Engineering of a Bacterial Biosensor for the Detection of Chlorate in Food


Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals Reagents and Strains
2.2. Chlorate Induction Assays
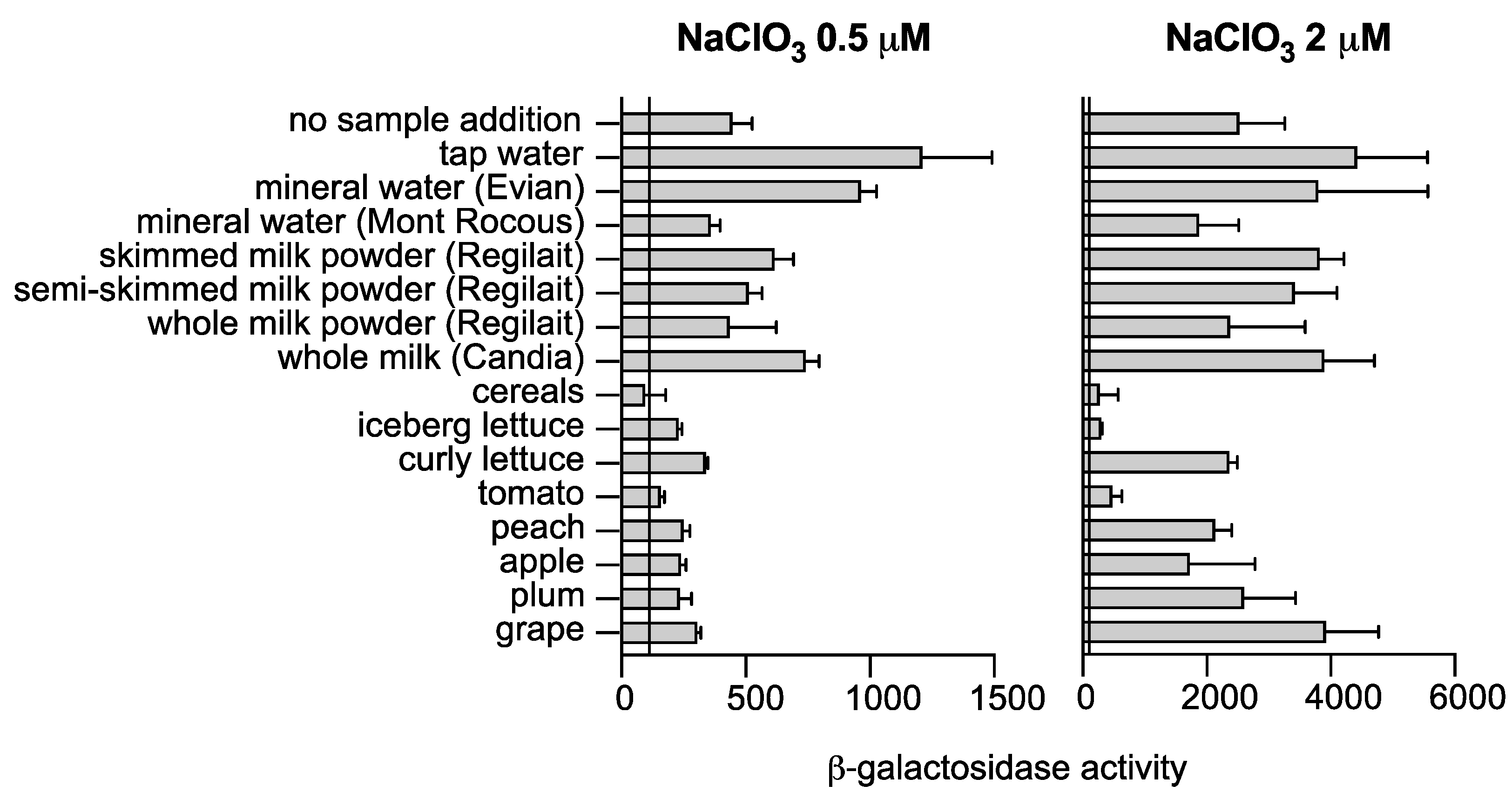
2.3. Chlorate Analysis in Food Samples
2.4. Statistical Analysis
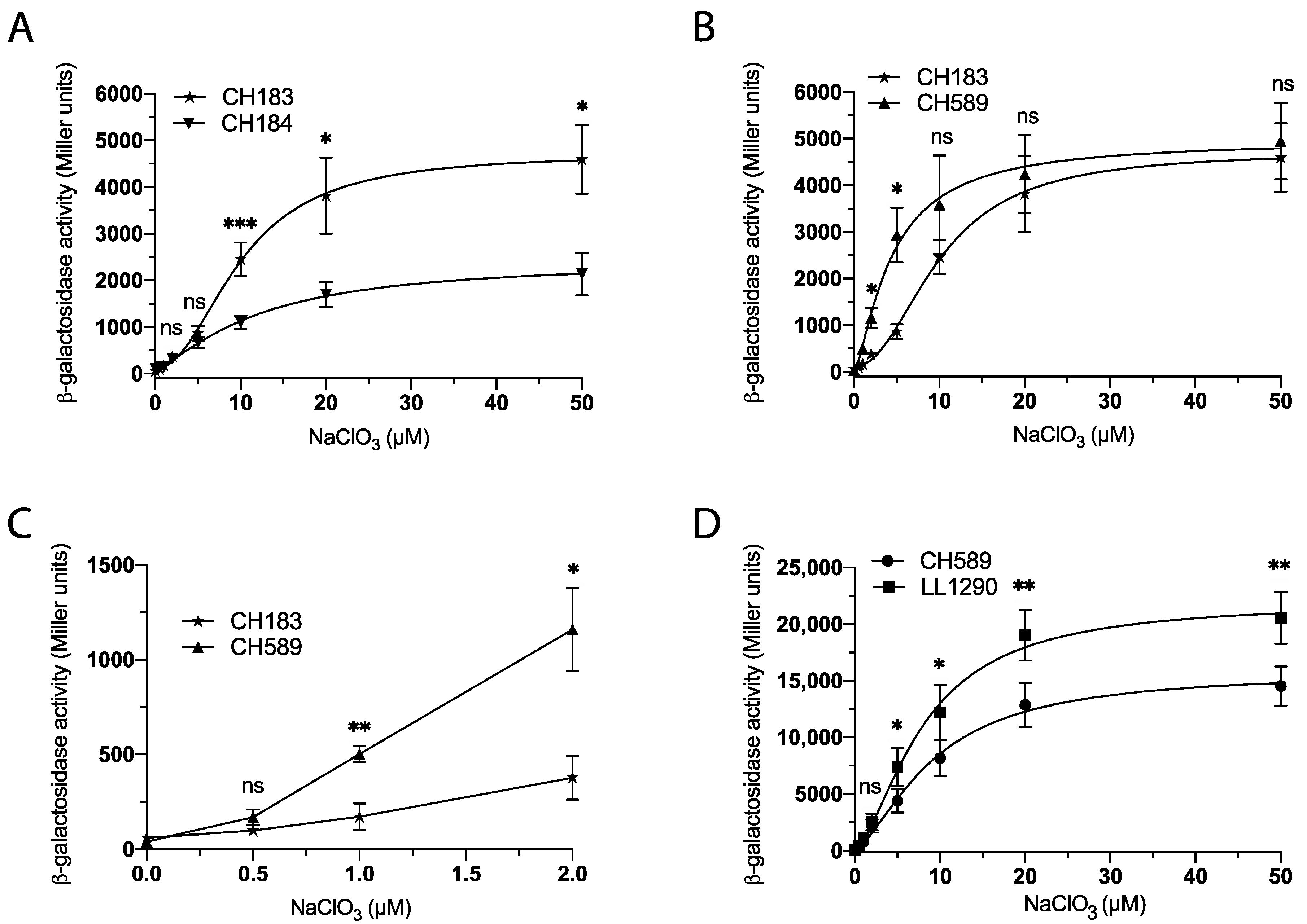
3. Results
3.1. M9 Minimal Medium Enhances Chlorate Detection and Reduces Nitrate Interference
3.2. Optimizing Chlorate Detection in E. coli Using Synthetic Biology
3.3. Optimizing Chlorate Detection in E. coli by Modifying the Culture Medium
3.4. Validation of a Bacterial Biosensor for the Detection of Chlorate in Food
4. Discussion
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Patents
References
- Risks for Public Health Related to the Presence of Chlorate in Food. EFSA J. 2015, 13, 4135. [CrossRef]
- Steffen, C.; Wetzel, E. Chlorate Poisoning: Mechanism of Toxicity. Toxicology 1993, 84, 217–231. [Google Scholar] [CrossRef] [PubMed]
- Youngblut, M.D.; Wang, O.; Barnum, T.P.; Coates, J.D. (Per)Chlorate in Biology on Earth and Beyond. Annu. Rev. Microbiol. 2016, 70, 435–457. [Google Scholar] [CrossRef] [PubMed]
- Mavroudakis, L.; Mavrakis, E.; Kouvarakis, A.; Pergantis, S.A. Determination of Chlorate, Perchlorate and Bromate Anions in Water Samples by Microbore Reversed-Phase Liquid Chromatography Coupled to Sonic-Spray Ionization Mass Spectrometry. Rapid Commun. Mass Spectrom. 2017, 31, 911–918. [Google Scholar] [CrossRef]
- Shriver-Lake, L.; Zabetakis, D.; Dressick, W.; Stenger, D.; Trammell, S. Paper-Based Electrochemical Detection of Chlorate. Sensors 2018, 18, 328. [Google Scholar] [CrossRef]
- Heinnickel, M.; Smith, S.C.; Koo, J.; O’Connor, S.M.; Coates, J.D. A Bioassay for the Detection of Perchlorate in the Ppb Range. Environ. Sci. Technol. 2011, 45, 2958–2964. [Google Scholar] [CrossRef]
- Alsaleh, S.A.; Barron, L.; Sturzenbaum, S. Perchlorate Detection via an Invertebrate Biosensor. Anal. Methods 2021, 13, 327–336. [Google Scholar] [CrossRef]
- Balabanov, V.P.; Khrulnova, S.A.; Kotova, V.Y.; Zavilgelsky, G.B. Ammonium Perchlorate Detection in Natural Environments Using Specific Lux Biosensors. Russ. J. Phys. Chem. B 2017, 11, 663–665. [Google Scholar] [CrossRef]
- Alefounder, P.R.; Ferguson, S.J. The Location of Dissimilatory Nitrite Reductase and the Control of Dissimilatory Nitrate Reductase by Oxygen in Paracoccus denitrificans. Biochem. J. 1980, 192, 231–240. [Google Scholar] [CrossRef]
- Giordano, G.; Violet, M.; Medani, C.; Pommier, J. A Common Pathway for the Activation of Several Molybdoenzymes in Escherichia coli K12. Biochim. Biophys. Acta (BBA)—Gen. Subj. 1984, 798, 216–225. [Google Scholar] [CrossRef]
- Glaser, J.H.; DeMoss, J.A. Phenotypic Restoration by Molybdate of Nitrate Reductase Activity in ChlD Mutants of Escherichia coli. J. Bacteriol. 1971, 108, 854–860. [Google Scholar] [CrossRef]
- Stewart, V. Nitrate Respiration in Relation to Facultative Metabolism in Enterobacteria. Microbiol. Rev. 1988, 52, 190–232. [Google Scholar] [CrossRef]
- Ezraty, B.; Gennaris, A.; Barras, F.; Collet, J.-F. Oxidative Stress, Protein Damage and Repair in Bacteria. Nat. Rev. Microbiol. 2017, 15, 385–396. [Google Scholar] [CrossRef]
- Aussel, L.; Ezraty, B. Methionine Redox Homeostasis in Protein Quality Control. Front. Mol. Biosci. 2021, 8, 665492. [Google Scholar] [CrossRef]
- Loiseau, L.; Vergnes, A.; Ezraty, B. Methionine Oxidation under Anaerobic Conditions in Escherichia coli. Mol. Microbiol. 2022, 118, 387–402. [Google Scholar] [CrossRef]
- Spero, M.A.; Jones, J.; Lomenick, B.; Chou, T.; Newman, D.K. Mechanisms of Chlorate Toxicity and Resistance in Pseudomonas Aeruginosa. Mol. Microbiol. 2022, 118, 321–335. [Google Scholar] [CrossRef]
- Vincent, M.S.; Vergnes, A.; Ezraty, B. Chlorate Contamination in Commercial Growth Media as a Source of Phenotypic Heterogeneity within Bacterial Populations. Microbiol. Spectr. 2023, 11, e0499122. [Google Scholar] [CrossRef]
- Thomason, L.C.; Costantino, N.; Court, D.L. E. Coli Genome Manipulation by P1 Transduction. Curr. Protoc. Mol. Biol. 2007, 79, 1.17.1–1.17.8. [Google Scholar] [CrossRef]
- Datsenko, K.A.; Wanner, B.L. One-Step Inactivation of Chromosomal Genes in Escherichia coli K-12 Using PCR Products. Proc. Natl. Acad. Sci. USA 2000, 97, 6640–6645. [Google Scholar] [CrossRef]
- El Hajj, S.; Henry, C.; Andrieu, C.; Vergnes, A.; Loiseau, L.; Brasseur, G.; Barré, R.; Aussel, L.; Ezraty, B. HprSR Is a Reactive Chlorine Species-Sensing, Two-Component System in Escherichia coli. J. Bacteriol. 2022, 204, e0044921. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.H. A Short Course in Bacterial Genetics: A Laboratory Manual and Handbook for Escherichia coli and Related Bacteria.; Cold Spring Harbor Laboratory Press: Long Island, NY, USA, 1992; ISBN 0879693495. [Google Scholar]
- Gennaris, A.; Ezraty, B.; Henry, C.; Agrebi, R.; Vergnes, A.; Oheix, E.; Bos, J.; Leverrier, P.; Espinosa, L.; Szewczyk, J.; et al. Repairing Oxidized Proteins in the Bacterial Envelope Using Respiratory Chain Electrons. Nature 2015, 528, 409–412. [Google Scholar] [CrossRef] [PubMed]
- Akboğa, D.; Saltepe, B.; Bozkurt, E.U.; Şeker, U.Ö.Ş. A Recombinase-Based Genetic Circuit for Heavy Metal Monitoring. Biosensors 2022, 12, 122. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wang, C.-W.; Wang, D.; Wei, N. A Whole-Cell Biosensor for Point-of-Care Detection of Waterborne Bacterial Pathogens. ACS Synth. Biol. 2021, 10, 333–344. [Google Scholar] [CrossRef]
- Fantino, J.-R.; Barras, F.; Denizot, F. Sposensor: A Whole-Bacterial Biosensor That Uses Immobilized Bacillus Subtilis Spores and a One-Step Incubation/Detection Process. Microb. Physiol. 2009, 17, 90–95. [Google Scholar] [CrossRef]
- Yamaji, K.; Taniguchi, R.; Urano, H.; Ogasawara, H. Roles of Methionine and Cysteine Residues of the Escherichia coli Sensor Kinase HprS in Reactive Chlorine Species Sensing. FEBS Lett. 2023, 597, 573–584. [Google Scholar] [CrossRef]
- Yancey, P.H.; Siebenaller, J.F. Co-Evolution of Proteins and Solutions: Protein Adaptation versus Cytoprotective Micromolecules and Their Roles in Marine Organisms. J. Exp. Biol. 2015, 218, 1880–1896. [Google Scholar] [CrossRef]
- Cheung, W.; Keski-Rahkonen, P.; Assi, N.; Ferrari, P.; Freisling, H.; Rinaldi, S.; Slimani, N.; Zamora-Ros, R.; Rundle, M.; Frost, G.; et al. A Metabolomic Study of Biomarkers of Meat and Fish Intake. Am. J. Clin. Nutr. 2017, 105, 600–608. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Genotype | Source |
|---|---|---|
| CH184 | MG1655 hiuH-lacZ (in lac locus) | El Hajj et al. (2022) [20] |
| CH183 | MG1655 msrP-lacZ (in lac locus) | El Hajj et al. (2022) [20] |
| CH589 | MG1655 ∆msrP KanR msrP-lacZ (in lac locus) | This study |
| LL1290 | MG1655 msrP-lacZ KanR (in hiuH-msrPQ locus) msrP-lacZ (in lac locus) | This study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vergnes, A.; Becam, J.; Loiseau, L.; Ezraty, B. Engineering of a Bacterial Biosensor for the Detection of Chlorate in Food. Biosensors 2023, 13, 629. https://doi.org/10.3390/bios13060629
Vergnes A, Becam J, Loiseau L, Ezraty B. Engineering of a Bacterial Biosensor for the Detection of Chlorate in Food. Biosensors. 2023; 13(6):629. https://doi.org/10.3390/bios13060629
Chicago/Turabian StyleVergnes, Alexandra, Jérôme Becam, Laurent Loiseau, and Benjamin Ezraty. 2023. "Engineering of a Bacterial Biosensor for the Detection of Chlorate in Food" Biosensors 13, no. 6: 629. https://doi.org/10.3390/bios13060629
APA StyleVergnes, A., Becam, J., Loiseau, L., & Ezraty, B. (2023). Engineering of a Bacterial Biosensor for the Detection of Chlorate in Food. Biosensors, 13(6), 629. https://doi.org/10.3390/bios13060629