Signal Enhancement in Oriented Immunosorbent Assays: A Balance between Accessibility of Antigen Binding Sites and Avidity

 , ,
, ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Antigens
2.3. Antibody Reduction Using 2-Mercaptoethylamine (2-MEA)
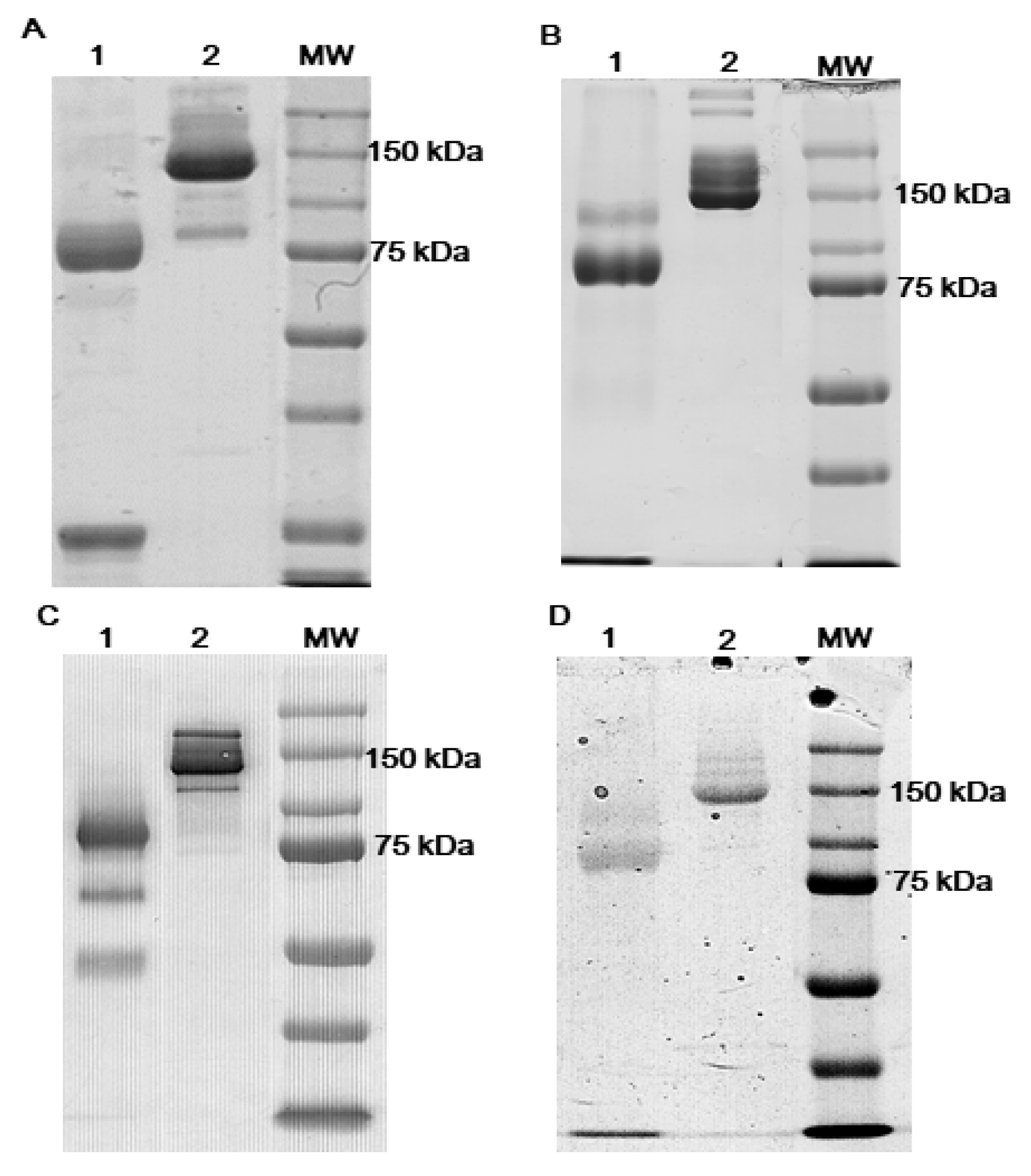
2.4. Native PAGE
2.5. ELISAs Performed with Oriented or Not Oriented Antibodies
3. Results
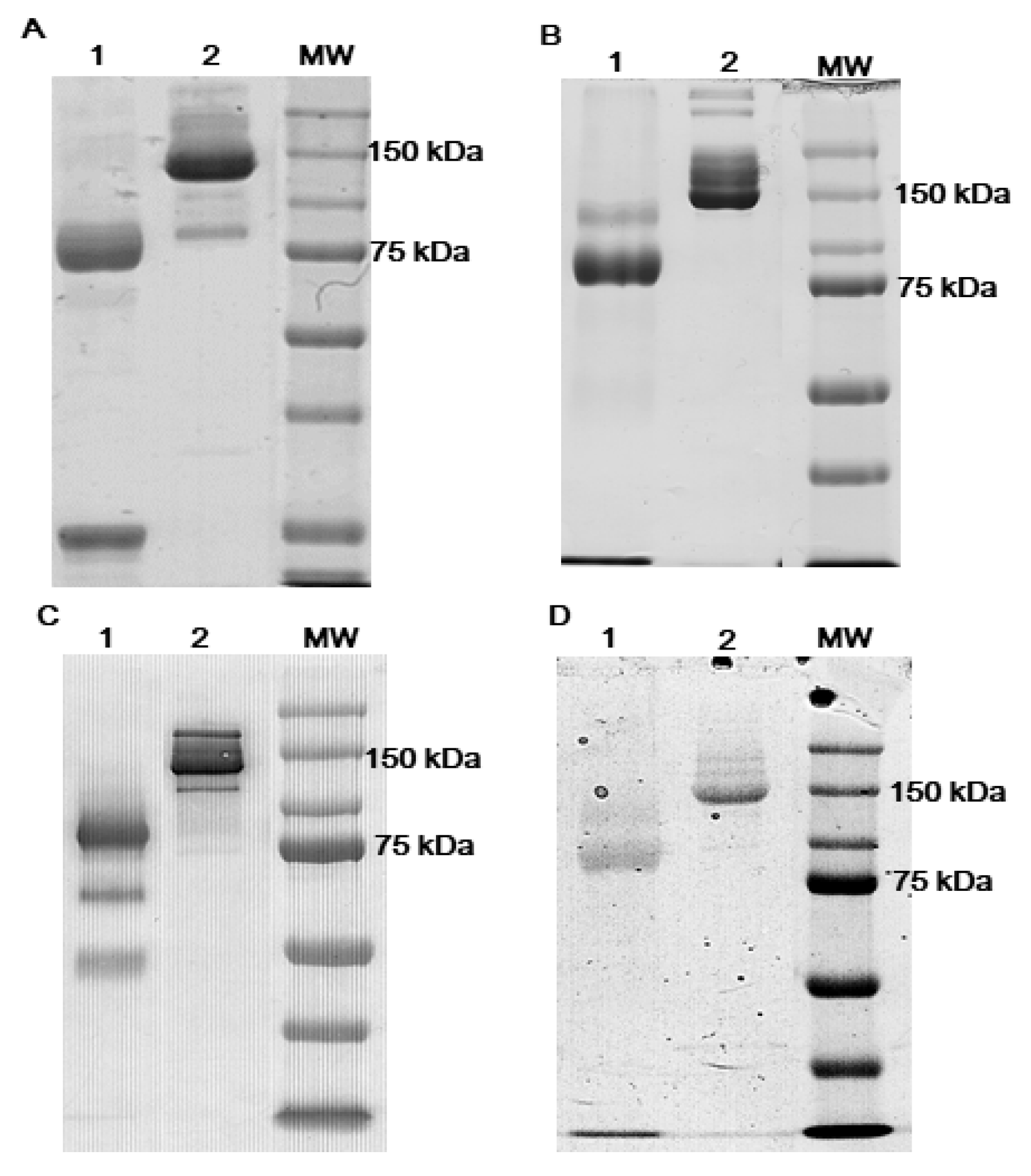
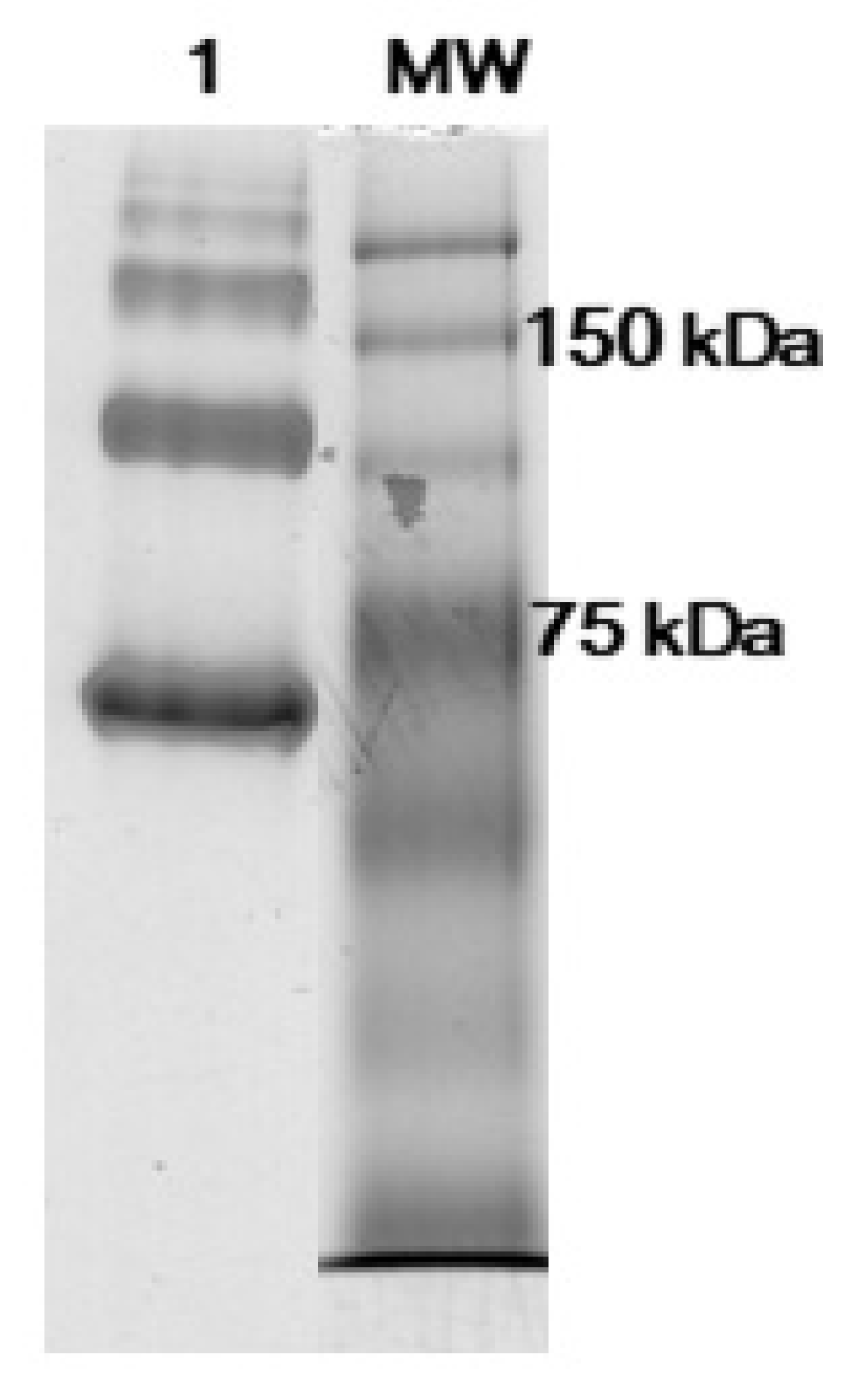
3.1. Antibodies Reduction
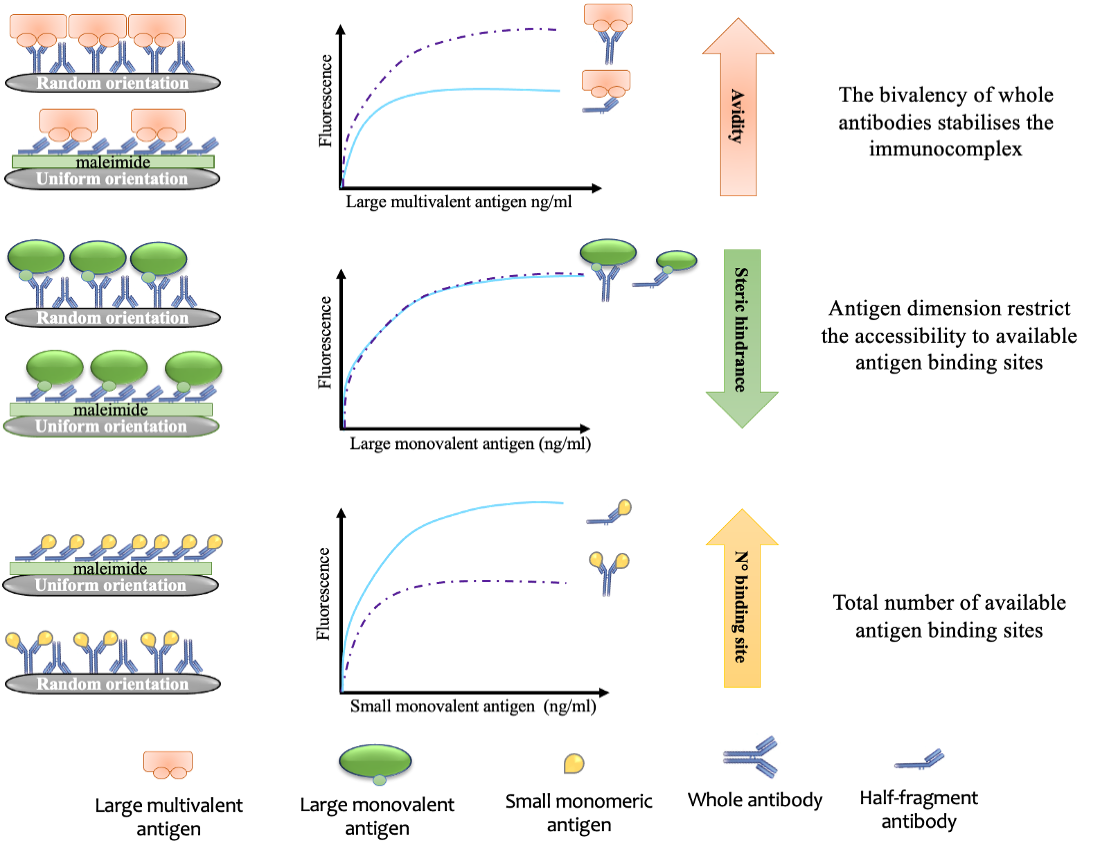
3.2. Signal Enhancement in Oriented ELISAs Using Monomeric Antigens
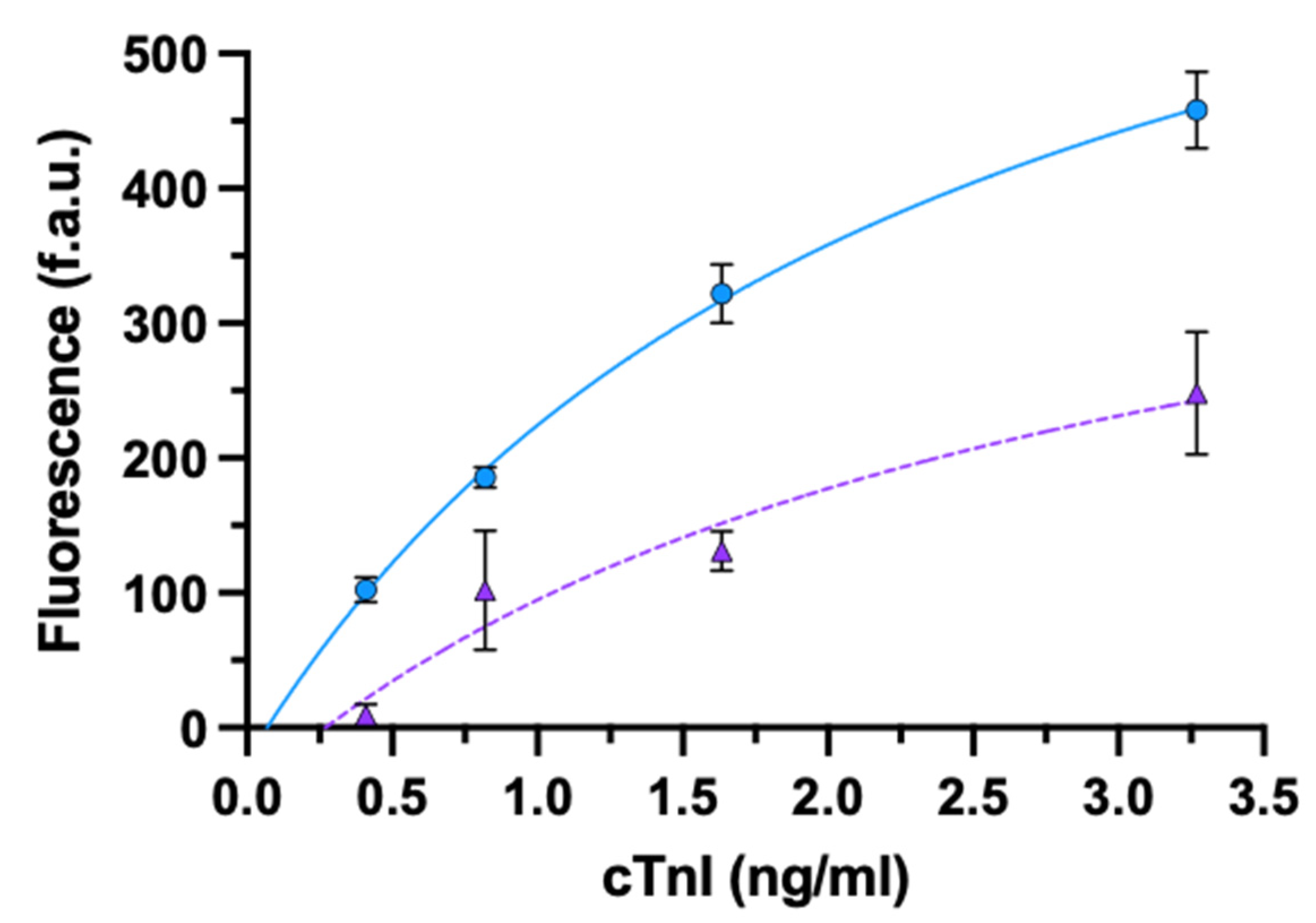
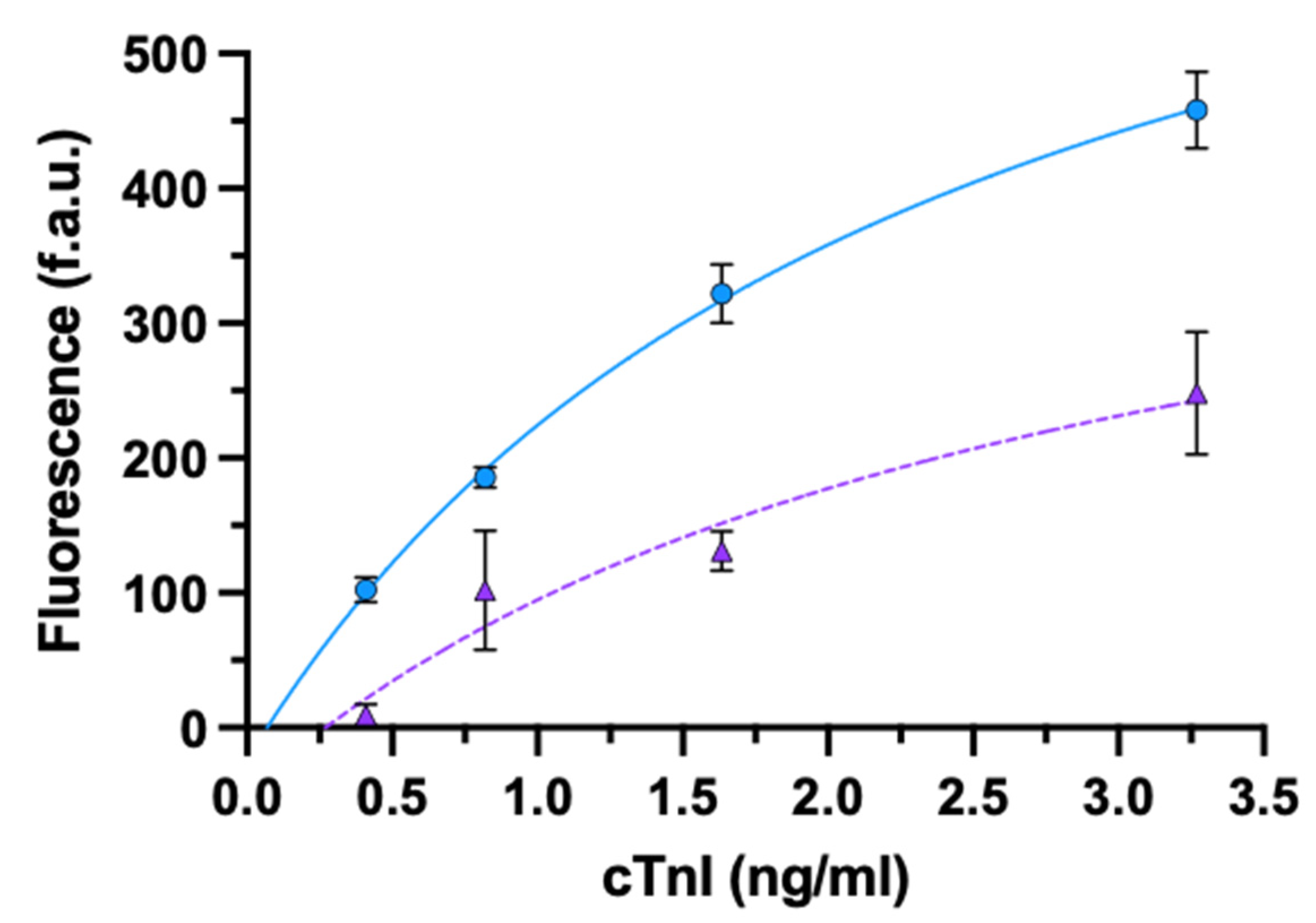
3.2.1. cTnI
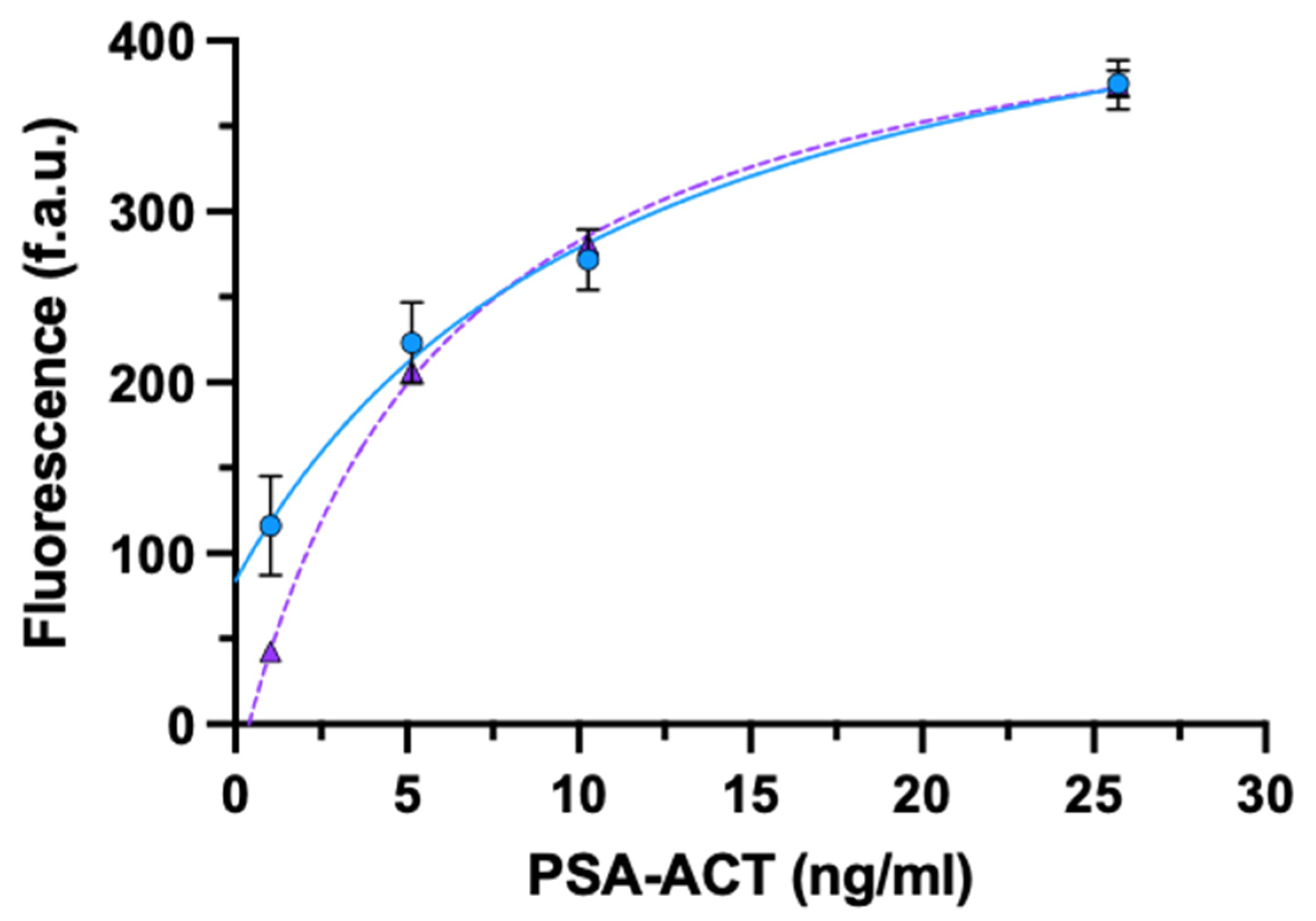
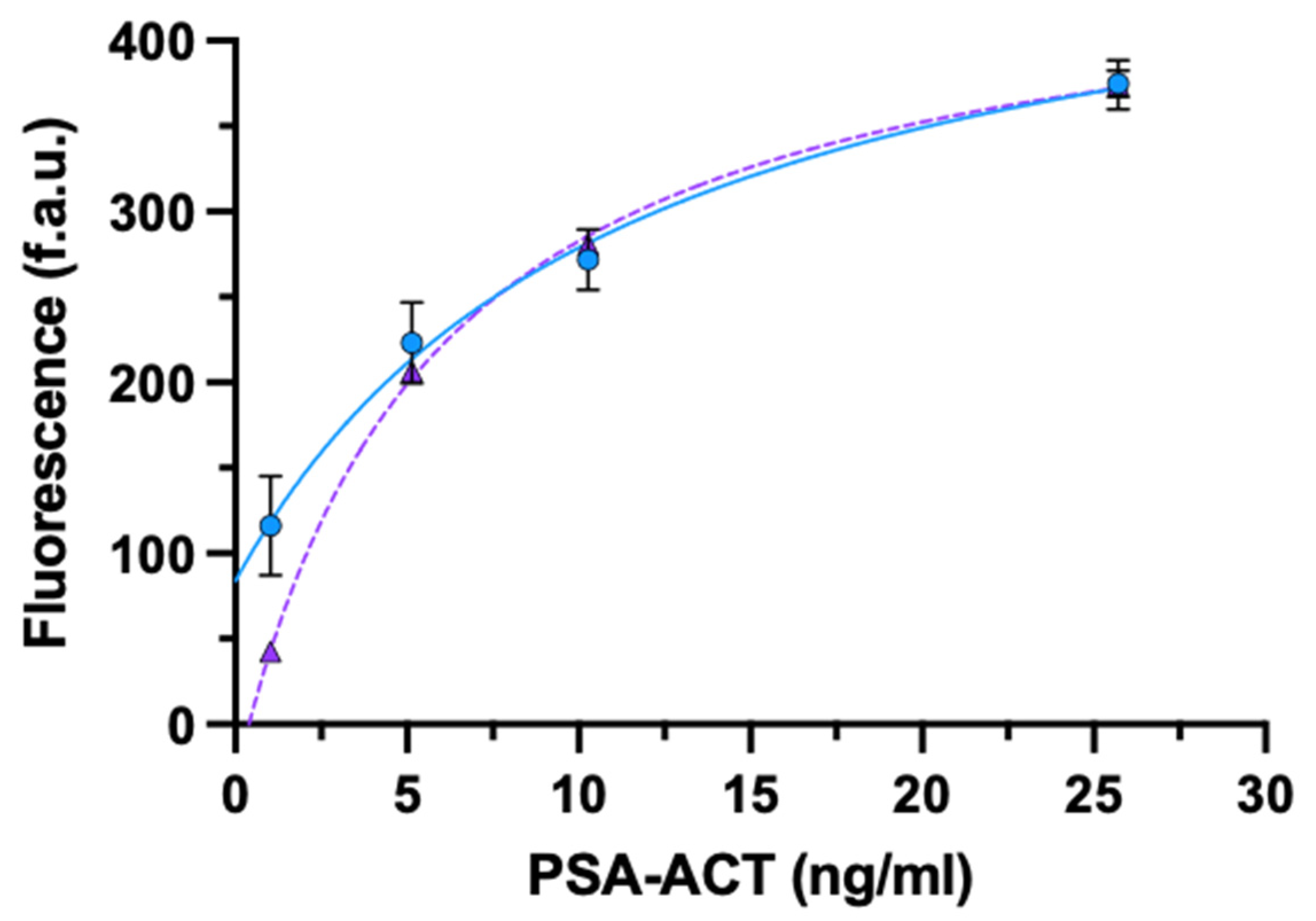
3.2.2. PSA-ACT
3.3. Signal Enhancement in Multimeric Antigens
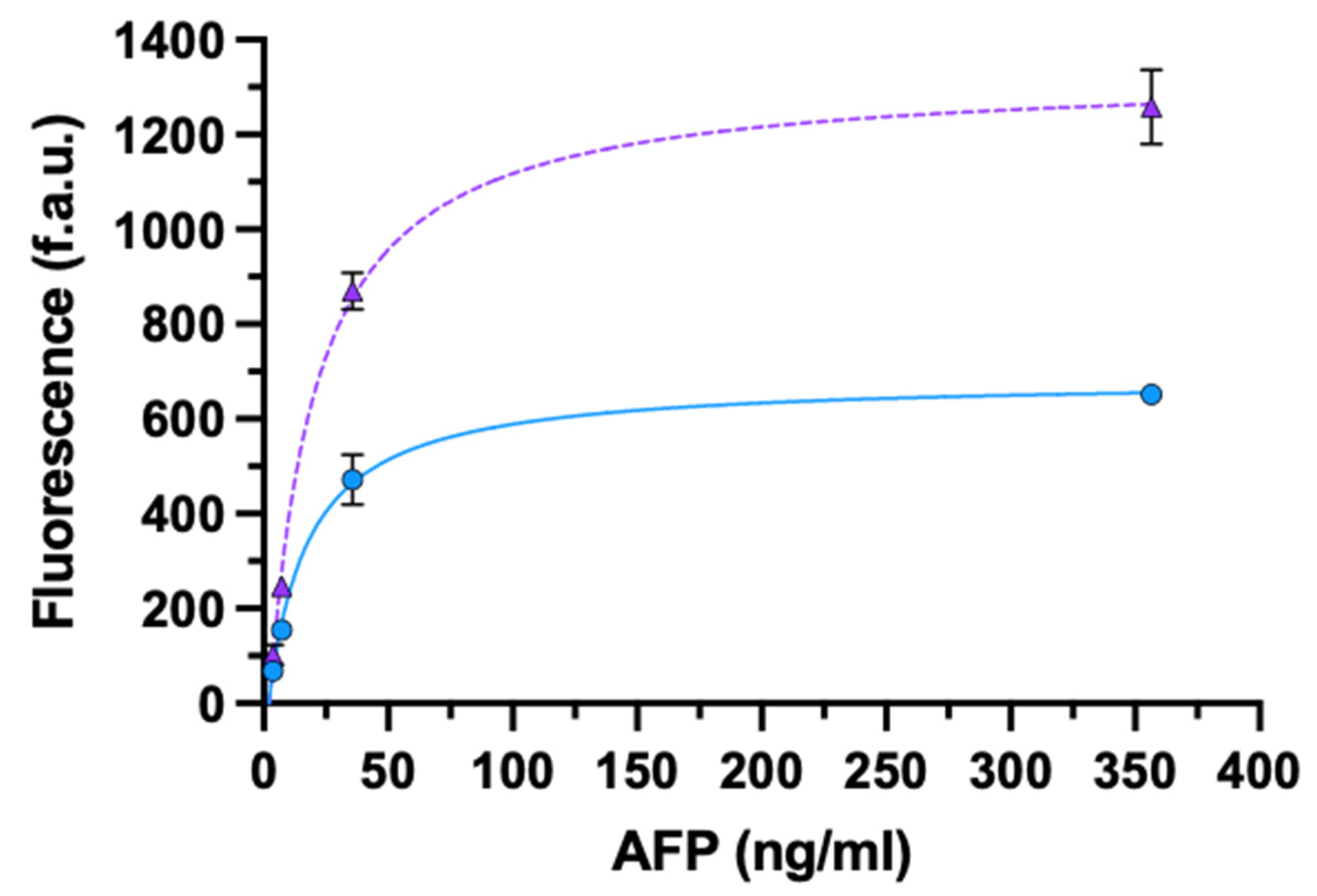
3.3.1. AFP
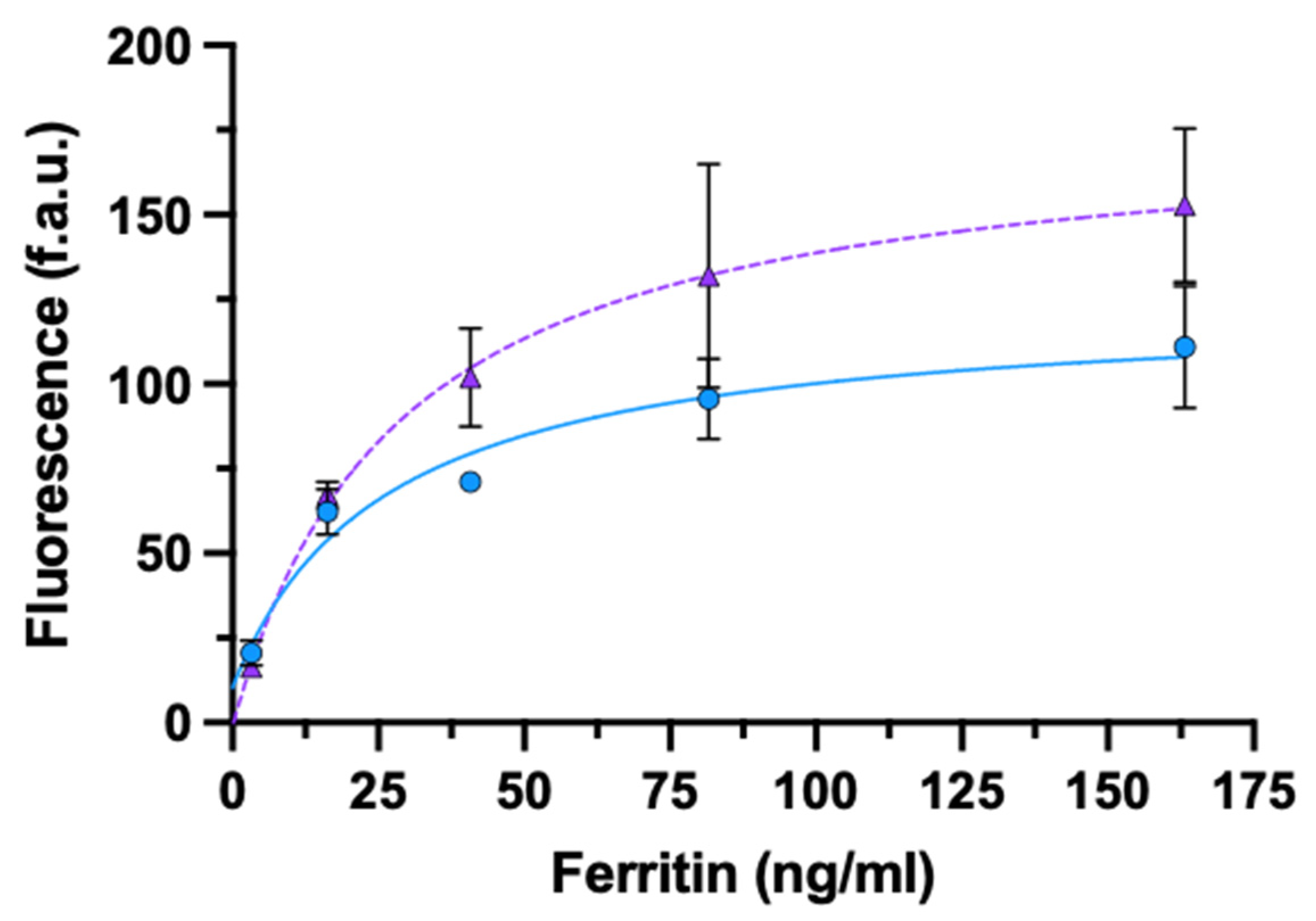
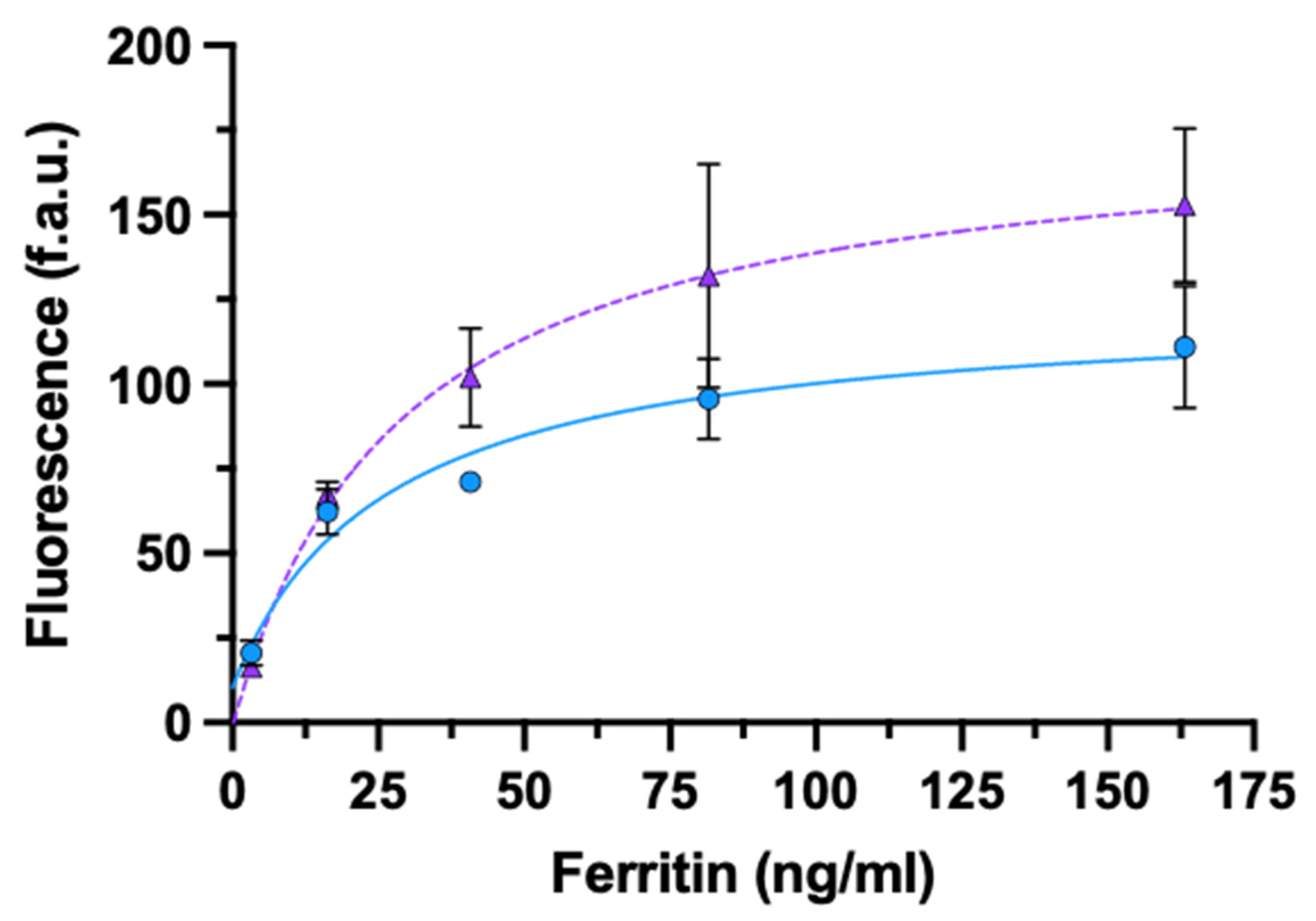
3.3.2. Ferritin
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cristea, C.; Florea, A.; Tertiș, M.; Săndulescu, R. Immunosensors. In Biosensors-Micro and Nanoscale Applications; Rinken, T., Ed.; IntechOpen Limited: London, UK, 2015; Volume 6, pp. 165–206. [Google Scholar] [CrossRef] [Green Version]
- Seo, M.-H.; Han, J.; Jin, Z.; Lee, D.-W.; Park, H.-S.; Kim, H.-S. Controlled and Oriented Immobilization of Protein by Site-Specific Incorporation of Unnatural Amino Acid. Anal. Chem. 2011, 83, 2841–2845. [Google Scholar] [CrossRef] [PubMed]
- Shen, G.; Cai, C.; Wang, K.; Lu, J. Improvement of Antibody Immobilization Using Hyperbranched Polymer and Protein A. Anal. Biochem. 2011, 409, 22–27. [Google Scholar] [CrossRef]
- Song, H.Y.; Zhou, X.; Hobley, J.; Su, X. Comparative Study of Random and Oriented Antibody Immobilization as Measured by Dual Polarization Interferometry and Surface Plasmon Resonance Spectroscopy. Langmuir ACS J. Surf. Colloids 2012, 28, 997–1004. [Google Scholar] [CrossRef] [PubMed]
- Tajima, N.; Takai, M.; Ishihara, K. Significance of Antibody Orientation Unraveled: Well-Oriented Antibodies Recorded High Binding Affinity. Anal. Chem. 2011, 83, 1969–1976. [Google Scholar] [CrossRef]
- Vashist, S.K.; Dixit, C.K.; MacCraith, B.D.; O’Kennedy, R. Effect of Antibody Immobilization Strategies on the Analytical Performance of a Surface Plasmon Resonance-Based Immunoassay. Analyst 2011, 136, 4431–4436. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Shen, Z.; Mernaugh, R. Recombinant Antibodies and Their Use in Biosensors. Anal. Bioanal. Chem. 2012, 402, 3027–3038. [Google Scholar] [CrossRef] [Green Version]
- Cho, I.-H.; Seo, S.-M.; Jeon, J.-W.; Paek, S.-H. Characterization for Binding Complex Formation with Site-Directly Immobilized Antibodies Enhancing Detection Capability of Cardiac Troponin I. J. Biomed. Biotechnol. 2009, 2009, 104094. [Google Scholar] [CrossRef] [PubMed]
- Butler, J.E.; Ni, L.; Nessler, R.; Joshi, K.S.; Suter, M.; Rosenberg, B.; Chang, J.; Brown, W.R.; Cantarero, L.A. The Physical and Functional Behavior of Capture Antibodies Adsorbed on Polystyrene. J. Immunol. Methods 1992, 150, 77–90. [Google Scholar] [CrossRef]
- Trilling, A.K.; Harmsen, M.M.; Ruigrok, V.J.B.; Zuilhof, H.; Beekwilder, J. The Effect of Uniform Capture Molecule Orientation on Biosensor Sensitivity: Dependence on Analyte Properties. Biosens. Bioelectron. 2013, 40, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Peluso, P.; Wilson, D.S.; Do, D.; Tran, H.; Venkatasubbaiah, M.; Quincy, D.; Heidecker, B.; Poindexter, K.; Tolani, N.; Phelan, M.; et al. Optimizing Antibody Immobilization Strategies for the Construction of Protein Microarrays. Anal. Biochem. 2003, 312, 113–124. [Google Scholar] [CrossRef]
- Antibody Interactions with Antigens. In Practical Immunology; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2002; pp. 71–114. ISBN 978-0-470-75747-5.
- Susini, V.; Caponi, L.; Rossi, V.L.; Sanesi, A.; Romiti, N.; Paolicchi, A.; Franzini, M. Sensitivity and Reproducibility Enhancement in Enzyme Immunosorbent Assays Based on Half Fragment Antibodies. Anal. Biochem. 2020, 114090. [Google Scholar] [CrossRef]
- Byers, D.M.; Kay, C.M. Hydrodynamic Properties of Bovine Cardiac Troponin-I and Troponin-T. J. Biol. Chem. 1983, 258, 2951–2954. [Google Scholar] [CrossRef]
- Prestigiacomo, A.F.; Chen, Z.; Stamey, T.A. A Universal Calibrator for Prostate Specific Antigen (PSA). Scand. J. Clin. Lab. Investig. Suppl. 1995, 221, 57–59. [Google Scholar] [CrossRef] [PubMed]
- Villoutreix, B.O.; Lilja, H.; Pettersson, K.; Lövgren, T.; Teleman, O. Structural Investigation of the Alpha-1-Antichymotrypsin: Prostate-Specific Antigen Complex by Comparative Model Building. Protein Sci. Publ. Protein Soc. 1996, 5, 836–851. [Google Scholar] [CrossRef] [Green Version]
- Aliau, S.; Marti, J.; Moretti, J. Bovine Alpha-Fetoprotein: Isolation and Characterization. Biochimie 1978, 60, 663–672. [Google Scholar] [CrossRef]
- Ruoslahti, E.; Seppälä, M. Studies of Carcino-Fetal Proteins: Physical and Chemical Properties of Human α-Fetoprotein. Int. J. Cancer 1971, 7, 218–225. [Google Scholar] [CrossRef]
- Gupta, R.; Alamrani, N.A.; Greenway, G.M.; Pamme, N.; Goddard, N.J. Method for Determining Average Iron Content of Ferritin by Measuring Its Optical Dispersion. Anal. Chem. 2019, 91, 7366–7372. [Google Scholar] [CrossRef] [Green Version]
- Esser, P. Principles in Adsorption to Polystyrene; Technical Bulletin; Thermo Fisher Scientific: Waltham, MA, USA, 2010; p. 7. [Google Scholar]
- Mackey, D.; Kelly, E.; Nooney, R.; O’Kennedy, R. Direct immunoassays and their performance—Theoretical modelling of the effects of antibody orientation and associated kinetics. Integr. Biol. 2018, 10, 598–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bush, D.B.; Knotts, T.A. The effects of antigen size, binding site valency, and flexibility on fab-antigen binding near solid surfaces. J. Chem. Phys. 2018, 149, 165102. [Google Scholar] [CrossRef]
- Takeda, N.; Yamashita, A.; Maeda, K.; Maeda, Y. Structure of the core domain of human cardiac troponin in the Ca2+-saturated form. Nature 2003, 424, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Lin, B.; Zhou, P.; Li, M. Molecular Analysis of AFP and HSA Interactions with PTEN Protein. Biomed. Res. Int. 2015, 2015, 256916. [Google Scholar] [CrossRef] [PubMed]
- Lawson, D.M.; Artymiuk, P.J.; Yewdall, S.J.; Smith, J.M.; Livingstone, J.C.; Treffry, A.; Luzzago, A.; Levi, S.; Arosio, P.; Cesareni, G.; et al. Solving the structure of human H ferritin by genetically engineering intermolecular crystal contacts. Nature 1991, 349, 541–544. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antigens | MW (kDa) | Stokes Radius (nm) | Epitope | Epitope Number |
|---|---|---|---|---|
| cTnI | 24 | 3 | N-terminus | 1 |
| PSA-ACT | 90 | >3 | n.a. | 1 |
| AFP | 70 | 3.26 | n.a. | 1–4 |
| Ferritin | 474 | 6 | n.a. | 12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Susini, V.; Fierabracci, V.; Barria, G.; Dodoli, L.; Caponi, L.; Paolicchi, A.; Franzini, M. Signal Enhancement in Oriented Immunosorbent Assays: A Balance between Accessibility of Antigen Binding Sites and Avidity. Biosensors 2021, 11, 493. https://doi.org/10.3390/bios11120493
Susini V, Fierabracci V, Barria G, Dodoli L, Caponi L, Paolicchi A, Franzini M. Signal Enhancement in Oriented Immunosorbent Assays: A Balance between Accessibility of Antigen Binding Sites and Avidity. Biosensors. 2021; 11(12):493. https://doi.org/10.3390/bios11120493
Chicago/Turabian StyleSusini, Vanessa, Vanna Fierabracci, Gaia Barria, Lisa Dodoli, Laura Caponi, Aldo Paolicchi, and Maria Franzini. 2021. "Signal Enhancement in Oriented Immunosorbent Assays: A Balance between Accessibility of Antigen Binding Sites and Avidity" Biosensors 11, no. 12: 493. https://doi.org/10.3390/bios11120493
APA StyleSusini, V., Fierabracci, V., Barria, G., Dodoli, L., Caponi, L., Paolicchi, A., & Franzini, M. (2021). Signal Enhancement in Oriented Immunosorbent Assays: A Balance between Accessibility of Antigen Binding Sites and Avidity. Biosensors, 11(12), 493. https://doi.org/10.3390/bios11120493