
Macro- and Micronutrient Cycling and Crucial Linkages to Geochemical Processes in Mangrove Ecosystems


Abstract
1. Introduction
2. Limiting Nutrients: N and P Cycling
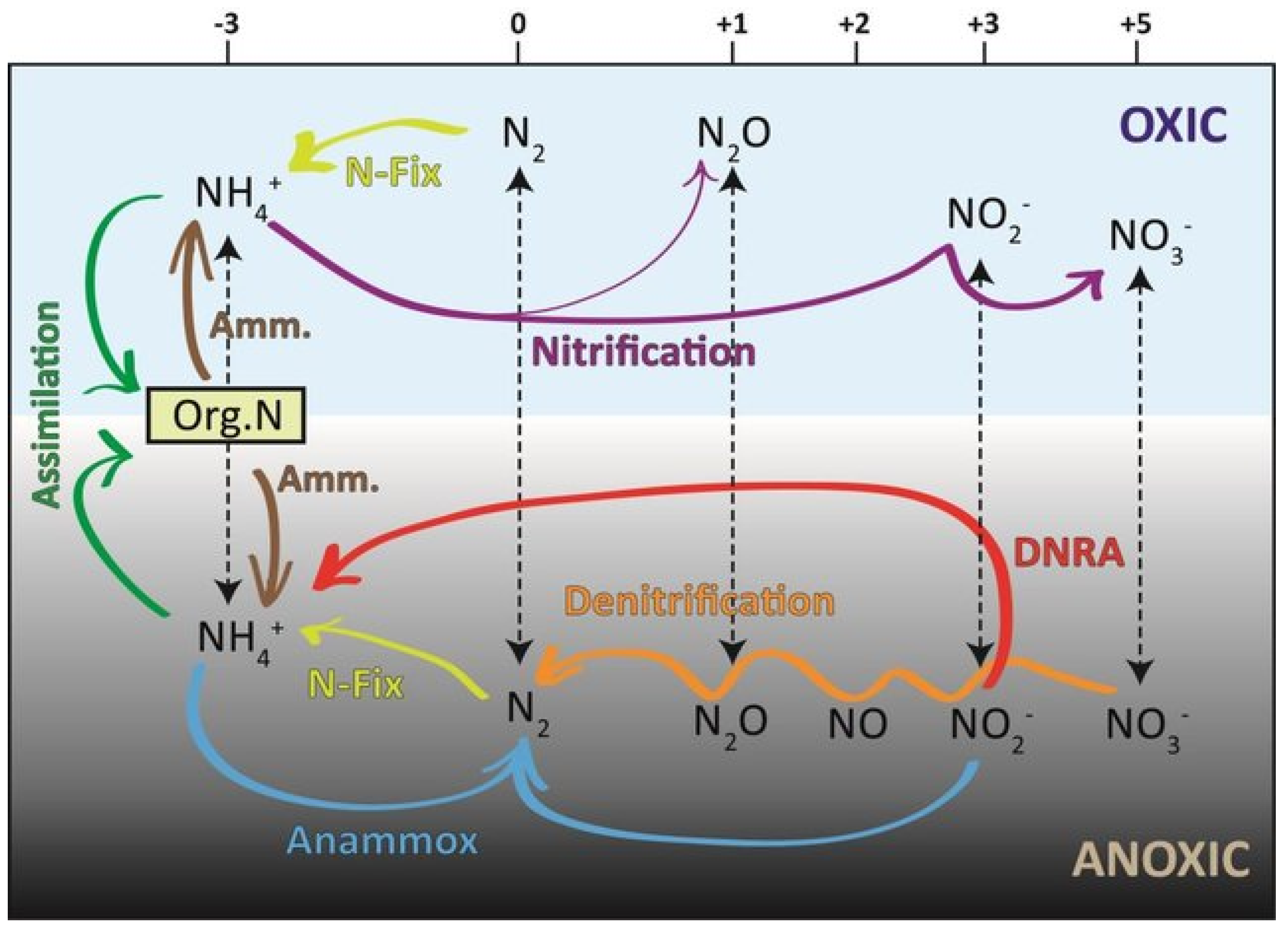
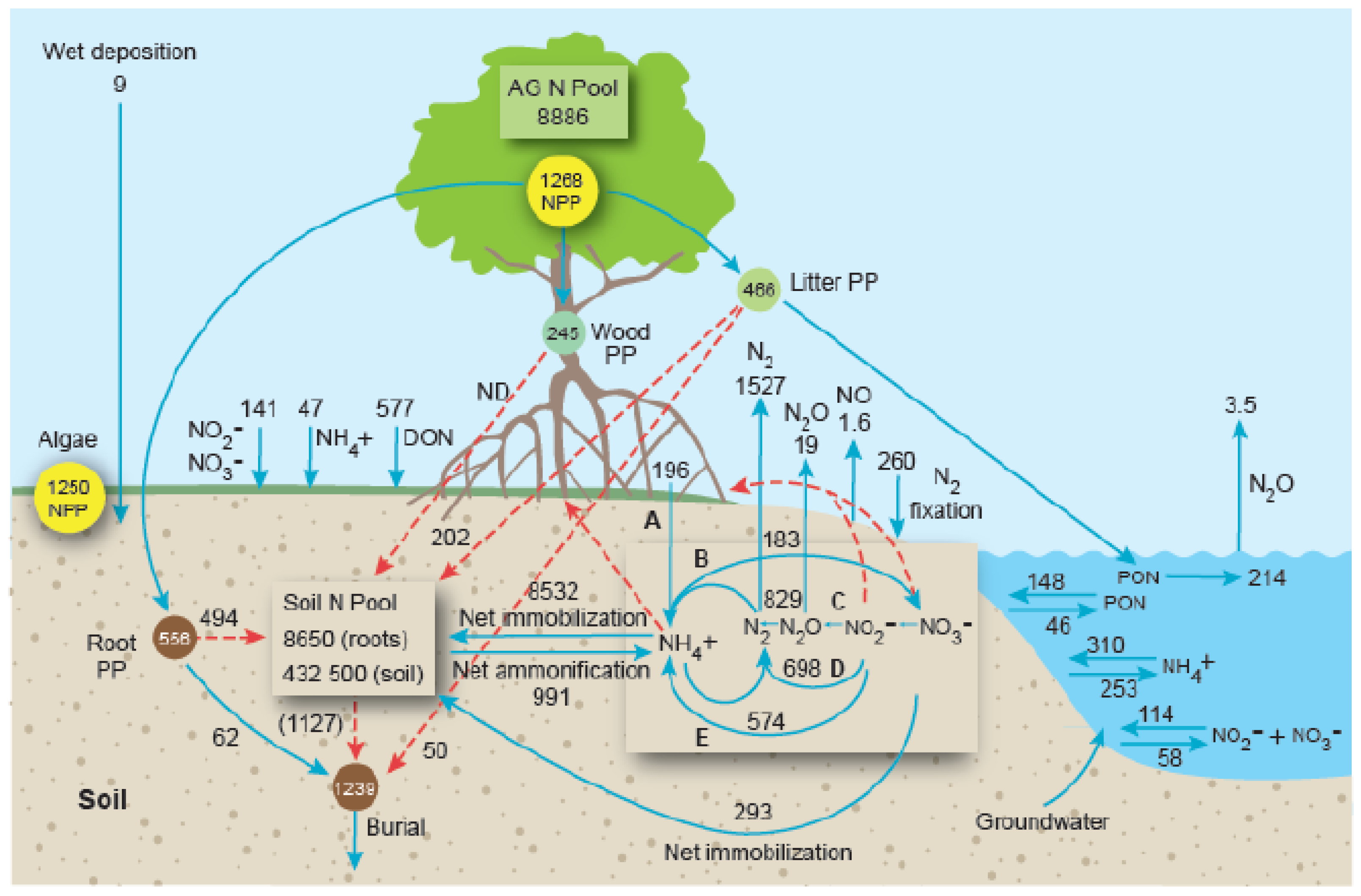
2.1. Nitrogen
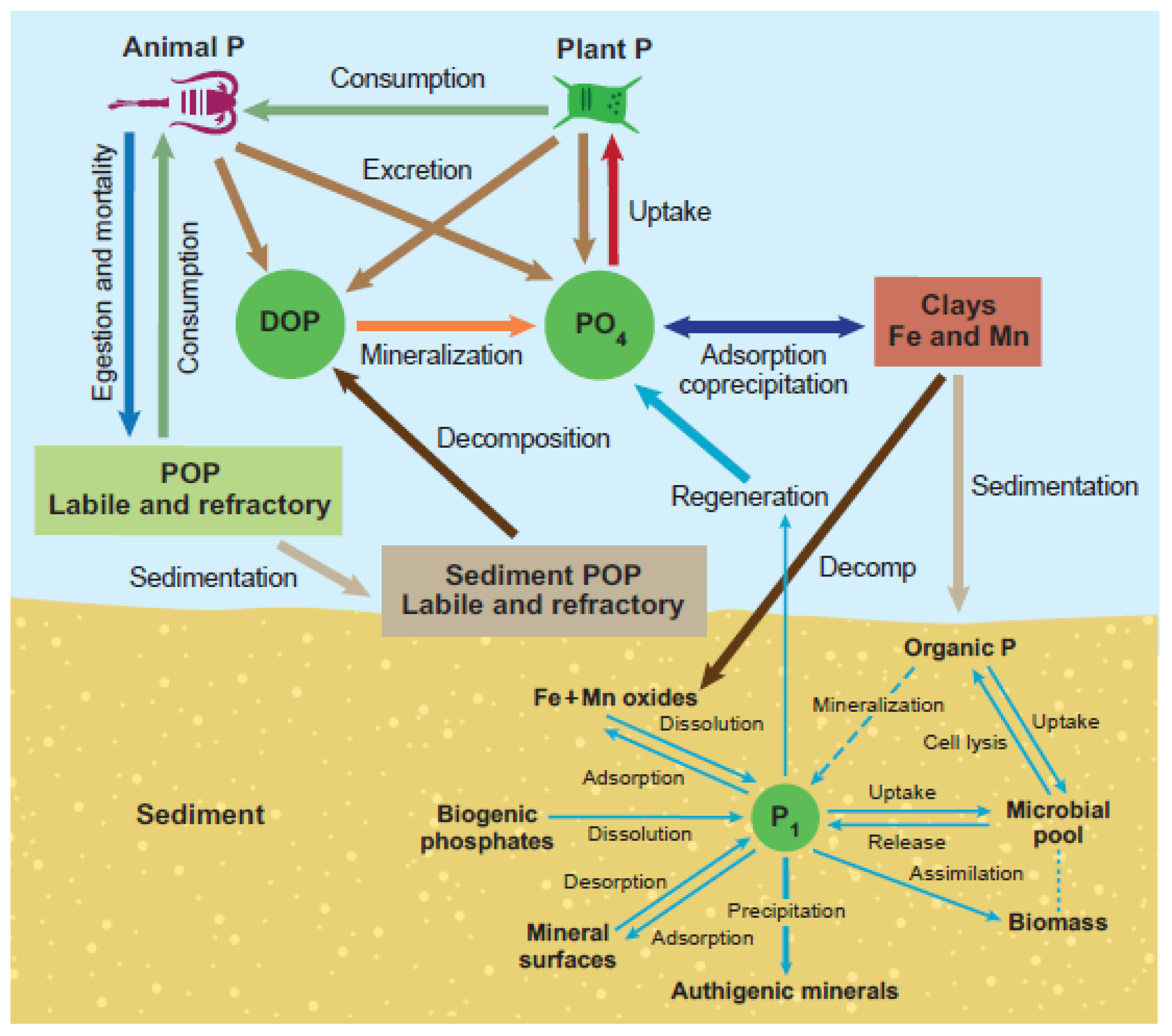
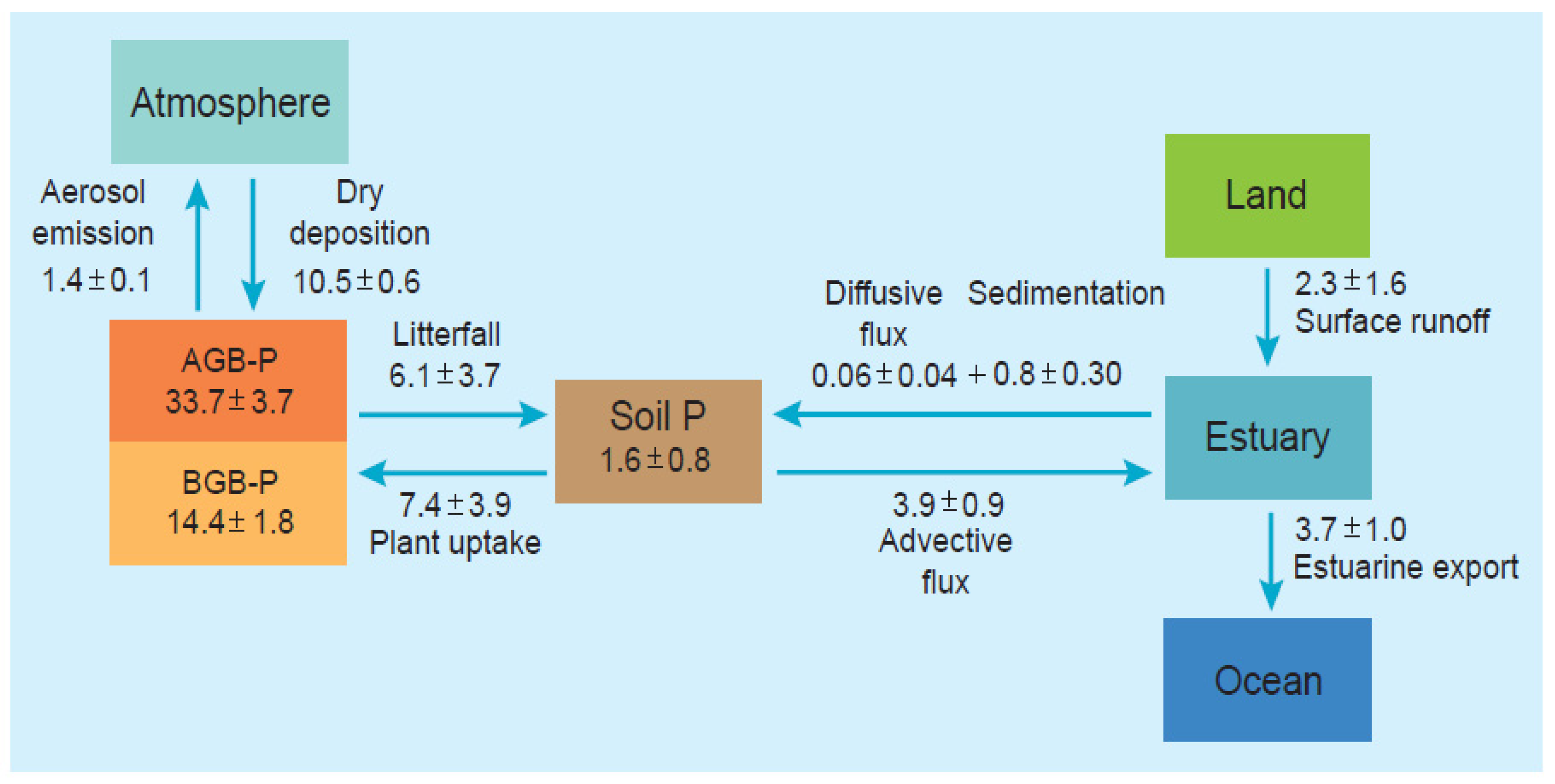
2.2. Phosphorus
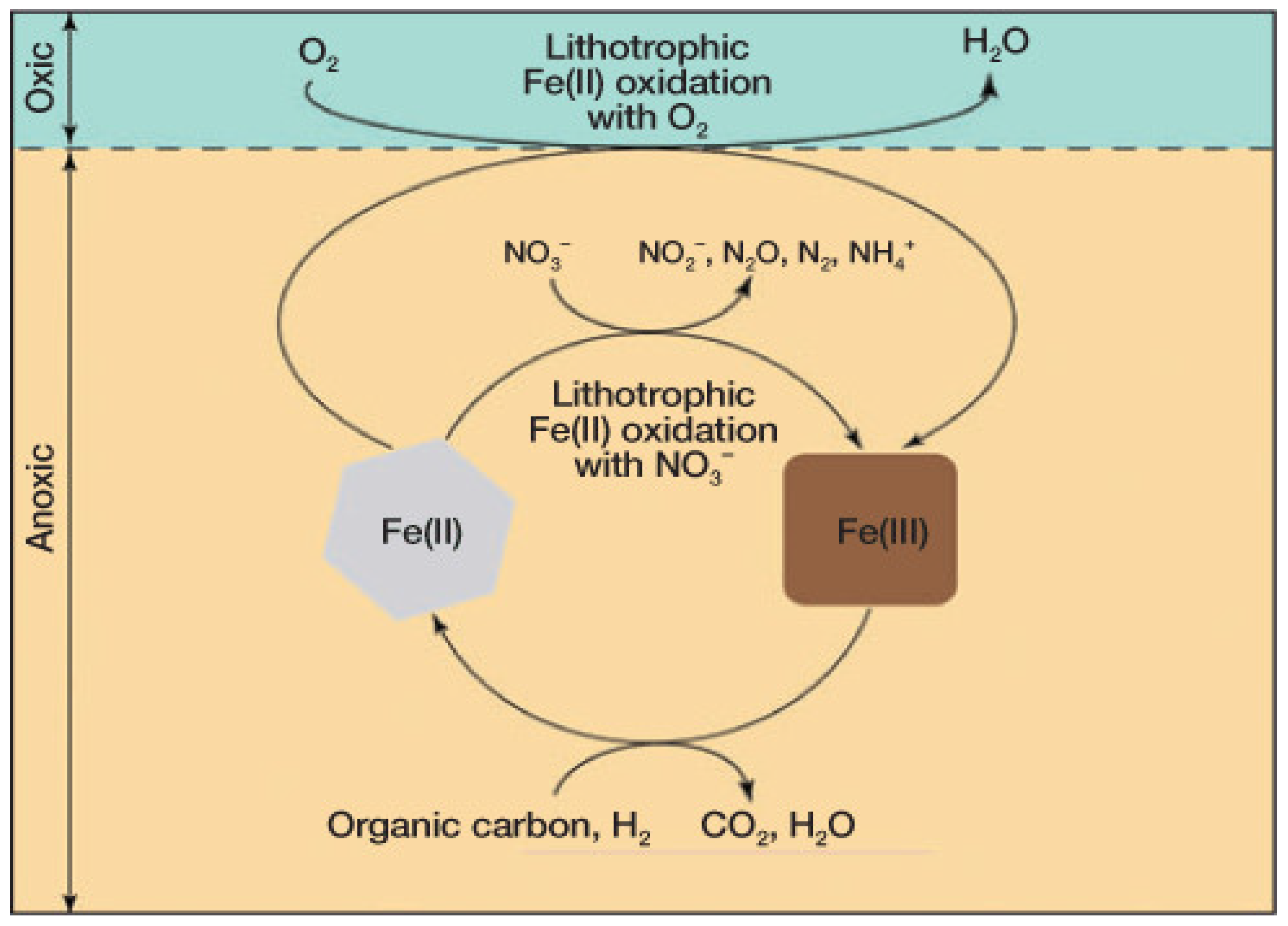
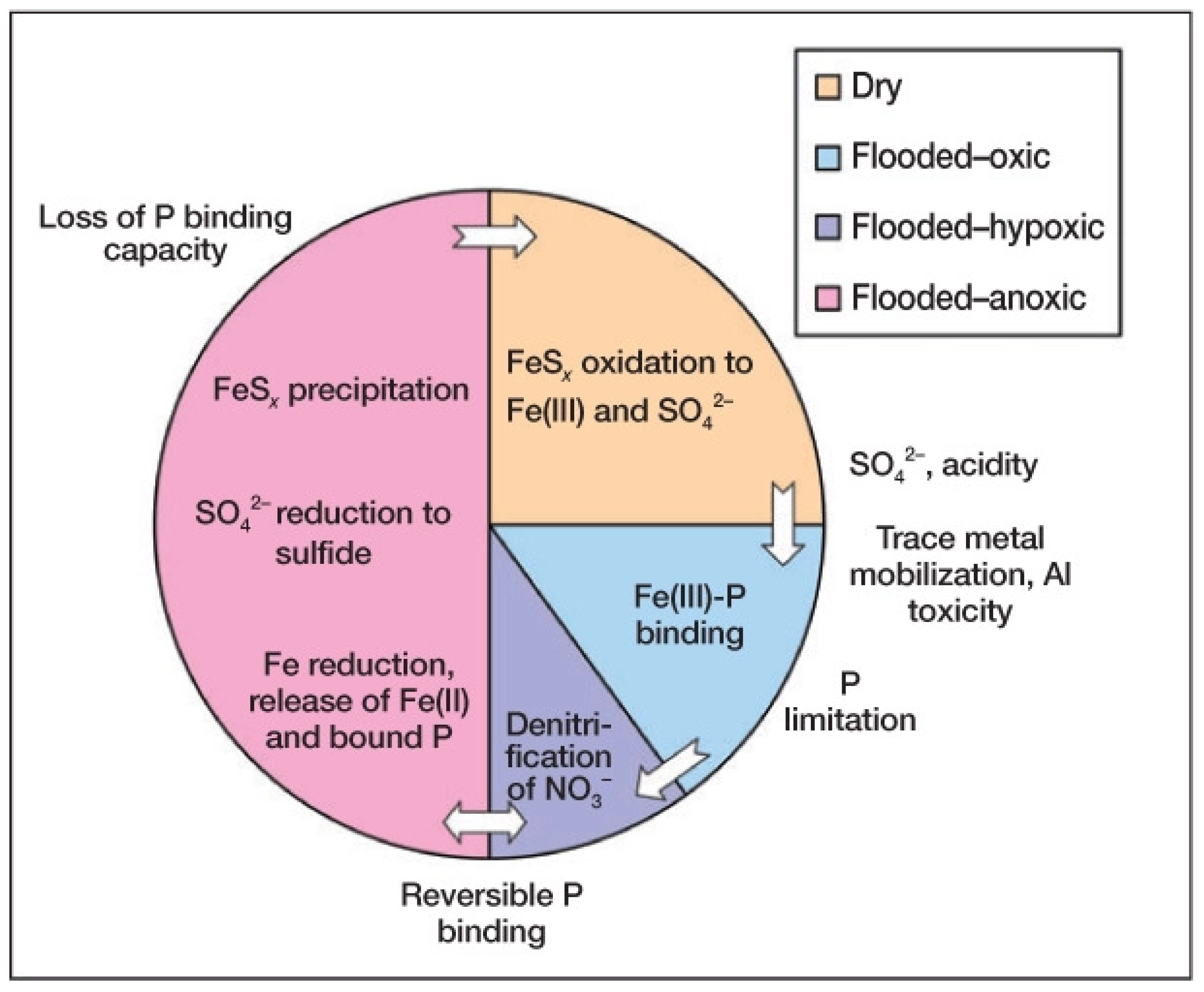
3. Coupled Biogeochemistry and Geochemistry: Sulfur (S) and Iron (Fe)
4. Abundant Macronutrients: Magnesium (Mg), Calcium (Ca), and Potassium (K)
5. Silicon Requirements
6. Micronutrients
6.1. Manganese (Mn)
6.2. Copper (Cu), Zinc (Zn) and Molybdenum (Mo)
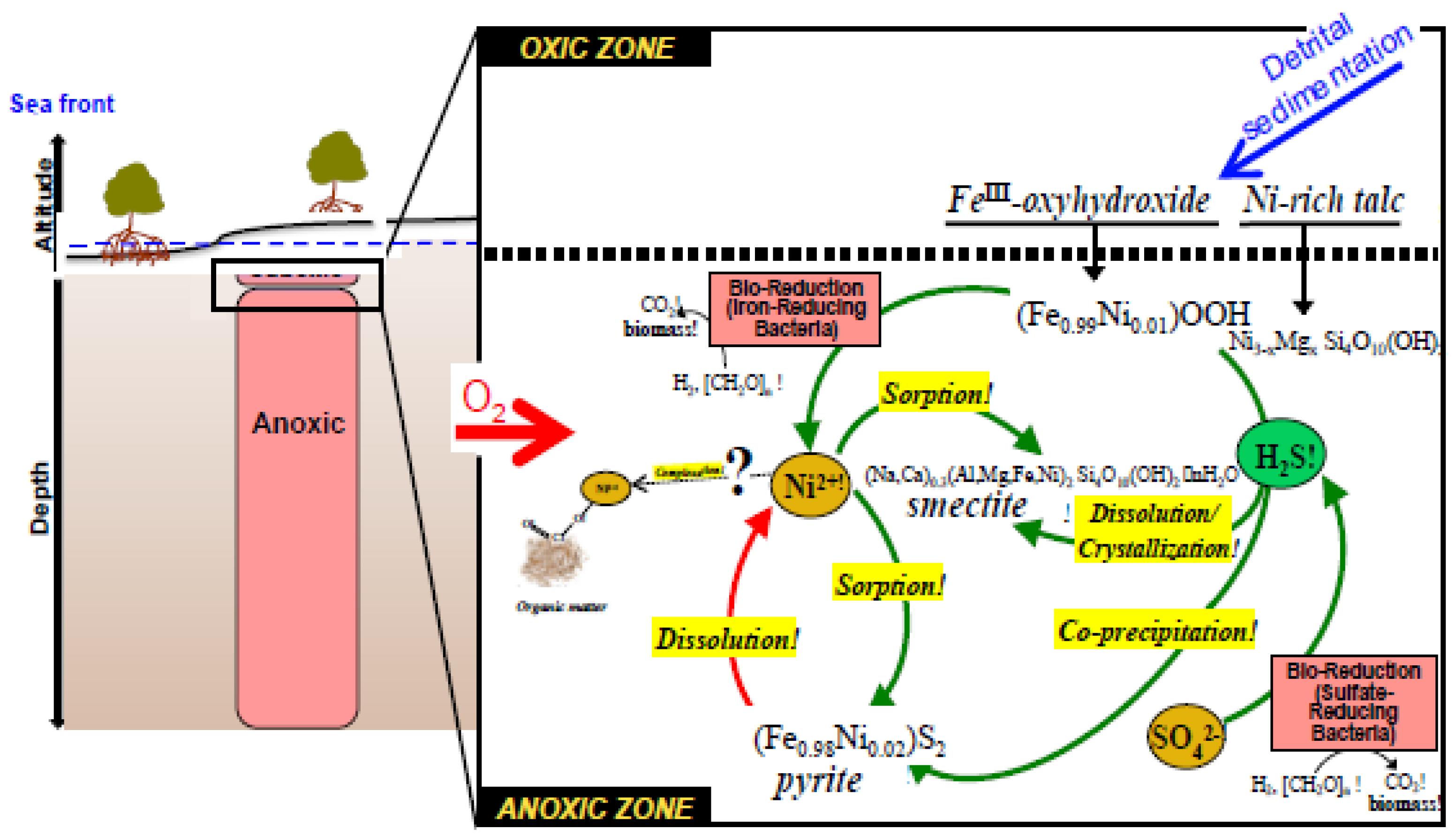
6.3. Nickel (Ni) and Cobalt (Co)
7. Discussion
7.1. An Emerging Picture
7.2. The Overlooked Role of Micronutrients
7.3. Perspectives for Further Research
8. Conclusions
- Nutrients are stored primarily in the soil pool but also in leaves and roots, in the former where nutrients are utilized in photosynthesis and in the latter where they are initially taken up from soils, respectively. Nutrient stocks in wood are low. The C:N:P stoichiometry of burial in mangrove soils is 95:7:1, which is considerably less than in mangrove matter (C:N:P = 3405:23:1) indicating preferential burial of N and P in relation to C.
- Depending on growth stage, species, and environmental conditions, mangrove growth can be limited not only by N and P, but also by low interstitial water concentrations of Fe, Cu, Zn and Mo.
- Only the N cycle is well understood, with little N export and high N retention and conservation; NH4+ immobilization into the soil pool is the largest N transformation process.
- Fe, Mn, S and Mo affect N metabolism and photosynthesis via organic matter diagenesis and P availability may be coupled to Fe-S redox oscillations during drying and flooding of soils. Fe is involved in nitrification, denitrification and anammox and Mo is involved in NO3− reduction and N2-fixation.
- Zn addition to plants results in increased accumulation of free amino acids, sugars, and antioxidants.
- Soil Mg, K, Mn, Zn and Ni pool sizes decrease as mangrove NP increases, suggesting increasing transformation and uptake, and more rapid turnover, than in less productive forests. Solid-phase Co shows the opposite pattern.
- The exchange of mangrove macro- and micronutrients with adjacent coastal waters have major impacts on nutrient cycles in the tropical coastal ocean. This includes not only N and P exchanges, but also Mn and Mo fluxes.
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Alongi, D.M. The Energetics of Mangrove Forests; Springer Science: Dordrecht, The Netherlands, 2009. [Google Scholar]
- Twilley, R.R.; Day, J.W., Jr. Mangrove wetlands. In Estuarine Ecology, 2nd ed.; Day, J.W., Jr., Crump, B.C., Kemp, W.M., Yañez-Arancibia, A., Eds.; Wiley-Blackwell: New York, NY, USA, 2013; pp. 165–202. [Google Scholar]
- Ball, M.C. Ecophysiology of mangroves. Trees 1988, 2, 129–142. [Google Scholar] [CrossRef]
- Bashan, Y.; Holguin, G. Plant growth-promoting bacteria: A potential tool for arid mangrove reforestation. Trees 2002, 16, 159–166. [Google Scholar] [CrossRef]
- Shuman, L.M. Mineral nutrition. In Plant-Environment Interactions, 2nd ed.; Wilkinson, R.E., Ed.; Marcel Dekker: New York, NY, USA, 2000; pp. 65–109. [Google Scholar]
- Reef, R.; Feller, I.C.; Lovelock, C.E. Nutrition of mangroves. Tree Physiol. 2010, 30, 1148–1160. [Google Scholar]
- Alongi, D.M.; Clough, B.F.; Dixon, P.; Tirendi, F. Nutrient partitioning and storage in arid-zone forests of the mangrove Rhizophora stylosa and Avicennia marina. Trees 2003, 17, 51–60. [Google Scholar] [CrossRef]
- Li, M.S. Nutrient dynamics of a Futian mangrove forest in Shenzhen, South China. Estuar. Coast. Shelf Sci. 1997, 45, 463–472. [Google Scholar] [CrossRef]
- Alongi, D.M. Nitrogen cycling and mass balance in the world’s mangrove forests. Nitrogen 2020, 1, 167–189. [Google Scholar] [CrossRef]
- Alongi, D.M. The dynamics of benthic nutrient pools and fluxes in tropical mangrove forests. J. Mar. Res. 1996, 54, 123–148. [Google Scholar] [CrossRef]
- Zhang, M.; Luo, Y.; Lin, L.; Lin, X.; Hetharua, B.; Zhao, W.; Zhou, M.; Zhan, Q.; Xu, H.; Zheng, T.; et al. Molecular and stable isotope evidence for the occurrence of nitrite-dependent anaerobic methane-oxidizing bacteria in mangrove sediment of Zhangjiang estuary, China. Appl. Microbiol. Biotechnol. 2018, 102, 2441–2454. [Google Scholar] [CrossRef] [PubMed]
- Seitzinger, S.; Harrison, J.A.; Böhlke, J.K.; Bouwman, A.F.; Lowrance, R.; Peterson, B.; Tobias, C.; Van Drecht, G. Denitrification across landscapes and waterscapes: A synthesis. Ecol. Appl. 2006, 16, 2064–2090. [Google Scholar] [CrossRef]
- Kristensen, E.; Connolly, R.M.; Otero, X.L.; Marchand, C.; Ferreira, T.O.; Rivera-Monroy, V.H. Biogeochemical cycles: Global approaches and perspectives. In Mangrove Ecosystems: A Global Biogeographic Perspective; Rivera-Monroy, V.H., Lee, S.Y., Kristensen, E., Twilley, R.R., Eds.; Springer: Cham, Switzerland, 2017; pp. 163–209. [Google Scholar]
- Alongi, D.M.; Boto, K.G.; Robertson, A.I. Nitrogen and phosphorus cycles. In Tropical Mangrove Ecosystems; Robertson, A.I., Alongi, D.M., Eds.; American Geophysical Union: Washington, DC, USA, 1992; pp. 251–292. [Google Scholar]
- Singh, G.; Chauhan, R.; Kumar Ranjan, R.; Balakrishna Prasad, M.; Ramanathan, A.L. Phosphorus dynamics in mangrove of India. Curr. Sci. 2015, 108, 1874–1881. [Google Scholar]
- Feller, I.C.; Lovelock, C.E.; McKee, K.L. Nutrient addition differentially affects ecological processes of Avicennia germinans in nitrogen versus phosphorus limited mangrove ecosystems. Ecosystems 2007, 10, 347–359. [Google Scholar] [CrossRef]
- El-Tarabily, K.A.; Youssef, T. Enhancement of morphological, anatomical, and physiological characteristics of seedlings of the mangrove Avicennia marina inoculated with a native phosphate-solubilizing isolate of Oceanobacillus picturae under greenhouse conditions. Plant Soil 2010, 332, 147–162. [Google Scholar] [CrossRef]
- Scharler, U.M.; Ulanowicz, R.E.; Fogel, M.L.; Wooler, M.J.; Jacobson-Meyer, M.E.; Lovelock, C.E.; Feller, I.C.; Frischer, M.; Lee, R.; McKee, K.; et al. Variable nutrient stoichiometry (carbon: Nitrogen: Phosphorus) across trophic levels determines community and ecosystem properties in an oligotrophic mangrove system. Oecologia 2015, 179, 863–876. [Google Scholar] [CrossRef]
- Xie, X.; Weng, B.; Cai, B.; Dong, Y.; Yan, C. Effects of arbuscular mycorrhizal inoculation and phosphorus supply on the growth and nutrient uptake of Kandelia obovata (Sheue, Liu & Yong) seedlings in autoclaved soil. Appl. Soil Ecol. 2014, 75, 162–171. [Google Scholar]
- Das, S.; Jana, T.L.; De, T.K. Vertical profile of phosphatase activity in the Sundarbans mangrove forest, North East coast of Bay of Bengal, India. Geomicrobiol. J. 2014, 31, 716–725. [Google Scholar] [CrossRef]
- Behera, B.C.; Singdevsachan, S.K.; Mishra, R.R.; Sethi, B.K.; Dutta, S.K.; Thatoi, H.N. Phosphate solubilizing bacteria from mangrove soils of Mahanadi River Delta, Odisha, India. World J. Agric. Res. 2016, 4, 18–23. [Google Scholar]
- Behera, B.C.; Yadav, H.; Singh, S.K.; Mishra, R.R.; Sethi, B.K.; Dutta, S.K.; Thatoi, H.N. Phosphate solubilization and acid phosphatase activity of Serratia sp., isolated from mangrove soil of Mahanadi river delta, Odisha, India. J. Genet. Eng. Biotechnol. 2017, 15, 169–178. [Google Scholar] [CrossRef]
- Abhijith, R.; Vennila, A.; Purushothaman, C.S.; Padua, S.; Shilta, M.T.; Mahesh, V. Influence of sediment chemistry on mangrove-phosphobacterial relationship. Int. J. Chem. Stud. 2018, 6, 1677–1686. [Google Scholar]
- Silva, C.A.R.; Mozeto, A.A.; Ovalle, R.C. Distribution and fluxes as microdetritus of phosphorus in red mangroves, Sepetiba Bay, Brazil. Mangrove Salt Marshes 1998, 2, 37–42. [Google Scholar] [CrossRef]
- Ramos e Silva, C.A.; Oliveira, S.R.; Régo, R.D.P.; Mozeto, A.A. Dynamics of phosphorus and nitrogen through litter fall and decomposition in a tropical mangrove forest. Mar. Environ. Res. 2007, 64, 524–534. [Google Scholar] [CrossRef]
- Nielsen, T.; Andersen, F.Ø. Phosphorus dynamics during decomposition of mangrove (Rhizophora apiculata) leaves in sediments. J. Exp. Mar. Biol. Ecol. 2003, 293, 73–88. [Google Scholar] [CrossRef]
- Ray, R.; Majumder, N.; Das Chowdhury, S.; Jana, T.K. Phosphorus budget of the Sundarbans mangrove ecosystem: Box model approach. Estuar. Coast. 2018, 41, 1036–1049. [Google Scholar] [CrossRef]
- Breithaupt, J.L.; Smoak, J.M.; Smith, T.J., III; Sanders, C.J. Temporal variability of carbon and nutrient burial, sediment accretion, and mass accumulation over the past century in a carbonate platform mangrove forest of the Florida Everglades. J. Geophys. Res. Biogeosci. 2014, 119, 2032–2048. [Google Scholar] [CrossRef]
- Sanders, C.J.; Santos, I.R.; Maher, D.T.; Breithaupt, J.L.; Smoak, J.M.; Ketterer, M.; Call, M.; Sanders, L.; Eyre, B.D. Examing 239+240Pu, 210Pb and historical events to determine carbon, nitrogen and phosphorus burial in mangrove sediments of Moreton Bay, Australia. J. Environ. Radioact. 2016, 151, 623–629. [Google Scholar] [CrossRef] [PubMed]
- Breithaupt, J.L.; Smoak, J.M.; Sanders, C.J.; Troxler, T.G. Spatial variability of organic carbon, CaCO3 and nutrient burial rates spanning a mangrove productivity gradient in the coastal Everglades. Ecosystems 2019, 22, 844–858. [Google Scholar] [CrossRef]
- Pérez, A.; Machado, W.; Gutierrez, D.; Smoak, J.M.; Breithaupt, J.L.; Saldarriaga, M.S.; Sanders, L.; Marotta, H.; Sanders, C.J. Carbon and nutrient accumulation in mangrove sediments affected by multiple environmental changes. J. Soil. Sediments 2020, 20, 2504–2509. [Google Scholar] [CrossRef]
- Pérez, A.; Machado, W.; Gutierrez, D.; Saldarriaga, M.S.; Sanders, C.J. Shrimp farming influence on carbon and nutrient accumulation within Peruvian mangrove sediments. Estuar. Coast. Shelf Sci. 2020, 243, 106879. [Google Scholar] [CrossRef]
- Matos, C.R.L.; Berrêdo, J.F.; Machado, W.; Sanders, C.J.; Metzger, E.; Cohen, M.C.L. Carbon and nutrient accumulation in tropical mangrove creeks, Amazon region. Mar. Geol. 2020, 429, 106317. [Google Scholar] [CrossRef]
- Saderne, V.; Cusack, M.; Serrano, O.; Almahasheer, H.; Krishnakumar, P.K.; Rabaoui, L.; Qurban, M.A.; Duarte, C.M. Role of vegetated coastal ecosystems as nitrogen and phosphorus filters and sinks in the coasts of Saudi Arabia. Environ. Res. Lett. 2020, 15, 034058. [Google Scholar] [CrossRef]
- Alongi, D.M. Carbon cycling in the world’s mangrove ecosystems revisited: Significance of non-steady state diagenesis and subsurface linkages between the forest floor and the coastal ocean. Forests 2020, 11, 977. [Google Scholar] [CrossRef]
- Gong, W.-K.; Ong, J.-E. Plant biomass and nutrient flux in a managed mangrove forest in Malaysia. Estuar. Coast. Shelf Sci. 1990, 31, 519–530. [Google Scholar] [CrossRef]
- Alongi, D.M.; Wattayakorn, G.; Tirendi, F.; Dixon, P. Nutrient capital in different aged forests of the mangrove Rhizophora apiculata. Bot. Mar. 2004, 47, 116–124. [Google Scholar] [CrossRef]
- Burgin, A.J.; Yang, W.H.; Hamilton, S.K.; Silver, W.L. Beyond carbon and nitrogen: How the microbial energy economy couples elemental cycles in diverse ecosystems. Front. Ecol. Environ. 2011, 9, 44–52. [Google Scholar] [CrossRef]
- Pan, F.; Liu, H.; Guo, Z.; Li, Z.; Wang, B.; Cai, Y.; Gao, A. Effects of tide and season changes on the iron-sulfur-phosphorus biogeochemistry in sediment porewater of a mangrove coast. J. Hydrol. 2019, 568, 686–702. [Google Scholar] [CrossRef]
- Du, J.; Liu, J.; Lu, H.; Hansell, D.; Zhang, Q.; Wang, W.; Yan, C. Effect of external phosphate addition on solid-phase iron distribution and iron accumulation in mangrove Kandelia obovata (S.L.). Environ. Sci. Pollut. Res. 2015, 22, 13506–13513. [Google Scholar] [CrossRef]
- Dai, M.; Liu, J.; Liu, W.; Lu, H.; Jia, H.; Hong, H.; Yan, C. Phosphorus effects on radial oxygen loss, root porosity and iron plaque in two mangrove seedlings under cadmium stress. Mar. Pollut. Bull. 2017, 119, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, E.; Alongi, D.M. Control by fiddler crabs (Uca vocans) and plant roots (Avicennia marina) on carbon, iron, and sulfur biogeochemistry in mangrove sediment. Limnol. Oceanogr. 2006, 51, 1557–1571. [Google Scholar] [CrossRef]
- Hinokidani, K.; Nakanishi, Y. Dissolved iron elution from mangrove ecosystem associated with polyphenols and a herbivorous snail. Ecol. Evol. 2019, 9, 672–6784. [Google Scholar] [CrossRef]
- Attri, K.; Kerkar, S.; Loka Bharathi, P.A. Ambient iron concentration regulates the sulfur reducing activity in the mangrove swamps of Diwar, Goa, India. Estuar. Coast. Shelf Sci. 2011, 95, 156–164. [Google Scholar] [CrossRef]
- Arrivabene, H.P.; Campos, C.Q.; Souza, I.C.; Wunderlin, D.A.; Milanez, C.R.D.; Machado, S.R. Differential bioaccumulation and translocation patterns in three mangrove plants experimentally exposed to iron. Consequences for environmental sensing. Environ. Pollut. 2016, 215, 302–313. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, K.P.; Loka Bharathi, P.A. Organic carbon and iron modulate nitrification rates in mangrove swamps of Goa, south west coast of India. Estaur. Coast. Shelf Sci. 2009, 84, 419–426. [Google Scholar] [CrossRef]
- Fernandes, S.O.; Gonsalves, M.-J.; Michotey, V.D.; Bonin, P.C.; Loka Bharathi, P.A. Denitrification activity is closely linked to the total ambient Fe concentrations in mangrove sediments of Goa, India. Estuar. Coast. Shelf Sci. 2013, 131, 64–74. [Google Scholar] [CrossRef]
- Guan, Q.S.; Cao, W.Z.; Wu, G.J.; Wang, F.F.; Jiang, C.; Tao, Y.R.; Gao, Y. Nitrogen loss through anaerobic ammonium oxidation coupled with iron reduction in a mangrove wetland. Eur. J. Soil Sci. 2018, 69, 732–741. [Google Scholar] [CrossRef]
- Guan, Q.; Zhang, Y.; Tao, Y.; Chang, C.-T.; Cao, W. Graphene functions as a conductive bridge to promote anaerobic ammonium oxidation coupled with iron reduction in mangrove sediment slurries. Geoderma 2019, 352, 181–184. [Google Scholar] [CrossRef]
- Zheng, Y.; Hou, L.; Zhang, Z.; Chen, F.; Gao, D.; Yin, G.; Han, P.; Dong, H.; Liang, X.; Yang, Y. Anaerobic ammonium oxidation (anammox) bacterial diversity, abundance, and activity in sediments in the Indus Estuary. Estuar. Coast. Shelf Sci. 2020, 243, 106925. [Google Scholar] [CrossRef]
- Raven, M.R.; Fike, D.A.; Gomes, M.L.; Webb, S.M. Chemical and isotopic evidence for organic matter sulfurization in redox gradients around mangrove roots. Front. Earth Sci. 2019, 7, 98. [Google Scholar] [CrossRef]
- Crémiere, A.; Strauss, H.; Sebilo, M.; Hong, W.-L.; Gros, O.; Schmidt, S.; Tocny, J.; Henry, F.; Gontharet, S.; Lavenman, A.M. Sulfur diagenesis under rapid accumulation of organic-rich sediments in a marine mangrove from Guadeloupe (French West Indies). Chem. Geol. 2017, 454, 67–79. [Google Scholar] [CrossRef]
- Alongi, D.M. Dissolved iron supply limits early growth of estuarine mangroves. Ecology 2010, 91, 3229–3241. [Google Scholar] [CrossRef]
- Almahasheer, H.; Duarte, C.M.; Irigoien, X. Nutrient limitation in Central Red Sea mangroves. Front. Mar. Sci. 2016, 3, 271. [Google Scholar] [CrossRef]
- Cuadros, J.; Andrade, G.; Ferreira, T.O.; de Moya Partiti, C.S.; Cohen, R.; Vidal-Torrado, P. The mangrove reactor: Fast clay transformation and potassium sink. Appl. Clay Sci. 2017, 140, 50–58. [Google Scholar] [CrossRef]
- Mantiquilla, J.A.; Salmasan, S.F.D.; Obelidhon, M.K.A.; Abad, R.G. Nutrient status of Nipa (Nypa fruticans Wurmb.) in selected areas of Mindanao, the Philippines. Banwa B 2019, 14, art012. [Google Scholar]
- Jayasekera, R. Chemical composition of the mangrove, Rhizophora mangle L. J. Plant. Physiol. 1991, 138, 119–121. [Google Scholar] [CrossRef]
- Hossain, M.; Othman, S.; Bujang, J.S.; Kusman, M. Macronutrients content in different parts of seedling, sapling and tree of Bruguiera parviflora of Kuala Selangor Nature Park Mangrove Forest in Malaysia. Khulna Univ. Stud. 2003, 5, 15–20. [Google Scholar]
- Medina, E.; Fernandez, W.; Barboza, F. Element uptake, accumulation, and resorption in leaves of mangrove species with different mechanisms of salt regulation. Web Ecol. 2015, 15, 3–13. [Google Scholar] [CrossRef]
- Nasrin, S.; Hossain, M.; Rahman, M.M. Adaptive responses to salinity: Nutrient resorption efficiency of Sonneratia apetala (Buch.-Ham.) along the salinity gradient in the Sundarbans of Bangladesh. Wetl. Ecol. Manag. 2019, 27, 343–351. [Google Scholar] [CrossRef]
- Nandy, P.; Das, S.; Ghose, M.; Spooner-Hart, R. Effects of salinity on photosynthesis, leaf anatomy, ion accumulation and photosynthetic nitrogen use efficiency in five Indian mangroves. Wetl. Ecol. Manag. 2007, 15, 347–357. [Google Scholar] [CrossRef]
- Ball, M.C.; Chow, W.S.; Anderson, J.M. Salinity-induced potassium deficiency causes loss of functional photosystem II in leaves of the grey mangrove, Avicennia marina, through depletion of the atrazine-binding polypeptide. Aust J. Plant. Physiol. 1987, 14, 351–361. [Google Scholar] [CrossRef]
- Osman, H.E.; AboHassan, A.A. Effect of NPK fertilization on growth and dry matter accumulation in mangrove [Avicennia marina (Forssk) vierh] grown in western Saudi Arabia. Meteorol. Environ. Arid Land Agric. Sci. 2010, 21, 57–70. [Google Scholar] [CrossRef]
- Yates, E.J.; Ashwath, N.; Midmore, D.J. Responses to nitrogen, phosphorus, potassium and sodium chloride by three mangrove species in pot culture. Trees 2002, 16, 120–125. [Google Scholar] [CrossRef]
- Alongi, D.M.; Wattayakorn, G.; Boyle, S.; Tirendi, F.; Payn, C.; Dixon, P. Influence of roots and climate on mineral and trace element storage and flux in tropical mangrove soils. Biogeochemistry 2004, 69, 105–123. [Google Scholar] [CrossRef]
- Ray, R.; Mandal, S.K.; González, A.G.; Pokrovsky, O.S.; Jana, T.K. Storage and recycling of major and trace elements in mangroves. Sci. Total Environ. 2021, 780, 146379. [Google Scholar] [CrossRef]
- Sahebi, M.; Hanafi, M.M.; Akmar, A.S.N.; Rafi, M.Y.; Azizi, P.; Idris, A.S. Serine-rich protein is a novel positive regulator for silicon accumulation in mangrove. Gene 2015, 556, 170–181. [Google Scholar] [CrossRef]
- Zhang, Q.; Liu, J.; Lu, H.; Zhao, S.; Wang, W.; Du, J.; Yan, C. Effects of silicon on growth, root anatomy, radial oxygen loss (ROL) and Fe/N plaque of Aegiceras corniculatum (L.) Blanco seedlings exposed to cadmium. Environ. Nanotechnol. Monit. Manag. 2015, 4, 6–11. [Google Scholar] [CrossRef]
- Alongi, D.M.; Ramanathan, A.L.; Kannan, L.; Tirendi, F.; Trott, L.A.; Bala Krishna Prasad, M. Influence of human-induced disturbance on benthic microbial metabolism in the Pichavaram mangroves, Vellar-Coleroon estuarine complex, India. Mar. Biol. 2005, 147, 1033–1044. [Google Scholar] [CrossRef]
- Alongi, D.M.; Pfitzner, J.; Trott, L.A.; Tirendi, F.; Dixon, P.; Klumpp, D.W. Rapid sediment accumulation and microbial mineralization in forests of the mangrove Kandelia candel in the Jiulongjiang estuary, China. Estuar. Coast. Shelf Sci. 2005, 63, 605–618. [Google Scholar] [CrossRef]
- Pan, F.; Liu, H.; Guo, Z.; Cai, Y.; Fu, Y.; Wu, J.; Wang, B.; Gao, A. Metal/metalloid and phosphorus characteristics in porewater associated with manganese geochemistry: A case study in the Jiulong River estuary, China. Environ. Pollut. 2019, 255, 113134. [Google Scholar] [CrossRef]
- Holloway, C.J.; Santos, I.R.; Tait, D.R.; Sanders, C.J.; Rose, A.L.; Schnetger, B.; Brumsack, H.-J.; Macklin, P.A.; Sippo, J.Z.; Maher, D.T. Manganese and iron release from mangrove porewaters: A significant component of oceanic budgets? Mar. Chem. 2016, 184, 43–52. [Google Scholar] [CrossRef]
- Shackira, A.M.; Puthur, J.T.; Salim, E.N. Acanthus ilicifolius L. a promising candidate for phytostabilization of zinc. Environ. Monit. Assess. 2017, 189, 282. [Google Scholar] [CrossRef]
- Hossain, M.; Othman, S.; Kusnan, M.; Bujang, J.S. Biological cycling of micro-nutrients (Cu, Fe and Zn) in Bruguiera parviflora dominated mangrove forest at Kuala Selangor Nature Park, Malaysia. Malays. Appl. Biol. 2008, 37, 63–68. [Google Scholar]
- De Araújo, J.M.C., Jr.; Ferreira, T.O.; Suarez-Abelenda, M.; Nόbrega, G.N.; Albuquerque, A.G.B.M.; de Bezerra, A.C.; Otero, X.L. The role of bioturbation by Ucides cordatus crab in the fractionation and bioavailability of trace metals in tropical semiarid mangroves. Mar. Pollut. 2016, 111, 194–202. [Google Scholar] [CrossRef]
- Jerlin, B.; Sharmila, S.; Kathiresan, K.; Kayalvizhi, K. Zinc solubilizing bacteria from rhizospheric soil of mangroves. Int. J. Microbiol. Biotechnol. 2017, 2, 148–155. [Google Scholar]
- MacFarlane, G.R.; Burchett, M.D. Toxicity, growth and accumulation relationships of copper, lead and zinc in the grey mangrove Avicennia marina (Forsk.) Vierh. Mar. Environ. Res. 2002, 54, 65–84. [Google Scholar] [CrossRef]
- Alongi, D.M. Micronutrients and mangroves: Experimental evidence for copper limitation. Limnol. Oceanogr. 2017, 62, 2759–2772. [Google Scholar] [CrossRef]
- Rahaman, W.; Singh, S.K.; Raghav, S. Dissolved Mo and U in rivers and estuaries of India: Implication to geochemistry of redox sensitive elements and their marine budgets. Chem. Geol. 2010, 278, 160–172. [Google Scholar] [CrossRef]
- Ravizza, G.; Turekian, K.K.; Hay, B.J. The geochemistry of rhenium and osmium in recent sediments from the Black Sea. Geochim. Cosmochim. Acta 1991, 55, 3741–3752. [Google Scholar] [CrossRef]
- Rahaman, W.; Goswami, V.; Singh, S.K.; Rai, V.K. Molybdenum isotopes in two Indian estuaries: Mixing characteristics and input to oceans. Geochim. Cosmochim. Acta 2014, 141, 407–422. [Google Scholar] [CrossRef]
- McManus, J.; Berelson, W.M.; Severmann, S.; Poulson, R.L.; Hammond, D.E.; Klinkhammer, G.P.; Holm, C. Molybdenium and uranium geochemistry in continental margin sediments: Paleoproxy potential. Geochim. Cosmochim. Acta 2006, 70, 4643–4662. [Google Scholar] [CrossRef]
- Wong, M.Y.; Mahowald, N.M.; Marino, R.; Williams, E.R.; Chellam, S.; Howarth, R.W. Natural atmospheric deposition of molybdenium: A global model and implications for tropical forests. Biogeochemistry 2020, 149, 159–174. [Google Scholar] [CrossRef]
- Noël, V.; Morin, G.; Juillot, F.; Marchand, C.; Brest, J.; Bargar, J.R.; Muñoz, M.; Marakovic, G.; Ardo, S.; Brown, G.E., Jr. Ni cycling in mangrove sediments in New Caledonia. Geochim. Cosmochim. Acta 2015, 169, 82–98. [Google Scholar] [CrossRef]
- Marchand, C.; Fernandez, J.-M.; Moreton, B. Trace metal geochemistry in mangrove sediments and their transfer to mangrove plants (New Caledonia). Sci. Total Environ. 2016, 562, 216–227. [Google Scholar] [CrossRef]
- Suzuki, K.N.; Machado, E.C.; Machado, W.; Bellido, A.V.; Bellido, L.F.; Osso, J.A., Jr.; Lopes, R.T. Selenium, chromium and cobalt diffusion into mangrove sediments: Radiotracer experiment evidence of coupled effects of bioturbation and rhizosphere. Water Air Soil Pollut. 2012, 223, 3887–3892. [Google Scholar] [CrossRef]
- Almahasheer, H.; Serrano, O.; Duarte, C.M.; Irigoien, X. Remobilization of heavy metals by mangrove leaves. Front. Mar. Sci. 2018, 5, 484. [Google Scholar] [CrossRef]
- Thanh-Nho, N.; Marchand, C.; Strady, E.; Huu-Phat, N.; Nhu-Trang, T.-T. Bioaccumulation of some trace elements in tropical mangrove plants and snails (Can Gio, Vietnam). Environ. Pollut. 2019, 248, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Schlesinger, W.H.; Bernhardt, E.S. Biogeochemistry: An Analysis of Global Change; Academic Press: London, UK, 2020. [Google Scholar]
- Reis, C.R.G.; Nardoto, G.B.; Oliveira, R.S. Global overview on nitrogen dynamics in mangroves and consequences of increasing nitrogen availability for these systems. Plant Soil 2017, 410, 1–19. [Google Scholar] [CrossRef]
- Alongi, D.M. Impact of global change on nutrient dynamics in mangrove forests. Forests 2018, 9, 596. [Google Scholar] [CrossRef]
- Thanh-Nho, N.; Marchand, C.; Strady, E.; Van Vinh, T.; Taillardat, P.; Cong-Hau, N.; Nhu-Trang, T.-T. Trace metal dynamics in a tropical mangrove tidal creek: Influence of porewater seepage (Can Gio, Vietnam). Front. Environ. Sci. 2020, 8, 139. [Google Scholar] [CrossRef]
- Sadat-Noori, M.; Glamore, W. Porewater exchange drives trace metal, dissolved organic carbon and total dissolved nitrogen export from a temperate mangrove wetland. J. Environ. Manag. 2019, 248, 109264. [Google Scholar] [CrossRef]
- Morel, F.M.M.; Hudson, R.J.M.; Price, N.M. Limitation of productivity by trace metals in the sea. Limnol. Oceanogr. 1991, 36, 1742–1755. [Google Scholar] [CrossRef]
- Zhang, J. Evidence of trace metal limited photosynthesis in eutrophic estuarine and coastal waters. Limnol. Oceanogr. 2000, 45, 1871–1878. [Google Scholar] [CrossRef]
- Sunda, W.G. Feedback interactions between trace metal nutrients and phytoplankton in the ocean. Front. Microbiol. 2012, 3, 204. [Google Scholar] [CrossRef]
- Paerl, H.W.; Crocker, K.M.; Prufert, L.E. Limitation of N2 fixation in coastal marine waters: Relative importance of molybdenum, iron, phosphorus, and organic matter availability. Limnol. Oceanogr. 1987, 32, 525–536. [Google Scholar] [CrossRef]
- Holmer, M.; Duarte, C.M.; Marbá, N. Iron additions reduce sulfate reduction rates and improve seagrass growth on organic-enriched carbonate sediments. Ecosystems 2005, 8, 721–730. [Google Scholar] [CrossRef]
- Brodersen, K.E.; Koren, K.; Mosshammer, M.; Ralph, P.J.; Kühl, M.; Santner, J. Seagrass-mediated phosphorus and iron solubilization in tropical sediments. Environ. Sci. Technol. 2017, 51, 14155–14163. [Google Scholar] [CrossRef]
- Ruiz-Helpern, S.; Macko, S.A.; Fourqurean, J.W. The effects of manipulation of sedimentary iron and organic matter on sediment biogeochemistry and seagrasses in a subtropical carbonate environment. Biogeochemistry 2008, 87, 113–126. [Google Scholar] [CrossRef]
- Van Langenhove, L.; Verryckt, L.; Bréchet, L.; Courtois, E.A.; Stahl, C.; Hofhansl, F.; Bauters, M.; Sardans, J.; Boeckx, P.; Fransen, E.; et al. Atmospheric deposition of elements and its relevance for nutrient budgets of tropical forests. Biogeochemistry 2020, 149, 175–193. [Google Scholar] [CrossRef]
- Barkley, A.E.; Prospero, J.M.; Mahowald, N.; Hamilton, D.S.; Popendorf, K.J.; Oehlert, A.M.; Pourmand, A.; Gatineau, A.; Panechou-Pulcherie, K.; Blackwelder, P.; et al. African biomass burning is a substantial source of phosphorus deposition to the Amazon, tropical Atlantic Ocean, and Southern Ocean. Proc. Nat. Acad. Sci. USA 2019, 116, 16216–16221. [Google Scholar] [CrossRef]
- Strzelec, M.; Proemse, B.C.; Gault-Ringold, M.; Boyd, P.W.; Perron, M.M.G.; Schofield, R.; Ryan, R.G.; Ristovski, Z.D.; Alroe, J.; Humphries, R.S.; et al. Atmospheric trace metal deposition near the Great Barrier Reef, Australia. Atmosphere 2020, 11, 390. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transformation Process | Mean ± 1 SE (Median) | Range |
|---|---|---|
| Denitrification | 26.3 ± 3.3 (3.9) | 0–443.5 |
| Gross ammonification | 301.6 ± 50.9 (267.4) | 77.4–898.8 |
| Net ammonification | 31.4 ± 6.2 (15.4) | 0.17–200.0 |
| Gross nitrification | 15.1 ± 5.81 (4.7) | 0–141.0 |
| Net nitrification | 5.8 ± 1.2 (1.9) | 0–30.8 |
| Anammox | 22.1 ± 5.5 (4.4) | 0–99.4 |
| DNRA | 18.2 ± 6.8 (4.5) | 0–108.6 |
| N2-fixation (soil) | 8.2 ± 1.7 (3.2) | 0–58.9 |
| N2-fixation (microbial mats) | 9.7 ± 2.6 (3.4) | 0–60.4 |
| N2-fixation (AB roots) | 31.8 ± 5.6 (26.3) | 2.8–73.4 |
| N2-fixation (BG roots) | 6.2 ± 1.7 (4.5) | 1–26.2 |
| N2-fixation (litter) | 1.2 ± 0.4 (0.5) | 0.2–3.3 |
| N2- fixation (dead leaves) | 0.8 ± 0.1 (0.8) | 0.4–1.2 |
| N2-fixation (stem bark) | 101.0 ± 36.4 (101.0) | 17.4–201.2 |
| Fluxes | Mean ± 1 SE (Median) | Range |
|---|---|---|
| N2O soil-air | 0.60 ± 0.17 (0.22) | −11.04 |
| DON soil-water | −18.29 ± 18.14 (0.00) | −743–19.6 |
| NO2− + NO3− soil-water | −4.46 ± 1.05 (−0.24) | −29.04–3.36 |
| NH4+ soil-water | −1.48 ± 2.20 (−0.16) | −88.7–55.19 |
| NH4+ import | −9.81 ± 4.35 (−1.57) | −100.76–−0.006 |
| NH4+ export | 8.00 ± 3.63 (0.39) | 0.03–62.40 |
| NH4+ net exchange | −1.45 ± 3.12 (−0.01) | −100.76–62.4 |
| DON import | −4.69 ± 2.17 (−2.87) | −21.84–−0.91 |
| DON export | 1.45 ± 0.33 (1.37) | 0.08–3.27 |
| DON net exchange | −1.46 ± 1.24 (0.08) | −21.84–3.27 |
| NO2− + NO3− import | −3.62 ± 1.52 (−0.73) | −28.80–−0.003 |
| NO2− + NO3− export | 1.84 ± 0.55 (0.29) | 0.05–11.90 |
| NO2− + NO3− net exchange | −0.73 ± 0.85 (0.08) | −28.8–11.90 |
| Component | Rhizophora stylosa | Rhizophora apiculata | Avicennia marina | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Mg | Ca | K | Mg | Ca | K | Mg | Ca | K | |
| Leaves | 13.8 | 9.4 | 6.3 | 7.3 | 19.6 | 8.2 | 11 | 5.9 | 8.8 |
| Branches | 16.5 | 1.2 | 1.1 | 1.2 | 14.5 | 2.6 | 1 | 9.2 | 2.7 |
| Stems | 16.6 | 1.3 | 1 | 0.5 | 4.7 | 0.8 | 1.2 | 7.1 | 2.8 |
| Prop roots | 7.6 | 9 | 3.1 | 0.9 | 5.4 | 1.2 | NA | ||
| BG live roots | 7.9 | 8.4 | 2.4 | 4.8 | 3.4 | 1.8 | 7 | 14.6 | 3.7 |
| BG dead roots | 11.6 | 60.8 | 4.8 | 6.5 | 7.5 | 1.2 | 6.5 | 14.9 | 3.9 |
| Soil (kg m−2) | 7.4 | 38.4 | 3 | 3.2 | 1.2 | 3.5 | 10 | 34.4 | 4.4 |
| Component | 25-Year-Old Forest | 5-Year-Old Forest | 3-Year-Old Forest | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Mg | Ca | K | Mg | Ca | K | Mg | Ca | K | |
| Inventory | 1.68 | 0.46 | 2.17 | 2.2 | 2.22 | 2.99 | 3.19 | 1.17 | 3.48 |
| Release | 0.29 | 0.25 | 0.1 | 0.03 | 0.06 | 0.02 | 0.04 | 0.15 | 0.05 |
| Turnover | 9.2 | 1.9 | 21.1 | 85 | 39.6 | 126.9 | 81.3 | 7.8 | 63.8 |
| AG NPP | 5.3 | 1.7 | 3.7 | ||||||
| Component | 25-Year-Old Forest | 5-Year-Old Forest | 3-Year-Old Forest | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mn | Cu | Zn | Mo | Mn | Cu | Zn | Mo | Mn | Cu | Zn | Mo | |
| Inventory | 89.5 | 3.9 | 12.7 | 1.6 | 230.5 | 3 | 19.2 | 3.1 | 120.5 | 2.8 | 19.8 | 4.1 |
| Release | 33.4 | 0.006 | 0.08 | 0.06 | 1.9 | 0.006 | 0.03 | 0.01 | 8.8 | 0.02 | 0.04 | 0.01 |
| Turnover | 3 | 610 | 162 | 28 | 120 | 470 | 733 | 320 | 14 | 147 | 505 | 430 |
| AG NPP | 5.3 | 1.7 | 3.7 | |||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alongi, D.M. Macro- and Micronutrient Cycling and Crucial Linkages to Geochemical Processes in Mangrove Ecosystems. J. Mar. Sci. Eng. 2021, 9, 456. https://doi.org/10.3390/jmse9050456
Alongi DM. Macro- and Micronutrient Cycling and Crucial Linkages to Geochemical Processes in Mangrove Ecosystems. Journal of Marine Science and Engineering. 2021; 9(5):456. https://doi.org/10.3390/jmse9050456
Chicago/Turabian StyleAlongi, Daniel M. 2021. "Macro- and Micronutrient Cycling and Crucial Linkages to Geochemical Processes in Mangrove Ecosystems" Journal of Marine Science and Engineering 9, no. 5: 456. https://doi.org/10.3390/jmse9050456
APA StyleAlongi, D. M. (2021). Macro- and Micronutrient Cycling and Crucial Linkages to Geochemical Processes in Mangrove Ecosystems. Journal of Marine Science and Engineering, 9(5), 456. https://doi.org/10.3390/jmse9050456