Biosensing Cytokine IL-6: A Comparative Analysis of Natural and Synthetic Receptors




Abstract
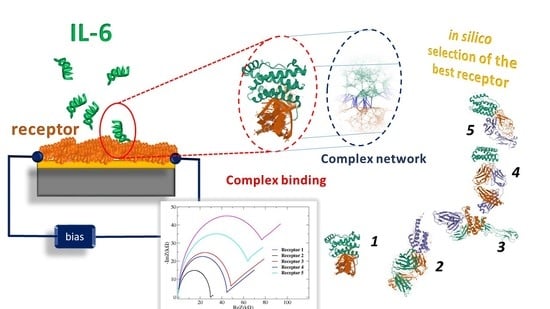
1. Introduction
2. Materials and Methods
2.1. Materials
- IL-6
- IL-6R
- SOMAmer SL1025
- Olokizumab (OKZ)-Fab Portion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| #Entry | Ligand | Receptor | KD(pM) |
|---|---|---|---|
| 1p9m | IL-6 | gp130, IL-6R | 80(IL-6R∙gp130) -900(IL-6R) [41] * |
| 1alu | IL-6 | n.a. | -- |
| 1n26 | n.a. | IL-6R | -- |
| 4zs7 | IL-6 | Fab 68F2 | 13–21 [44] |
| 4cni | IL-6 | Fab OKZ | 10 [18] |
| 4o9h | IL-6 | Fab 61H7 | 3.3–6.3 [44] |
| 4ni9 | IL-6 | SOMAmer SL1025 | 200 [51] |
- 61H7 & 68F2- humanized Camelid-Fab
2.2. Methods
3. Results
3.1. Contact Maps
3.2. ToCI
3.3. Gedanken-Experiment
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Appendix A


References
- Mehta, P.; McAuley, D.F.; Brown, M.; Sanchez, E.; Tattersall, R.S.; Manson, J.J. COVID-19: Consider cytokine storm syndromes and immunosuppression. Lancet 2020, 395, P1033–P1034. [Google Scholar] [CrossRef]
- Moore, J.B.; June, C.H. Cytokine release syndrome in severe COVID-19. Science 2020, 368, 473–474. [Google Scholar] [CrossRef] [PubMed]
- Hunter, C.A.; Jones, S.A. IL-6 as a keystone cytokine in health and disease. Nat. Immunol. 2015, 16, 448–457. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.A.; Scheller, J.; Rose-John, S. Therapeutic strategies for the clinical blockade of IL-6/gp130 signaling. J. Clin. Investig. 2011, 121, 3375–3383. [Google Scholar] [CrossRef]
- Kontzias, A.; Kotlyar, A.; Laurence, A.; Changelian, P.; O’Shea, J.J. Jakinibs: A new class of kinase inhibitors in cancer and autoimmune disease. Curr. Opin. Pharmacol. 2012, 12, 464–470. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, P.C.; Castell, J.V.; Andus, T. Interleukin-6 and the acute phase response. Biochem. J. 1990, 265, 621–636. [Google Scholar] [CrossRef]
- Sato, K.; Tsuchiya, M.; Saldanha, J.; Koishihara, Y.; Ohsugi, Y.; Kishimoto, T.; Bendig, M.M. Reshaping a human antibody to inhibit the interleukin 6-dependent tumor cell growth. Cancer Res. 1993, 53, 851–856. [Google Scholar]
- Sciascia, S.; Aprà, F.; Baffa, A.; Baldovino, S.; Boero, D.; Bonora, S.; Calcagno, A.; Cecchi, I.; Cinnirella, G.; Converso, M.; et al. Pilot prospective open, single-arm multicentre study on off-label use of tocilizumab in patients with severe COVID-19. Clin. Exp. Rheumatol. 2020, 38, 529–532. [Google Scholar]
- Toniati, P.; Piva, S.; Cattalini, M.; Garrafa, E.; Regola, F.; Castelli, F.; Franceschini, F.; Airò, P.; Bazzani, C.; Beindorf, E.A.; et al. Tocilizumab for the treatment of severe COVID-19 pneumonia with hyperinflammatory syndrome and acute respiratory failure: A single center study of 100 patients in Brescia, Italy. Autoimmun. Rev. 2020, 19, 102568. [Google Scholar] [CrossRef]
- Cavalli, G.; De Luca, G.; Campochiaro, C.; Della Torre, E.; Ripa, M.; Canetti, D.; Oltolini, C.; Castiglioni, B.; Tassan Din, C.; Boffini, N.; et al. Interleukin-1 blockade with high-dose anakinra in patients with COVID-19, acute respiratory distress syndrome, and hyperinflammation: A retrospective cohort study. Lancet Rheumatol. 2020, 2, e325–e331. [Google Scholar] [CrossRef]
- Hosseini, F.; Hassannia, H.; Mahdian-Shakib, A.; Jadidi-Niaragh, F.; Enderami, S.E.; Fattahi, M.; Anissian, A.; Mirshafiey, A.; Kokhaei, P. Targeting of crosstalk between tumor and tumor microenvironment by β-D mannuronic acid (M2000) in murine breast cancer model. Cancer Med. 2017, 6, 640–650. [Google Scholar] [CrossRef]
- Taniguchi, K.; Karin, M. NF-κB, inflammation, immunity and cancer: Coming of age. Nat. Rev. Immunol. 2018, 18, 309–324. [Google Scholar] [CrossRef] [PubMed]
- Bromberg, J.; Wang, T.C. Inflammation and cancer: IL-6 and STAT3 complete the link. Cancer Cell 2009, 15, 79–80. [Google Scholar] [CrossRef] [PubMed]
- Grivennikov, S.; Karin, M. Autocrine IL-6 signaling: A key event in tumorigenesis? Cancer Cell 2008, 13, 7–9. [Google Scholar] [CrossRef] [PubMed]
- Ghandadi, M.; Sahebkar, A. Interleukin-6: A critical cytokine in Cancer multidrug resistance. Curr. Pharm. Des. 2016, 22, 518–526. [Google Scholar] [CrossRef] [PubMed]
- Kishimoto, T. Interleukin-6: From basic science to medicine—40 years in immunology. Annu. Rev. Immunol. 2005, 23, 1–21. [Google Scholar] [CrossRef]
- Kumari, N.; Dwarakanath, B.S.; Das, A.; Bhatt, A.N. Role of interleukin-6 in cancer progression and therapeutic resistance. Tumor Biol. 2016, 37, 11553–11572. [Google Scholar] [CrossRef]
- Shaw, S.; Bourne, T.; Meier, C.; Carrington, B.; Gelinas, R.; Henry, A.; Popplewell, A.; Adams, R.; Baker, T.; Rapecki, S.; et al. Discovery and characterization of olokizumab: A humanized antibody targeting interleukin-6 and neutralizing gp130-signaling. mAbs 2014, 6, 773–781. [Google Scholar] [CrossRef]
- Baraket, A.; Lee, M.; Zine, N.; Sigaud, M.; Bausells, J.; Errachid, A. A fully integrated electrochemical biosensor platform fabrication process for cytokines detection. Biosens. Bioelect. 2017, 93, 170–175. [Google Scholar] [CrossRef]
- Tertiş, M.; Ciui, B.; Suciu, M.; Săndulescu, R.; Cristea, C. Label-free electrochemical aptasensor based on gold and polypyrrole nanoparticles for interleukin 6 detection. Electrochim. Acta 2017, 258, 1208–1218. [Google Scholar] [CrossRef]
- Tertis, M.; Leva, P.I.; Bogdan, D.; Suciu, M.; Graur, F.; Cristea, C. Impedimetric aptasensor for the label-free and selective detection of Interleukin-6 for colorectal cancer screening. Biosens. Bioelectr. 2019, 137, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Wang, S.; Jin, H.; Bao, W.; Huang, S.; Wang, J. An electrochemical impedance sensor for the label-free ultrasensitive detection of interleukin-6 antigen. Sens. Actuators B Chem. 2013, 178, 310–315. [Google Scholar] [CrossRef]
- Li, T.; Yang, M. Electrochemical sensor utilizing ferrocene loaded porous polyelectrolyte nanoparticles as label for the detection of protein biomarker IL-6. Sens. Actuators B Chem. 2011, 158, 361–365. [Google Scholar] [CrossRef]
- Huang, J.; Harvey, J.; Fam, W.D.; Nimmo, M.A.; Tok, I.A. Novel biosensor for Interleukin-6 detection. Procedia Eng. 2013, 60, 195–200. [Google Scholar] [CrossRef]
- Crivianu-Gaita, V.; Thompson, M. Aptamers, antibody scFv, and antibody Fab’fragments: An overview and comparison of three of the most versatile biosensor biorecognition elements. Biosens. Bioelectr. 2016, 85, 32–45. [Google Scholar] [CrossRef]
- Lu, H.; Tonge, P.J. Drug–target residence time: Critical information for lead optimization. Curr. Opin. Chem. Biol. 2010, 14, 467–474. [Google Scholar] [CrossRef]
- Bonanni, B.; Kamruzzahan, A.S.M.; Bizzarri, A.R.; Rankl, C.; Gruber, H.J.; Hinterdorfer, P.; Cannistraro, S. Single molecule recognition between cytochrome C 551 and gold-immobilized azurin by force spectroscopy. Biophys. J. 2005, 89, 2783–2791. [Google Scholar] [CrossRef]
- Gelinas, A.D.; Davies, D.R.; Edwards, T.E.; Rohloff, J.C.; Carter, J.D.; Zhang, C.; Gupta, S.; Ishikawa, Y.; Hirota, M.; Jarvis, T.C.; et al. Crystal structure of interleukin-6 in complex with a modified nucleic acid ligand. J. Biol. Chem. 2014, 289, 8720–8734. [Google Scholar] [CrossRef]
- Tuerk, C.; Gold, L. Systematic evolution of ligands by exponential enrichment: RNA ligands to bacteriophage T4 DNA polymerase. Science 1990, 249, 505–510. [Google Scholar] [CrossRef]
- Jo, H.; Ban, C. Aptamer–nanoparticle complexes as powerful diagnostic and therapeutic tools. Exp. Mol. Med. 2016, 48, e230. [Google Scholar] [CrossRef]
- Tombelli, S.; Minunni, M.; Mascini, M. Aptamers-based assays for diagnostics, environmental and food analysis. Biomol. Eng. 2007, 24, 191–200. [Google Scholar] [CrossRef]
- Cataldo, R.; De Nunzio, G.; Millithaler, J.F.; Alfinito, E. Aptamers Which Target Proteins: What Proteotronics Suggests to Pharmaceutics. Curr. Pharm. Des. 2020, 26, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Russo Krauss, I.; Merlino, A.; Randazzo, A.; Novellino, E.; Mazzarella, L.; Sica, F. High-resolution structures of two complexes between thrombin and thrombin-binding aptamer shed light on the role of cations in the aptamer inhibitory activity. Nucleic Acids Res. 2012, 40, 8119–8128. [Google Scholar] [CrossRef] [PubMed]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The protein data bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Bini, A.; Mascini, M.; Mascini, M.; Turner, A.P. Selection of thrombin-binding aptamers by using computational approach for aptasensor application. Biosens. Bioelectron. 2011, 26, 4411–4416. [Google Scholar] [CrossRef] [PubMed]
- Cataldo, R.; Ciriaco, F.; Alfinito, E. A validation strategy for in silico generated aptamers. Comput. Biol. Chem. 2018, 77, 123–130. [Google Scholar] [CrossRef]
- Cataldo, R.; Giotta, L.; Guascito, M.R.; Alfinito, E. Assessing the quality of in silico produced biomolecules: The discovery of a new conformer. J. Phys. Chem. B 2019, 123, 1265–1273. [Google Scholar] [CrossRef]
- Alfinito, E.; Pousset, J.; Reggiani, L. Proteotronics: Development of Protein-Based Electronics; CRC Press: Boca Raton, FL, USA, 2015; pp. 1–270. [Google Scholar]
- Gold, L.; Ayers, D.; Bertino, J.; Bock, C.; Bock, A.; Brody, E.N.; Carter, J.; Dalby, A.; Eaton, B.E.; Fitzwater, T.; et al. Aptamer-based multiplexed proteomic technology for biomarker discovery. PLoS ONE 2010, 1, e0015004. [Google Scholar] [CrossRef]
- Albert, R.; Barabási, A.L. Statistical mechanics of complex networks. Rev. Mod. Phys. 2002, 74, 47. [Google Scholar] [CrossRef]
- Boulanger, M.J.; Chow, D.C.; Brevnova, E.E.; Garcia, K.C. Hexameric structure and assembly of the interleukin-6/IL-6 α-receptor/gp130 complex. Science 2003, 300, 2101–2104. [Google Scholar] [CrossRef]
- White, U.; M Stephens, J. The gp130 receptor cytokine family: Regulators of adipocyte development and function. Curr. Pharm. Des. 2011, 17, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Klarenbeek, A.; Blanchetot, C.; Schragel, G.; Sadi, A.S.; Ongenae, N.; Hemrika, W.; Wijidenes, H.; Spinelli, S.; Desmyter, A.; Cambillau, C.; et al. Combining somatic mutations present in different in vivo affinity-matured antibodies isolated from immunized Lama glama yields ultra-potent antibody therapeutics. PEDS 2016, 29, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Blanchetot, C.; De Jonge, N.; Desmyter, A.; Ongenae, N.; Hofman, E.; Klarenbeek, A.; Sadi, A.; Hultberg, A.; Spinelli, S.; Loris, R.; et al. Structural mimicry of receptor interaction by antagonistic interleukin-6 (IL-6) antibodies. J. Biol. Chem. 2016, 291, 13846–13854. [Google Scholar] [CrossRef] [PubMed]
- Somers, W.; Stahl, M.; Seehra, J.S. 1.9 Å crystal structure of interleukin 6: Implications for a novel mode of receptor dimerization and signaling. EMBO J. 1997, 16, 989–997. [Google Scholar] [CrossRef]
- Kobilka, B.K.; Deupi, X. Conformational complexity of G-protein-coupled receptors. Trends Pharmacol. Sci. 2007, 28, 397–406. [Google Scholar] [CrossRef]
- Varghese, J.N.; Moritz, R.L.; Lou, M.Z.; Van Donkelaar, A.; Ji, H.; Ivancic, N.; Branson, K.M.; Hall, N.E.; Simpson, R.J. Structure of the extracellular domains of the human interleukin-6 receptor α-chain. Proc. Natl. Acad. Sci. USA 2002, 99, 15959–15964. [Google Scholar] [CrossRef]
- Kovacevic, K.D.; Gilbert, J.C.; Jilma, B. Pharmacokinetics, pharmacodynamics and safety of aptamers. Adv. Drug Deliv. Rev. 2018, 134, 36–50. [Google Scholar] [CrossRef]
- Gelinas, A.D.; Davies, D.R.; Janjic, N. Embracing proteins: Structural themes in aptamer–protein complexes. Curr. Opin. Struct. Biol. 2016, 36, 122–132. [Google Scholar] [CrossRef]
- Gupta, S.; Hirota, M.; Waugh, S.M.; Murakami, I.; Suzuki, T.; Muraguchi, M.; Shibamori, M.; Ishikawa, Y.; Jarvis, C.T.; Carter, J.D.; et al. Chemically modified DNA aptamers bind interleukin-6 with high affinity and inhibit signaling by blocking its interaction with interleukin-6 receptor. J. Biol. Chem. 2014, 289, 8706–8719. [Google Scholar] [CrossRef]
- Hirota, M.; Murakami, I.; Ishikawa, Y.; Suzuki, T.; Sumida, S.I.; Ibaragi, S.; Janjic, N.; Schneider, D.J. Chemically modified interleukin-6 aptamer inhibits development of collagen-induced arthritis in cynomolgus monkeys. Nucleic Acid Ther. 2016, 26, 10–19. [Google Scholar] [CrossRef]
- Nasonov, E.; Fatenejad, S.; Korneva, E.; Krechikova, D.; Maslyansky, A.; Plaksina, T.; Stanislav, M.; Stoilov, R.; Tyabut, T.; Yakushin, S.; et al. Safety and Efficacy of Olokizumab in a Phase III Trial of Patients with Moderately to Severely Active Rheumatoid Arthritis Inadequately Controlled by Methotrexate—CREDO1 Study. In Arthritis & Rheumatology; Wiley: Hoboken, NJ, USA, 2019; Volume 71, Available online: https://acrabstracts.org/abstract/safety-and-efficacy-of-olokizumab-in-a-phase-iii-trial-of-patients-with-moderately-to-severely-active-rheumatoid-arthritis-inadequately-controlled-by-methotrexate-credo1-study/ (accessed on 17 June 2020).
- Alfinito, E.; Reggiani, L.; Cataldo, R.; De Nunzio, G.; Giotta, L.; Guascito, M.R. Modeling the microscopic electrical properties of thrombin binding aptamer (TBA) for label-free biosensors. Nanotechnology 2017, 28, 065502. [Google Scholar] [CrossRef] [PubMed]
- Bizzarri, A.R.; Cannistraro, S. The application of atomic force spectroscopy to the study of biological complexes undergoing a biorecognition process. Chem. Soc. Rev. 2010, 39, 734–749. [Google Scholar] [CrossRef] [PubMed]
- Alfinito, E.; Reggiani, L. Mechanisms responsible for the photocurrent in bacteriorhodopsin. Phys. Rev. E 2015, 91, 032702. [Google Scholar] [CrossRef] [PubMed]
- Alfinito, E.; Millithaler, J.F.; Reggiani, L.; Zine, N.; Jaffrezic-Renault, N. Human olfactory receptor 17-40 as an active part of a nanobiosensor: A microscopic investigation of its electrical properties. RSC Adv. 2011, 1, 123–127. [Google Scholar] [CrossRef]
- Bochet, L.; Meulle, A.; Imbert, S.; Salles, B.; Valet, P.; Muller, C. Cancer-associated adipocytes promotes breast tumor radioresistance. Biochem. Biophys. Res. Commun. 2011, 411, 102–106. [Google Scholar] [CrossRef]
- Hirano, T.; Ishihara, K.; Hibi, M. Roles of STAT3 in mediating the cell growth, differentiation and survival signals relayed through the IL-6 family of cytokine receptors. Oncogene 2000, 19, 2548–2556. [Google Scholar] [CrossRef]
- Guo, Y.; Xu, F.; Lu, T.; Duan, Z.; Zhang, Z. Interleukin-6 signaling pathway in targeted therapy for cancer. Cancer Treat. Rev. 2012, 38, 904–910. [Google Scholar] [CrossRef]
- Wolf, J.; Rose-John, S.; Garbers, C. Interleukin-6 and its receptors: A highly regulated and dynamic system. Cytokine 2014, 70, 11–20. [Google Scholar] [CrossRef]







| Receptor | s(A-D) | s(A-D*) | s(A-C) | s(B) |
|---|---|---|---|---|
| IL-6R & gp130 | 2.7 | 3.4 | 3.3 | 3.0 |
| gp130 | 4.1 | 4.4 | 4.9 | 5.1 |
| IL-6R | 3.1 | 3.0 | 5.2 | 3.5 |
| OKZ-Fab | 7.4 | 1.9 | 5.3 | 5.7 |
| SL1025-Apt | 1.6 | 3.0 | 2.2 | 2.4 |
| 61H7-Fab | 1.2 | 6.7 | 4.4 | 4.2 |
| 68F2-Fab | 3.4 | 4.2 | 4.4 | 4.1 |
| Receptor | rcomp(D1) | rrec(DLOD) | κ | Δ/N |
|---|---|---|---|---|
| IL-6R | 16.4 | 1 | 16.4 | 15.4 |
| OKZ-Fab | 88.0 | 0.32 | 275 | 86.1 |
| SL1025-Apt | 227 | 15.9 | 14.3 | 211 |
| 61H7-Fab 68F2-Fab | 200 35.9 | 0.38 0.33 | 530 110 | 200 35.9 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alfinito, E.; Beccaria, M.; Ciccarese, M. Biosensing Cytokine IL-6: A Comparative Analysis of Natural and Synthetic Receptors. Biosensors 2020, 10, 106. https://doi.org/10.3390/bios10090106
Alfinito E, Beccaria M, Ciccarese M. Biosensing Cytokine IL-6: A Comparative Analysis of Natural and Synthetic Receptors. Biosensors. 2020; 10(9):106. https://doi.org/10.3390/bios10090106
Chicago/Turabian StyleAlfinito, Eleonora, Matteo Beccaria, and Mariangela Ciccarese. 2020. "Biosensing Cytokine IL-6: A Comparative Analysis of Natural and Synthetic Receptors" Biosensors 10, no. 9: 106. https://doi.org/10.3390/bios10090106
APA StyleAlfinito, E., Beccaria, M., & Ciccarese, M. (2020). Biosensing Cytokine IL-6: A Comparative Analysis of Natural and Synthetic Receptors. Biosensors, 10(9), 106. https://doi.org/10.3390/bios10090106