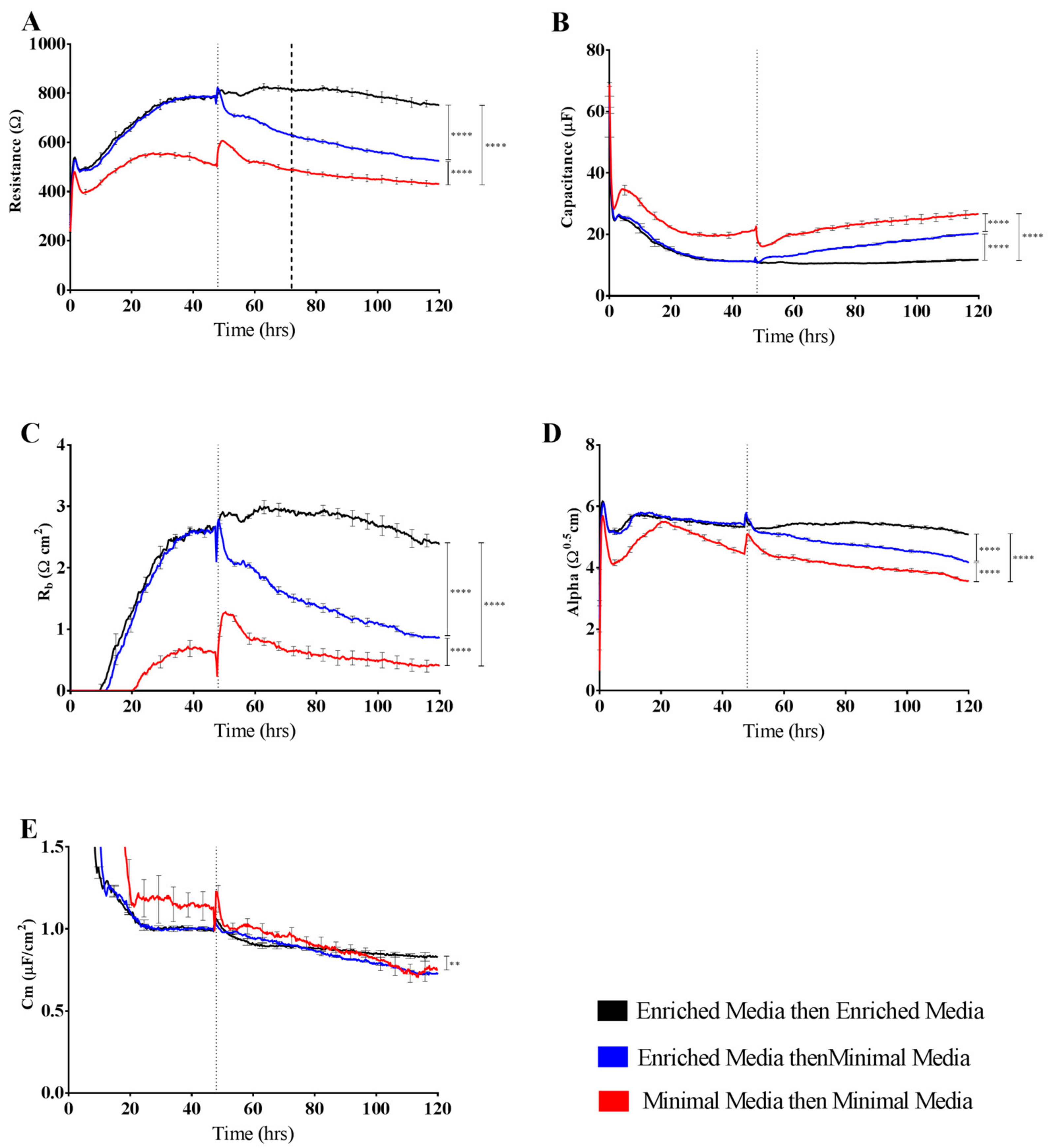
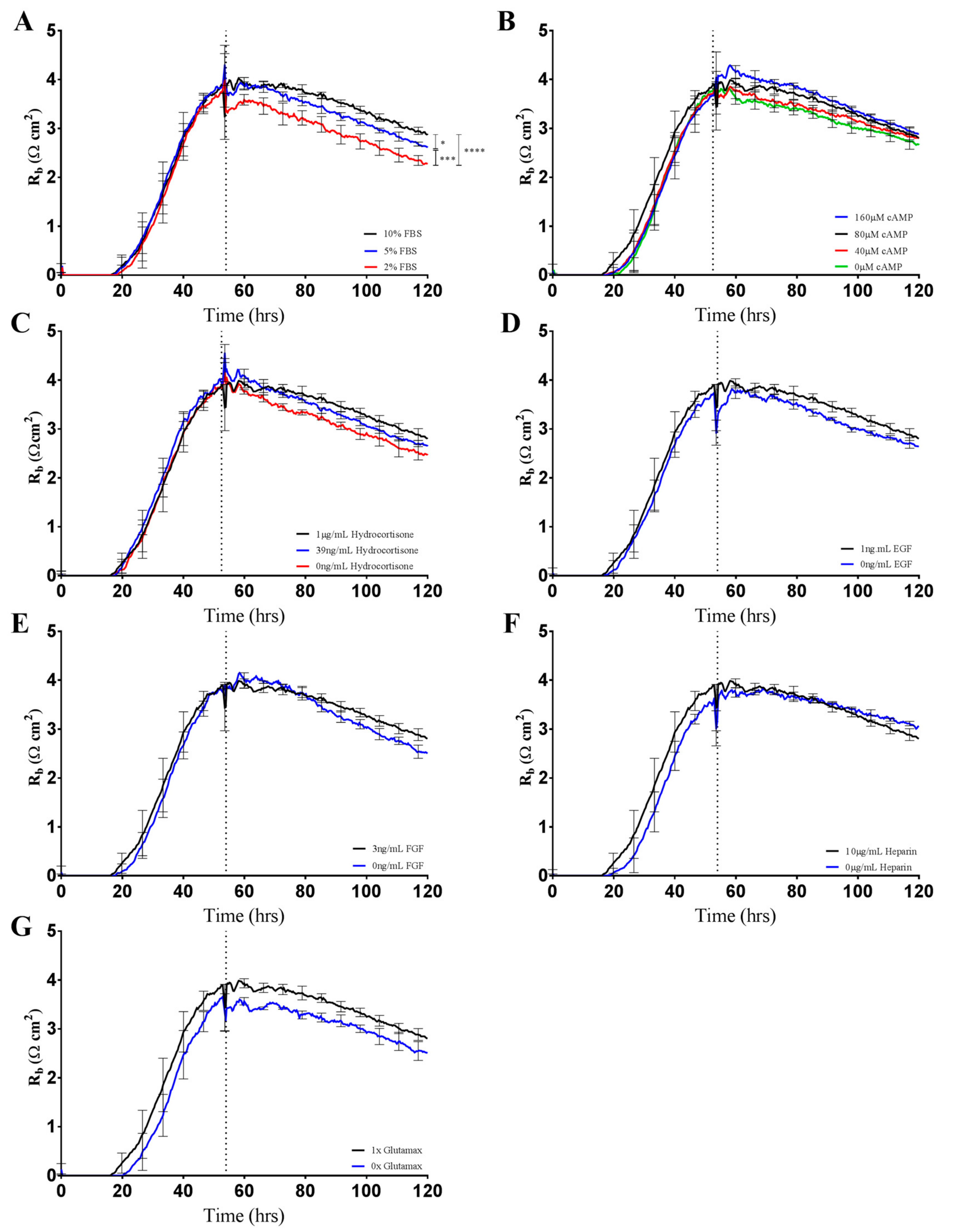
It is predictable that the formation and maintenance of a strong endothelial barrier in vitro will be dependent on the optimal use of growth factors and supplements in the cell-culture media and that this will vary depending on cell type and source. We hypothesised that our immortalised brain endothelial cells cultured in media enriched with various growth factors and 10% FBS (Enriched Media) would exhibit a stronger and more stable barrier than that produced from a less enriched media (Minimal Media). This was indeed observed where Enriched Media achieved an endothelial resistance of ~800 Ω, whereas the Minimal Media only achieved a maximal resistance of ~500 Ω (
Figure 4A). It was unclear as to which components of the barrier were influenced to a greater extent by the Enriched Media or which factors within the media caused the greater effect. This is where the power of ECIS really excelled, as it clearly demonstrates that the paracellular barrier (R
b) is dramatically weaker in the cells grown in the Minimal Media. The endothelial cells grown in the Enriched Media also begin to form their paracellular barrier faster and achieve a stronger barrier, which is maintained for at least 50–60 h (stronger for longer). The basal adhesion (α) is weaker for the cells grown in the Minimal Media, but this effect is far less dramatic than that of R
b. The real-time power of ECIS is highlighted in
Figure 4, where the brain endothelial cells cultured in the Enriched Media are switched to the Minimal Media. In this scenario, there is an immediate and continuous loss of paracellular barrier strength and basal adhesion. This not only highlights the importance of real-time measurements for observing immediate or acute changes in barrier function but reveals that the maintenance of the stronger barrier is a highly active process continually requiring the nutrients present in the Enriched Media [
16,
17].
Typically, optimisation of BBB endothelial in vitro models has relied on transendothelial electrical resistance (TEER) as an indication of increased paracellular resistance [
10,
11,
18,
19]. In particular, epithelial voltohmmeter (EVOM) has been extensively employed as a means of quantitatively describing barrier integrity based on ohmic resistance. The extensive limitations of EVOM are reviewed elsewhere [
20], however it is worth noting that EVOM TEER measurements provide measurements only based on the overall resistive properties of an endothelial cell monolayer grown on porous membranes. Consequently, EVOM recordings do not allow for the separation of paracellular and basal resistance, thus limiting the interpretation of subsequent TEER measurements [
20,
21]. ECIS, therefore, as a means of establishing the conditions required to generate a strong brain endothelial barrier, provides a more in-depth and relevant analysis of cell barrier properties (described by [
2]).
ECIS technology further allowed us to determine the contribution of each Enriched Media additive to the increased barrier resistance by brain endothelial cells (
Figure 5). Unexpectedly, serum concentration appeared to be the only additive that significantly altered endothelial barrier strength (
Figure 5A). Serum-dependent barrier function is well established throughout the literature [
10,
11,
18,
22,
23]. However, serum addition is frequently reported to negatively affect blood brain barrier integrity, with high concentrations of serum and the presence of growth factors within the culture medium reportedly capable of inhibiting the formation of brain endothelial tight junctions [
24]. In a unique study, Nitz et al. demonstrated that the disruption of tight junctions by serum was highly side specific, meaning that apically applied serum did not decrease barrier function to the same extent as basal administration [
24]. Physiologically, apical membranes of brain endothelium are exposed to serum at much higher concentrations compared to the basal compartments. The side-specific nature of serum-mediated junctional disruption likely reflects membrane-specific receptor–serum interactions at the basal surface that mediate vascular leakiness.
Measuring endothelial barrier resistance with ECIS allows impedance measurements to be acquired when cells are grown on a flat, nonporous surface coated with appropriate extracellular matrix proteins. Therefore, unlike typical EVOM measurements, where cells are required to be grown on porous filters, ECIS ensures serum is only exposed to the apical surface of brain endothelium, thus eliminating any potential for basal–serum interactions that possibly promote loss of barrier integrity. A recent systematic review of immortalised brain endothelial cell lines showed that out of 49 studies investigating endothelial barrier resistance, 48 used transwell-based systems [
25]. Most of these studies had serum present on both sides of the polarised endothelial cells. High serum levels on the basolateral aspect of the endothelium is not physiological and potentially may introduce a barrier artefact in some studies. There is also a need for a better understanding of side-specific serum effects on brain endothelial cells. ECIS technology using solid-state plates negates the potential limitations of transwell-based resistance measuring systems, making it an ideal tool for these studies. The 96-well ECIS plates also offer the capacity to assess a wide range of culture and growth conditions simultaneously, ideal for optimisation.
The ECIS modelling clearly shows that the paracellular barrier is stronger for longer in the Enriched Media. Such a sizable difference should be reflected at the molecular level in the expression of key junctional proteins that govern the strength of the paracellular space [
26]. Others have recommended that changes occurring in R
b measurements should be critically assessed by immunocytochemical staining of the endothelial cell monolayer [
3] to validate and confirm the modelled parameters. We therefore assessed the expression and location of key junctional proteins (CD144, ZO-1, β-catenin, and α-catenin) which are known to positively contribute to paracellular barrier strength (
Figure 6 and
Figure 7). There was a striking and obvious difference in the location and expression level of each of these key proteins after 72 h of culture in the respective media. Overall, the expression is higher in the Enriched Media, with pronounced junctional staining evident for CD144, α-catenin, and β-catenin. There is also a greater number of cells present, which means there is a greater volume of paracellular space present under these conditions (
Figure 6B). This, in theory, provides more path for current flow between the cells. Therefore, in order for the R
b to be higher in the Enriched conditions, it means that the barrier must be tighter. In the scenario, where the media was switched from Enriched to Minimal Media, there is a substantial loss of R
b. There is also a concordant reduction in the expression of CD144, ZO-1, α-catenin, and β-catenin. The most obvious differences of these are for CD144 and ZO-1. Although these are subtler than the differences where the cells are grown entirely in the Minimal Media, the changes are consistent with the modelled differences in R
b.
We conclude that ECIS provides a powerful multiparametric method of assessing endothelial barrier function, and that this solid-substrate impedance sensing technology has multiple advantages over other technologies. As our study requires an understanding of the changes occurring in both the paracellular space and contribution from the basal adhesion, other technologies such as EVOM or xCELLigence RTCA would not have sufficed. The utility of ECIS has provided us with a powerful means of assessing multiple aspects of endothelial barrier formation, maintenance, stability, and the optimal conditions required for the generation of a strong barrier (stronger for longer). Importantly, the investigation highlighted the importance of multifrequency impedance sensing in monitoring endothelial barrier function, with emphasis placed on the biological context of ECIS technology.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






