First Vibrational Fingerprint of Parietaria judaica Protein via Surface-Enhanced Raman Spectroscopy

 , ,
, ,  ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Production of Recombinant Allergenic Proteins
2.2. Ag Dendrites Synthesis and SERS Sample Preparation and Characterization
2.3. Raman and SERS Measurements
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Par j 1 | Parietaria judaica 1 |
| Par j 2 | Parietaria judaica 2 |
| Par j 4 | Parietaria judaica 4 |
| SERS | Surface-enhanced Raman spectroscopy |
References
- Ishizaka, K.; Ishizaka, T. Identification of γE-Antibodies as a Carrier of Reaginic Activity. J. Immunol. 1967, 99, 1187–1198. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Chen, J.; Ye, L.; Cai, Z.; Sun, J.; Ji, K. Anti-IgE Therapy for IgE-Mediated Allergic Diseases: From Neutralizing IgE Antibodies to Eliminating IgE+ B Cells. Clin. Transl. Allergy 2018, 8, 27. [Google Scholar] [CrossRef]
- Izmailovich, M.; Semenova, Y.; Abdushukurova, G.; Mukhamejanova, A.; Dyussupova, A.; Faizova, R.; Gazaliyeva, M.; Akhvlediani, L.; Glushkova, N.; Kalmakhanov, S.; et al. Molecular Aspects of Allergen-Specific Immunotherapy in Patients with Seasonal Allergic Rhinitis. Cells 2023, 12, 383. [Google Scholar] [CrossRef] [PubMed]
- Ciprandi, G.; Puccinelli, P.; Incorvaia, C.; Masieri, S. Parietaria Allergy: An Intriguing Challenge for the Allergist. Medicina 2018, 54, 106. [Google Scholar] [CrossRef] [PubMed]
- Colombo, P.; Bonura, A.; Costa, M.A.; Izzo, V.; Passantino, R.; Locorotondo, G.; Amoroso, S.; Geraci, D. The Allergens of Parietaria. Int. Arch. Allergy Immunol. 2003, 130, 173–179. [Google Scholar] [CrossRef]
- Colombo, P.; Kennedy, D.; Ramsdale, T.; Costa, M.A.; Duro, G.; Izzo, V.; Salvadori, S.; Guerrini, R.; Cocchiara, R.; Mirisola, M.G.; et al. Identification of an Immunodominant IgE Epitope of the Parietaria judaica Major Allergen. J. Immunol. 1998, 160, 2780–2785. [Google Scholar] [CrossRef]
- Bonura, A.; Gulino, L.; Trapani, A.; Di Felice, G.; Tinghino, R.; Amoroso, S.; Geraci, D.; Valenta, R.; Westritschnig, K.; Scala, E.; et al. Isolation, Expression and Immunological Characterization of a Calcium-Binding Protein from Parietaria Pollen. Mol. Immunol. 2008, 45, 2465–2473. [Google Scholar] [CrossRef]
- Molecular Allergology User’s Guide 2.0. Available online: https://hub.eaaci.org/resources_documents/molecular-allergology-users-guide-2-0/ (accessed on 16 July 2024).
- Chen, M.C.; Lord, R.C. Laser-Excited Raman Spectroscopy of Biomolecules. VIII. Conformational Study of Bovine Serum Albumin. J. Am. Chem. Soc. 1976, 98, 990–992. [Google Scholar] [CrossRef]
- David, C.; d’Andrea, C.; Lancelot, E.; Bochterle, J.; Guillot, N.; Fazio, B.; Maragò, O.M.; Sutton, A.; Charnaux, N.; Neubrech, F.; et al. Raman and IR Spectroscopy of Manganese Superoxide Dismutase, a Pathology Biomarker. Vib. Spectrosc. 2012, 62, 50–58. [Google Scholar] [CrossRef]
- Moskovits, M. Surface-Enhanced Spectroscopy. Rev. Mod. Phys. 1985, 57, 783–826. [Google Scholar] [CrossRef]
- Ru, E.L.; Etchegoin, P. Principles of Surface-Enhanced Raman Spectroscopy: And Related Plasmonic Effects; Elsevier: Amsterdam, The Netherlands, 2008; ISBN 978-0-08-093155-5. [Google Scholar]
- Lo Faro, M.J.; Leonardi, A.A.; Morganti, D.; Sciuto, E.L.; Irrera, A.; Fazio, B. Surface-Enhanced Raman Scattering for Biosensing Platforms: A Review. Radiat. Eff. Defects Solids 2022, 177, 1209–1221. [Google Scholar] [CrossRef]
- Nie, S.; Emory, S.R. Probing Single Molecules and Single Nanoparticles by Surface-Enhanced Raman Scattering. Science 1997, 275, 1102–1106. [Google Scholar] [CrossRef] [PubMed]
- Fazio, B.; D’Andrea, C.; Foti, A.; Messina, E.; Irrera, A.; Donato, M.G.; Villari, V.; Micali, N.; Maragò, O.M.; Gucciardi, P.G. SERS Detection of Biomolecules at Physiological pH via Aggregation of Gold Nanorods Mediated by Optical Forces and Plasmonic Heating. Sci. Rep. 2016, 6, 26952. [Google Scholar] [CrossRef] [PubMed]
- Kneipp, K.; Wang, Y.; Kneipp, H.; Perelman, L.T.; Itzkan, I.; Dasari, R.R.; Feld, M.S. Single Molecule Detection Using Surface-Enhanced Raman Scattering (SERS). Phys. Rev. Lett. 1997, 78, 1667–1670. [Google Scholar] [CrossRef]
- D’Andrea, C.; Cazzaniga, F.A.; Bistaffa, E.; Barucci, A.; de Angelis, M.; Banchelli, M.; Farnesi, E.; Polykretis, P.; Marzi, C.; Indaco, A.; et al. Impact of Seed Amplification Assay and Surface-Enhanced Raman Spectroscopy Combined Approach on the Clinical Diagnosis of Alzheimer’s Disease. Transl. Neurodegener. 2023, 12, 35. [Google Scholar] [CrossRef]
- Srivastava, S.; Wang, W.; Zhou, W.; Jin, M.; Vikesland, P.J. Machine Learning-Assisted Surface-Enhanced Raman Spectroscopy Detection for Environmental Applications: A Review. Environ. Sci. Technol. 2024, 58, 20830–20848. [Google Scholar] [CrossRef]
- Morganti, D.; Rizzo, M.G.; Spata, M.O.; Guglielmino, S.; Fazio, B.; Battiato, S.; Conoci, S. Temporal Convolutional Network on Raman Shift for Human Osteoblast Cells Fingerprint Analysis. Intell.-Based Med. 2024, 10, 100183. [Google Scholar] [CrossRef]
- Dorofeeva, Y.; Colombo, P.; Blanca, M.; Mari, A.; Khanferyan, R.; Valenta, R.; Focke-Tejkl, M. Expression and Characterization of Recombinant Par j 1 and Par j 2 Resembling the Allergenic Epitopes of Parietaria judaica Pollen. Sci. Rep. 2019, 9, 15043. [Google Scholar] [CrossRef]
- Faro, M.J.L.; D’andrea, C.; Leonardi, A.A.; Morganti, D.; Irrera, A.; Fazio, B. Fractal Silver Dendrites as 3D SERS Platform for Highly Sensitive Detection of Biomolecules in Hydration Conditions. Nanomaterials 2019, 9, 1630. [Google Scholar] [CrossRef]
- Licciardi, M.; Montana, G.; Bondì, M.L.; Bonura, A.; Scialabba, C.; Melis, M.; Fiorica, C.; Giammona, G.; Colombo, P. An Allergen-Polymeric Nanoaggregate as a New Tool for Allergy Vaccination. Int. J. Pharm. 2014, 465, 275–283. [Google Scholar] [CrossRef]
- Chen, C.-Y.; Hsiao, P.-H. Silver-Assisted Chemical Etching on Silicon with Polyvinylpyrrolidone-Mediated Formation of Silver Dendrites. ChemPhysChem 2015, 16, 540–545. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, G.; De Luca, G.; Franco, D.; Morganti, D.; Rizzo, M.G.; Bonavita, A.; Neri, G.; Fazio, E.; Neri, F.; Fazio, B.; et al. Structural and Antibacterial Studies of Novel ZnO and ZnxMn(1−x)O Nanostructured Titanium Scaffolds for Biomedical Applications. Biomater. Adv. 2023, 145, 213193. [Google Scholar] [CrossRef] [PubMed]
- Stumvoll, S.; Westritschnig, K.; Lidholm, J.; Spitzauer, S.; Colombo, P.; Duro, G.; Kraft, D.; Geraci, D.; Valenta, R. Identification of Cross-Reactive and Genuine Parietaria judaica Pollen Allergens. J. Allergy Clin. Immunol. 2003, 111, 974–979. [Google Scholar] [CrossRef]
- Rygula, A.; Majzner, K.; Marzec, K.M.; Kaczor, A.; Pilarczyk, M.; Baranska, M. Raman Spectroscopy of Proteins: A Review. J. Raman Spectrosc. 2013, 44, 1061–1076. [Google Scholar] [CrossRef]
- Jenkins, A.L.; Larsen, R.A.; Williams, T.B. Characterization of Amino Acids Using Raman Spectroscopy. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2005, 61, 1585–1594. [Google Scholar] [CrossRef]
- Lo Faro, M.J.; Ruello, G.; Leonardi, A.A.; Morganti, D.; Irrera, A.; Priolo, F.; Gigan, S.; Volpe, G.; Fazio, B. Visualization of Directional Beaming of Weakly Localized Raman from a Random Network of Silicon Nanowires. Adv. Sci. 2021, 8, 2100139. [Google Scholar] [CrossRef]
- Benevides, J.M.; Overman, S.A.; Thomas, G.J., Jr. Raman Spectroscopy of Proteins. Curr. Protoc. Protein Sci. 2003, 33, 17.8.1–17.8.35. [Google Scholar] [CrossRef]
- Maiti, N.C.; Apetri, M.M.; Zagorski, M.G.; Carey, P.R.; Anderson, V.E. Raman Spectroscopic Characterization of Secondary Structure in Natively Unfolded Proteins: α-Synuclein. J. Am. Chem. Soc. 2004, 126, 2399–2408. [Google Scholar] [CrossRef]
- Zhu, G.; Zhu, X.; Fan, Q.; Wan, X. Raman Spectra of Amino Acids and Their Aqueous Solutions. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2011, 78, 1187–1195. [Google Scholar] [CrossRef]
- Kuhar, N.; Sil, S.; Umapathy, S. Potential of Raman Spectroscopic Techniques to Study Proteins. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2021, 258, 119712. [Google Scholar] [CrossRef]
- Kolasinski, K.W.; Tamarov, K.; Swanson, J.D.; Unger, B.A.; Ernst, A.T.; Aindow, M.; Kiviluoto, R.; Lehto, V.-P.; Riikonen, J. Injection Metal-Assisted Catalytic Etching (MACE) of Si Powder: Discovery of Low-Load MACE and Pore Distribution Tunability Using Ag, Au, Pd, Pt and Cu Catalysts. Meet. Abstr. 2020, MA2020-02, 1219. [Google Scholar] [CrossRef]
- Peng, K.; Fang, H.; Hu, J.; Wu, Y.; Zhu, J.; Yan, Y.; Lee, S. Metal-Particle-Induced, Highly Localized Site-Specific Etching of Si and Formation of Single-Crystalline Si Nanowires in Aqueous Fluoride Solution. Chem. A Eur. J. 2006, 12, 7942–7947. [Google Scholar] [CrossRef] [PubMed]
- Morganti, D.; Leonardi, A.A.; Lo Faro, M.J.; Leonardi, G.; Salvato, G.; Fazio, B.; Musumeci, P.; Livreri, P.; Conoci, S.; Neri, G.; et al. Ultrathin Silicon Nanowires for Optical and Electrical Nitrogen Dioxide Detection. Nanomaterials 2021, 11, 1767. [Google Scholar] [CrossRef]
- Huang, Z.; Geyer, N.; Werner, P.; de Boor, J.; Gösele, U. Metal-Assisted Chemical Etching of Silicon: A Review. Adv. Mater. 2011, 23, 285–308. [Google Scholar] [CrossRef]
- Morganti, D.; Faro, M.J.L.; Leonardi, A.A.; Fazio, B.; Conoci, S.; Irrera, A. Luminescent Silicon Nanowires as Novel Sensor for Environmental Air Quality Control. Sensors 2022, 22, 8755. [Google Scholar] [CrossRef]
- Leonardi, A.A.; Sciuto, E.L.; Lo Faro, M.J.; Morganti, D.; Midiri, A.; Spinella, C.; Conoci, S.; Irrera, A.; Fazio, B. Molecular Fingerprinting of the Omicron Variant Genome of SARS-CoV-2 by SERS Spectroscopy. Nanomaterials 2022, 12, 2134. [Google Scholar] [CrossRef]
- McPeak, K.M.; Jayanti, S.V.; Kress, S.J.P.; Meyer, S.; Iotti, S.; Rossinelli, A.; Norris, D.J. Plasmonic Films Can Easily Be Better: Rules and Recipes. ACS Photonics 2015, 2, 326–333. [Google Scholar] [CrossRef]
- Polyanskiy, M.N. Refractiveindex.Info Database of Optical Constants. Sci. Data 2024, 11, 94. [Google Scholar] [CrossRef] [PubMed]
- Baronio, C.M.; Barth, A. The Amide I Spectrum of Proteins—Optimization of Transition Dipole Coupling Parameters Using Density Functional Theory Calculations. J. Phys. Chem. B 2020, 124, 1703–1714. [Google Scholar] [CrossRef]
- Van Wart, H.E.; Lewis, A.; Scheraga, H.A.; Saeva, F.D. Disulfide Bond Dihedral Angles from Raman Spectroscopy. Proc. Natl. Acad. Sci. USA 1973, 70, 2619–2623. [Google Scholar] [CrossRef]
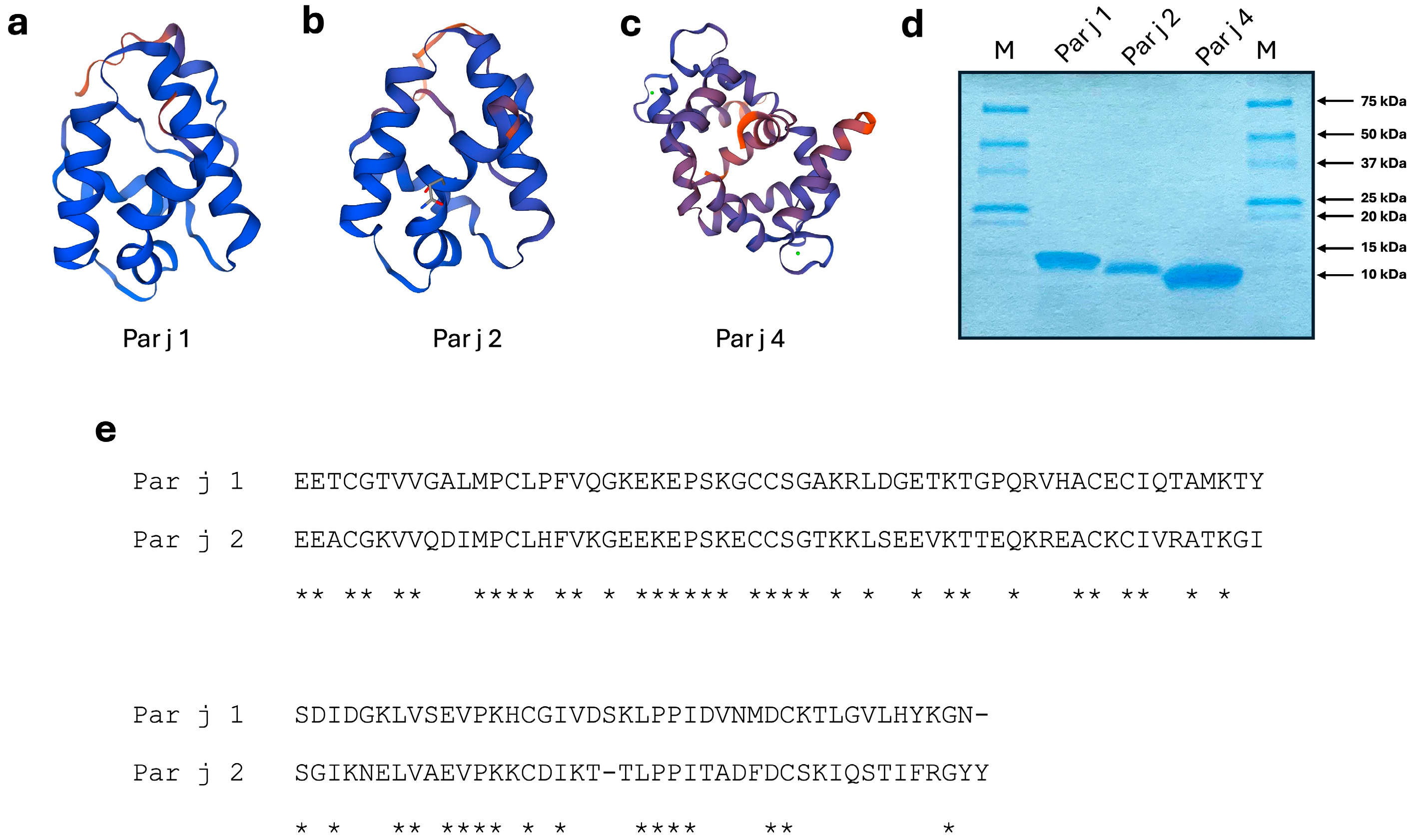

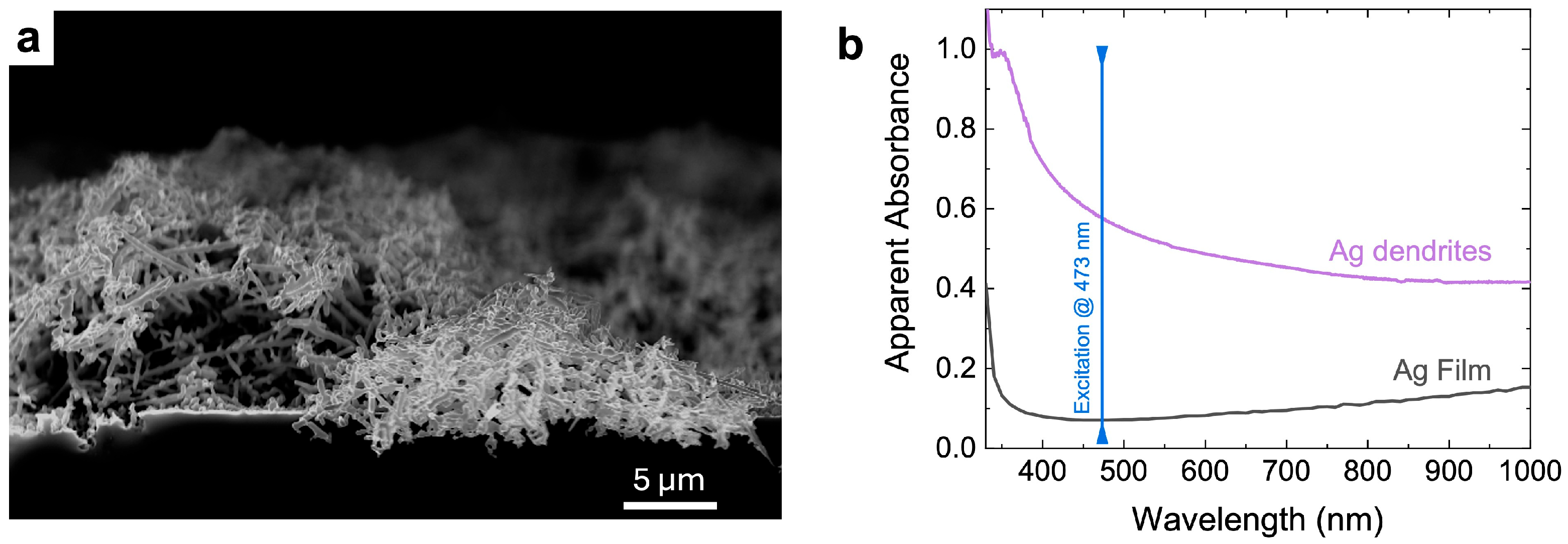
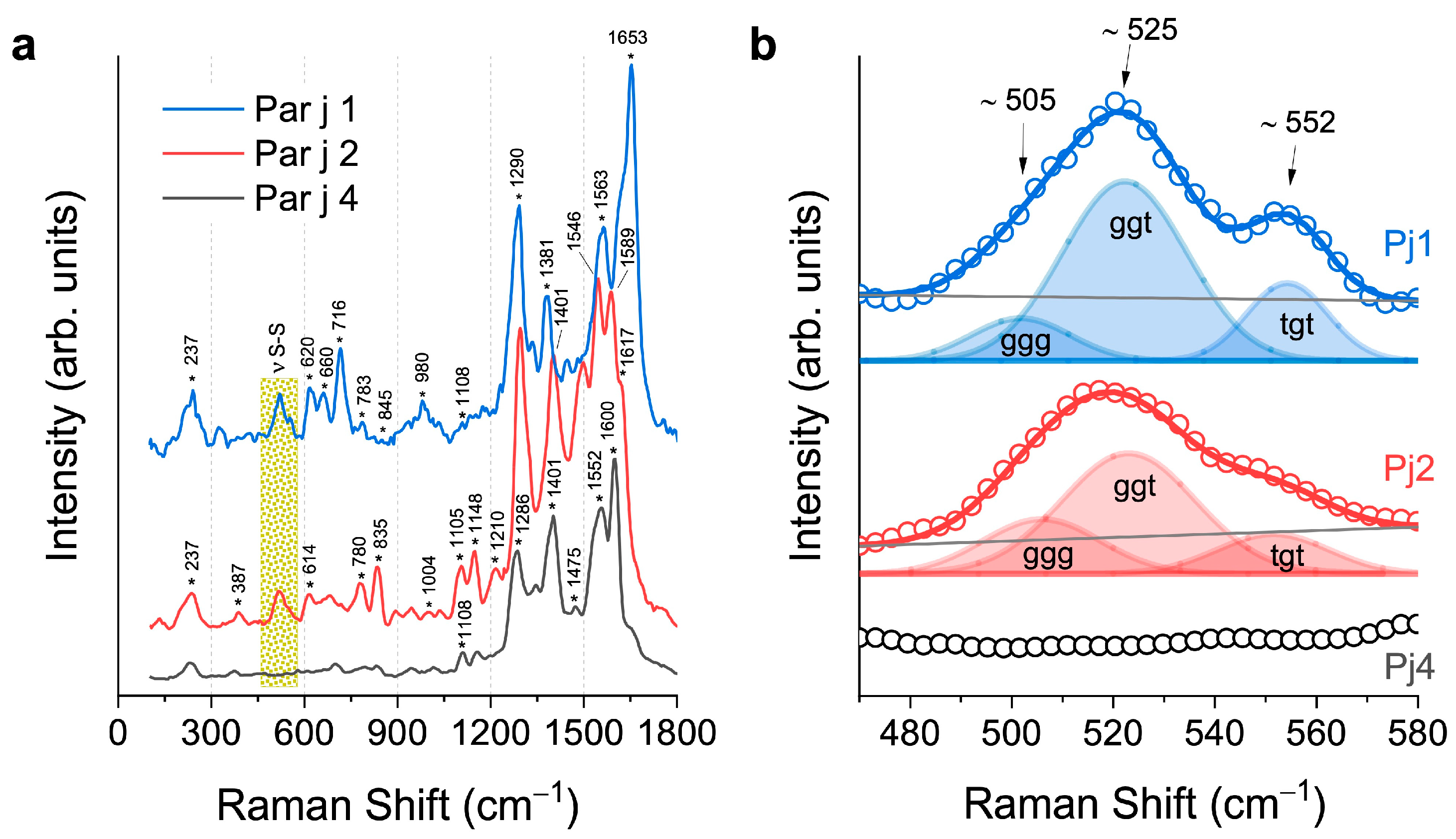

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Frequency (cm–1) | Mode Assignment | Reference | Protein |
|---|---|---|---|
| 511 | CaF2 | Par j 1, 2, 4 | |
| 560 | Ile, Thr | [31] | Par j 4 |
| 645 | OCN bending NH bending out-of-plane | [32] | Par j 1, 2 |
| 808 | CaF2 | Par j 1, 2 | |
| 929–935 | C-C stretching CaF2, PBS | [15] | Par j 1, 2, 4 |
| 1000 | Phe ring (C-C) breathing mode | [10,15,31,32] | Par j 1, 2 |
| 1061 | C-C and CN stretching Lys (C wagging) | [31] | Par j 4 |
| 1114–1125 | CN stretching | [15] | Par j 1, 2, 4 |
| 1237–1338 | CH2 twisting, CH bending Amide III | [15,31,32] | Par j 1, 2, 4 |
| 1450 | CH2 and CH3 scissoring | [31] | Par j 1, 2, 4 |
| 1611 | Phe Ring stretching | [10] | Par j 1, 2, 4 |
| 1659 | Amide I | [31] | Par j 1, 2, 4 |
| Parietaria judaica 1 | ||
|---|---|---|
| Frequency (cm–1) | Mode Assignment | |
| 237 | Ag-O Str. | |
| 508–557 | S-S str.(disulfide bonds) | [31,32] |
| 520 | Si-Si sym str, | |
| 620 | Phe COO- deformation | [31] |
| 625–800 | ring (C-C) deformation, OCN bend, NH bend out-of-plane | [31,32] |
| 715–730 | Tyr | [15,31] |
| 783 | Lys | [15] |
| 845 | Leu Lys (Cγ twist., Cε wagg., Cβ wagg.) | [15] |
| 850 | Val His (C-H out of plane) Leu Ala Nt–Cα; O–Ct–O; Cα–Ct–O | [15] |
| 897 | CC stretching or CCNsym stretching | [31] |
| 931 | Ile Arg (Nη1–Cζ–Nη2 sym st; Cδ–Nε; Cβ–Cγ; Nε–Cζ) | [15] |
| 965 | CC stretching or CCNsym stretching | [31] |
| 982 | His Lys | [15] |
| 1010 | Phe ring (C-C) breathing | [10,15,31,32] |
| 1108 | His (NtH3+ asym. rock.) Thr | [15] |
| 1144 | Lys (NtH3+ asym rock; Ct Cα-Hα) | [15] |
| 1172 | CN str | [31] |
| 1268–1400 | Amide III | [15,31] |
| 1316 | CH2 twst., CH bend.; His (NtH3+ asym. rock, Cβ-rock., Ct-Cα-H) | [15,31] |
| 1330 | CH2 twst., CH bend. | [31] |
| 1438–1467 | CH2 and CH3 scissoring | [31] |
| 1480–1580 | Amide II | [15,32] |
| 1600–1660 | Amide I | [31] |
| Parietaria judaica 2 | ||
| Frequency (cm–1) | Mode Assignment | |
| 237 | Ag-O Str. | |
| 508–557 | S-S str.(disulfide bonds) | [31,32] |
| 630–800 | OCN bend, NH bend out-of-plane | [32] |
| 780 | Lys | [15] |
| 835 | Phe, Tyr | [15] |
| 890–980 | CC stretching or CNsym stretching | [32] |
| 1085 | CN str. | [31] |
| 1124 | Val, Ser | [15] |
| 1148 | His Lys (NtH3+ asym rock; Ct Cα-Hα) | [15] |
| 1162 | Lys | [15] |
| 1210 | Lys, Tyr, Phe | [15,31] |
| 1220–1400 | Amide III | [32] |
| 1320 | CH2 twst., CH bend.; His (NtH3+ asym. rock, Cβ-rock., Ct-Cα-H) | [15,31] |
| 1340 | CH2 twst., CH bend. | [31] |
| 1386 | Cys | [15] |
| 1401 | His (Nt–Cα–Hα, Cβ–Cα–H, Cβ-rock) | [15] |
| 1480–1580 | Amide II | [15,32] |
| 1589 | Phe | [31] |
| 1600 | Tyr + Phe ring stretching | [31] |
| 1617 | Phe ring stretching | [10] |
| 1600–1660 | Amide I | [31] |
| Parietaria judaica 4 | ||
| Frequency (cm–1) | Mode Assignment | |
| 237 | Ag-O Str. | |
| 792 | NH bend out-of-plane | [32] |
| 810 | Ser, Met | [15] |
| 835 | Phe | [15] |
| 860 | Ala (Nt–Cα; O–Ct–O; Cα–Ct–O sym bend; Cα–Ct) Arg (Cζ–Nη2) Ser | [15] |
| 910–950 | CC stretching or CNsym stretching | [31,32] |
| 1004 | Phe ring (C-C) breathing | [10,15,31,32] |
| 1100 | NH3+ rocking Lys (Cε–Nζ; Cı–Cε) Arg (Cγ–Cδ) Gly (N–Cα) | [15,31] |
| 1225–1275 | Amide III | [31,32] |
| 1300–1350 | CH def., CH2 twst., CH bend. | [15,32] |
| 1401 | His (Nt–Cα–Hα, Cβ–Cα–H, Cβ-rock) | [15] |
| 1475 | CH2 and CH3 scissoring | [31,32] |
| 1482–1533 | Amide II | [15,32] |
| 1552 | Amide II | [15] |
| 1600–1660 | Amide I | [15] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morganti, D.; Longo, V.; Leonardi, A.A.; Irrera, A.; Colombo, P.; Fazio, B. First Vibrational Fingerprint of Parietaria judaica Protein via Surface-Enhanced Raman Spectroscopy. Biosensors 2025, 15, 182. https://doi.org/10.3390/bios15030182
Morganti D, Longo V, Leonardi AA, Irrera A, Colombo P, Fazio B. First Vibrational Fingerprint of Parietaria judaica Protein via Surface-Enhanced Raman Spectroscopy. Biosensors. 2025; 15(3):182. https://doi.org/10.3390/bios15030182
Chicago/Turabian StyleMorganti, Dario, Valeria Longo, Antonio Alessio Leonardi, Alessia Irrera, Paolo Colombo, and Barbara Fazio. 2025. "First Vibrational Fingerprint of Parietaria judaica Protein via Surface-Enhanced Raman Spectroscopy" Biosensors 15, no. 3: 182. https://doi.org/10.3390/bios15030182
APA StyleMorganti, D., Longo, V., Leonardi, A. A., Irrera, A., Colombo, P., & Fazio, B. (2025). First Vibrational Fingerprint of Parietaria judaica Protein via Surface-Enhanced Raman Spectroscopy. Biosensors, 15(3), 182. https://doi.org/10.3390/bios15030182