
Development of Functional Fluorescent Molecular Probes for the Detection of Biological Substances

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
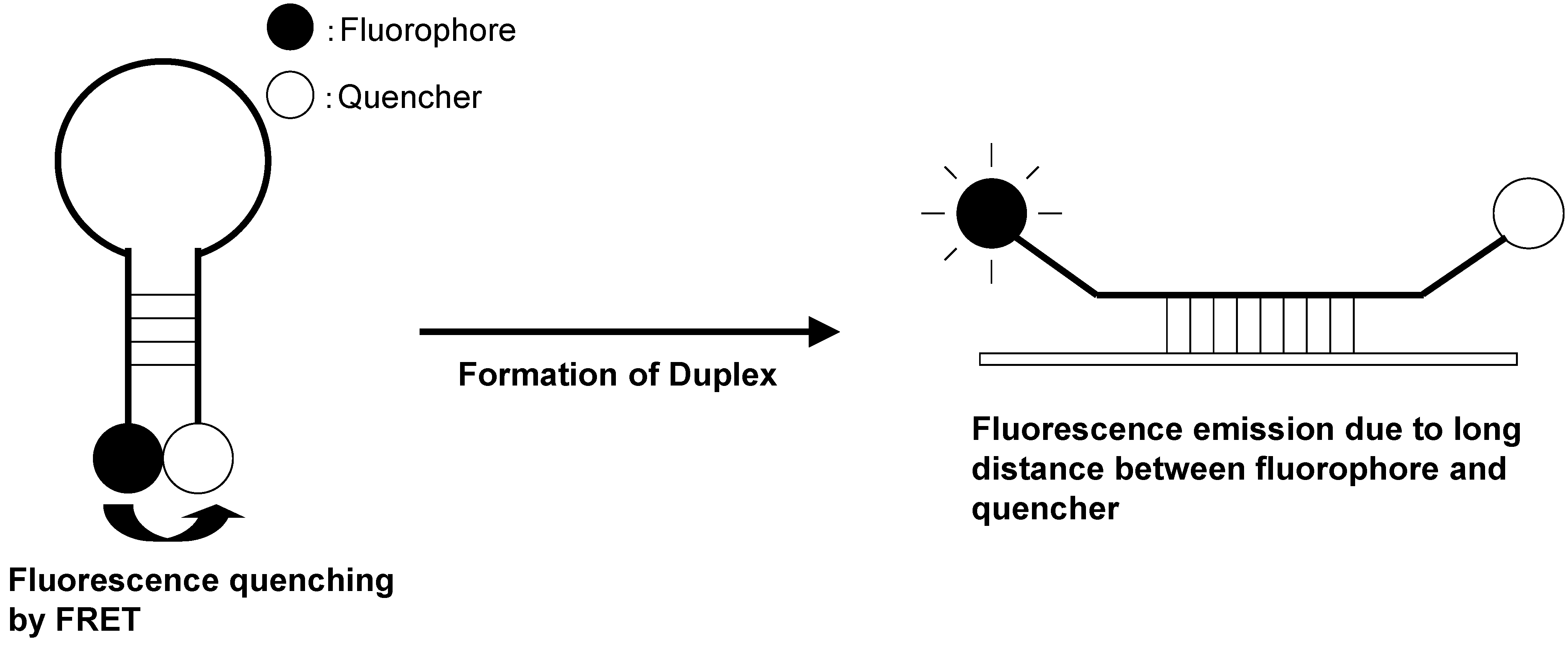{kind=link}
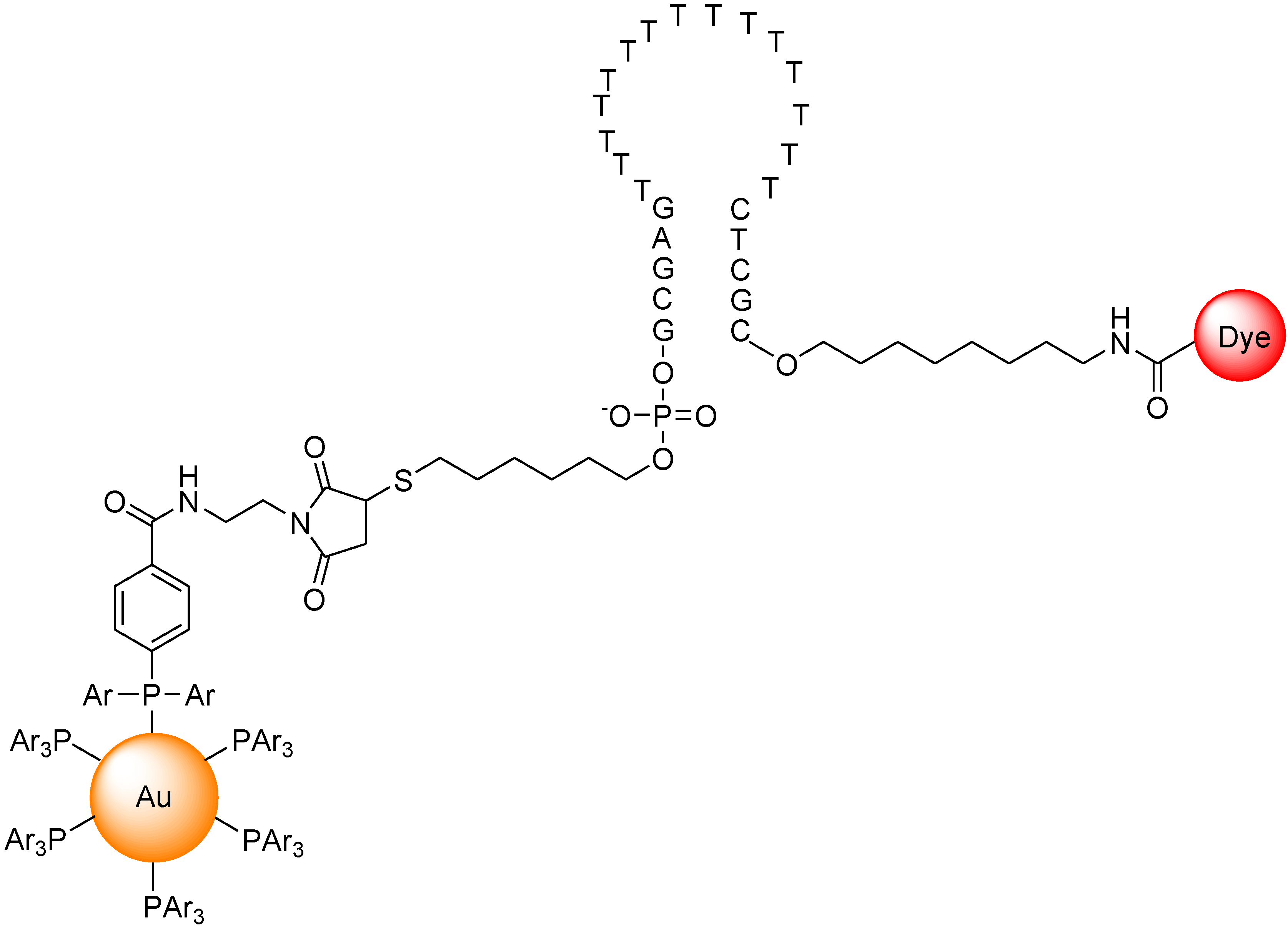{kind=link}
Abstract
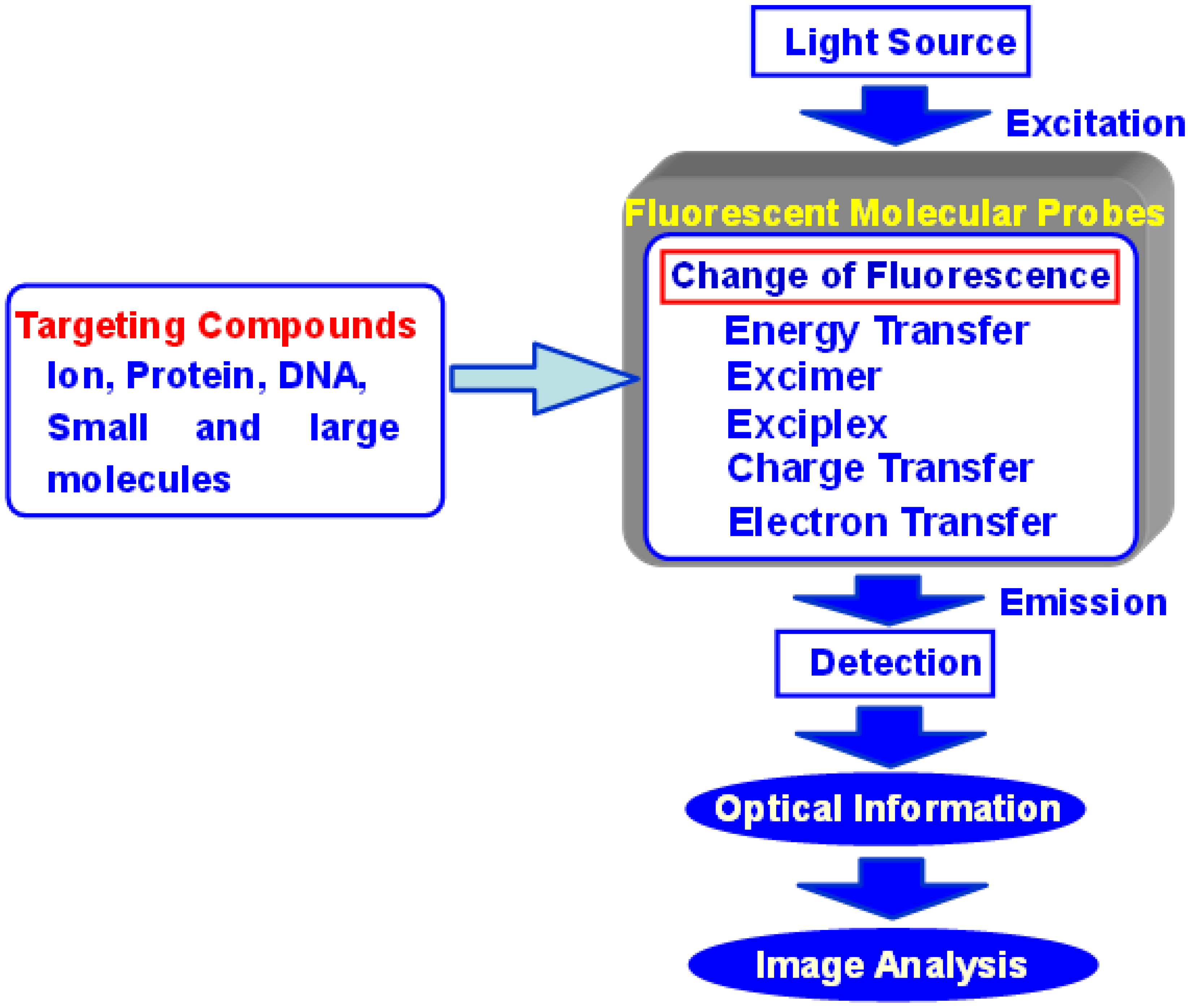
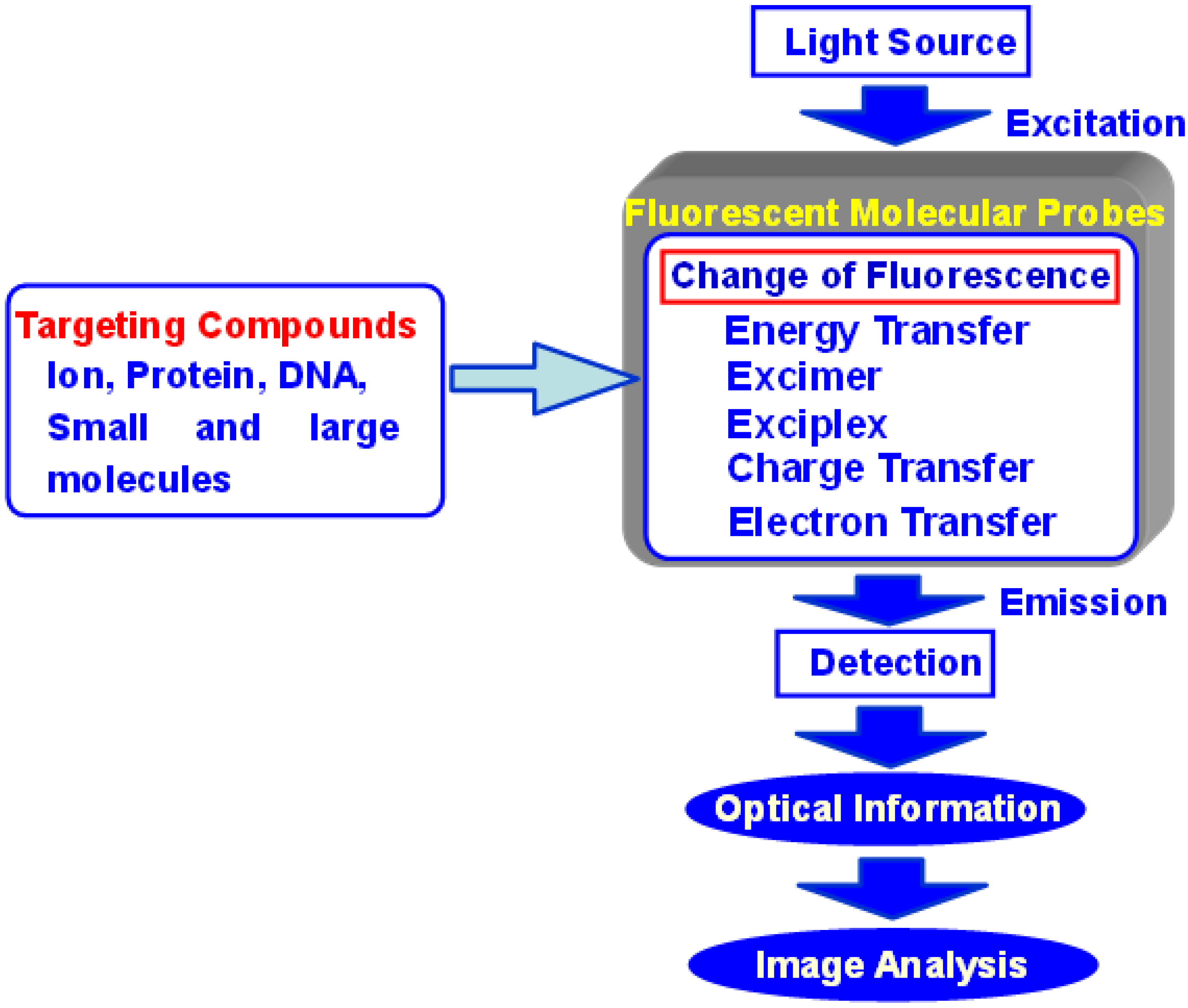
:1. Introduction

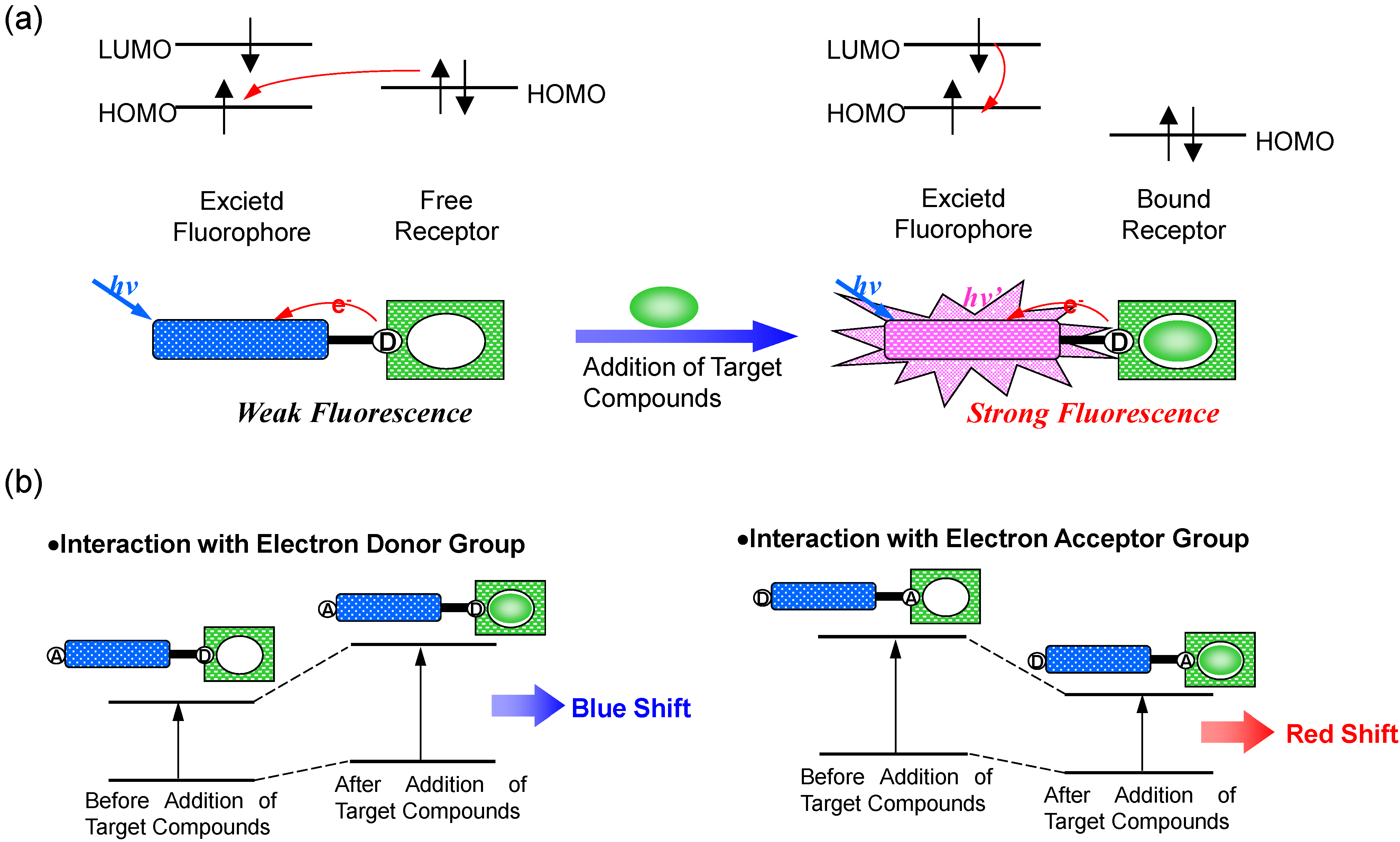
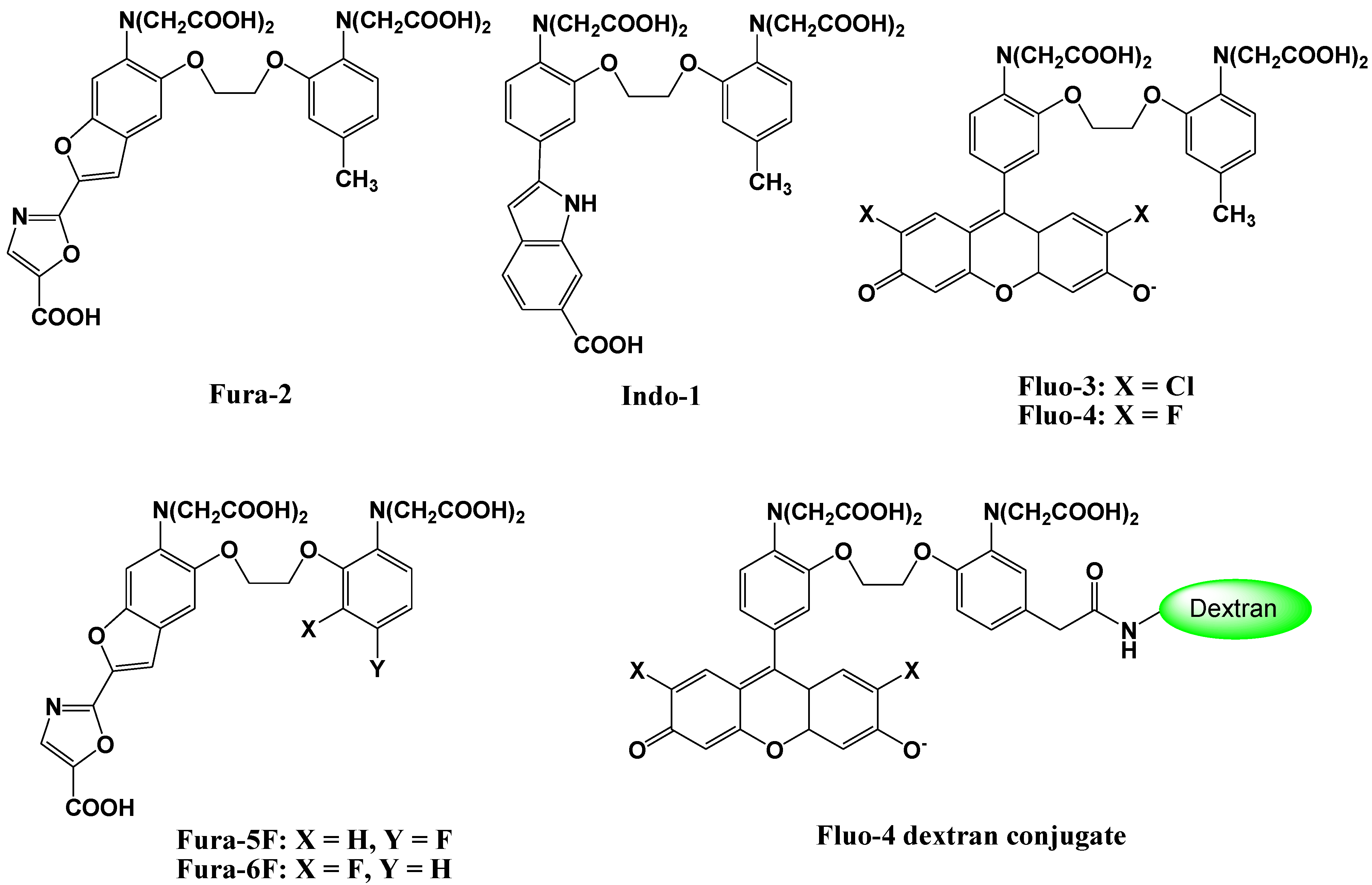
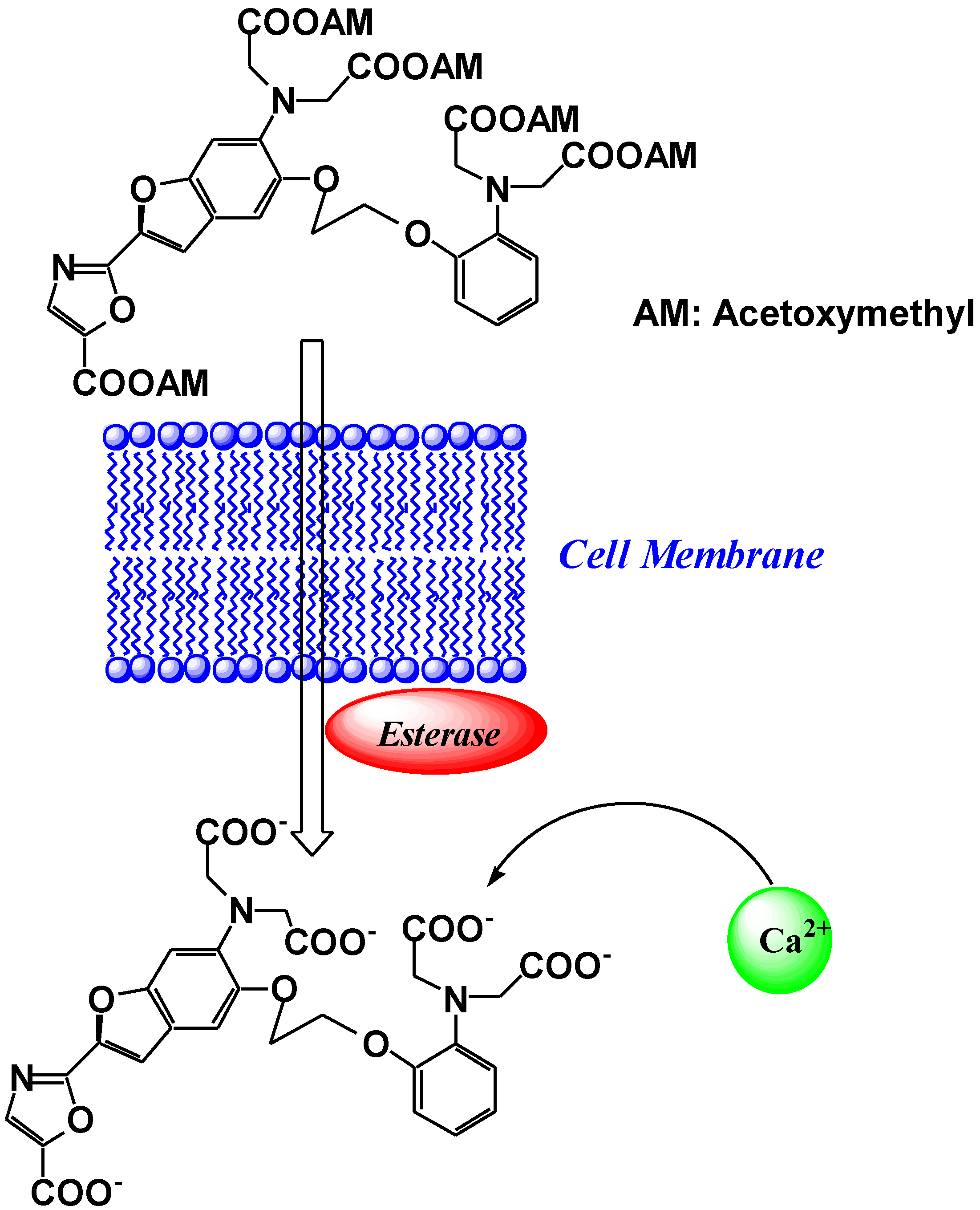
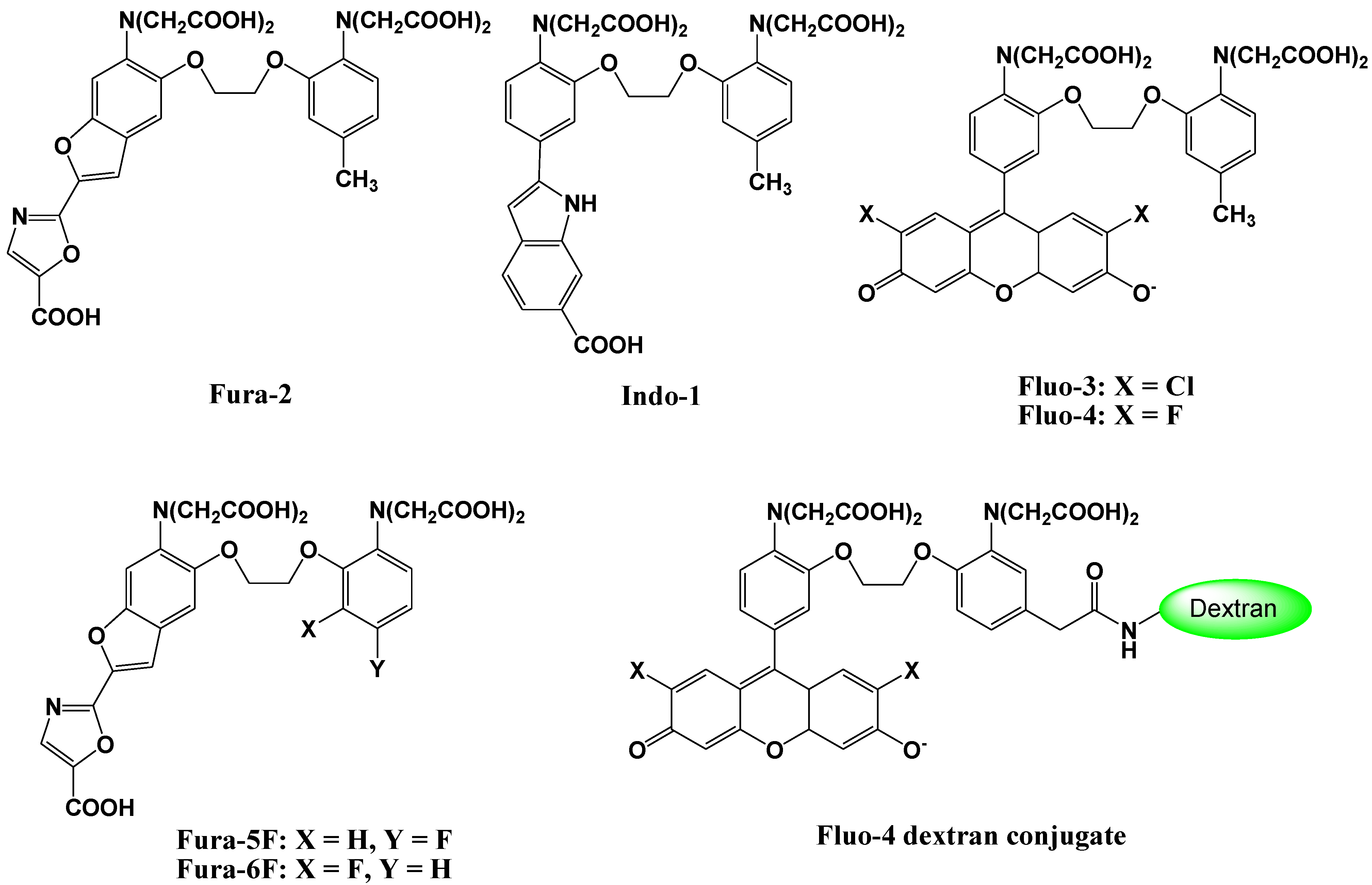
2. Ca2+ Selective Fluorescent Sensors
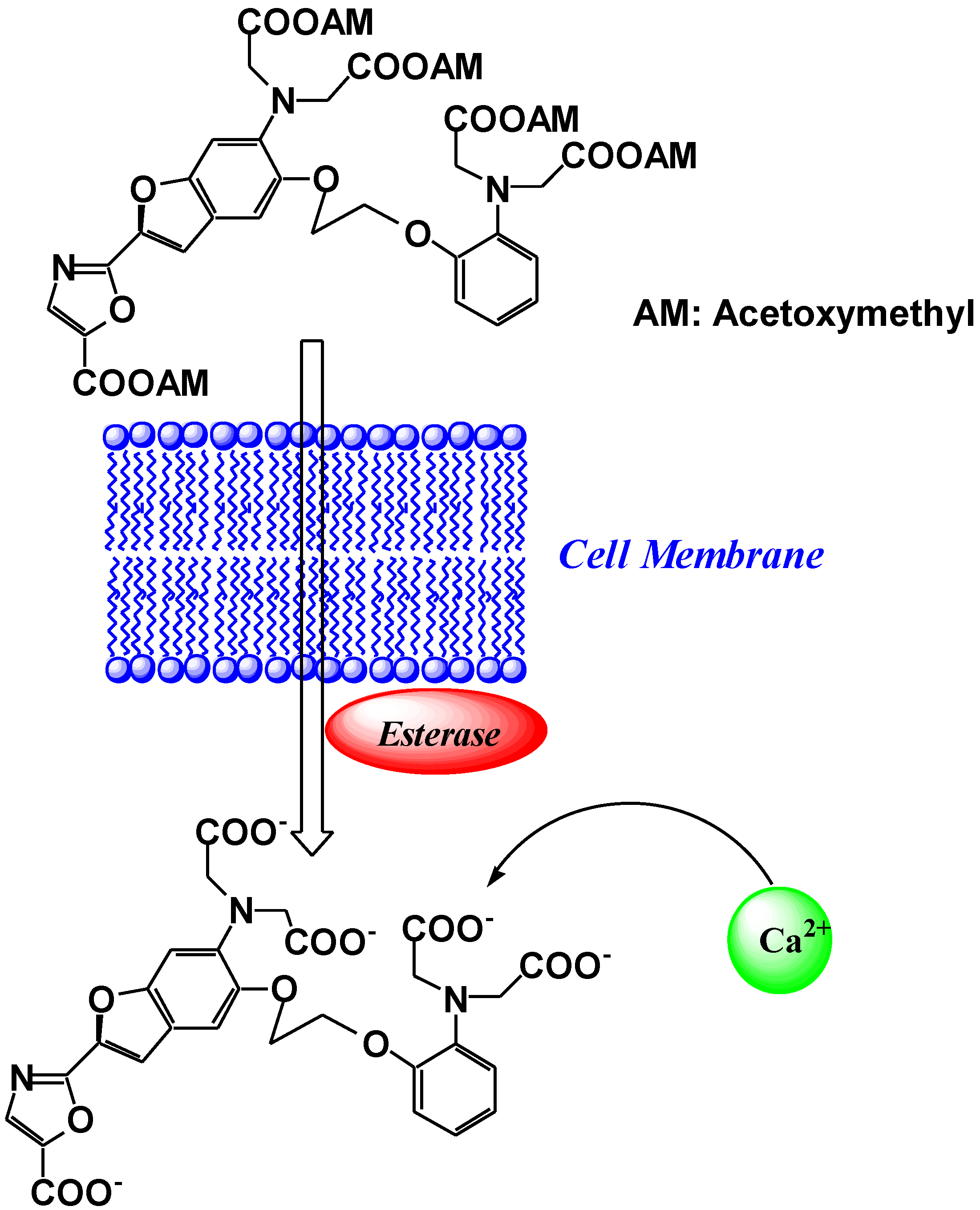
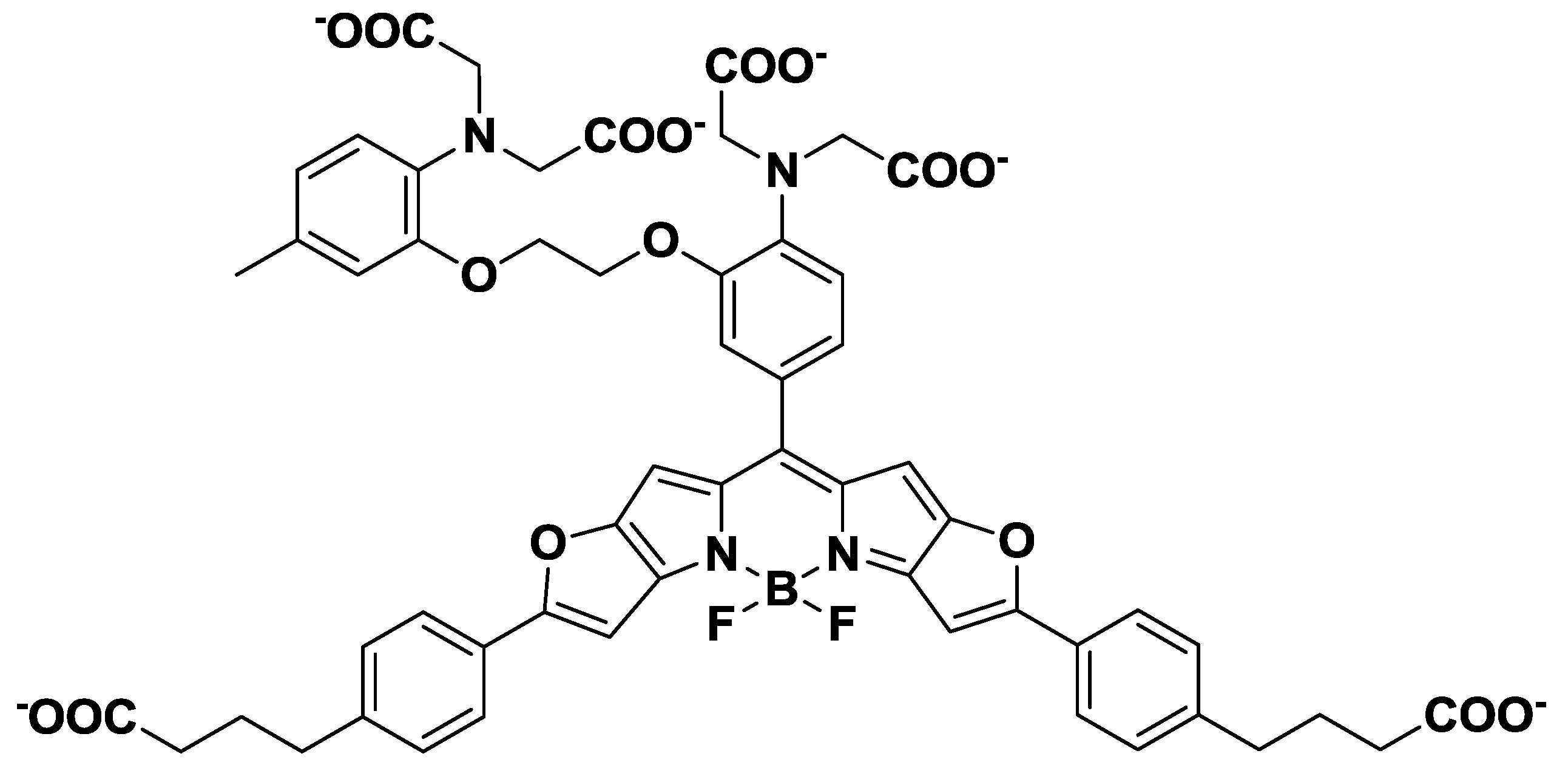
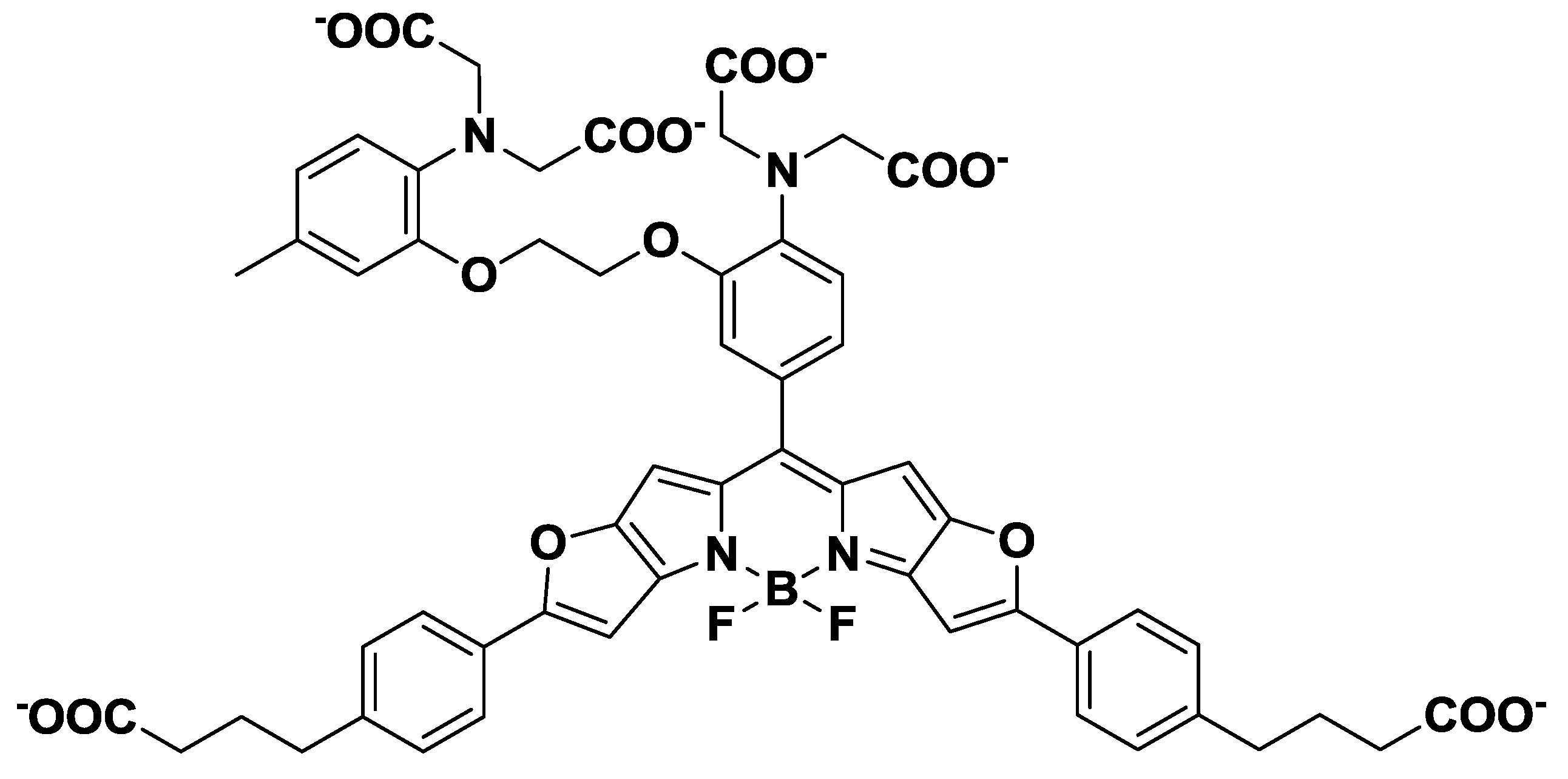
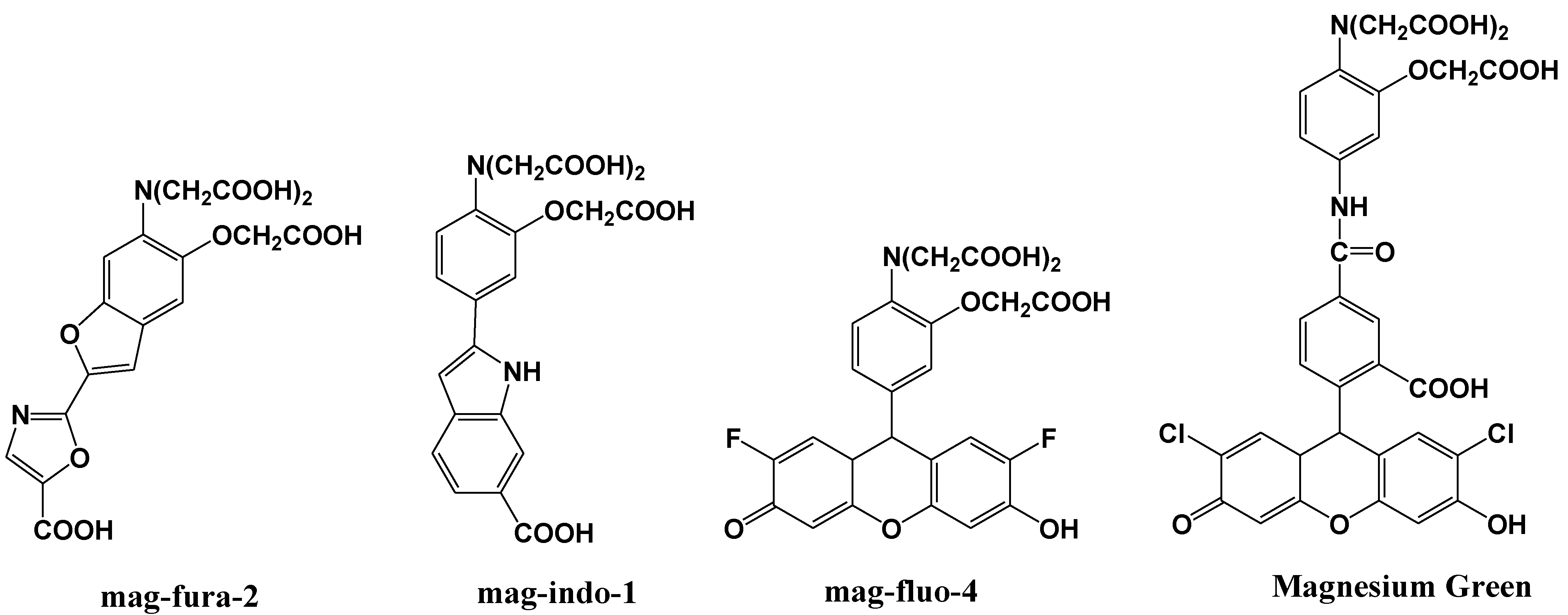
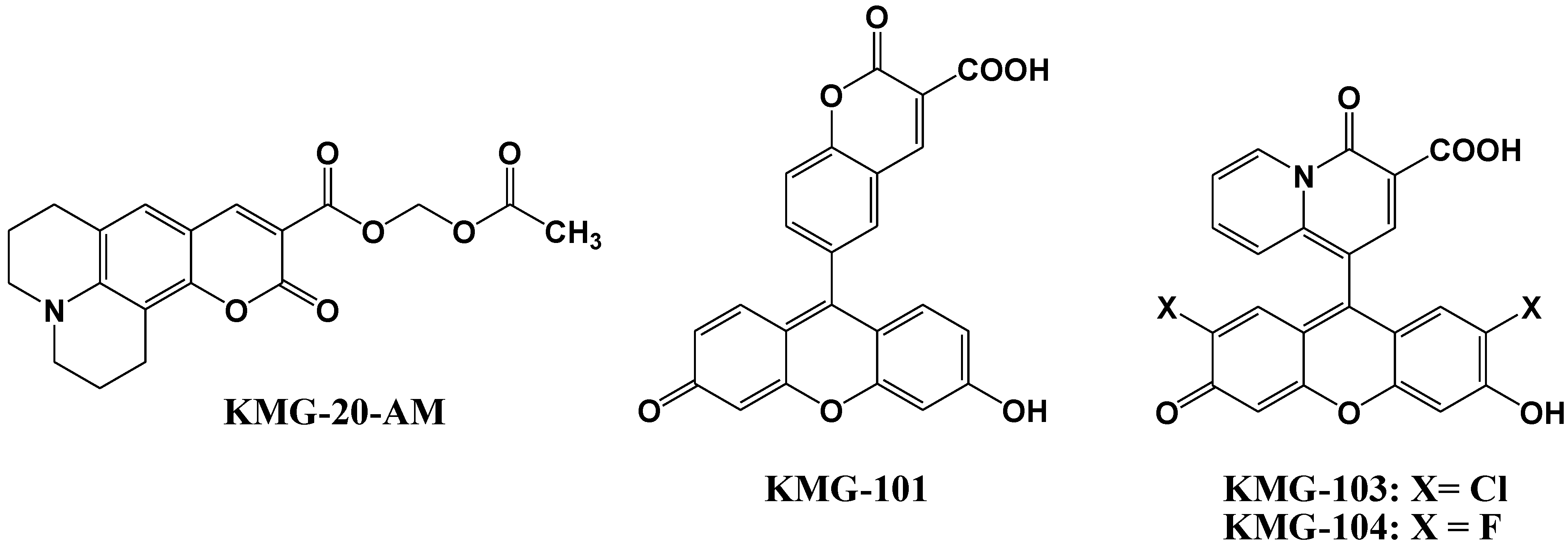
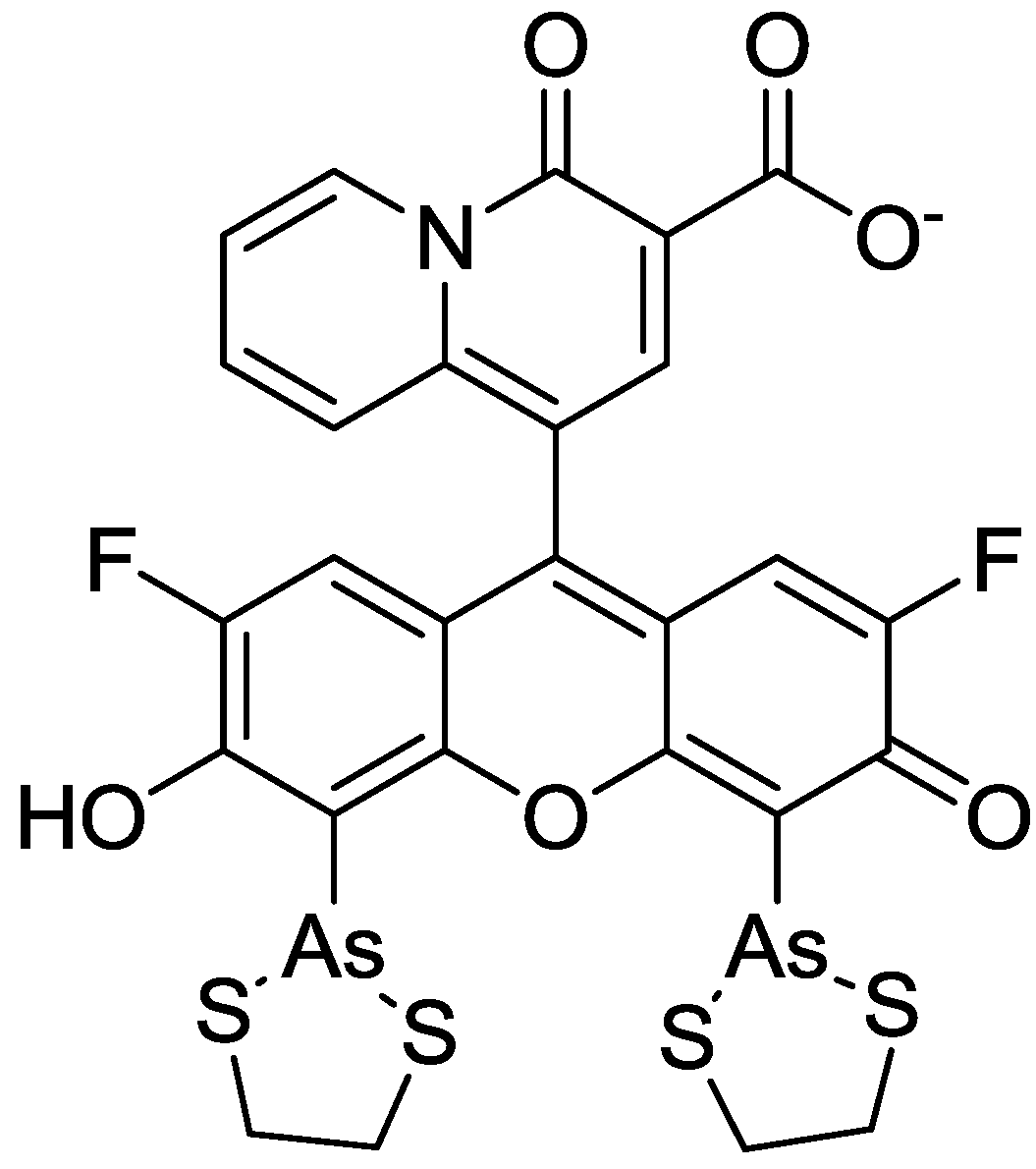
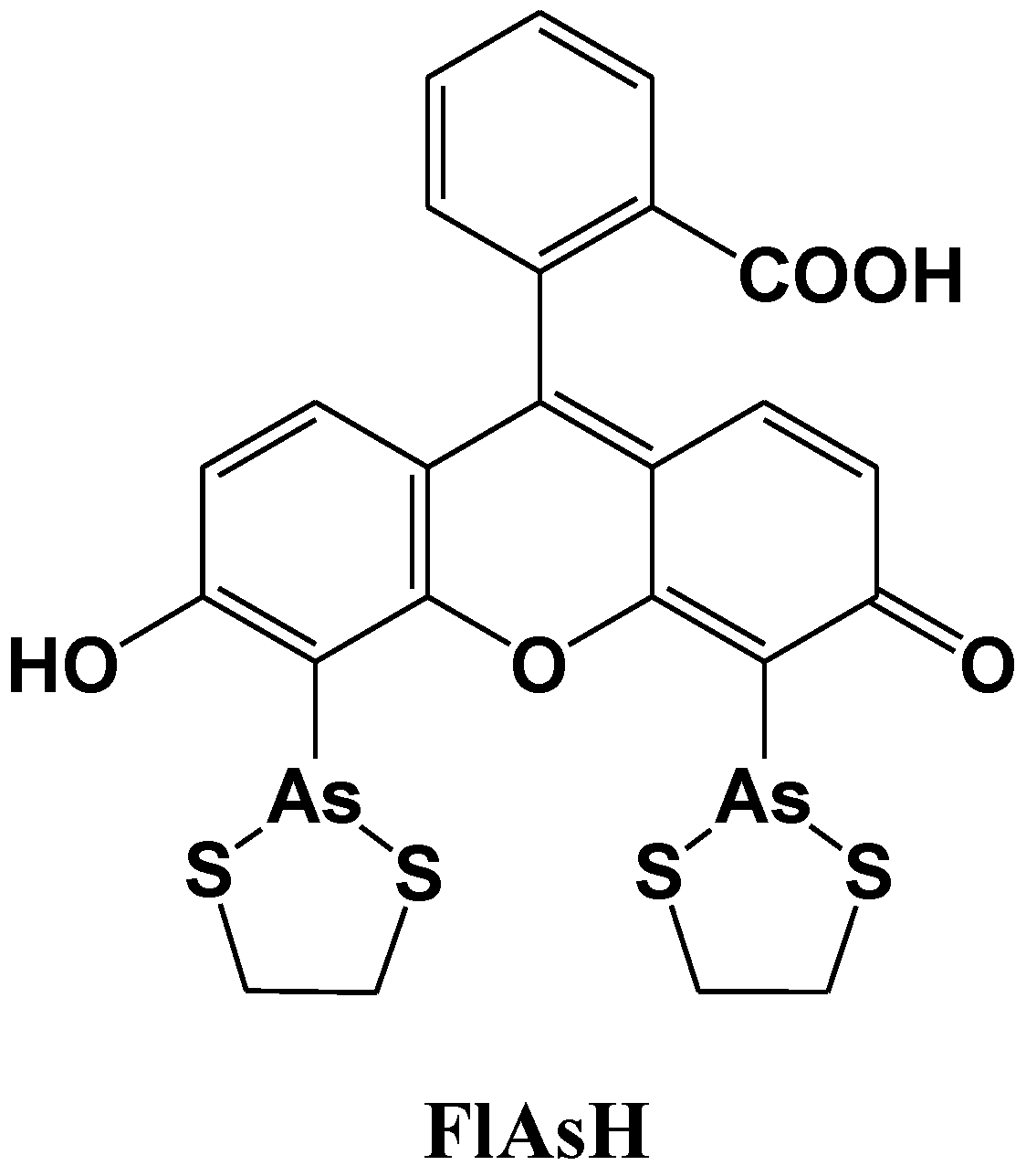
3. Mg2+ Selective Fluorescent Molecular Probes
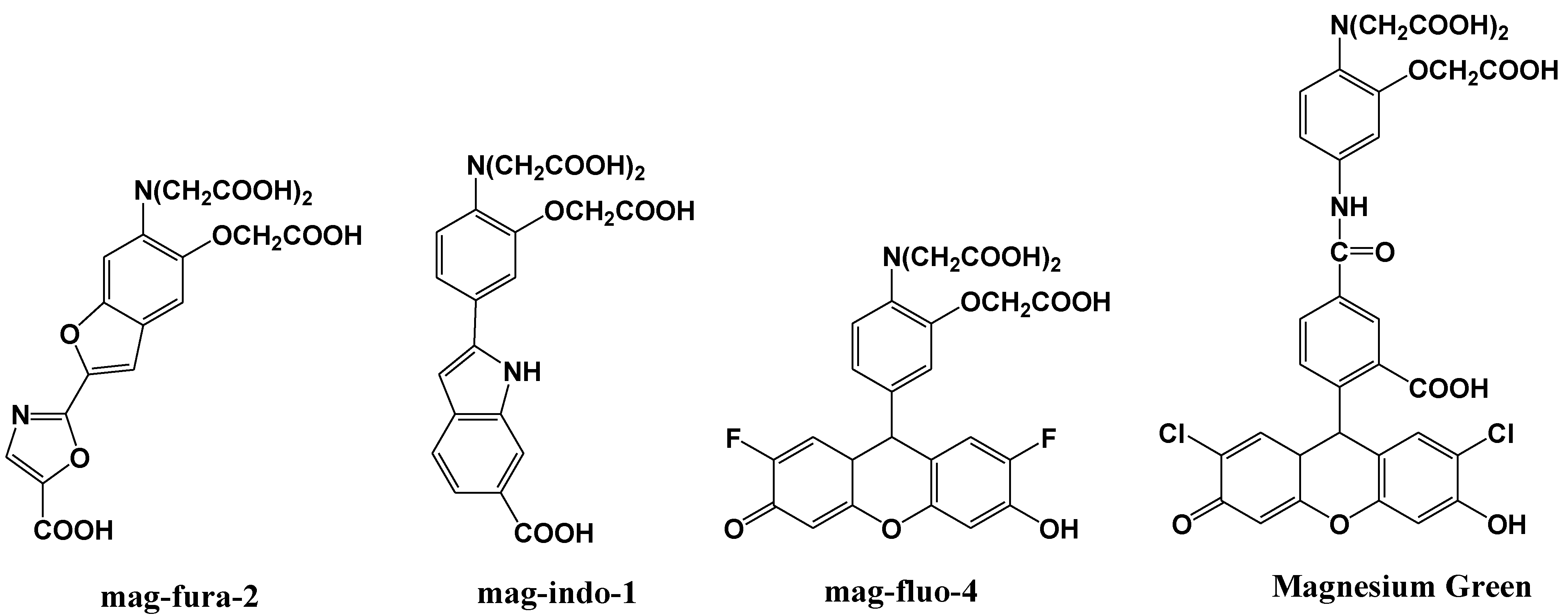
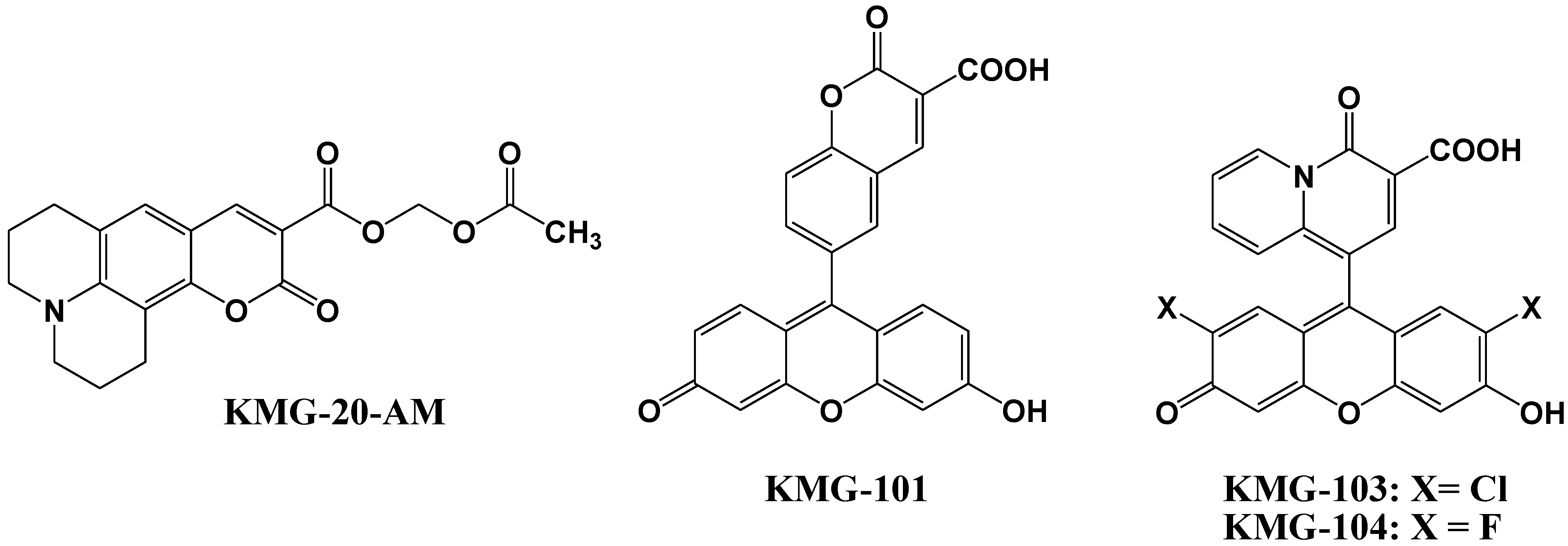
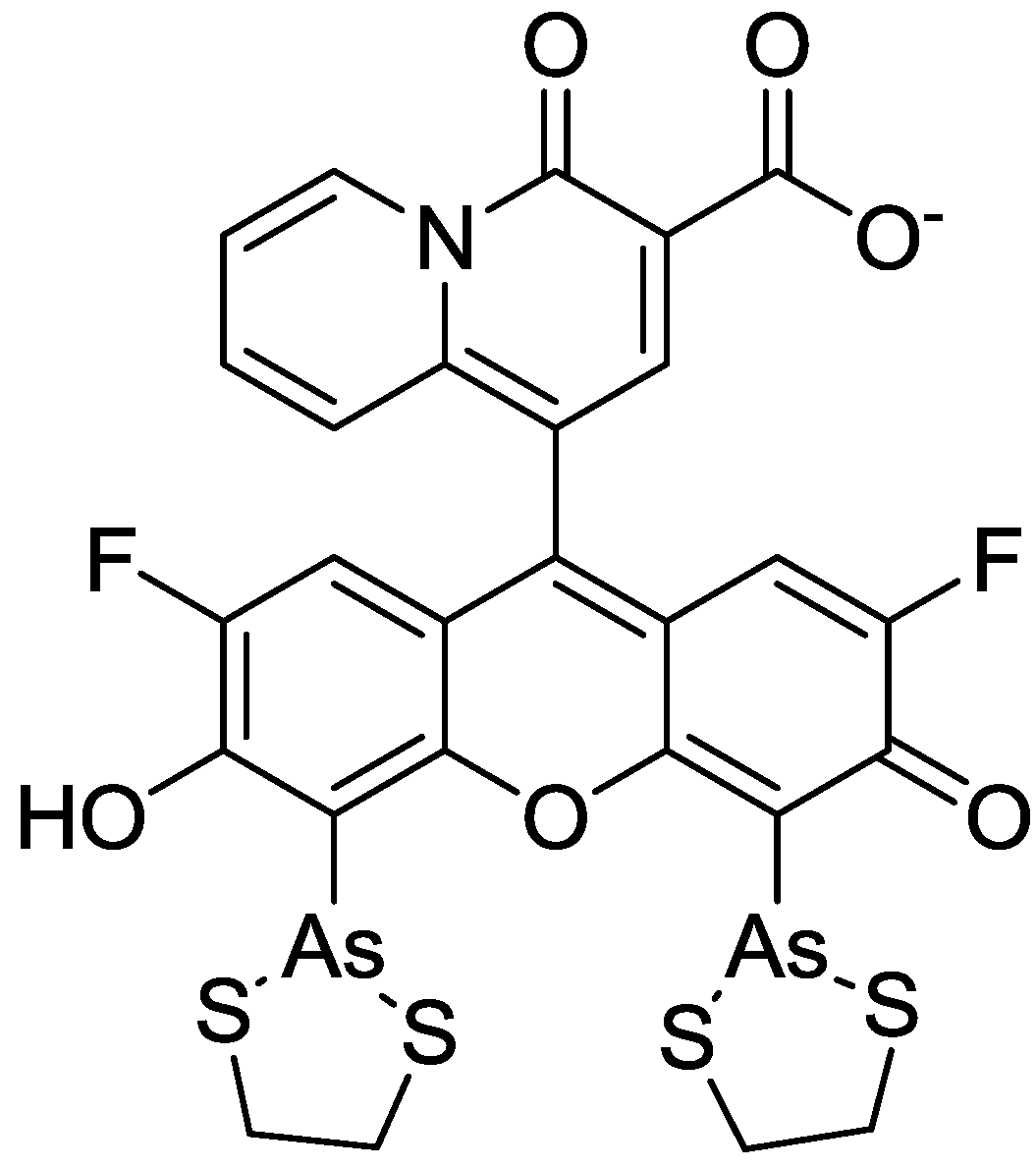
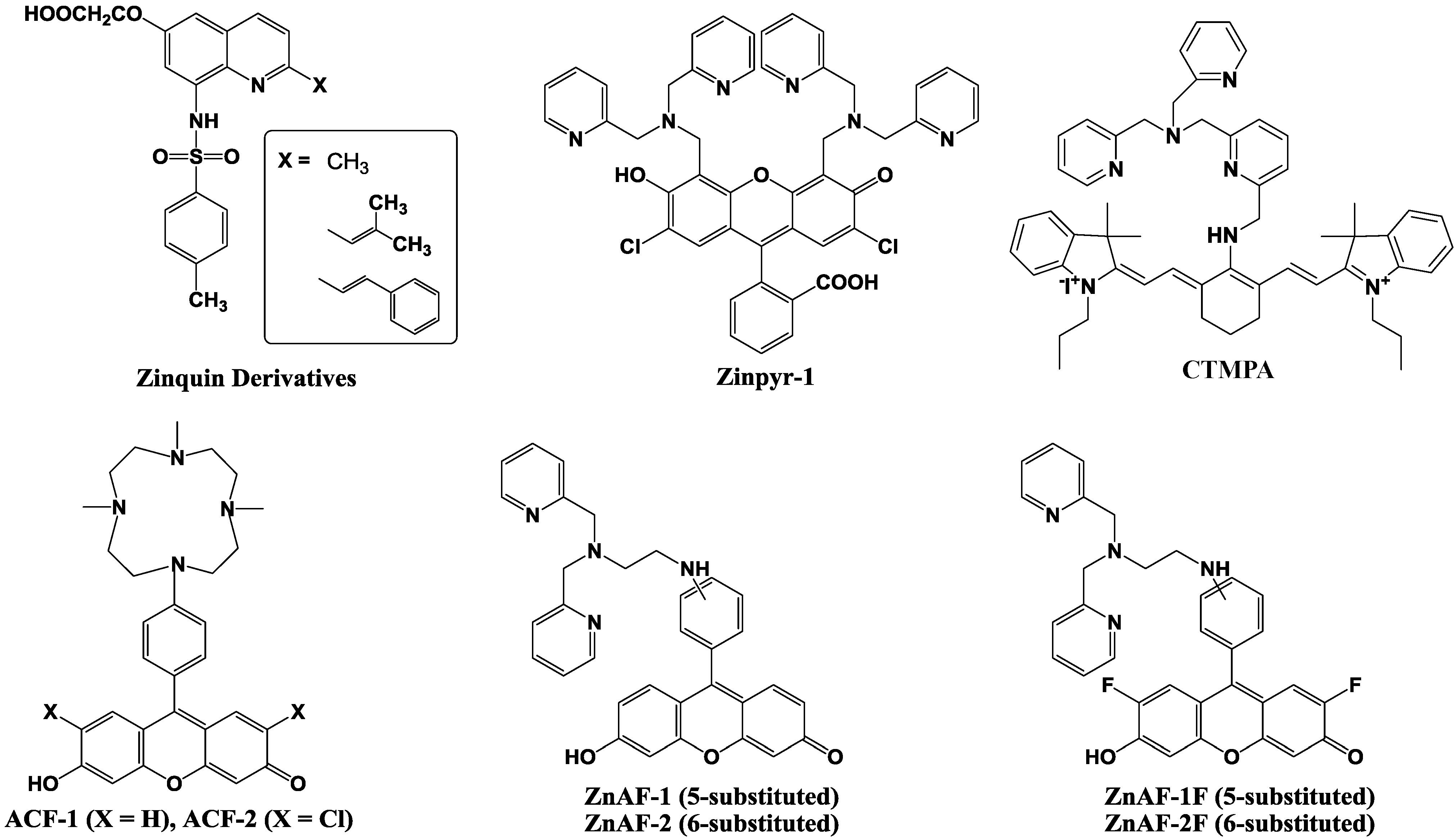
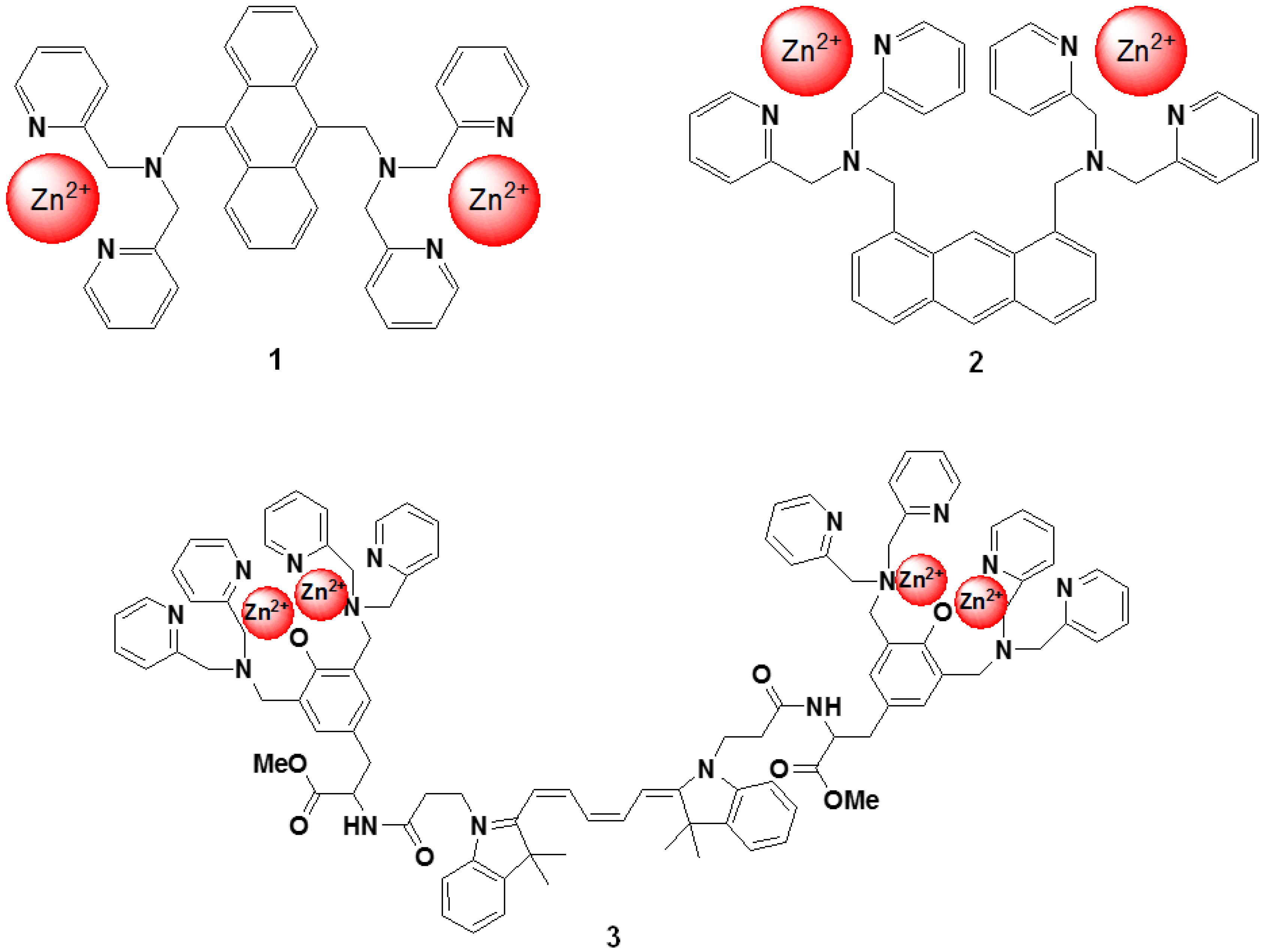
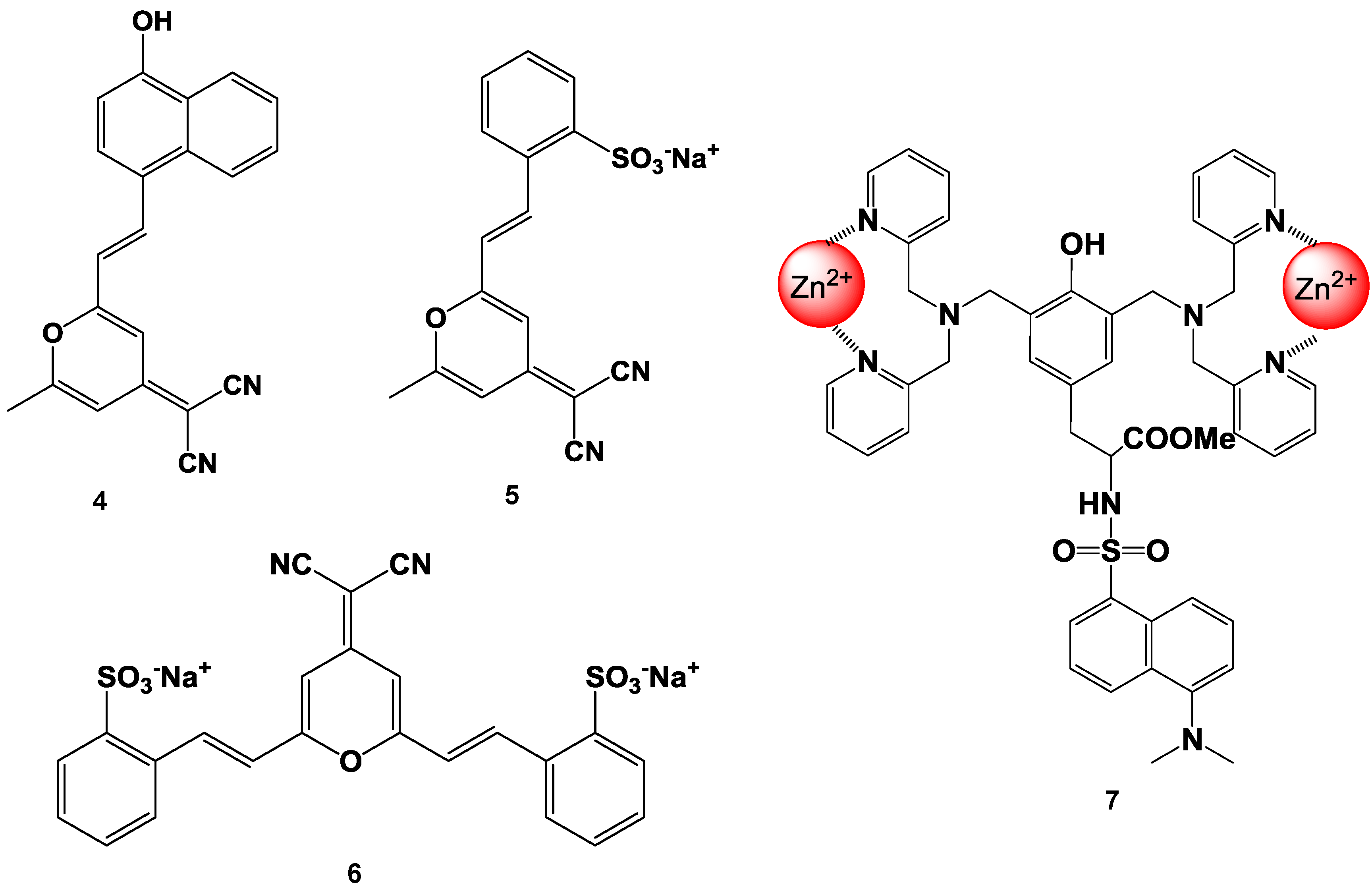
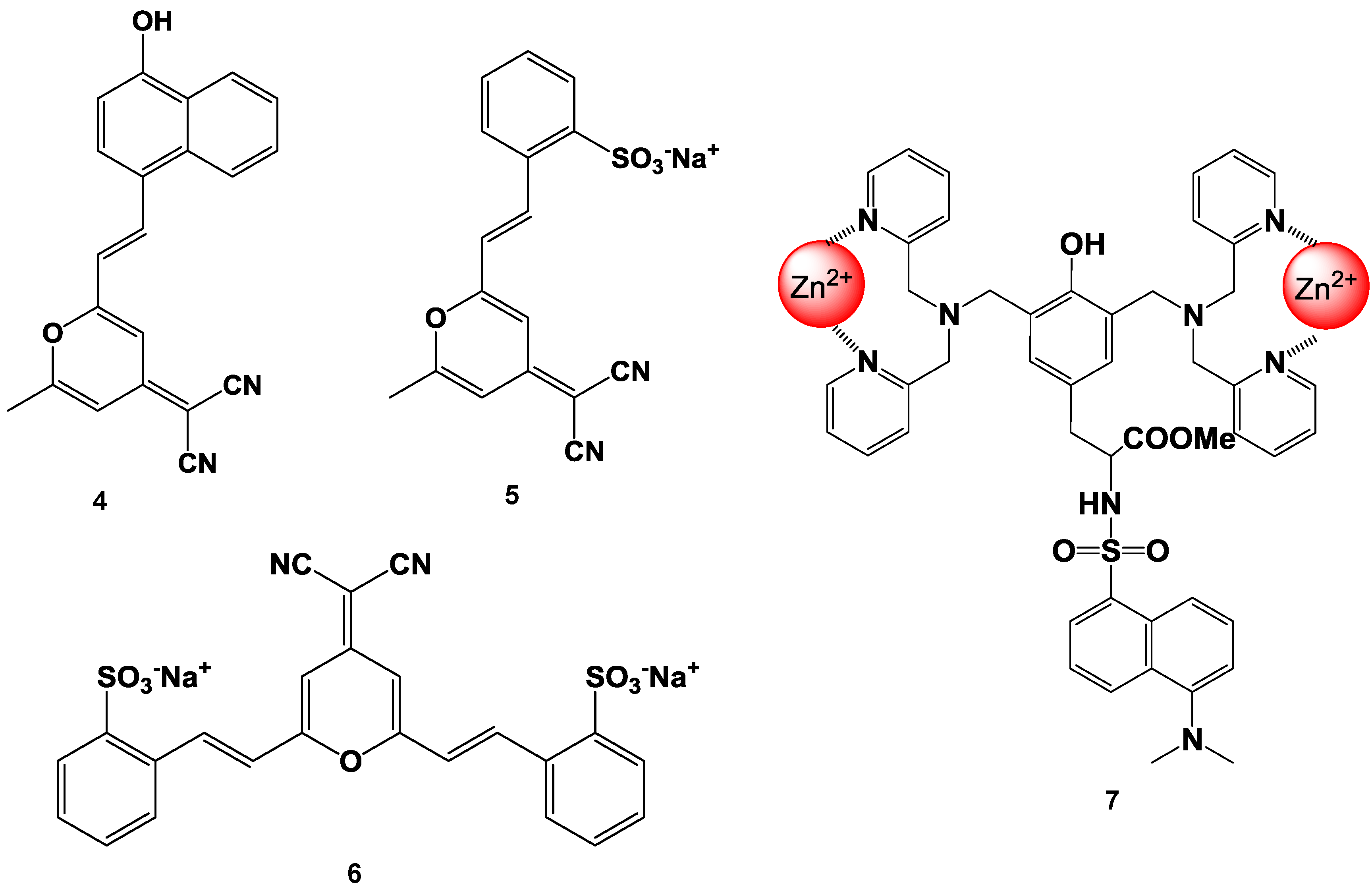
4. Zn2+ Selective Fluorescent Molecular Probes
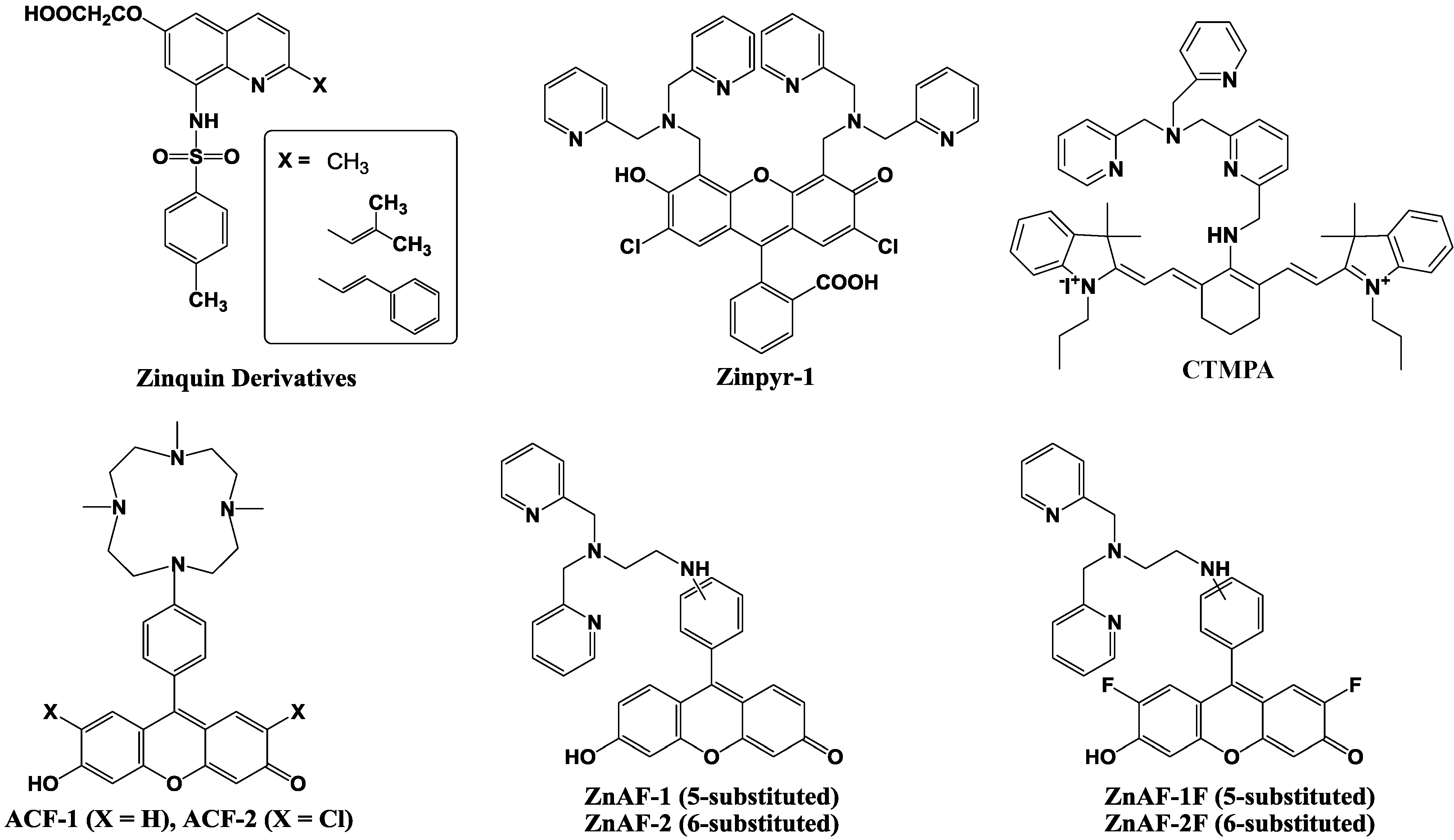
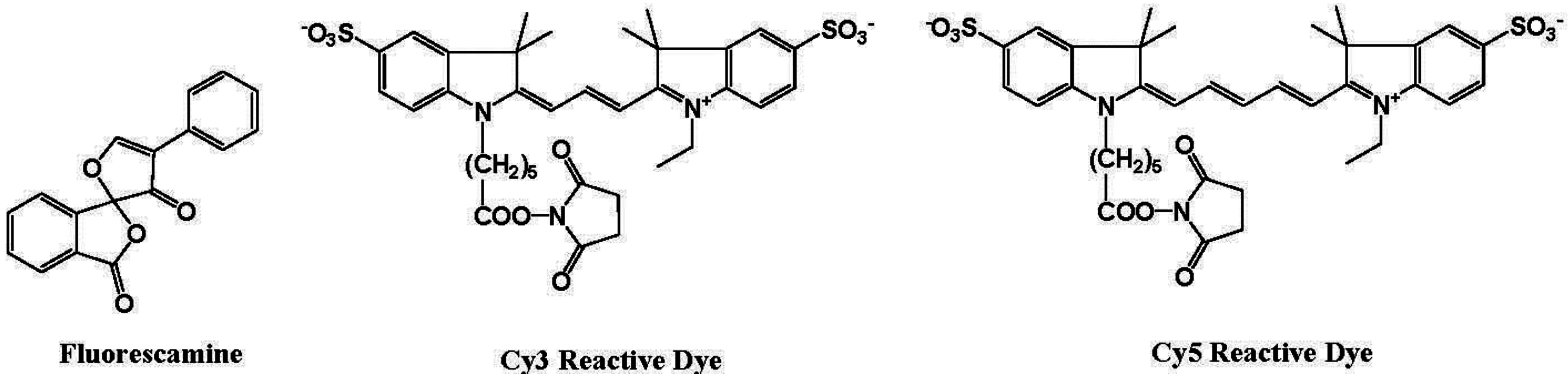
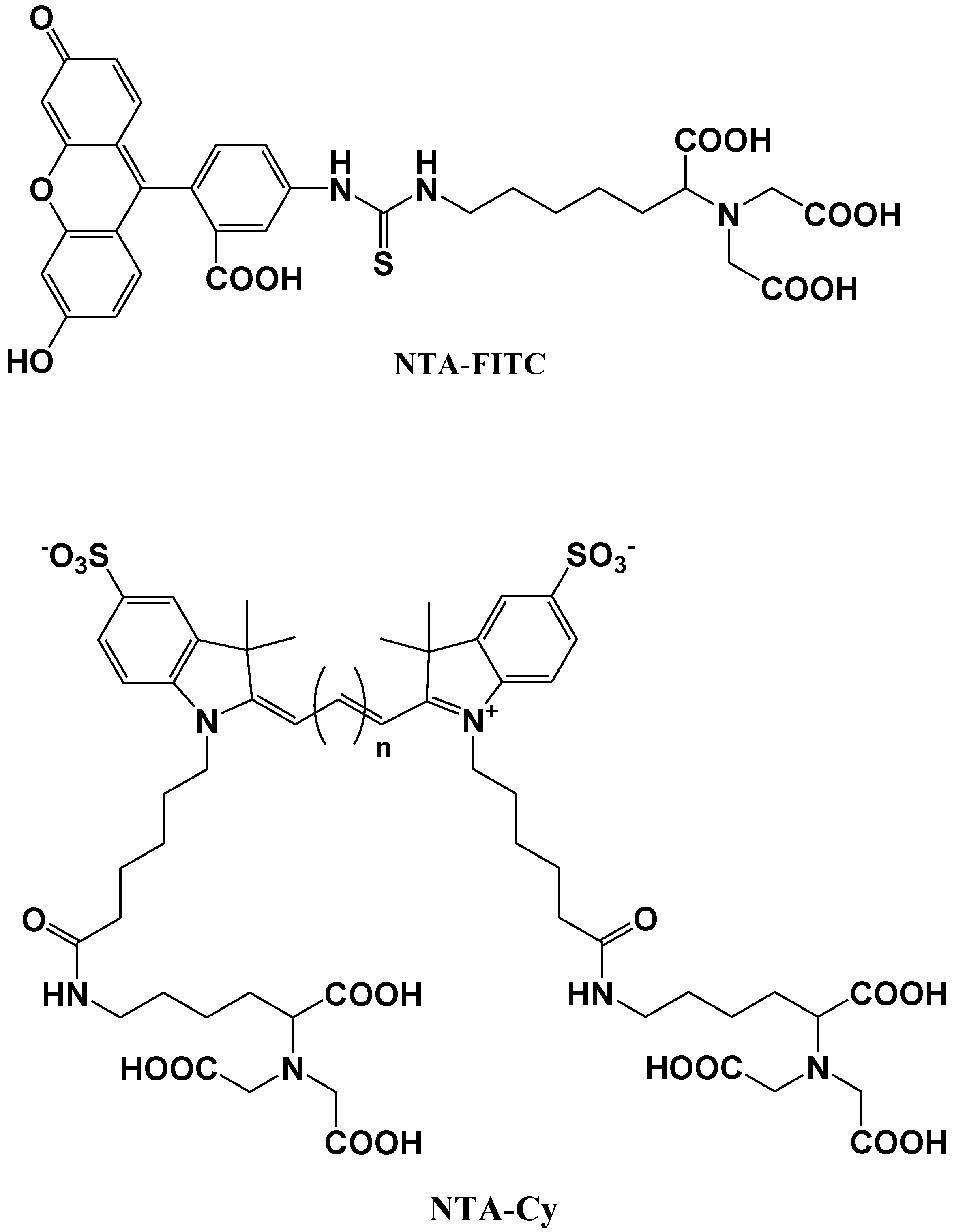
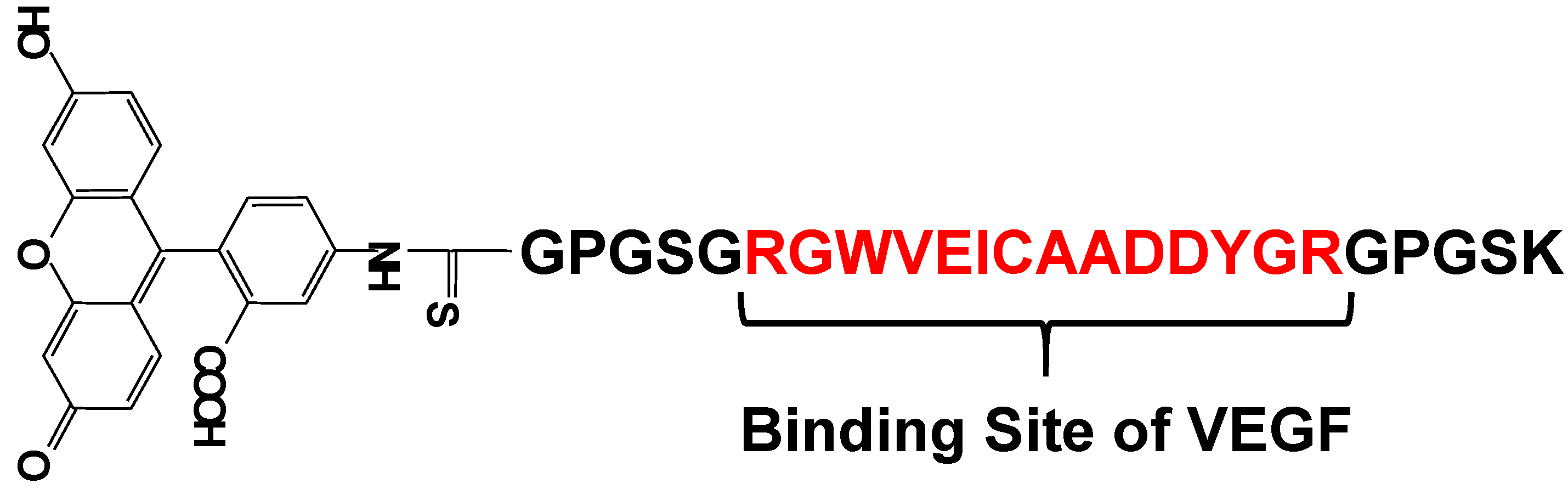
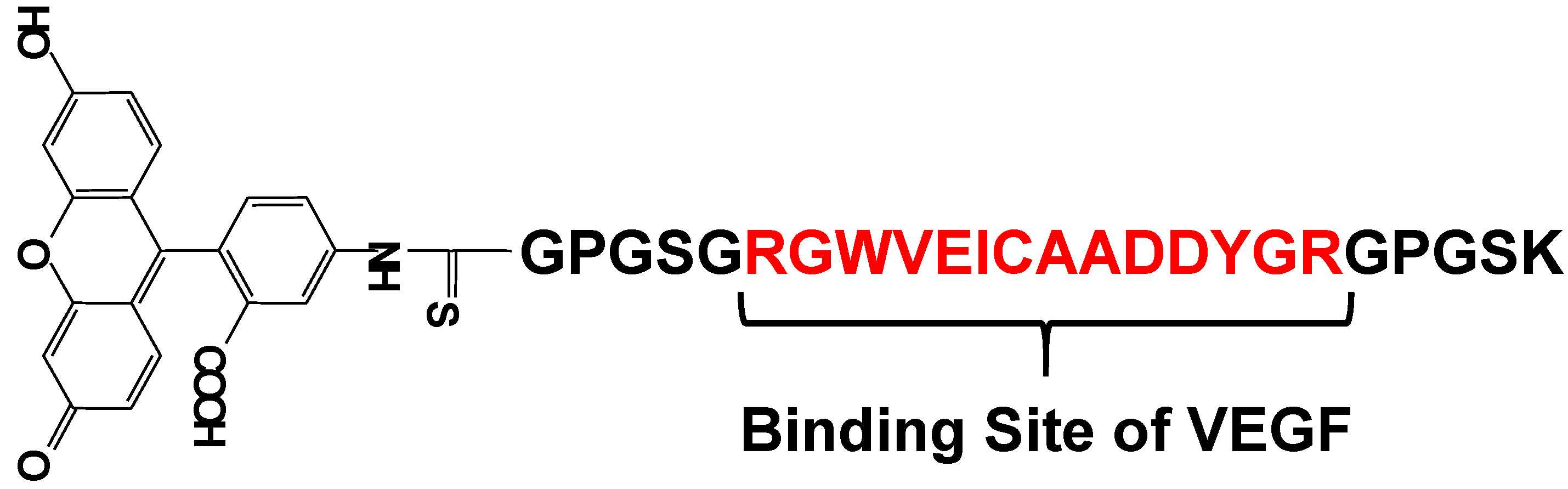
5. Protein Selective Detection Using Fluorescent Probes
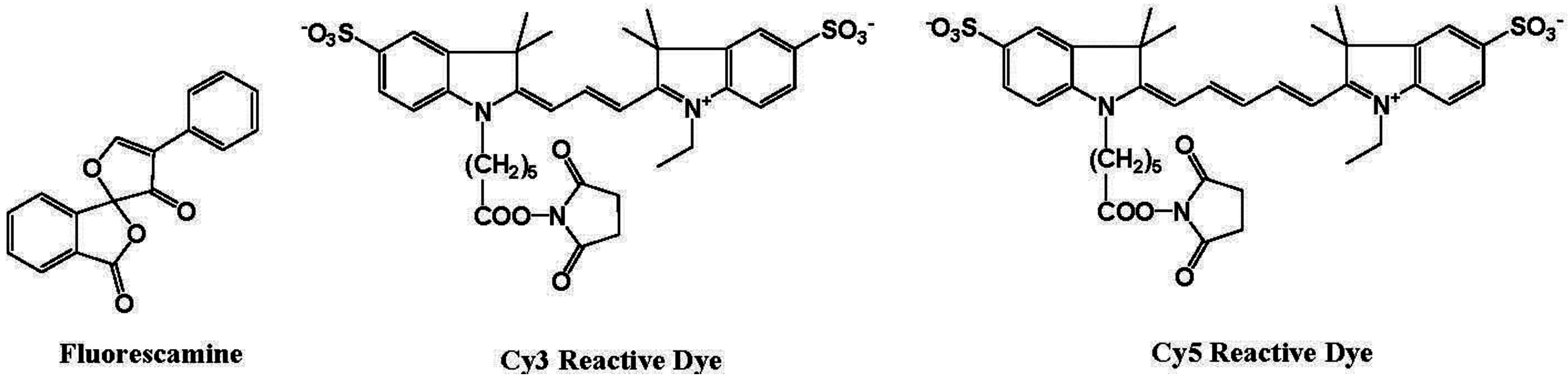



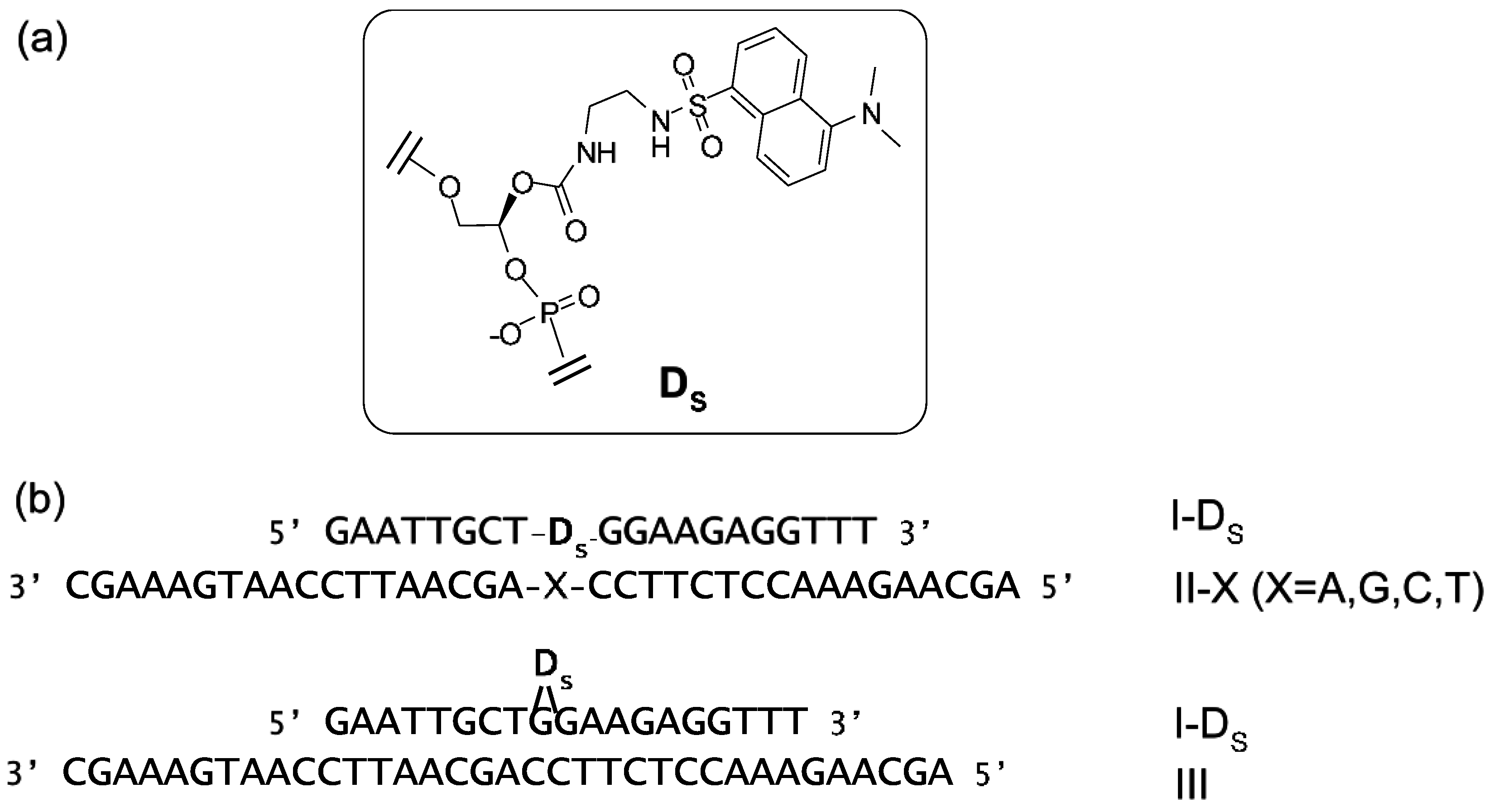
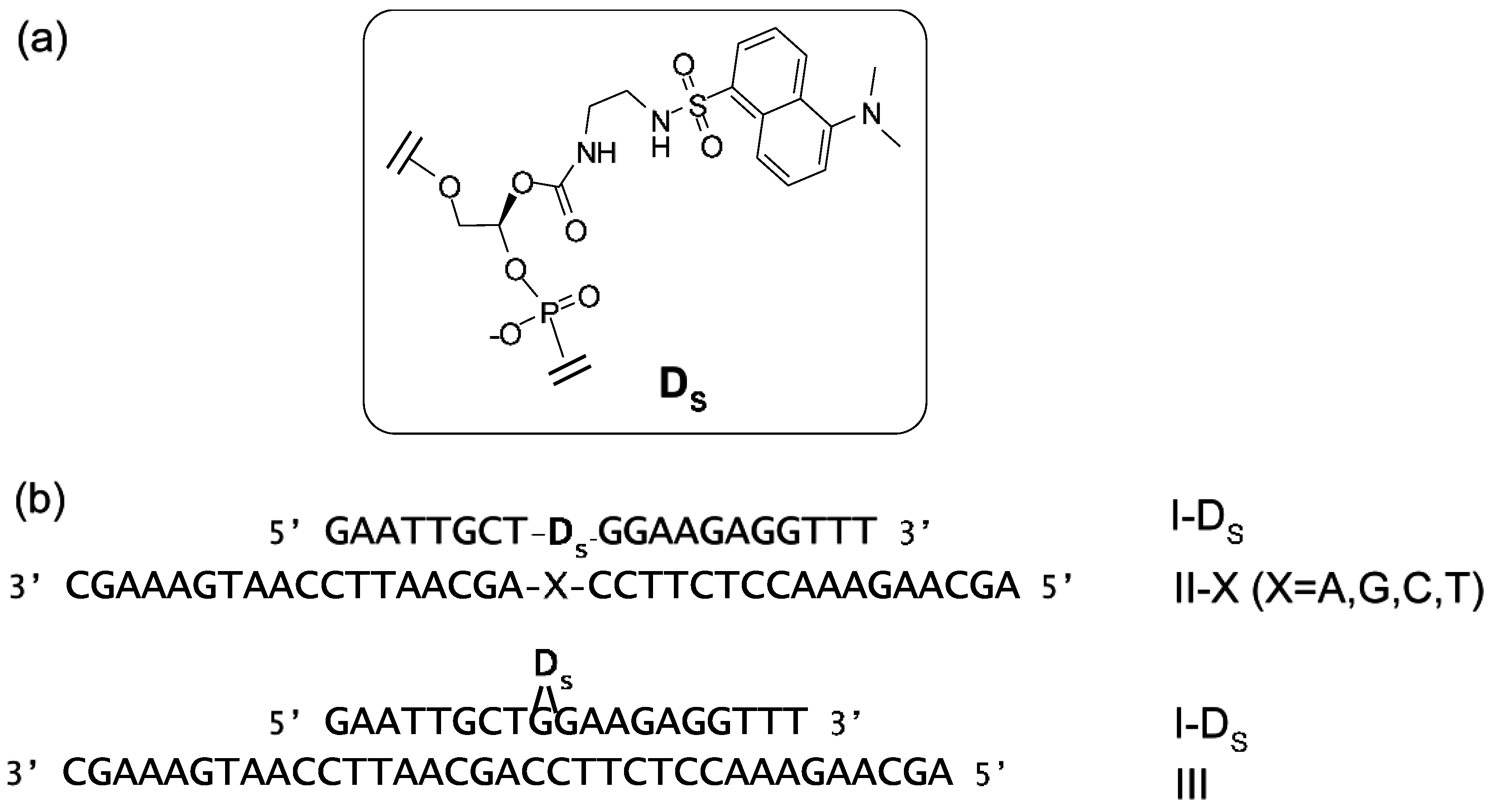
6. Fluorescent Probes for the Measurement of DNA and RNA
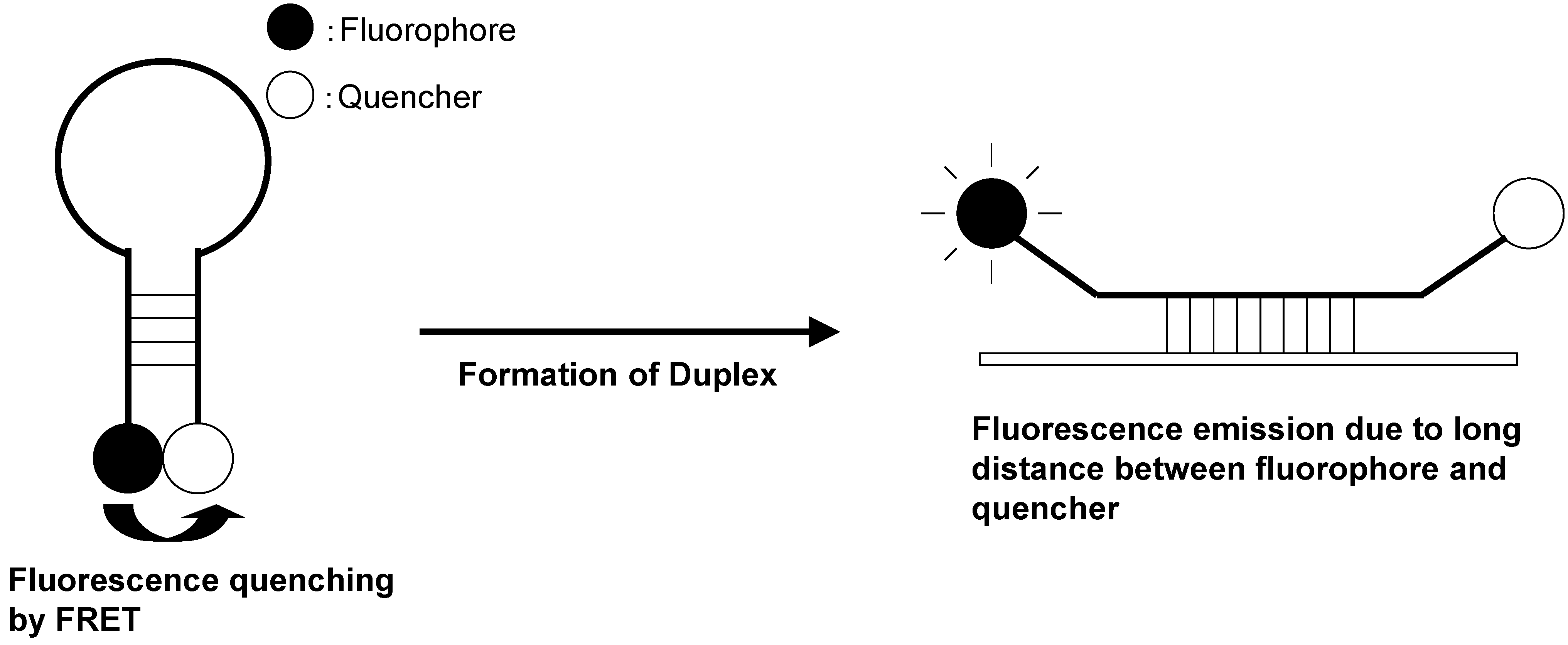
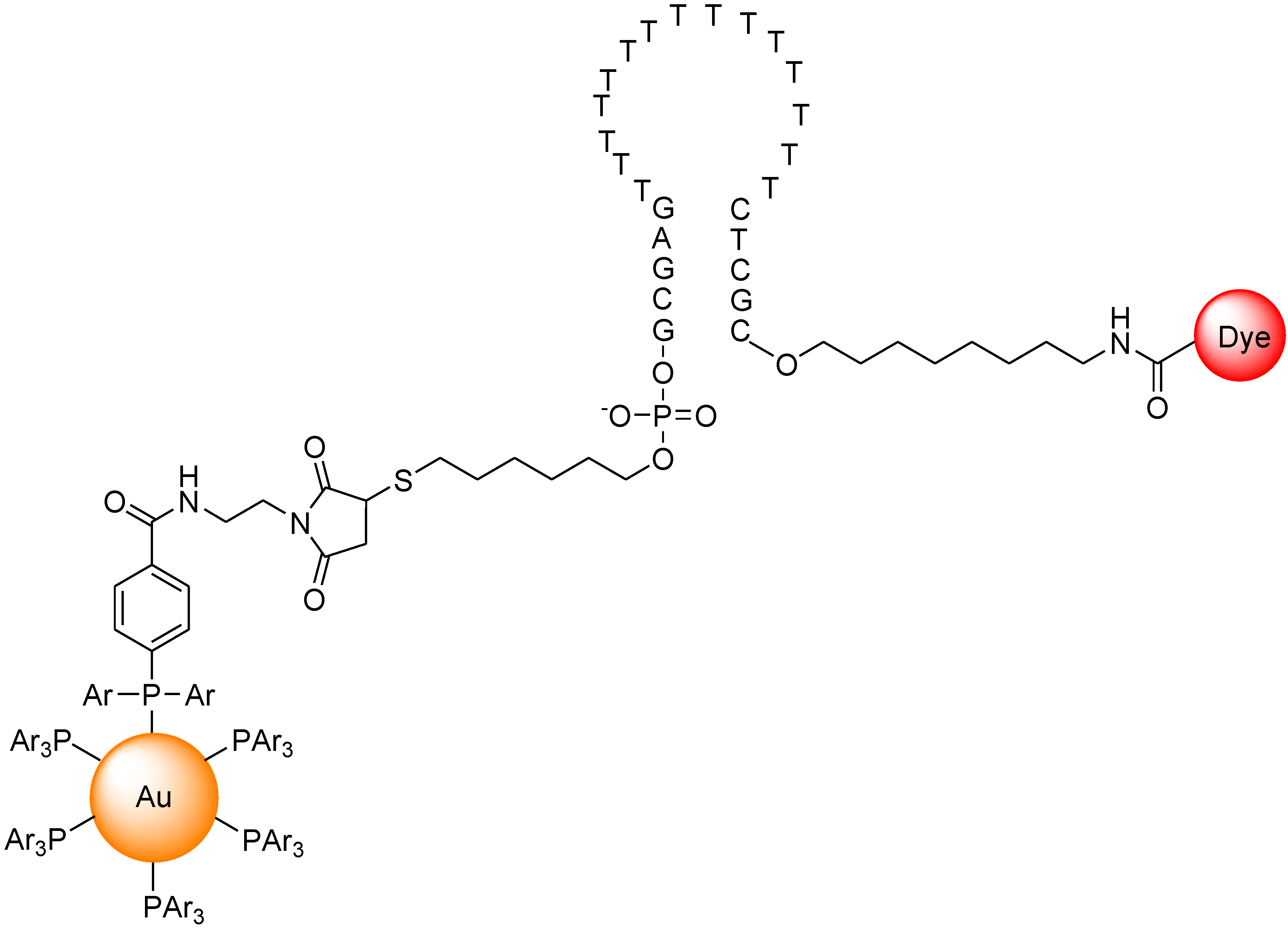
7. Conclusions
Conflicts of Interest
References
- Ishikawa, H.; Suzuki, K.; Nakanishi, M.; Inokai, A. Bioimaging for Molecular Dynamics and Cellular Functions; Kyoritsushuppan Co., Ltd.: Tokyo, Japan, 1998. [Google Scholar]
- Rurack, K.; Resch-Genger, U. Rigidization, preorientation and electronic decoupling—The ‘magic triangle’ for the design of highly efficient fluorescent sensors and switches. Chem. Soc. Rev. 2002, 31, 116–127. [Google Scholar] [CrossRef] [PubMed]
- Weller, A. Mechanism and Spin-Dynamics of Photoinduced Electron-Transfer Reactions. Zeitschrift fur Physikalische Chemie-Wiesbaden 1982, 130, 129–138. [Google Scholar] [CrossRef]
- Fabbrizzi, L.; Lichelli, M.; Pallavicini, P.; Sacchi, D.; Taglietti, A. Sensing of transition metals through fluorescence quenching or enhancement: A review. Analyst 1996, 121, 1763–1768. [Google Scholar] [CrossRef]
- Bergonzi, R.; Fabbrizzi, L.; Lichelli, M.; Mangano, C. Molecular switches of fluorescence operating through metal centred redox couples. Coord. Chem. Rev. 1998, 170, 31–46. [Google Scholar] [CrossRef]
- Ganguly, A.; Ghosh, S.; Kar, S.; Guchhait, N. Selective fluorescence sensing of Cu(II) and Zn(II) using a simple Schiff base ligand: Naked eye detection and elucidation of photoinduced electron transfer (PET) mechanism. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2015, 143, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Valeur, B.; Schulman, S.G. Molecular Luminescence Spectroscopy, Part 3; Wiley: New York, NY, USA, 1993. [Google Scholar]
- Löhr, H.-G.; Vögtle, F. Chromo- and fluoroionophores. A new class of dye reagents. Acc. Chem. Res. 1985, 18, 65–72. [Google Scholar] [CrossRef]
- Manna, A.K.; Balamurugan, D.; Cheung, M.S.; Dunietz, B.D. Unraveling the Mechanism of Photoinduced Charge Transfer in Carotenoid-Porphyrin-C-60 Molecular Triad. J. Phys. Chem. Lett. 2015, 6, 1231–1237. [Google Scholar] [CrossRef]
- Hopfield, J.J. Photoinduced charge-transfer—Critical test of mechanism and range of biological electron-transfer processes. Biophys. J. 1977, 18, 311–321. [Google Scholar] [CrossRef]
- Campbell, A.K. Intracellular Calcium; John Wiley and Sons: New York, NY, USA, 1983. [Google Scholar]
- Bootman, M.D.; Lipp, P.; Berridge, M.J. The organisation and functions of local Ca2+ signals. J. Cell Sci. 2001, 114, 2213–2222. [Google Scholar] [PubMed]
- Grynkiewicz, G.; Poenie, M.; Tsien, R.Y. A new generation of Ca2+ indicators with greatly improved fluorescence properties. J. Biol. Chem. 1985, 260, 3440–3450. [Google Scholar] [PubMed]
- Tsien, R.Y. New calcium indicators and buffers with high selectivity against magnesium and protons: Design, synthesis, and properties of prototype structures. Biochemistry 1980, 19, 2396–2404. [Google Scholar] [CrossRef] [PubMed]
- Roe, M.W.; Lemasters, J.J.; Herman, B. Assessment of Fura-2 for measurements of cytosolic free calcium. Cell Calcium 1990, 11, 63–73. [Google Scholar] [CrossRef]
- Berridge, M.J.; Lipp, P.; Bootman, M.D. The versatility and universality of calcium signalling. Nat. Rev. Mol. Cell Biol. 2000, 1, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Augustine, G.J.; Santamaria, F.; Tanaka, K. Local Calcium Signaling in Neurons. Neuron 2003, 40, 331–346. [Google Scholar] [CrossRef]
- Leite, M.F.; Thrower, E.D.; Echevarria, W.; Koulen, P.; Hirata, K.; Bennett, A.M.; Ehrlich, B.E.; Nathanson, M.H. Nuclear and cytosolic calcium are regulated independently. Proc. Natl. Acad. Sci. USA 2003, 100, 2975–2980. [Google Scholar] [CrossRef] [PubMed]
- Csordas, G.; Hajnoczky, G. Plasticity of Mitochondrial Calcium Signaling. J. Biol. Chem. 2003, 278, 42273–42282. [Google Scholar] [CrossRef] [PubMed]
- Naziroglu, M.; Ovey, I.S. Involvement of apoptosis and calcium accumulation through TRPV1 channels in neurobiology of epilepsy. Neuroscience 2015, 293, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Pietrzak-Nowacka, M.; Safranow, K.; Bober, J.; Olszewska, M.; Birkenfeld, B.; Nowosiad, M.; Ciechanowski, K. Clinical research Calcium-phosphate metabolism parameters and erythrocyte Ca2+ concentration in autosomal dominant polycystic kidney disease patients with normal renal function. Arch. Med. Sci. 2013, 9, 837–842. [Google Scholar] [CrossRef] [PubMed]
- Thompson, M.D.; Mei, Y.; Weisbrod, R.M.; Silver, M.; Shukla, P.C.; Bolotina, V.M.; Cohen, R.A.; Tong, X.Y. Glutathione adducts on sarcoplasmic/endoplasmic reticulum Ca2+ ATPase Cys-674 regulate endothelial cell calcium stores and angiogenic function as well as promote ischemic blood flow recovery. J. Biol. Chem. 2014, 289, 19907–19916. [Google Scholar] [CrossRef] [PubMed]
- Minta, A.; Kao, J.P.; Tsien, R.Y. Fluorescent indicators for cytosolic calcium based on rhodamine and fluorescein chromophores. J. Biol. Chem. 1989, 264, 8171–8178. [Google Scholar] [PubMed]
- Staehlke, S.; Koertge, A.; Nebe, B. Intracellular calcium dynamics dependent on defined microtopographical features of titanium. Biomaterials 2015, 46, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Narasimhan, G.; de Alba-Aguayo, D.R.; Mondragon-Flores, R.; Gonzalez-Pozos, S.; Miranda-Saturnino, M.J.; Sridharan, M.; Rueda, A. Acute Administration of Chitosan Nanoparticles Increases Ca2+ Leak in Rat Cardiomyocytes. J. Nano Res. 2014, 28, 29–38. [Google Scholar] [CrossRef]
- Lai, Y.Z.; Kisaalita, W.S. Performance Evaluation of 3D Polystyrene 96-Well Plates with Human Neural Stem Cells in a Calcium Assay. JALA 2012, 17, 284–292. [Google Scholar] [CrossRef] [PubMed]
- Gee, K.R.; Archer, E.A.; Lapham, L.A.; Leovard, M.E.; Zhou, Z.-L.; Diwu, J.Z. New ratiometric fluorescent calcium indicators with moderately attenuated binding affinities. Bioorg. Med. Chem. Lett. 2000, 10, 1515–1518. [Google Scholar] [CrossRef]
- Martin, V.V.; Beierlein, M.; Morgan, J.L.; Rothe, A.; Gee, K.R. Novel fluo-4 analogs for fluorescent calcium measurements. Cell Calcium 2004, 36, 509–514. [Google Scholar] [CrossRef] [PubMed]
- Ozmen, B.; Akkaya, E.U. Infrared fluorescence sensing of submicromolar calcium: Pushing the limits of photoinduced electron transfer. Tetrahedron Lett. 2000, 41, 9185–9188. [Google Scholar] [CrossRef]
- Akkaya, E.U.; Turkyilmaz, S. A squaraine-based near IR fluorescent chemosensor for Calcium. Tetrahedron Lett. 1997, 38, 4513–4516. [Google Scholar] [CrossRef]
- Matsui, A.; Umezawa, K.; Shindo, Y.; Fujii, T.; Citterio, D.; Oka, K.; Suzuki, K. A near-infrared fluorescent calcium probe: A new tool for intracellular multicolour Ca2+ imaging. Chem. Commun. 2011, 47, 10407–10409. [Google Scholar] [CrossRef] [PubMed]
- Fatholahi, M.; LaNoue, K.; Romani, A.; Scarpa, A. Relationship between Total and Free Cellular Mg2+ during Metabolic Stimulation of Rat Cardiac Myocytes and Perfused Hearts. Arch. Biochem. Biophys. 2000, 374, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Gaussin, V.; Gailly, P.; Gillis, J.M.; Hue, L. Fructose-induced increase in intracellular free Mg2+ ion concentration in rat hepatocytes: Relation with the enzymes of glycogen metabolism. Biochem. J. 1997, 326, 823–827. [Google Scholar] [PubMed]
- Romani, A.M.; Scarpa, A. Regulation of cell magnesium. Arch. Biochem. Biophys. 1992, 298, 1–12. [Google Scholar] [CrossRef]
- London, R.E. Methods for Measurement of Intracellular Magnesium:NMR and Fluorescence. Annu. Rev. Physiol. 1991, 53, 241–258. [Google Scholar] [CrossRef] [PubMed]
- Murphy, E.; Freudenrich, C.C.; Levy, L.A.; London, R.E.; Lieberman, M. Monitoring cytosolic free magnesium in cultured chicken heart cells by use of the fluorescent indicator Furaptra. Proc. Natl. Acad. Sci. USA 1989, 86, 2981–2984. [Google Scholar] [CrossRef] [PubMed]
- Touyz, R.M.; Schiffrin, E.L. Angiotensin II and Vasopressin Modulate Intracellular Free Magnesium in Vascular Smooth Muscle Cells through Na+-dependent Protein Kinase C Pathways. J. Biol. Chem. 1996, 271, 24353–24358. [Google Scholar] [CrossRef] [PubMed]
- Tashiro, M.; Inoue, H.; Konishi, M. Physiological Pathway of Magnesium Influx in Rat Ventricular Myocytes. Biophys. J. 2014, 107, 2049–2058. [Google Scholar] [CrossRef] [PubMed]
- Gouadon, E.; Lecerf, F.; German-Fattal, M. Differential Effects of Cyclosporin A and Tacrolimus on Magnesium Influx in Caco2 Cells. J. Pharm. Pharmceu. Sci. 2012, 15, 389–398. [Google Scholar]
- Csernoch, L.; Bernengo, J.C.; Szentesi, P.; Jacquemond, V. Measurements of Intracellular Mg2+ Concentration in Mouse Skeletal Muscle Fibers with the Fluorescent Indicator Mag-Indo-1. Biophys. J. 1998, 75, 957–967. [Google Scholar] [CrossRef]
- Cheng, C.; Reynolds, I.J. Subcellular localization of glutamate-stimulated intracellular magnesium concentration changes in cultured rat forebrain neurons using confocal microscopy. Neurosciences 2000, 95, 973–979. [Google Scholar] [CrossRef]
- Meena, L.S.; Dhakate, S.R.; Sahare, P.D. Elucidation of Mg2+ binding activity of adenylate kinase from Mycobacterium tuberculosis H(37)Rv using fluorescence studies. Biotech. Appl. Biochem. 2012, 59, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.C.; Broedersz, C.P.; Rowat, A.C.; Wedig, T.; Herrmann, H.; MacKintosh, F.C.; Weitz, D.A. Divalent Cations Crosslink Vimentin Intermediate Filament Tail Domains to Regulate Network Mechanics. J. Mol. Biol. 2010, 399, 637–644. [Google Scholar] [CrossRef] [PubMed]
- Shmigol, A.V.; Eisner, D.A.; Wray, B. Simultaneous measurements of changes in sarcoplasmic reticulum and cytosolic [Ca2+] in rat uterine smooth muscle cells. J. Physiol. 2001, 531, 707–713. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.D.; Hollingworth, S.; Baylor, S.M. Properties of tri- and tetracarboxylate Ca2+ indicators in frog skeletal muscle fibers. Biophys. J. 1996, 70, 896–916. [Google Scholar] [CrossRef]
- Gongqin, S.; Raymond, J.A.B. Requirement for an Additional Divalent Metal Cation to Activate Protein Tyrosine Kinase. Biochemistry 1997, 36, 2139–2146. [Google Scholar]
- Budinger, G.R.S.; Duranteau, J.; Chandel, N.S.; Schumacker, P.T. Hibernation during Hypoxia in Cardiomyocytes: Role of mitochondria as the O2sensor. J. Biol. Chem. 1998, 273, 3320–3326. [Google Scholar] [CrossRef] [PubMed]
- Haugland, R.P. Handbook of Fluorescent Probes and Research Chemicals, 7th ed.; Molecular Probes Inc.: Leiden, The Netherlands, 2002. [Google Scholar]
- Pesco, J.; Salmon, J.; Vigo, J.; Viallet, P. Mag-indo1 Affinity for Ca2+, Compartmentalization and Binding to Proteins: The Challenge of Measuring Mg2+ Concentrations in Living Cells. Anal. Biochem. 2001, 290, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Komatsu, H.; Ikeda, T.; Saito, N.; Araki, S.; Citterio, D.; Hisamoto, H.; Kitamura, Y.; Kubota, T.; Nakagawa, J.; Oka, K.; Suzuki, K. Design and Synthesis of Mg2+-Selective Fluoroionophores Based on a Coumarin Derivative and Application for Mg2+ Measurement in a Living Cell. Anal. Chem. 2002, 74, 1423–1428. [Google Scholar] [CrossRef] [PubMed]
- Park, E.J.; Brasuel, M.; Martin, A.P.; Kopelman, R. Ratiometric Optical PEBBLE Nanosensors for Real-Time Magnesium Ion Concentrations inside Viable Cells. Anal. Chem. 2003, 75, 3784–3791. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, H.; Iwasawa, N.; Citterio, D.; Suzuki, Y.; Kubota, T.; Tokuno, K.; Kitamura, Y.; Oka, K.; Suzuki, K. Design and Synthesis of Highly Sensitive and Selective Fluorescein-Derived Magnesium Fluorescent Probes and Application to Intracellular 3D Mg2+ Imaging. J. Am. Chem. Soc. 2004, 126, 16353–16360. [Google Scholar] [CrossRef] [PubMed]
- Fujii, T.; Shindo, Y.; Hotta, K.; Citterio, D.; Nishiyama, S.; Suzuki, K.; Oka, K. Design and Synthesis of a FlAsH-Type Mg2+ Fluorescent Probe for Specific Protein Labeling. J. Am. Chem. Soc. 2014, 136, 2374–2381. [Google Scholar] [CrossRef] [PubMed]
- Frausto da Silva, J.J.R.; Williams, R.J.P. The Biological Chemistry of the Elements; Oxford University Press: Oxford, UK, 1991. [Google Scholar]
- Mills, C.F. Zinc in Human Biology, Humannutrition Reviews; Springer-Verlag: Berlin, Germany, 1988. [Google Scholar]
- Xie, X.; Smart, T.J. A physiological role for endogenous zinc in rat hippocampal synaptic neurotransmission. Nature 1991, 349, 521–523. [Google Scholar] [CrossRef] [PubMed]
- Koh, J.-Y.; Choi, D.W. Zinc and brain injury. Annu. Rev. Neurosci. 1998, 21, 347–375. [Google Scholar]
- Bush, A.I.; Pettingel, W.H.; Multhaup, G.; Paradis, M.D.; Vonsattel, J.-P.; Gusella, J.F.; Beyreuther, K.; Masters, C.L.; Tanzi, R.E. Rapid induction of Alzheimer A beta amyloid formation by zinc. Science 1994, 265, 1464–1467. [Google Scholar] [CrossRef] [PubMed]
- Hendrickson, K.M.; Rodopoulos, T.; Pittet, P.-A.; Mahadevan, I.; Lincoln, S.F.; Ward, A.D.; Kurucsev, T.; Duckworth, P.A.; Forbes, I.J.; Zalewski, P.D.; Betts, W.H. Complexation of zinc(II) and other divalent metal ions by the fluorophore 2-methyl-8-(toluene-p-sulfonamido)-6-quinolyloxyacetic acid in 50% aqueous ethanol. J. Chem. Soc. Dalton Trans. 1997, 3879–3882. [Google Scholar] [CrossRef]
- Fahrni, C.J.; O’Halloran, T.V. Aqueous Coordination Chemistry of Quinoline-Based Fluorescence Probes for the Biological Chemistry of Zinc. J. Am. Chem. Soc. 1999, 121, 11448–11458. [Google Scholar] [CrossRef]
- Qian, W.J.; Aspinwall, C.A.; Battiste, M.A.; Kennedy, R.T. Detection of Secretion from Single Pancreatic β-Cells Using Extracellular Fluorogenic Reactions and Confocal Fluorescence Microscopy. Anal. Chem. 2000, 72, 711–717. [Google Scholar] [CrossRef] [PubMed]
- Golan, Y.; Berman, B.; Assaraf, Y.G. Heterodimerization, Altered Subcellular Localization, and Function of Multiple Zinc Transporters in Viable Cells Using Bimolecular Fluorescence Complementation. J. Biol. Chem. 2015, 290, 9050–9063. [Google Scholar] [CrossRef] [PubMed]
- Lasry, I.; Golan, Y.; Berman, B.; Amram, N.; Glaser, F.; Assaraf, Y.G. In Situ Dimerization of Multiple Wild Type and Mutant Zinc Transporters in Live Cells Using Bimolecular Fluorescence Complementation. J. Biol. Chem. 2014, 289, 7275–7292. [Google Scholar] [CrossRef] [PubMed]
- Kinber, M.C.; Mahadevan, I.B.; Lincoln, S.F.; Ward, A.D.; Tiekink, E.R.T. The Synthesis and Fluorescent Properties of Analogues of the Zinc(II) Specific Fluorophore Zinquin Ester. J. Org. Chem. 2000, 65, 8204–8209. [Google Scholar] [CrossRef]
- Walkup, G.K.; Burdette, S.C.; Lippard, S.J.; Tsien, R.Y. A New Cell-Permeable Fluorescent Probe for Zn2+. J. Am. Chem. Soc. 2000, 122, 5644–5645. [Google Scholar] [CrossRef]
- Sinha, S.; Mukherjee, T.; Mathew, J.; Mukhopadhyay, S.K.; Ghosh, S. Triazole-based Zn2+-specific molecular marker for fluorescence bioimaging. Anal. Chim. Acta 2014, 822, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Buccella, D.; Lippard, S.J. Visualization of Peroxynitrite-Induced Changes of Labile Zn2+ in the Endoplasmic Reticulum with Benzoresorufin-Based Fluorescent Probes. J. Am. Chem. Soc. 2013, 135, 13512–13520. [Google Scholar] [CrossRef] [PubMed]
- Hirano, T.; Kikuchi, K.; Urano, Y.; Higuchi, T.; Nagano, T. Novel Zinc Fluorescent Probes Excitable with Visible Light for Biological Applications. Angew. Chem. Int. Ed. 2000, 39, 1052–1054. [Google Scholar] [CrossRef]
- Hirano, T.; Kikuchi, K.; Urano, Y.; Higuchi, T.; Nagano, T. Highly Zinc-Selective Fluorescent Sensor Molecules Suitable for Biological Applications. J. Am. Chem. Soc. 2000, 122, 12399–12400. [Google Scholar] [CrossRef]
- Hirano, T.; Kikuchi, K.; Urano, Y.; Nagano, T. Improvement and Biological Applications of Fluorescent Probes for Zinc, ZnAFs. J. Am. Chem. Soc. 2002, 124, 6555–6562. [Google Scholar] [CrossRef] [PubMed]
- Takeda, A.; Suzuki, M.; Tamano, H.; Takada, S.; Ide, K.; Oku, N. Involvement of glucocorticoid-mediated Zn2+ signaling in attenuation of hippocampal CA1 LTP by acute stress. Neurochem. Int. 2012, 60, 394–399. [Google Scholar] [CrossRef] [PubMed]
- Nydegger, I.; Rumschik, S.M.; Kay, A.R. Zinc Is Externalized Rather than Released during Synaptic Transmission. ACS Chem. Neurosci. 2010, 1, 728–736. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.Q.; Kim, G.H.; Yoon, J.; Shin, I. Synthesis of a highly Zn2+-selective cyanine-based probe and its use for tracing endogenous zinc ions in cells and organisms. Nat. Protocol. 2014, 9, 1245–1254. [Google Scholar] [CrossRef] [PubMed]
- Bhown, A.S.; Cornelius, T.W.; Volanakis, J.E.; Bennett, J.C. A comparison of fluorescamine and o-phthaldialdehyde as effective blocking reagents in protein sequence analyses by the Beckman sequencer. Anal. Biochem. 1983, 131, 337–340. [Google Scholar] [CrossRef]
- Amano, H.; Ohuchi, Y.; Katayama, Y.; Maeda, M. A New Fluorescent Reagent for the Detection of Proteins Having Histidine-tag (his-tag). Anal. Sci. 2001, 17, i1469–i1472. [Google Scholar]
- Kapanidis, A.N.; Ebright, Y.W.; Ebright, R.H. Site-Specific Incorporation of Fluorescent Probes into Protein: Hexahistidine-Tag-Mediated Fluorescent Labeling with (Ni2+:Nitrilotriacetic Acid)n−Fluorochrome Conjugates. J. Am. Chem. Soc. 2001, 123, 12123–12125. [Google Scholar] [CrossRef] [PubMed]
- Adams, S.R.; Campbell, R.E.; Gross, L.A.; Martin, B.R.; Walkup, G.K.; Yao, Y.; Llopis, J.; Tsien, R.Y. New Biarsenical Ligands and Tetracysteine Motifs for Protein Labeling in Vitro and in Vivo: Synthesis and Biological Applications. J. Am. Chem. Soc. 2002, 124, 6063–6076. [Google Scholar] [CrossRef] [PubMed]
- Ojida, A.; Mito-oka, Y.; Inoue, M.; Hamachi, I. First Artificial Receptors and Chemosensors toward Phosphorylated Peptide in Aqueous Solution. J. Am. Chem. Soc. 2002, 124, 6256–6258. [Google Scholar] [CrossRef] [PubMed]
- Ojida, A.; Honda, K.; Shinmi, D.; Kiyonaka, S.; Mori, Y.; Hamachi, I. Oligo-Asp Tag/Zn(II) Complex Probe as a New Pair for Labeling and Fluorescence Imaging of Proteins. J. Am. Chem. Soc. 2006, 128, 10452–10459. [Google Scholar] [CrossRef] [PubMed]
- Honda, K.; Fujishima, S.; Ojida, A.; Hamachi, I. Pyrene Excimer-Based Dual-Emission Detection of a Oligoaspartate Tag-Fused Protein by Using a ZnII–DpaTyr Probe. ChemBioChem 2007, 8, 1370–1372. [Google Scholar] [CrossRef] [PubMed]
- Ojida, A.; Sakamoto, T.; Inoue, M.; Fujishima, S.; Lippens, G.; Hamachi, I. Fluorescent BODIPY-Based Zn(II) Complex as a Molecular Probe for Selective Detection of Neurofibrillary Tangles in the Brains of Alzheimer’s Disease Patients. J. Am. Chem. Soc. 2009, 131, 6543–6548. [Google Scholar] [CrossRef] [PubMed]
- Somura, M.; Takiyama, K.; Kinoshita-Kikuta, E.; Kinoshita, E.; Koike, T. A Phos-tag-based fluorescence resonance energy transfer system for the analysis of the kinase reaction of a substrate peptide. Anal. Methods 2011, 3, 1303–1309. [Google Scholar] [CrossRef]
- Suzuki, Y.; Yokoyama, K. Design and Synthesis of Intramolecular Charge Transfer-Based Fluorescent Reagents for the Highly-Sensitive Detection of Proteins. J. Am. Chem. Soc. 2005, 127, 17799–17802. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Yokoyama, K. Design and synthesis of ICT-based fluorescent probe for high-sensitivity protein detection and application to rapid protein staining for SDS-PAGE. Proteomics 2008, 8, 2785–2790. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Takagi, N.; Chimuro, T.; Shinohara, A.; Sakaguchi, N.; Hiratsuka, A.; Yokoyama, K. Design and synthesis of new fluorescent probe for rapid and highly sensitive detection of proteins via electrophoretic gel stain. Electrophoresis 2011, 32, 1403–1413. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Takagi, N.; Sano, T.; Chimuro, T. Design and synthesis of a novel fluorescent protein probe for easy and rapid electrophoretic gel staining by using a commonly available UV-based fluorescent imaging system. Electrophoresis 2013, 34, 2464–2472. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Yokoyama, K. Development of a Fluorescent Peptide for the Detection of Vascular Endothelial Growth Factor (VEGF). ChemBioChem 2009, 10, 1793–1795. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Yokoyama, K. Construction of a more sensitive fluorescence sensing material for the detection of vascular endothelial growth factor, a biomarker for angiogenesis, prepared by combining a fluorescent peptide and a nanopillar substrate. Biosens. Bioelectron. 2011, 26, 3696–3699. [Google Scholar] [CrossRef] [PubMed]
- Niemeyer, C.M.; Blohm, D. DNA-Mikroarrays. Angew. Chem. 1999, 111, 3039–3043. [Google Scholar] [CrossRef]
- Suzuki, Y.; Kowata, K.; Komatsu, Y. Development of dansyl-modified oligonucleotide probes responding to structural changes in a duplex. Bioorg. Med. Chem. Lett. 2013, 23, 6123–6126. [Google Scholar] [CrossRef] [PubMed]
- Shibata, A.; Abe, H.; Furukawa, K.; Tsuneda, S.; Ito, Y. Reduction-Triggered Fluorescence Probe for Peptide-Templated Reactions. Chem. Pharmceu. Bull. 2009, 57, 1223–1226. [Google Scholar] [CrossRef]
- Tyagi, S.; Kramer, F.R. Molecular Beacons: Probes that Fluoresce upon Hybridization. Nat. Biotechnol. 1996, 14, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, S.; Bratu, D.P.; Kramer, F.R. Multicolor molecular beacons for allele discrimination. Nat. Biotechnol. 1998, 16, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Leone, G.; van Schijndel, H.; van Gemen, B.; Kramer, F.R.; Schoen, C.D. Molecular beacon probes combined with amplification by NASBA enable homogeneous, real-time detection of RNA. Nucleic Acids Res. 1998, 26, 2150–2155. [Google Scholar] [CrossRef] [PubMed]
- Steemers, F.J.; Ferguson, J.A.; Walt, D.R. Screening unlabeled DNA targets with randomly ordered fiber-optic gene arrays. Nat. Biotechnol. 2000, 18, 91–94. [Google Scholar] [PubMed]
- Darby, R.A.J.; Sollogoub, M.; McKeen, C.; Brown, L.; Risitano, A.; Brown, N.; Barton, C.; Brown, T.; Fox, K.R. High throughput measurement of duplex, triplex and quadruplex melting curves using molecular beacons and a LightCycler. Nucleic Acids Res. 2002, 30. [Google Scholar] [CrossRef]
- Dubertret, B.; Calame, M.; Libchaber, A.J. Single-mismatch detection using gold-quenched fluorescent oligonucleotides. Nat. Biotech. 2001, 19, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Ping, J.F.; Zhou, Y.B.; Wu, Y.Y.; Papper, V.; Boujday, S.; Marks, R.S.; Steele, T.W.J. Recent advances in aptasensors based on graphene and graphene-like nanomaterials. Biosens. Bioelectron. 2015, 64, 373–385. [Google Scholar] [CrossRef] [PubMed]
- Han, S.X.; Jia, X.; Ma, J.L.; Zhu, Q. Molecular Beacons: A Novel Optical Diagnostic Tool. Arch. Immunol. Therapiae Exp. 2013, 61, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Ostergaard, M.E.; Hrdlicka, P.J. Pyrene-functionalized oligonucleotides and locked nucleic acids (LNAs): Tools for fundamental research, diagnostics, and nanotechnology. Chem. Soc. Rev. 2011, 40, 5771–5788. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suzuki, Y.; Yokoyama, K. Development of Functional Fluorescent Molecular Probes for the Detection of Biological Substances. Biosensors 2015, 5, 337-363. https://doi.org/10.3390/bios5020337
Suzuki Y, Yokoyama K. Development of Functional Fluorescent Molecular Probes for the Detection of Biological Substances. Biosensors. 2015; 5(2):337-363. https://doi.org/10.3390/bios5020337
Chicago/Turabian StyleSuzuki, Yoshio, and Kenji Yokoyama. 2015. "Development of Functional Fluorescent Molecular Probes for the Detection of Biological Substances" Biosensors 5, no. 2: 337-363. https://doi.org/10.3390/bios5020337
APA StyleSuzuki, Y., & Yokoyama, K. (2015). Development of Functional Fluorescent Molecular Probes for the Detection of Biological Substances. Biosensors, 5(2), 337-363. https://doi.org/10.3390/bios5020337