
Effect of Alumina Particles on the Osteogenic Ability of Osteoblasts
 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Al2O3, Ti, and UHMWPE Particles
2.2. Cell Culture
2.3. MTT Assay
2.4. Lactate Dehydrogenase Activity (LDH) Assay
2.5. Alkaline Phosphatase (ALP) Activity
2.6. Protein Isolation and Western Blotting
2.7. RNA Isolation and Real-Time RT-PCR
2.8. Luciferase Reporter Assay
2.9. Sirius Red S Staining
2.10. Statistical Analysis
3. Results
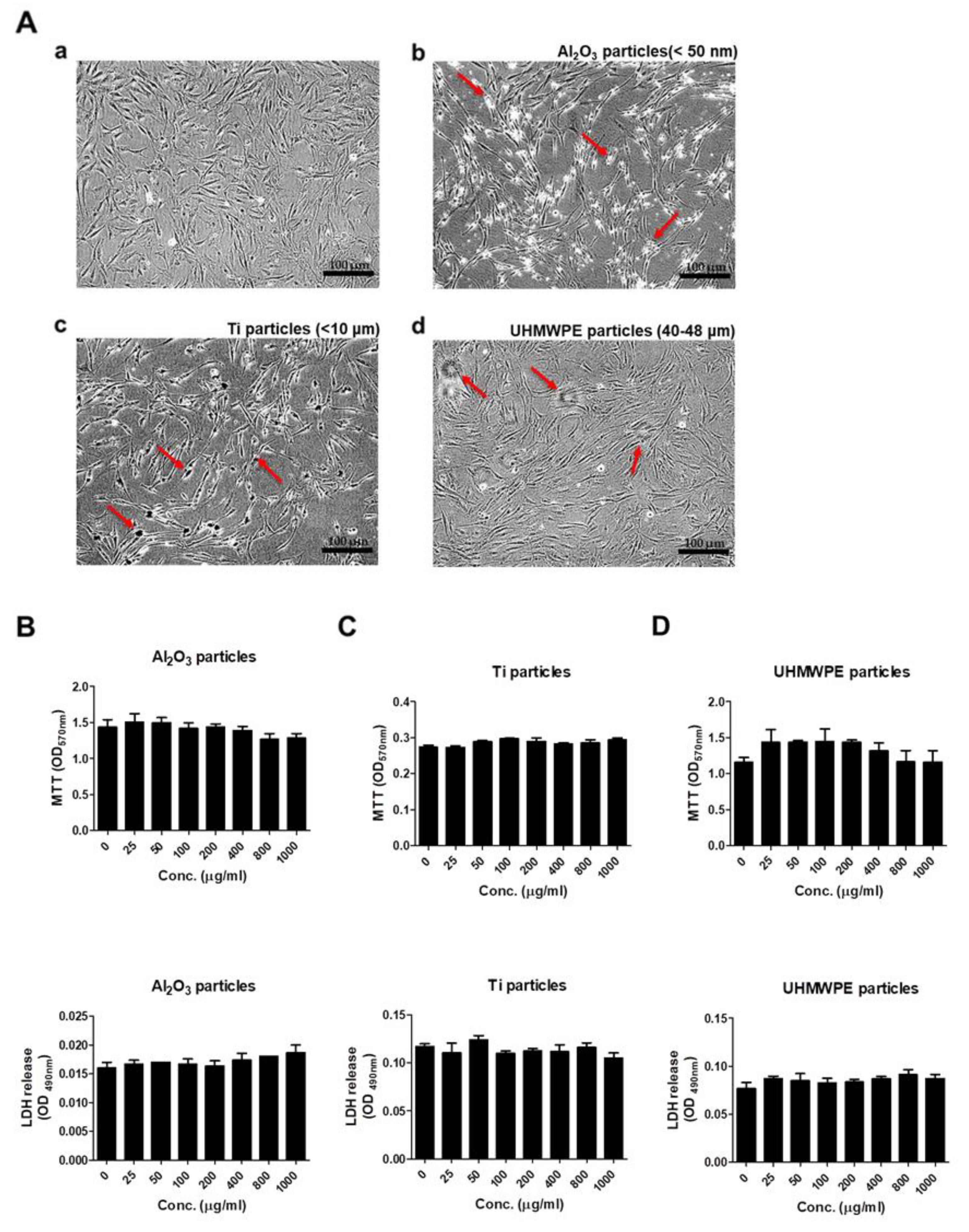
3.1. Comparison of Cell Morphology, Viability, and Cytotoxicity after Stimulating Osteoprogenitors with Different Types of Wear Particles
3.2. Al2O3 Particles Suppress ALP Activity of Osteoprogenitors
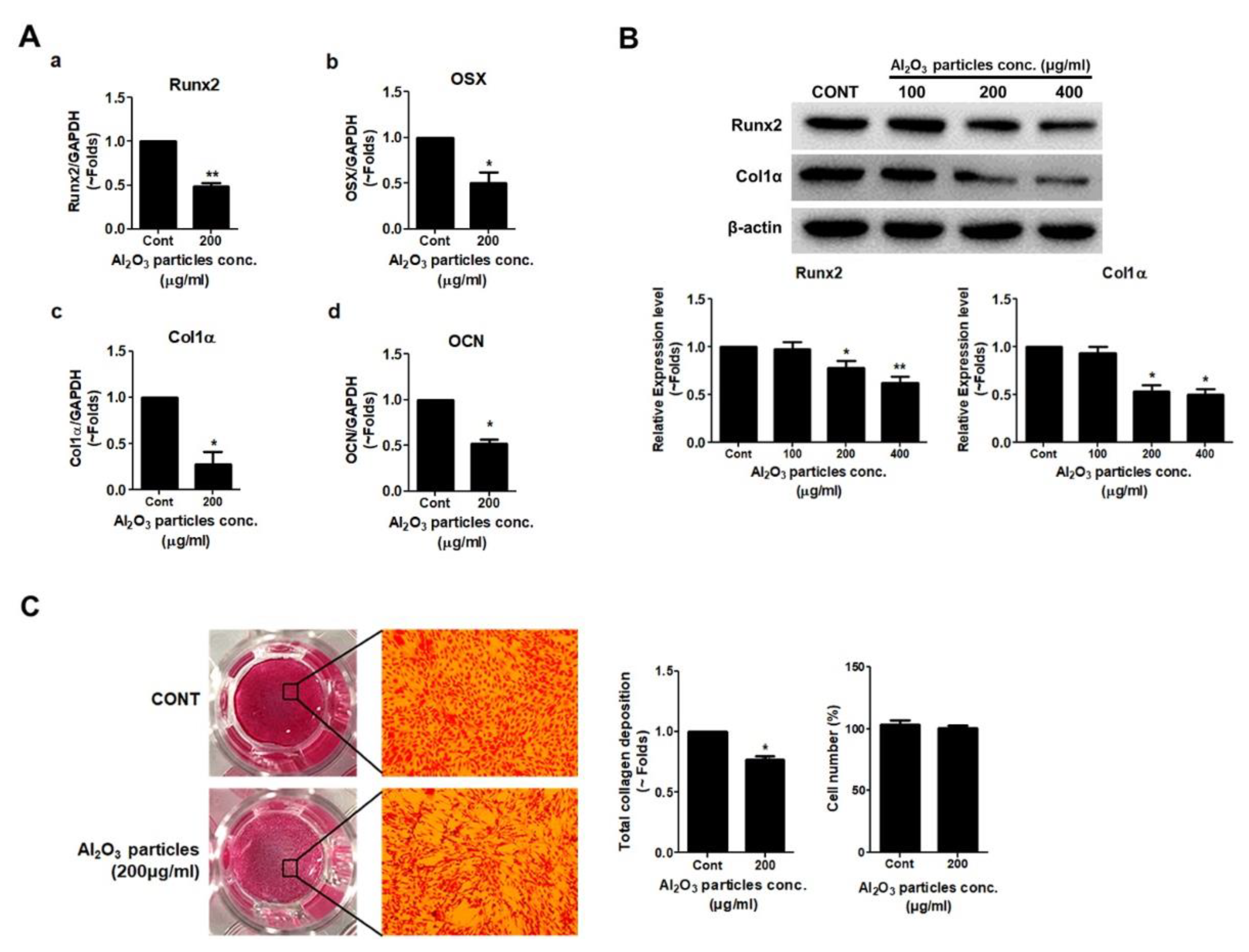
3.3. Effects of Al2O3 Particles on the Osteogenic Activity of Osteoprogenitors
3.4. Al2O3 Particles Induce Inflammation in Osteoprogenitors
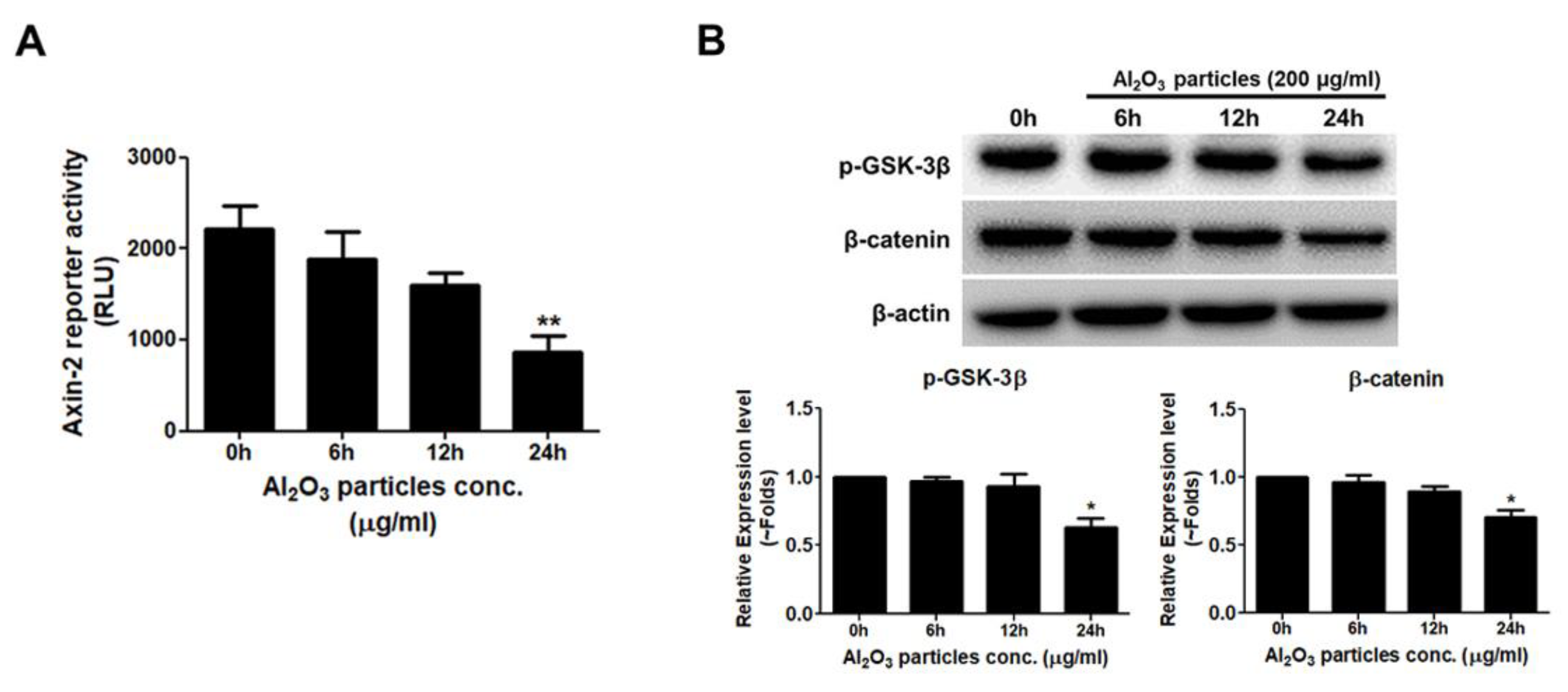
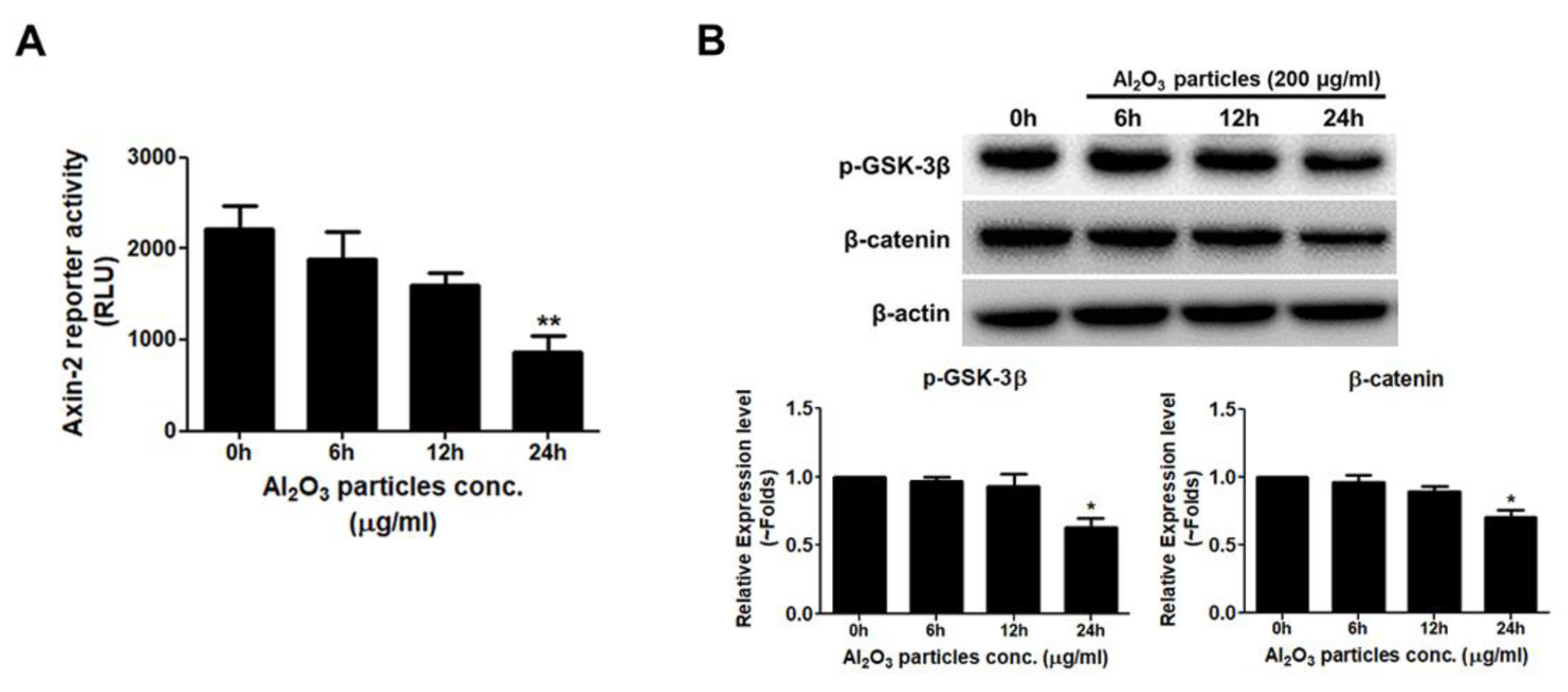
3.5. Al2O3 Particles Reduce the Activity of the WNT/β-Catenin Signal Pathway
3.6. Al2O3 Particles Increase the mRNA Expression Level of the WNT/β-Catenin Signaling Pathway Antagonists
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Marsh, M.; Newman, S. Trends and developments in hip and knee arthroplasty technology. J. Rehabil. Assist. Technol. Eng. 2021, 8, 2055668320952043. [Google Scholar] [CrossRef] [PubMed]
- Kurtz, S.; Ong, K.; Lau, E.; Mowat, F.; Halpern, M. Projections of primary and revision hip and knee arthroplasty in the United States from 2005 to 2030. J. Bone Jt. Surg. Am. 2007, 89, 780–785. [Google Scholar] [CrossRef]
- Kim, A.M.; Kang, S.; Park, J.H.; Yoon, T.H.; Kim, Y. Geographic variation and factors associated with rates of knee arthroplasty in Korea-a population based ecological study. BMC Musculoskelet. Disord. 2019, 20, 400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saini, M.; Singh, Y.; Arora, P.; Arora, V.; Jain, K. Implant biomaterials: A comprehensive review. World J. Clin. Cases 2015, 3, 52–57. [Google Scholar] [CrossRef]
- Kim, Y.H.; Choi, Y.; Kim, J.S. Osteolysis in well-functioning fixed- and mobile-bearing TKAs in younger patients. Clin. Orthop. Relat. Res. 2010, 468, 3084–3093. [Google Scholar] [CrossRef] [Green Version]
- Jagga, S.; Sharma, A.R.; Bhattacharya, M.; Chakraborty, C.; Lee, S.S. Influence of single nucleotide polymorphisms (SNPs) in genetic susceptibility towards periprosthetic osteolysis. Genes Genom. 2019, 41, 1113–1125. [Google Scholar] [CrossRef]
- Bozic, K.J.; Kurtz, S.M.; Lau, E.; Ong, K.; Vail, T.P.; Berry, D.J. The epidemiology of revision total hip arthroplasty in the United States. J. Bone Jt. Surg. Am. 2009, 91, 128–133. [Google Scholar] [CrossRef]
- Kurtz, S.; Mowat, F.; Ong, K.; Chan, N.; Lau, E.; Halpern, M. Prevalence of primary and revision total hip and knee arthroplasty in the United States from 1990 through 2002. J. Bone Jt. Surg. Am. 2005, 87, 1487–1497. [Google Scholar] [CrossRef]
- Bozic, K.J.; Kurtz, S.M.; Lau, E.; Ong, K.; Chiu, V.; Vail, T.P.; Rubash, H.E.; Berry, D.J. The epidemiology of revision total knee arthroplasty in the United States. Clin. Orthop. Relat. Res. 2010, 468, 45–51. [Google Scholar] [CrossRef] [Green Version]
- Callaghan, J.J.; O’Rourke, M.R.; Saleh, K.J. Why knees fail: Lessons learned. J. Arthroplast. 2004, 19, 31–34. [Google Scholar] [CrossRef]
- Harris, W.H. The problem is osteolysis. Clin. Orthop. Relat. Res. 1995, 311, 46–53. [Google Scholar]
- Ingham, E.; Fisher, J. The role of macrophages in osteolysis of total joint replacement. Biomaterials 2005, 26, 1271–1286. [Google Scholar] [CrossRef]
- Ren, W.; Yang, S.Y.; Fang, H.W.; Hsu, S.; Wooley, P.H. Distinct gene expression of receptor activator of nuclear factor-kappaB and rank ligand in the inflammatory response to variant morphologies of UHMWPE particles. Biomaterials 2003, 24, 4819–4826. [Google Scholar] [CrossRef]
- Bozic, K.J.; Ong, K.; Lau, E.; Kurtz, S.M.; Vail, T.P.; Rubash, H.E.; Berry, D.J. Risk of complication and revision total hip arthroplasty among Medicare patients with different bearing surfaces. Clin. Orthop. Relat. Res. 2010, 468, 2357–2362. [Google Scholar] [CrossRef] [Green Version]
- Hundric-Hašpl, Ž.; Pecina, M.; Haspl, M.; Tomicic, M.; Jukic, I. Plasma cytokines as markers of aseptic prosthesis loosening. Clin. Orthop. Relat. Res. 2006, 453, 299–304. [Google Scholar] [CrossRef]
- Stea, S.; Visentin, M.; Granchi, D.; Ciapetti, G.; Donati, M.E.; Sudanese, A.; Zanotti, C.; Toni, A. Cytokines and osteolysis around total hip prostheses. Cytokine 2000, 12, 1575–1579. [Google Scholar] [CrossRef]
- Zhang, L.; Jia, T.H.; Chong, A.C.; Bai, L.; Yu, H.; Gong, W.; Wooley, P.H.; Yang, S.Y. Cell-based osteoprotegerin therapy for debris-induced aseptic prosthetic loosening on a murine model. Gene 2010, 17, 1262–1269. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.R.; Jagga, S.; Chakraborty, C.; Lee, S.S. Fibroblast-Like-Synoviocytes Mediate Secretion of Pro-Inflammatory Cytokines via ERK and JNK MAPKs in Ti-Particle-Induced Osteolysis. Materials 2020, 13, 3628. [Google Scholar] [CrossRef]
- Jagga, S.; Sharma, A.R.; Lee, Y.H.; Nam, J.S.; Lee, S.S. Sclerostin-Mediated Impaired Osteogenesis by Fibroblast-Like Synoviocytes in the Particle-Induced Osteolysis Model. Front. Mol. Biosci. 2021, 8, 666295. [Google Scholar] [CrossRef]
- Mandelin, J.; Li, T.F.; Liljestrom, M.; Kroon, M.E.; Hanemaaijer, R.; Santavirta, S.; Konttinen, Y.T. Imbalance of RANKL/RANK/OPG system in interface tissue in loosening of total hip replacement. J. Bone Jt. Surg. Br. 2003, 85, 1196–1201. [Google Scholar] [CrossRef] [Green Version]
- Vermes, C.; Chandrasekaran, R.; Jacobs, J.J.; Galante, J.O.; Roebuck, K.A.; Glant, T.T. The effects of particulate wear debris, cytokines, and growth factors on the functions of MG-63 osteoblasts. J. Bone Jt. Surg. Am. 2001, 83, 201–211. [Google Scholar] [CrossRef]
- Lassus, J.; Salo, J.; Jiranek, W.A.; Santavirta, S.; Nevalainen, J.; Matucci-Cerinic, M.; Horak, P.; Konttinen, Y. Macrophage activation results in bone resorption. Clin. Orthop. Relat. Res. 1998, 352, 7–15. [Google Scholar] [CrossRef]
- Nam, J.S.; Sharma, A.R.; Jagga, S.; Lee, D.H.; Sharma, G.; Nguyen, L.T.; Lee, Y.H.; Chang, J.D.; Chakraborty, C.; Lee, S.S. Suppression of osteogenic activity by regulation of WNT and BMP signaling during titanium particle induced osteolysis. J. Biomed. Mater. Res. A 2017, 105, 912–926. [Google Scholar] [CrossRef]
- Vermes, C.; Glant, T.T.; Hallab, N.J.; Fritz, E.A.; Roebuck, K.A.; Jacobs, J.J. The potential role of the osteoblast in the development of periprosthetic osteolysis: Review of in vitro osteoblast responses to wear debris, corrosion products, and cytokines and growth factors. J. Arthroplast. 2001, 16, 95–100. [Google Scholar] [CrossRef]
- Chiu, R.; Smith, K.E.; Ma, G.K.; Ma, T.; Smith, R.L.; Goodman, S.B. Polymethylmethacrylate particles impair osteoprogenitor viability and expression of osteogenic transcription factors Runx2, osterix, and Dlx5. J. Orthop. Res. 2010, 28, 571–577. [Google Scholar] [CrossRef]
- Liang, M.H.; Chuang, D.M. Differential roles of glycogen synthase kinase-3 isoforms in the regulation of transcriptional activation. J. Biol. Chem. 2006, 281, 30479–30484. [Google Scholar] [CrossRef] [Green Version]
- Gu, Y.; Wang, Z.; Shi, J.; Wang, L.; Hou, Z.; Guo, X.; Tao, Y.; Wu, X.; Zhou, W.; Liu, Y.; et al. Titanium particle-induced osteogenic inhibition and bone destruction are mediated by the GSK-3beta/beta-catenin signal pathway. Cell Death Dis. 2017, 8, e2878. [Google Scholar] [CrossRef] [Green Version]
- Gordon, M.D.; Nusse, R. Wnt signaling: Multiple pathways, multiple receptors, and multiple transcription factors. J. Biol. Chem. 2006, 281, 22429–22433. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Z.; Zylstra-Diegel, C.R.; Schumacher, C.A.; Baker, J.J.; Carpenter, A.C.; Rao, S.; Yao, W.; Guan, M.; Helms, J.A.; Lane, N.E.; et al. Wntless functions in mature osteoblasts to regulate bone mass. Proc. Natl. Acad. Sci. USA 2012, 109, E2197–E2204. [Google Scholar] [CrossRef] [Green Version]
- Gibon, E.; Amanatullah, D.F.; Loi, F.; Pajarinen, J.; Naabeshima, A.; Yao, Z.; Hamadouche, M.; Goodman, S.B. The biological response to orthopaedic implants for joint replacement: Part I: Metals. J. Biomed. Mater. Res. Appl. Biomater. 2017, 105, 2162–2173. [Google Scholar] [CrossRef] [Green Version]
- Klinder, A.; Seyfarth, A.; Hansmann, D.; Bader, R.; Jonitz-Heincke, A. Inflammatory Response of Human Peripheral Blood Mononuclear Cells and Osteoblasts Incubated With Metallic and Ceramic Submicron Particles. Front. Immunol. 2018, 9, 831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dean, D.D.; Schwartz, Z.; Liu, Y.; Blanchard, C.R.; Agrawal, C.M.; Mabrey, J.D.; Sylvia, V.L.; Lohmann, C.H.; Boyan, B.D. The effect of ultra-high molecular weight polyethylene wear debris on MG63 osteosarcoma cells in vitro. J. Bone Jt. Surg. Am. 1999, 81, 452–461. [Google Scholar] [CrossRef] [PubMed]
- Chiu, R.; Ma, T.; Smith, R.L.; Goodman, S.B. Ultrahigh molecular weight polyethylene wear debris inhibits osteoprogenitor proliferation and differentiation in vitro. J. Biomed. Mater. Res. 2009, 89, 242–247. [Google Scholar] [CrossRef]
- Chiu, R.; Ma, T.; Smith, R.L.; Goodman, S.B. Kinetics of polymethylmethacrylate particle-induced inhibition of osteoprogenitor differentiation and proliferation. J. Orthop. Res. 2007, 25, 450–457. [Google Scholar] [CrossRef]
- Pinzone, J.J.; Hall, B.M.; Thudi, N.K.; Vonau, M.; Qiang, Y.W.; Rosol, T.J.; Shaughnessy, J.D., Jr. The role of Dickkopf-1 in bone development, homeostasis, and disease. Blood 2009, 113, 517–525. [Google Scholar] [CrossRef] [Green Version]
- Sharma, G.; Sharma, A.R.; Seo, E.M.; Nam, J.S. Genetic polymorphism in extracellular regulators of Wnt signaling pathway. Biomed. Res. Int 2015, 2015, 847529. [Google Scholar] [CrossRef]
- Abu-Amer, Y.; Darwech, I.; Clohisy, J.C. Aseptic loosening of total joint replacements: Mechanisms underlying osteolysis and potential therapies. Arthritis Res. 2007, 9 (Suppl S1), S6. [Google Scholar] [CrossRef] [Green Version]
- Chiu, R.; Ma, T.; Smith, R.L.; Goodman, S.B. Polymethylmethacrylate particles inhibit osteoblastic differentiation of MC3T3-E1 osteoprogenitor cells. J. Orthop. Res. 2008, 26, 932–936. [Google Scholar] [CrossRef]
- Lei, P.; Dai, Z.; Zhang, Y.S.; Liu, H.; Niu, W.; Li, K.; Wang, L.; Hu, Y.; Xie, J. Macrophage inhibits the osteogenesis of fibroblasts in ultrahigh molecular weight polyethylene (UHMWPE) wear particle-induced osteolysis. J. Orthop. Surg. Res. 2019, 14, 80. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.Y.; Ren, W.; Park, Y.; Sieving, A.; Hsu, S.; Nasser, S.; Wooley, P.H. Diverse cellular and apoptotic responses to variant shapes of UHMWPE particles in a murine model of inflammation. Biomaterials 2002, 23, 3535–3543. [Google Scholar] [CrossRef]
- Lee, S.S.; Sharma, A.R.; Choi, B.S.; Jung, J.S.; Chang, J.D.; Park, S.; Salvati, E.A.; Purdue, E.P.; Song, D.K.; Nam, J.S. The effect of TNFalpha secreted from macrophages activated by titanium particles on osteogenic activity regulated by WNT/BMP signaling in osteoprogenitor cells. Biomaterials 2012, 33, 4251–4263. [Google Scholar] [CrossRef]
- Nevelos, J.E.; Ingham, E.; Doyle, C.; Fisher, J.; Nevelos, A.B. Analysis of retrieved alumina ceramic components from Mittelmeier total hip prostheses. Biomaterials 1999, 20, 1833–1840. [Google Scholar] [CrossRef]
- Bader, R.; Bergschmidt, P.; Fritsche, A.; Ansorge, S.; Thomas, P.; Mittelmeier, W. Alternative materials and solutions in total knee arthroplasty for patients with metal allergy. Orthopade 2008, 37, 136–142. [Google Scholar] [CrossRef]
- Malem, D.; Nagy, M.T.; Ghosh, S.; Shah, B. Catastrophic failure of ceramic-on-ceramic total hip arthroplasty presenting as squeaking hip. BMJ Case Rep. 2013, 2013, bcr-2013. [Google Scholar] [CrossRef] [Green Version]
- Toni, A.; Traina, F.; Stea, S.; Sudanese, A.; Visentin, M.; Bordini, B.; Squarzoni, S. Early diagnosis of ceramic liner fracture. Guidelines based on a twelve-year clinical experience. J. Bone Jt. Surg. Am. 2006, 88 (Suppl. S4), 55–63. [Google Scholar] [CrossRef]
- Regis, D.; Sandri, A.; Bartolozzi, P. Delayed diagnosis of low-symptomatic ceramic acetabular liner fracture in ceramic-on-ceramic total hip arthroplasty. Orthopedics 2008, 31, 1–3. [Google Scholar] [CrossRef]
- Jeffers, J.R.; Walter, W.L. Ceramic-on-ceramic bearings in hip arthroplasty: State of the art and the future. J. Bone Jt. Surg Br. 2012, 94, 735–745. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Wang, C.; Li, Z.; Wang, H.; He, J.; Zhu, J.; Zhang, Y.; Shen, C.; Xiao, F.; Gao, Y.; et al. Nano-sized Al2O3 particle-induced autophagy reduces osteolysis in aseptic loosening of total hip arthroplasty by negative feedback regulation of RANKL expression in fibroblasts. Cell Death Dis. 2018, 9, 840. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.F.; Zheng, Y.F.; Qin, L. A comprehensive biological evaluation of ceramic nanoparticles as wear debris. Nanomedicine 2011, 7, 975–982. [Google Scholar] [CrossRef]
- Rodrigo, A.; Valles, G.; Saldana, L.; Rodriguez, M.; Martinez, M.E.; Munuera, L.; Vilaboa, N. Alumina particles influence the interactions of cocultured osteoblasts and macrophages. J. Orthop. Res. 2006, 24, 46–54. [Google Scholar] [CrossRef]
- Lohmann, C.H.; Schwartz, Z.; Koster, G.; Jahn, U.; Buchhorn, G.H.; MacDougall, M.J.; Casasola, D.; Liu, Y.; Sylvia, V.L.; Dean, D.D.; et al. phagocytosis of wear debris by osteoblasts affects differentiation and local factor production in a manner dependent on particle composition. Biomaterials 2000, 21, 551–561. [Google Scholar] [CrossRef]
- Hatton, A.; Nevelos, J.; Nevelos, A.; Banks, R.; Fisher, J.; Ingham, E. Alumina–alumina artificial hip joints. Part I: A histological analysis and characterisation of wear debris by laser capture microdissection of tissues retrieved at revision. Biomaterials 2002, 23, 3429–3440. [Google Scholar] [CrossRef]
- Hatton, A.; Nevelos, J.; Matthews, J.; Fisher, J.; Ingham, E. Effects of clinically relevant alumina ceramic wear particles on TNF-α production by human peripheral blood mononuclear phagocytes. Biomaterials 2003, 24, 1193–1204. [Google Scholar] [CrossRef]
- Katagiri, T.; Takahashi, N. Regulatory mechanisms of osteoblast and osteoclast differentiation. Oral. Dis. 2002, 8, 147–159. [Google Scholar] [CrossRef]
- Bitar, D.; Parvizi, J. Biological response to prosthetic debris. World J. Orthop. 2015, 6, 172–189. [Google Scholar] [CrossRef]
- Deng, Z.L.; Sharff, K.A.; Tang, N.; Song, W.X.; Luo, J.; Luo, X.; Chen, J.; Bennett, E.; Reid, R.; Manning, D.; et al. Regulation of osteogenic differentiation during skeletal development. Front Biosci. 2008, 13, 2001–2021. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′-3′) |
|---|---|
| Cox2 | F: AAGTGCGGTGCAAACTTTCT R: TCTCGGTGGCTGGTAGTGA |
| Runx2 | F: GGAAAGGAGGCACAAAGAAGCCAT R: AGTCCATTGGTGCTTGAGAAGGGA |
| Osterix (OSX) | F: TTCTCCTGGCAAAGACGGAC R: AGGAAGCTGAAGTCATAACCGCCA |
| Collagen 1α (Col1α) | F: TGCTTGTGACGAGCTATCAG R: GAGGACAGGGAGGATCAAGT |
| Osteocalcin (OCN) | F: TTCAAAAGAAGTGCTGGAAAAGGT R: GATCATCTCTACCTGAGTGTCTTT |
| DKK1 | F: TCAGGTCCATTCTGGCCAACTCTT R: TGGGCATTCCCTCCCTTCCAATAA |
| DKK2 | F: ATGGCAGAATCTAGGAAGGCCACA R: CGAACCCTTCTTGCGTTGTTTGGT |
| DKK3 | F: AGCTGATGGAAGACACTCAGCACA R: TCCTGGTGCACATGGACTGTGTTA |
| DKK4 | F: ATGGTACTGGTGACCTTGCTTGGA R: TCCGCGGAGCTCTTGATGTTGTTA |
| sFRP1 | F: ACGAGTTGAAGTCAGAGGCCATC R: ACAGTCGGCACCGTTCTTCAG |
| sFRP2 | F: ATCCTGGAGACAAAGAGCAAGACC R: TGACCAGATACCGGAGCGTTGATG |
| sFRP3 | F: TGCAAATGTAAGCCTGTCAGAGC R: TCCACAACGGCGGTCACATC |
| sFRP4 | F: GTGGCGTTCAAGGATGATGCTTC R: TTACTGCGACTGGTGCGACTG |
| sFRP5 | F: CCCTGGACAACGACCTCTGC R: CACAAAGTCACTGGAGCACATCTG |
| Sclerostin (SOST) | F: AAAGGGAAGGGAGTGTGGAACGAA R: CGCAGGCTTTACATTTGGGTGGAA |
| WIF | F: CCACCTGAGGAGAGCTTGTACC R: TGGCATTCTTTGTTGGGCTTTCC |
| WISE | F: ACTGGATCGGAGGAGGCTATGG R: TGTGGCTGGACTCGTTGTGC |
| GAPDH | F: TCGTGGATCTGACGTGCCGCCTG R: CACCACCCTGTTGCTGTAGCCGTAT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, A.R.; Lee, Y.-H.; Gankhuyag, B.; Chakraborty, C.; Lee, S.-S. Effect of Alumina Particles on the Osteogenic Ability of Osteoblasts. J. Funct. Biomater. 2022, 13, 105. https://doi.org/10.3390/jfb13030105
Sharma AR, Lee Y-H, Gankhuyag B, Chakraborty C, Lee S-S. Effect of Alumina Particles on the Osteogenic Ability of Osteoblasts. Journal of Functional Biomaterials. 2022; 13(3):105. https://doi.org/10.3390/jfb13030105
Chicago/Turabian StyleSharma, Ashish Ranjan, Yeon-Hee Lee, Buyankhishig Gankhuyag, Chiranjib Chakraborty, and Sang-Soo Lee. 2022. "Effect of Alumina Particles on the Osteogenic Ability of Osteoblasts" Journal of Functional Biomaterials 13, no. 3: 105. https://doi.org/10.3390/jfb13030105
APA StyleSharma, A. R., Lee, Y.-H., Gankhuyag, B., Chakraborty, C., & Lee, S.-S. (2022). Effect of Alumina Particles on the Osteogenic Ability of Osteoblasts. Journal of Functional Biomaterials, 13(3), 105. https://doi.org/10.3390/jfb13030105