
The Apoptosis Inhibitor Protein Survivin Is a Critical Cytoprotective Resistor against Silica-Based Nanotoxicity
 ,
,  , , and
, , and 
Abstract
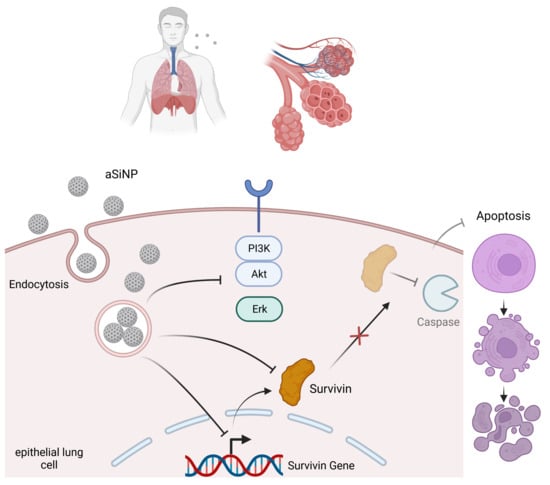
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Nanoparticles, Antibodies (Ab), and Reagents
2.2. Characterization of Amorphous Silica Nanoparticles (aSiNPs)
2.3. Transmission Electron Microscopy (TEM)
2.4. Physicochemical Characterization Methods
2.5. Endotoxin Detection in aSiNPs Dispersions
2.6. Preparation of aSiNPs for Apical Exposure
2.7. Nanoparticle Uptake Studies
2.8. Cultivation of Model Cell Lines ISO-HAS-1 and H441
2.9. Plasmids
2.10. Cells Transfection and Luciferase Assay
2.11. Apoptosis and Viability Assays
2.12. Origin of Human and Mouse Tissue
2.13. Tissue Preparation and Immunohistochemistry (IHC)
2.14. Microscopy and Fluorescence Imaging of Cells
2.15. Protein Extraction, Immunoblot Analysis, and Immunofluorescence
2.16. Statistical Analysis
3. Results
3.1. ‘Characterization of the Nano–Bio Interface I’—Nano-scaled Amorphous Silica Nanoparticles
3.2. ‘Characterization of the Nano–Bio Interface II’—A Biological Model to Mimic the Alveolar-Capillary Barrier of the Lung
3.3. ‘Effects at the Nano–Bio Interface’—aSiNPs Induce Cell Death upon Exposure to Alveolar-Capillary Barrier Model Cells
3.4. ‘Signaling at the Nano–Bio Interface’—High Concentrations of aSiNP15 Attenuate Pro-Survival Pathways
3.5. ‘Survival and Death at the Nano–Bio Interface’—aSiNP-Induced Cell Death Is Regulated by the Anti-Apoptotic Protein Survivin
3.6. ‘Probing Physiological Relevance of the Nano–Bio Interface’—Survivin Is Expressed in Healthy Mouse and Human Lung Tissue
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Davies, J.C.T. PEN 13-Nanotechnology Oversight. Available online: https://www.wilsoncenter.org/publication/pen-13-nanotechnology-oversight (accessed on 14 January 2019).
- Liljenström, C.L.D.; Finnveden, G. Silicon-Based Nanomaterials in a Life-Cycle Perspective, Including a Case Study on Self-Cleaning Coatings. 2013. Available online: https://www.researchgate.net/publication/280264076_Silicon-based_nanomaterials_in_a_life-cycle_perspective_including_a_case_study_on_self-cleaning_coatings (accessed on 14 January 2019).
- Vance, M.; Lawson, N.C.; Rupal, M.; Beck, P.; Burgess, J.O. Color and Gloss of Nano-Filled Resin-Modified Glass Ionomers and Resin Composites. J. Esthet. Restor. Dent. 2015, 27, 293–299. [Google Scholar] [CrossRef]
- Croissant, J.G.; Fatieiev, Y.; Khashab, N.M. Degradability and Clearance of Silicon, Organosilica, Silsesquioxane, Silica Mixed Oxide, and Mesoporous Silica Nanoparticles. Adv. Mater. 2017, 29, 1604634. [Google Scholar] [CrossRef]
- Dekkers, S.; Krystek, P.; Peters, R.J.; Lankveld, D.P.; Bokkers, B.G.; van Hoeven-Arentzen, P.H.; Bouwmeester, H.; Oomen, A.G. Presence and risks of nanosilica in food products. Nanotoxicology 2011, 5, 393–405. [Google Scholar] [CrossRef]
- Leung, C.C.; Yu, I.T.; Chen, W. Silicosis. Lancet 2012, 379, 2008–2018. [Google Scholar] [CrossRef]
- Barnes, C.A.; Elsaesser, A.; Arkusz, J.; Smok, A.; Palus, J.; Lesniak, A.; Salvati, A.; Hanrahan, J.P.; Jong, W.H.; Dziubaltowska, E.; et al. Reproducible comet assay of amorphous silica nanoparticles detects no genotoxicity. Nano Lett. 2008, 8, 3069–3074. [Google Scholar] [CrossRef]
- Arts, J.H.; Muijser, H.; Duistermaat, E.; Junker, K.; Kuper, C.F. Five-day inhalation toxicity study of three types of synthetic amorphous silicas in Wistar rats and post-exposure evaluations for up to 3 months. Food Chem. Toxicol. 2007, 45, 1856–1867. [Google Scholar] [CrossRef]
- Barik, T.K.; Sahu, B.; Swain, V. Nanosilica-from medicine to pest control. Parasitol. Res. 2008, 103, 253–258. [Google Scholar] [CrossRef]
- Fadeel, B.; Garcia-Bennett, A.E. Better safe than sorry: Understanding the toxicological properties of inorganic nanoparticles manufactured for biomedical applications. Adv. Drug Deliv. Rev. 2010, 62, 362–374. [Google Scholar] [CrossRef]
- da Cruz Schneid, A.; Albuquerque, L.J.C.; Mondo, G.B.; Ceolin, M.; Picco, A.S.; Cardoso, M.B. Colloidal stability and degradability of silica nanoparticles in biological fluids: A review. J. Sol.-Gel Sci. Technol. 2022, 102, 41–62. [Google Scholar] [CrossRef]
- Higashisaka, Y.Y.K.; Tsutsumi, Y. Applications and Safety of Nanomaterials Used in the Food Industry. Food Saf. 2015, 3, 39–47. [Google Scholar] [CrossRef]
- Napierska, D.; Thomassen, L.C.; Rabolli, V.; Lison, D.; Gonzalez, L.; Kirsch-Volders, M.; Martens, J.A.; Hoet, P.H. Size-dependent cytotoxicity of monodisperse silica nanoparticles in human endothelial cells. Small 2009, 5, 846–853. [Google Scholar] [CrossRef]
- Thomassen, L.C.; Aerts, A.; Rabolli, V.; Lison, D.; Gonzalez, L.; Kirsch-Volders, M.; Napierska, D.; Hoet, P.H.; Kirschhock, C.E.; Martens, J.A. Synthesis and characterization of stable monodisperse silica nanoparticle sols for in vitro cytotoxicity testing. Langmuir 2010, 26, 328–335. [Google Scholar] [CrossRef]
- Choi, H.S.; Ashitate, Y.; Lee, J.H.; Kim, S.H.; Matsui, A.; Insin, N.; Bawendi, M.G.; Semmler-Behnke, M.; Frangioni, J.V.; Tsuda, A. Rapid translocation of nanoparticles from the lung airspaces to the body. Nat. Biotechnol. 2010, 28, 1300–1303. [Google Scholar] [CrossRef]
- Xie, G.; Sun, J.; Zhong, G.; Shi, L.; Zhang, D. Biodistribution and toxicity of intravenously administered silica nanoparticles in mice. Arch. Toxicol. 2010, 84, 183–190. [Google Scholar] [CrossRef]
- Fruijtier-Polloth, C. The toxicological mode of action and the safety of synthetic amorphous silica-a nanostructured material. Toxicology 2012, 294, 61–79. [Google Scholar] [CrossRef]
- Ahamed, M. Silica nanoparticles-induced cytotoxicity, oxidative stress and apoptosis in cultured A431 and A549 cells. Hum. Exp. Toxicol. 2013, 32, 186–195. [Google Scholar] [CrossRef]
- Lankoff, A.; Arabski, M.; Wegierek-Ciuk, A.; Kruszewski, M.; Lisowska, H.; Banasik-Nowak, A.; Rozga-Wijas, K.; Wojewodzka, M.; Slomkowski, S. Effect of surface modification of silica nanoparticles on toxicity and cellular uptake by human peripheral blood lymphocytes in vitro. Nanotoxicology 2013, 7, 235–250. [Google Scholar] [CrossRef]
- Yang, Y.X.; Song, Z.M.; Cheng, B.; Xiang, K.; Chen, X.X.; Liu, J.H.; Cao, A.; Wang, Y.; Liu, Y.; Wang, H. Evaluation of the toxicity of food additive silica nanoparticles on gastrointestinal cells. J. Appl. Toxicol. 2014, 34, 424–435. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Wheatley, S.P.; Altieri, D.C. Survivin at a glance. J. Cell Sci. 2019, 132, 223826. [Google Scholar] [CrossRef]
- Unruhe, B.; Schroder, E.; Wunsch, D.; Knauer, S.K. An Old Flame Never Dies: Survivin in Cancer and Cellular Senescence. Gerontology 2016, 62, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Rauch, A.; Carlstedt, A.; Emmerich, C.; Mustafa, A.M.; Goder, A.; Knauer, S.K.; Linnebacher, M.; Heinzel, T.; Kramer, O.H. Survivin antagonizes chemotherapy-induced cell death of colorectal cancer cells. Oncotarget 2018, 9, 27835–27850. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Lu, W.; Yang, J.; Edwards, M.; Jiang, S. Survivin as a biological biomarker for diagnosis and therapy. Expert. Opin. Biol. Ther. 2021, 21, 1429–1441. [Google Scholar] [CrossRef] [PubMed]
- Rahimmi, A.; Fathi, F.; Nikkhoo, B.; Soleimani, F.; Khademerfan, M. Over-expression of survivin could prevent the oxidative stress and toxicity of rotenone in SH-SY5Y cells. Iran. J. Basic. Med. Sci. 2022, 25, 842–849. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Chen, J.K.; Conway, E.M.; Harris, R.C. Survivin mediates renal proximal tubule recovery from AKI. J. Am. Soc. Nephrol. 2013, 24, 2023–2033. [Google Scholar] [CrossRef]
- Knauer, S.K.; Heinrich, U.R.; Bier, C.; Habtemichael, N.; Docter, D.; Helling, K.; Mann, W.J.; Stauber, R.H. An otoprotective role for the apoptosis inhibitor protein survivin. Cell Death Dis. 2010, 1, e51. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Assadiasl, S.; Mousavi, M.J.; Amirzargar, A. Antiapoptotic Molecule Survivin in Transplantation: Helpful or Harmful? J. Transpl. 2018, 2018, 6492034. [Google Scholar] [CrossRef]
- Lee, B.S.; Kim, S.H.; Jin, T.; Choi, E.Y.; Oh, J.; Park, S.; Lee, S.H.; Chung, J.H.; Kang, S.M. Protective effect of survivin in Doxorubicin-induced cell death in h9c2 cardiac myocytes. Korean Circ. J. 2013, 43, 400–407. [Google Scholar] [CrossRef]
- Kasper, J.Y.; Feiden, L.; Hermanns, M.I.; Bantz, C.; Maskos, M.; Unger, R.E.; Kirkpatrick, C.J. Pulmonary surfactant augments cytotoxicity of silica nanoparticles: Studies on an in vitro air-blood barrier model. Beilstein J. Nanotechnol. 2015, 6, 517–528. [Google Scholar] [CrossRef]
- Knauer, S.K.; Bier, C.; Habtemichael, N.; Stauber, R.H. The Survivin-Crm1 interaction is essential for chromosomal passenger complex localization and function. EMBO Rep. 2006, 7, 1259–1265. [Google Scholar] [CrossRef]
- Knauer, S.K.; Kramer, O.H.; Knosel, T.; Engels, K.; Rodel, F.; Kovacs, A.F.; Dietmaier, W.; Klein-Hitpass, L.; Habtemichael, N.; Schweitzer, A.; et al. Nuclear export is essential for the tumor-promoting activity of survivin. FASEB J. 2007, 21, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Fetz, V.; Bier, C.; Habtemichael, N.; Schuon, R.; Schweitzer, A.; Kunkel, M.; Engels, K.; Kovacs, A.F.; Schneider, S.; Mann, W.; et al. Inducible NO synthase confers chemoresistance in head and neck cancer by modulating survivin. Int. J. Cancer 2009, 124, 2033–2041. [Google Scholar] [CrossRef] [PubMed]
- Kramer, O.H.; Knauer, S.K.; Zimmermann, D.; Stauber, R.H.; Heinzel, T. Histone deacetylase inhibitors and hydroxyurea modulate the cell cycle and cooperatively induce apoptosis. Oncogene 2008, 27, 732–740. [Google Scholar] [CrossRef] [PubMed]
- Kramer, O.H.; Knauer, S.K.; Greiner, G.; Jandt, E.; Reichardt, S.; Guhrs, K.H.; Stauber, R.H.; Bohmer, F.D.; Heinzel, T. A phosphorylation-acetylation switch regulates STAT1 signaling. Genes. Dev. 2009, 23, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Engels, K.; Knauer, S.K.; Loibl, S.; Fetz, V.; Harter, P.; Schweitzer, A.; Fisseler-Eckhoff, A.; Kommoss, F.; Hanker, L.; Nekljudova, V.; et al. NO signaling confers cytoprotectivity through the survivin network in ovarian carcinomas. Cancer Res. 2008, 68, 5159–5166. [Google Scholar] [CrossRef]
- Matter, F.L.; Laura, A. Niederberger, Markus From colloidal dispersions to aerogels: How to master nanoparticle gelation. Nano Today 2020, 30, 100827. [Google Scholar] [CrossRef]
- Sonwani, S.; Madaan, S.; Arora, J.; Suryanarayan, S.; Rangra, D.; Mongia, N.; Vats, T.; Saxena, P. Inhalation Exposure to Atmospheric Nanoparticles and Its Associated Impacts on Human Health: A Review. Front. Sustain. Cities 2021, 3, 690444. [Google Scholar] [CrossRef]
- Kapalczynska, M.; Kolenda, T.; Przybyla, W.; Zajaczkowska, M.; Teresiak, A.; Filas, V.; Ibbs, M.; Blizniak, R.; Luczewski, L.; Lamperska, K. 2D and 3D cell cultures—A comparison of different types of cancer cell cultures. Arch. Med. Sci. 2018, 14, 910–919. [Google Scholar] [CrossRef]
- Hermanns, M.I.; Kasper, J.; Dubruel, P.; Pohl, C.; Uboldi, C.; Vermeersch, V.; Fuchs, S.; Unger, R.E.; Kirkpatrick, C.J. An impaired alveolar-capillary barrier in vitro: Effect of proinflammatory cytokines and consequences on nanocarrier interaction. J. R. Soc. Interface 2010, 7 (Suppl. S1), S41–S54. [Google Scholar] [CrossRef]
- Pryhuber, G.S.; Khalak, R.; Zhao, Q. Regulation of surfactant proteins A and B by TNF-alpha and phorbol ester independent of NF-kappa B. Am. J. Physiol. 1998, 274, L289–L295. [Google Scholar] [CrossRef]
- Kasper, M.; Barth, K. Potential contribution of alveolar epithelial type I cells to pulmonary fibrosis. Biosci. Rep. 2017, 37, BSR20171301. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.Z.; Fang, X.C.; Wang, D.; Tang, F.D.; Wang, X.D. Involvement of type II pneumocytes in the pathogenesis of chronic obstructive pulmonary disease. Respir. Med. 2010, 104, 1391–1395. [Google Scholar] [CrossRef] [PubMed]
- Salomon, J.J.; Muchitsch, V.E.; Gausterer, J.C.; Schwagerus, E.; Huwer, H.; Daum, N.; Lehr, C.M.; Ehrhardt, C. The cell line NCl-H441 is a useful in vitro model for transport studies of human distal lung epithelial barrier. Mol. Pharm. 2014, 11, 995–1006. [Google Scholar] [CrossRef]
- Khubchandani, K.R.; Snyder, J.M. Surfactant protein A (SP-A): The alveolus and beyond. FASEB J. 2001, 15, 59–69. [Google Scholar] [CrossRef]
- Wert, S.E.; Dey, C.R.; Blair, P.A.; Kimura, S.; Whitsett, J.A. Increased expression of thyroid transcription factor-1 (TTF-1) in respiratory epithelial cells inhibits alveolarization and causes pulmonary inflammation. Dev. Biol. 2002, 242, 75–87. [Google Scholar] [CrossRef]
- Kuo, W.T.; Odenwald, M.A.; Turner, J.R.; Zuo, L. Tight junction proteins occludin and ZO-1 as regulators of epithelial proliferation and survival. Ann. N. Y. Acad. Sci. 2022, 1514, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Docter, D.; Bantz, C.; Westmeier, D.; Galla, H.J.; Wang, Q.; Kirkpatrick, J.C.; Nielsen, P.; Maskos, M.; Stauber, R.H. The protein corona protects against size- and dose-dependent toxicity of amorphous silica nanoparticles. Beilstein J. Nanotechnol. 2014, 5, 1380–1392. [Google Scholar] [CrossRef]
- Tenzer, S.; Docter, D.; Kuharev, J.; Musyanovych, A.; Fetz, V.; Hecht, R.; Schlenk, F.; Fischer, D.; Kiouptsi, K.; Reinhardt, C.; et al. Rapid formation of plasma protein corona critically affects nanoparticle pathophysiology. Nat. Nanotechnol. 2013, 8, 772–781. [Google Scholar] [CrossRef]
- Cao, Z.; Liao, Q.; Su, M.; Huang, K.; Jin, J.; Cao, D. AKT and ERK dual inhibitors: The way forward? Cancer Lett. 2019, 459, 30–40. [Google Scholar] [CrossRef]
- Belyanskaya, L.L.; Hopkins-Donaldson, S.; Kurtz, S.; Simoes-Wust, A.P.; Yousefi, S.; Simon, H.U.; Stahel, R.; Zangemeister-Wittke, U. Cisplatin activates Akt in small cell lung cancer cells and attenuates apoptosis by survivin upregulation. Int. J. Cancer 2005, 117, 755–763. [Google Scholar] [CrossRef]
- Ye, Q.; Cai, W.; Zheng, Y.; Evers, B.M.; She, Q.B. ERK and AKT signaling cooperate to translationally regulate survivin expression for metastatic progression of colorectal cancer. Oncogene 2014, 33, 1828–1839. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Meng, Q.; Liu, L.Z.; You, Y.P.; Liu, N.; Jiang, B.H. Regulation of survivin by PI3K/Akt/p70S6K1 pathway. Biochem. Biophys. Res. Commun. 2010, 395, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Croissant, J.G.; Fatieiev, Y.; Almalik, A.; Khashab, N.M. Mesoporous Silica and Organosilica Nanoparticles: Physical Chemistry, Biosafety, Delivery Strategies, and Biomedical Applications. Adv. Heal. Mater. 2018, 7, 1700831. [Google Scholar] [CrossRef]
- Stahl, P.; Kollenda, S.; Sager, J.; Schmidt, L.; Schroer, M.A.; Stauber, R.H.; Epple, M.; Knauer, S.K. Tuning Nanobodies’ Bioactivity: Coupling to Ultrasmall Gold Nanoparticles Allows the Intracellular Interference with Survivin. Small 2023, 19, e2300871. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Zhen, H.; Zhang, J.; Zhang, W.; Zhang, R.; Cheng, X.; Guo, G.; Mao, X.; Wang, J.; Zhang, X. Survivin promotes glioma angiogenesis through vascular endothelial growth factor and basic fibroblast growth factor in vitro and in vivo. Mol. Carcinog. 2012, 51, 586–595. [Google Scholar] [CrossRef] [PubMed]
- Koike, H.; Morikawa, Y.; Sekine, Y.; Matsui, H.; Shibata, Y.; Suzuki, K. Survivin is associated with cell proliferation and has a role in 1a,25-dihydroxyvitamin D3 induced cell growth inhibition in prostate cancer. J. Urol. 2011, 185, 1497–1503. [Google Scholar] [CrossRef] [PubMed]
- Wolanin, K.; Piwocka, K. Role of survivin in mitosis. Postep. Biochem. 2007, 53, 10–18. [Google Scholar]
- Na, T.Y.; Schecterson, L.; Mendonsa, A.M.; Gumbiner, B.M. The functional activity of E-cadherin controls tumor cell metastasis at multiple steps. Proc. Natl. Acad. Sci. USA 2020, 117, 5931–5937. [Google Scholar] [CrossRef]
- Liu, L.; Shi, G.P. CD31: Beyond a marker for endothelial cells. Cardiovasc. Res. 2012, 94, 3–5. [Google Scholar] [CrossRef]
- Gao, J.; Liu, X.; Yang, F.; Liu, T.; Yan, Q.; Yang, X. By inhibiting Ras/Raf/ERK and MMP-9, knockdown of EpCAM inhibits breast cancer cell growth and metastasis. Oncotarget 2015, 6, 27187–27198. [Google Scholar] [CrossRef]
- Huang, L.; Yang, Y.; Yang, F.; Liu, S.; Zhu, Z.; Lei, Z.; Guo, J. Functions of EpCAM in physiological processes and diseases (Review). Int. J. Mol. Med. 2018, 42, 1771–1785. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.C.; Wu, C.T.; Chen, C.S.; Chang, Y.L. E-cadherin expression in surgically-resected non-small cell lung cancers--a clinicopathological study. Thorac. Cardiovasc. Surg. 2000, 48, 294–299. [Google Scholar] [CrossRef] [PubMed]
- Benlhabib, H.; Mendelson, C.R. Epigenetic regulation of surfactant protein A gene (SP-A) expression in fetal lung reveals a critical role for Suv39h methyltransferases during development and hypoxia. Mol. Cell Biol. 2011, 31, 1949–1958. [Google Scholar] [CrossRef] [PubMed]
- Sano, H.; Kuroki, Y. The lung collectins, SP-A and SP-D, modulate pulmonary innate immunity. Mol. Immunol. 2005, 42, 279–287. [Google Scholar] [CrossRef] [PubMed]
- La Rosa, S.; Chiaravalli, A.M.; Placidi, C.; Papanikolaou, N.; Cerati, M.; Capella, C. TTF1 expression in normal lung neuroendocrine cells and related tumors: Immunohistochemical study comparing two different monoclonal antibodies. Virchows Arch. 2010, 457, 497–507. [Google Scholar] [CrossRef]
- Pusztaszeri, M.P.; Seelentag, W.; Bosman, F.T. Immunohistochemical expression of endothelial markers CD31, CD34, von Willebrand factor, and Fli-1 in normal human tissues. J. Histochem. Cytochem. 2006, 54, 385–395. [Google Scholar] [CrossRef]
- Vallet, C.; Aschmann, D.; Beuck, C.; Killa, M.; Meiners, A.; Mertel, M.; Ehlers, M.; Bayer, P.; Schmuck, C.; Giese, M.; et al. Functional Disruption of the Cancer-Relevant Interaction between Survivin and Histone H3 with a Guanidiniocarbonyl Pyrrole Ligand. Angew. Chem. Int. Ed. Engl. 2020, 59, 5567–5571. [Google Scholar] [CrossRef]
- Fukuda, S.; Pelus, L.M. Survivin, a cancer target with an emerging role in normal adult tissues. Mol. Cancer Ther. 2006, 5, 1087–1098. [Google Scholar] [CrossRef]
- Gil-Kulik, P.; Krzyzanowski, A.; Dudzinska, E.; Karwat, J.; Chomik, P.; Swistowska, M.; Kondracka, A.; Kwasniewska, A.; Cioch, M.; Jojczuk, M.; et al. Potential Involvement of BIRC5 in Maintaining Pluripotency and Cell Differentiation of Human Stem Cells. Oxid. Med. Cell Longev. 2019, 2019, 8727925. [Google Scholar] [CrossRef]
- Asweto, C.O.; Wu, J.; Alzain, M.A.; Hu, H.; Andrea, S.; Feng, L.; Yang, X.; Duan, J.; Sun, Z. Cellular pathways involved in silica nanoparticles induced apoptosis: A systematic review of in vitro studies. Env. Toxicol. Pharmacol. 2017, 56, 191–197. [Google Scholar] [CrossRef]
- Kusaczuk, M.; Kretowski, R.; Naumowicz, M.; Stypulkowska, A.; Cechowska-Pasko, M. Silica nanoparticle-induced oxidative stress and mitochondrial damage is followed by activation of intrinsic apoptosis pathway in glioblastoma cells. Int. J. Nanomed. 2018, 13, 2279–2294. [Google Scholar] [CrossRef] [PubMed]
- Al Samri, M.T.; Biradar, A.V.; Alsuwaidi, A.R.; Balhaj, G.; Al-Hammadi, S.; Shehab, S.; Al-Salam, S.; Tariq, S.; Pramathan, T.; Benedict, S.; et al. In vitro biocompatibility of calcined mesoporous silica particles and fetal blood cells. Int. J. Nanomed. 2012, 7, 3111–3121. [Google Scholar] [CrossRef]
- Lehman, S.E.; Morris, A.S.; Mueller, P.S.; Salem, A.K.; Grassian, V.H.; Larsen, S.C. Silica Nanoparticle-Generated ROS as a Predictor of Cellular Toxicity: Mechanistic Insights and Safety by Design. Env. Sci. Nano 2016, 3, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Siegrist, S.; Kettiger, H.; Fasler-Kan, E.; Huwyler, J. Selective stimulation of the JAK/STAT signaling pathway by silica nanoparticles in human endothelial cells. Toxicol. Vitr. 2017, 42, 308–318. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Dunphy, D.R.; Jiang, X.; Meng, H.; Sun, B.; Tarn, D.; Xue, M.; Wang, X.; Lin, S.; Ji, Z.; et al. Processing pathway dependence of amorphous silica nanoparticle toxicity: Colloidal vs pyrolytic. J. Am. Chem. Soc. 2012, 134, 15790–15804. [Google Scholar] [CrossRef]
- Altomare, D.A.; Khaled, A.R. Homeostasis and the importance for a balance between AKT/mTOR activity and intracellular signaling. Curr. Med. Chem. 2012, 19, 3748–3762. [Google Scholar] [CrossRef]
- Nitschkowski, D.; Marwitz, S.; Kotanidou, S.A.; Reck, M.; Kugler, C.; Rabe, K.F.; Ammerpohl, O.; Goldmann, T. Live and let die: Epigenetic modifications of Survivin and Regucalcin in non-small cell lung cancer tissues contribute to malignancy. Clin. Epigenetics 2019, 11, 157. [Google Scholar] [CrossRef]
- Munscher, A.; Prochnow, S.; Gulati, A.; Sauter, G.; Lorincz, B.; Blessmann, M.; Hanken, H.; Bottcher, A.; Clauditz, T.S. Survivin expression in head and neck squamous cell carcinomas is frequent and correlates with clinical parameters and treatment outcomes. Clin. Oral. Investig. 2019, 23, 361–367. [Google Scholar] [CrossRef]
- Zhou, J.; Guo, X.; Chen, W.; Wang, L.; Jin, Y. Targeting survivin sensitizes cervical cancer cells to radiation treatment. Bioengineered 2020, 11, 130–140. [Google Scholar] [CrossRef]
- Knauer, S.K.; Unruhe, B.; Karczewski, S.; Hecht, R.; Fetz, V.; Bier, C.; Friedl, S.; Wollenberg, B.; Pries, R.; Habtemichael, N.; et al. Functional characterization of novel mutations affecting survivin (BIRC5)-mediated therapy resistance in head and neck cancer patients. Hum. Mutat. 2013, 34, 395–404. [Google Scholar] [CrossRef]
- Arkun, Y. Dynamic Modeling and Analysis of the Cross-Talk between Insulin/AKT and MAPK/ERK Signaling Pathways. PLoS ONE 2016, 11, e0149684. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Breder-Bonk, C.; Docter, D.; Barz, M.; Strieth, S.; Knauer, S.K.; Gül, D.; Stauber, R.H. The Apoptosis Inhibitor Protein Survivin Is a Critical Cytoprotective Resistor against Silica-Based Nanotoxicity. Nanomaterials 2023, 13, 2546. https://doi.org/10.3390/nano13182546
Breder-Bonk C, Docter D, Barz M, Strieth S, Knauer SK, Gül D, Stauber RH. The Apoptosis Inhibitor Protein Survivin Is a Critical Cytoprotective Resistor against Silica-Based Nanotoxicity. Nanomaterials. 2023; 13(18):2546. https://doi.org/10.3390/nano13182546
Chicago/Turabian StyleBreder-Bonk, Christina, Dominic Docter, Matthias Barz, Sebastian Strieth, Shirley K. Knauer, Désirée Gül, and Roland H. Stauber. 2023. "The Apoptosis Inhibitor Protein Survivin Is a Critical Cytoprotective Resistor against Silica-Based Nanotoxicity" Nanomaterials 13, no. 18: 2546. https://doi.org/10.3390/nano13182546
APA StyleBreder-Bonk, C., Docter, D., Barz, M., Strieth, S., Knauer, S. K., Gül, D., & Stauber, R. H. (2023). The Apoptosis Inhibitor Protein Survivin Is a Critical Cytoprotective Resistor against Silica-Based Nanotoxicity. Nanomaterials, 13(18), 2546. https://doi.org/10.3390/nano13182546