Anatomy and Ultrastructure of the Cyprid Temporary Adhesive System in Two Species of Acorn Barnacle




Abstract
1. Introduction
2. Materials and Methods
2.1. Organism Acquisition
2.2. Sample Preparation
2.3. Serial Block Face-Scanning Electron Microscopy
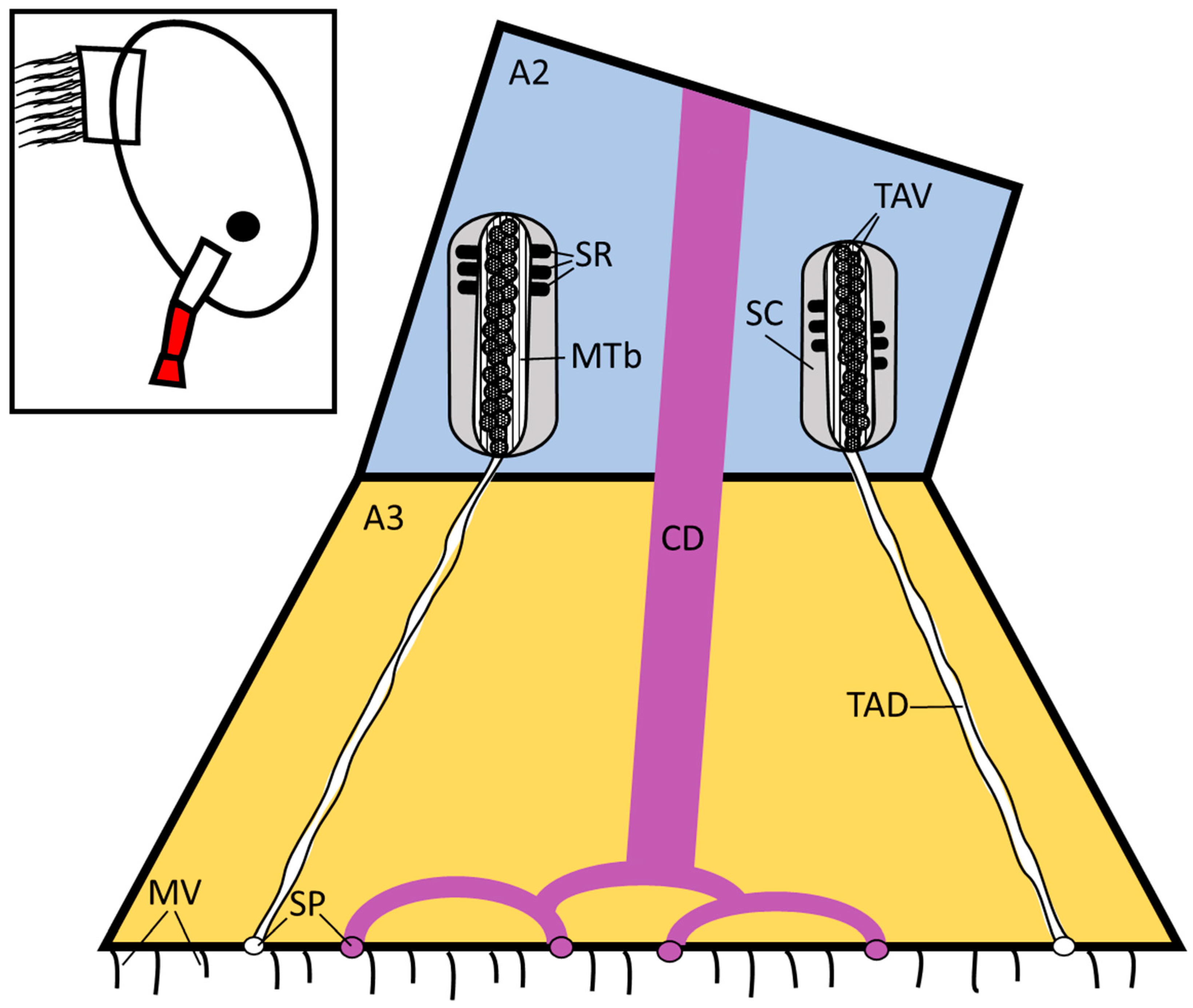
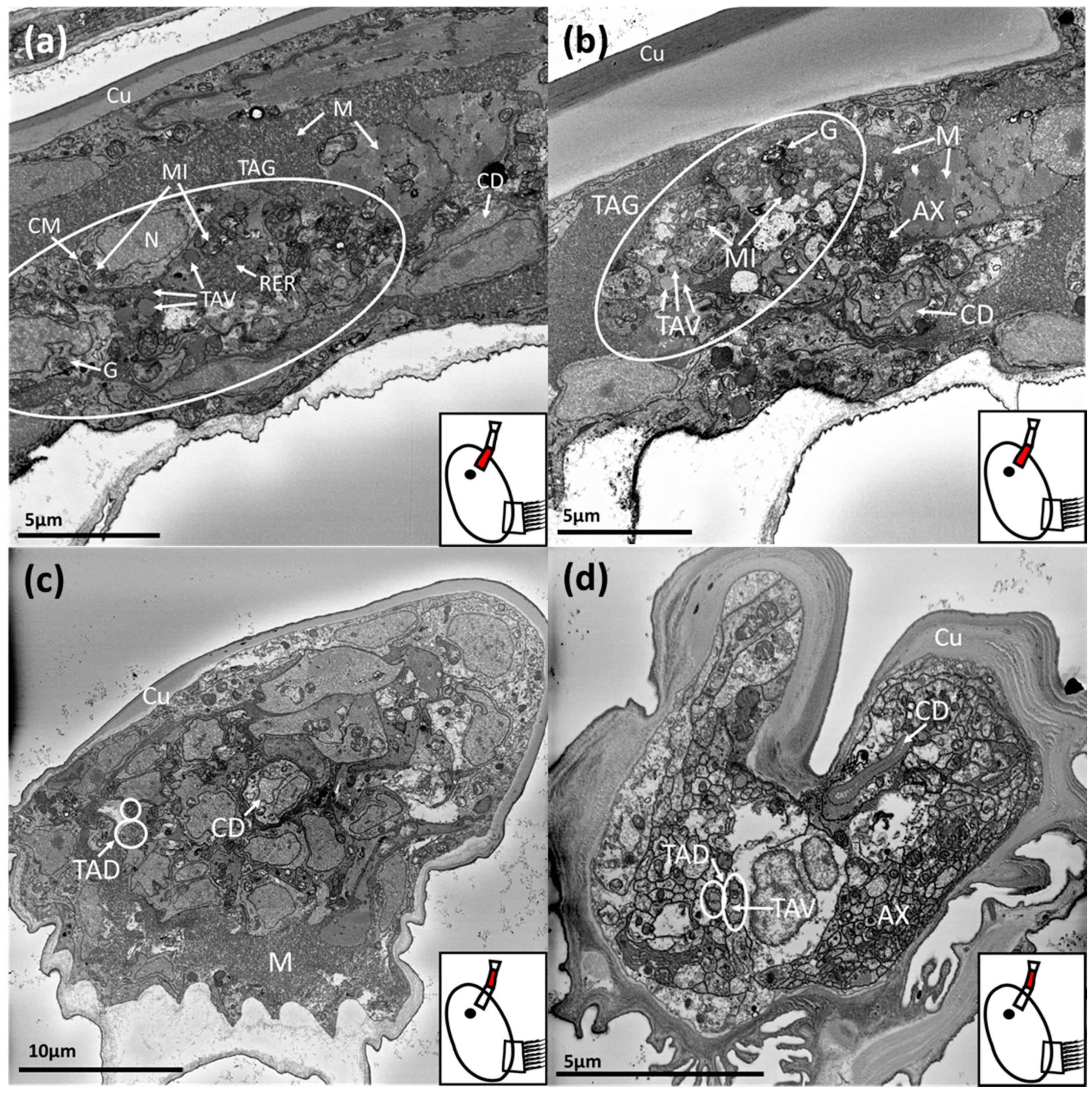
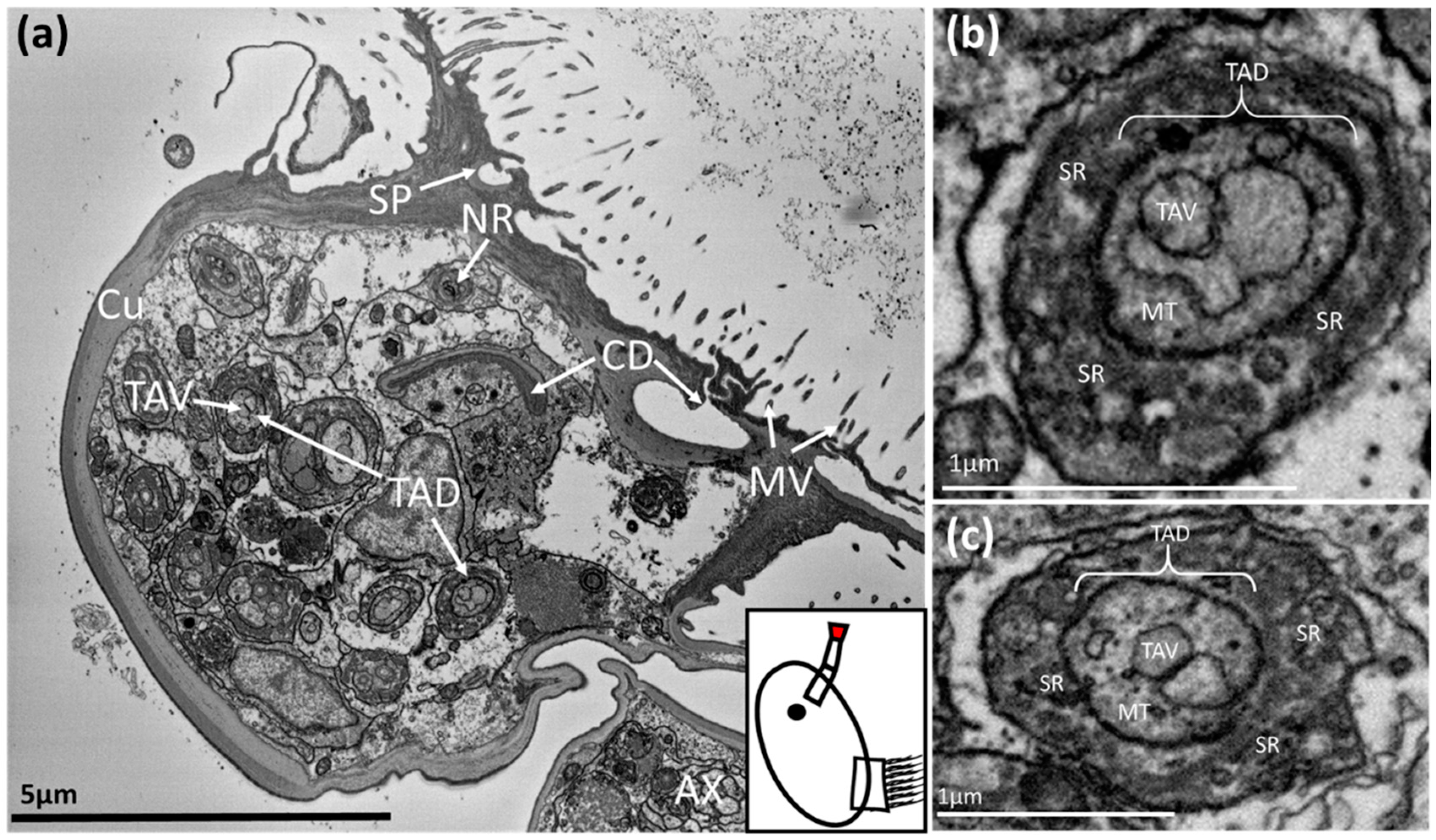
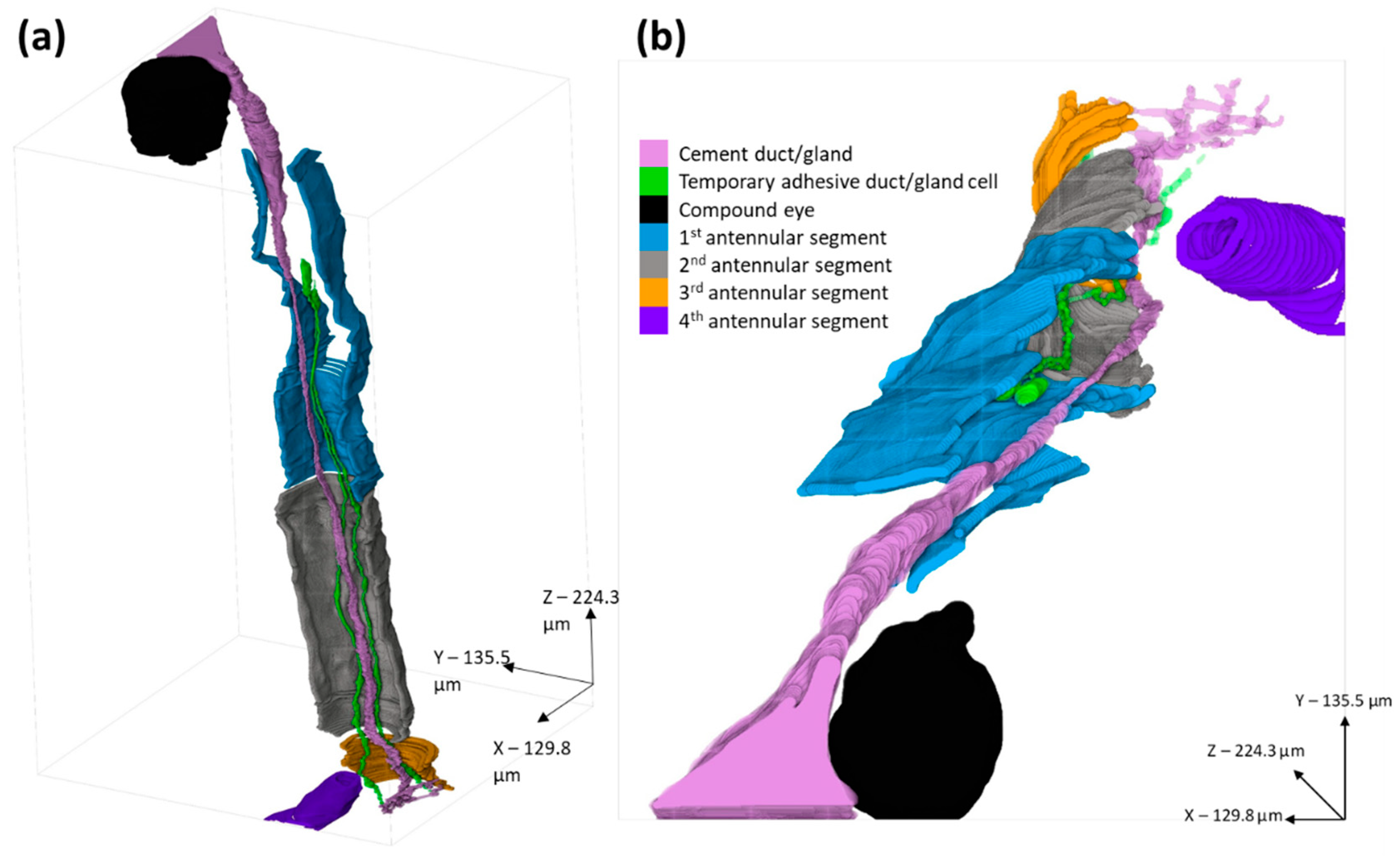
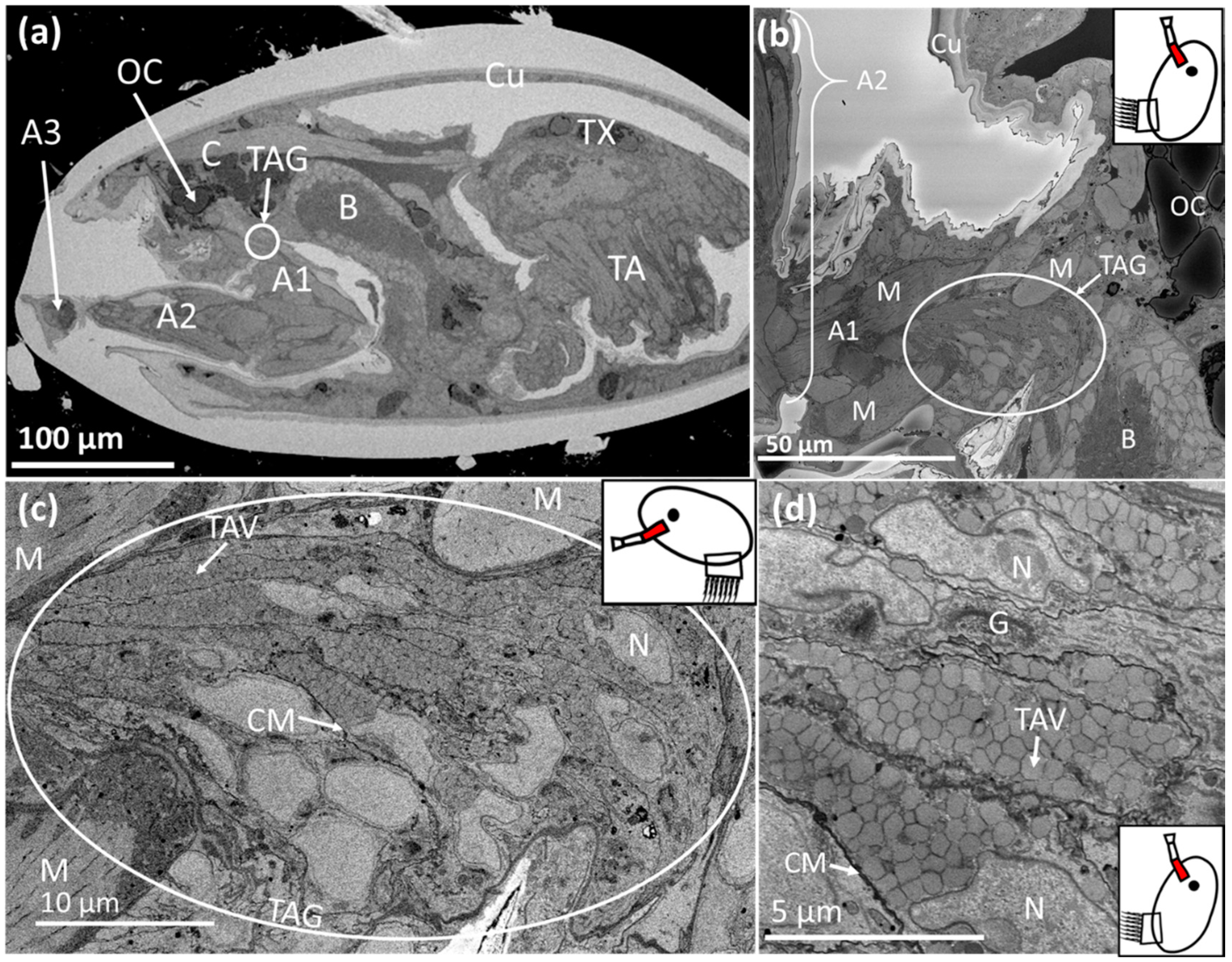
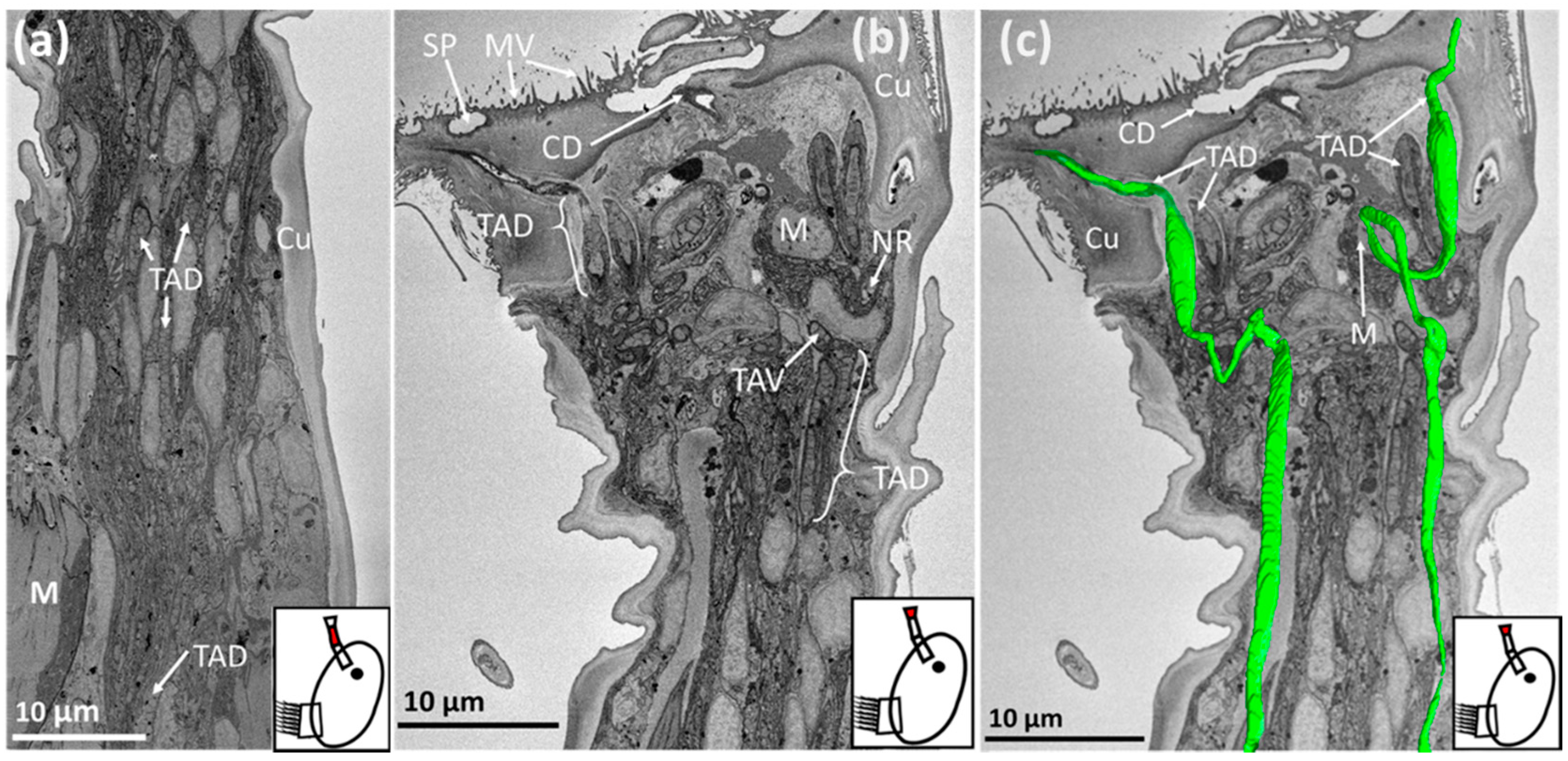
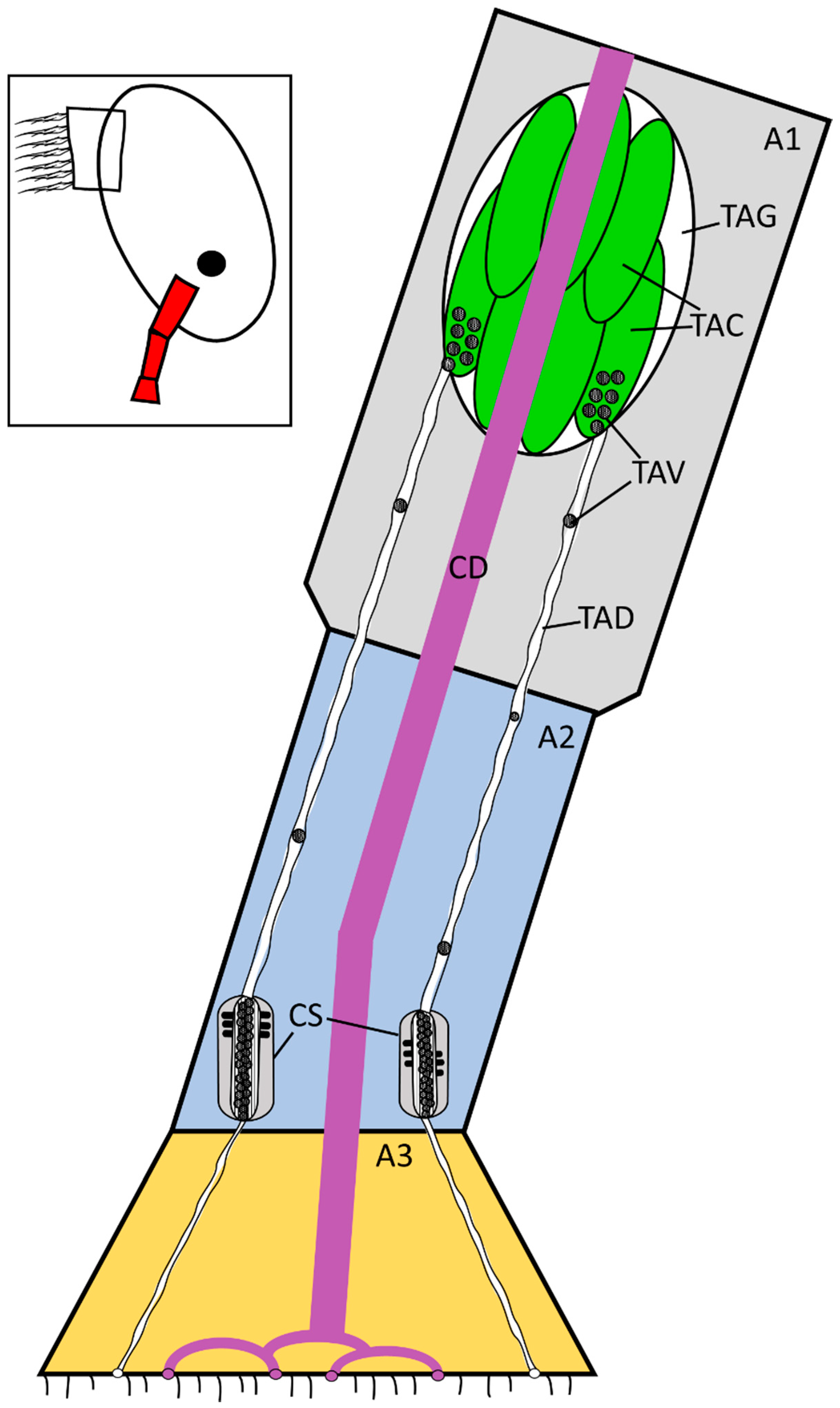
3. Results
3.1. Balanus amphitrite
3.2. Megabalanus coccopoma
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Crisp, D.J. Settlement responses in marine organisms. In Adaptations to Environment: Essays on the Physiology of Marine Animals; Newell, R.C., Ed.; Butterworths: London, UK, 1976; pp. 83–124. [Google Scholar]
- Walker, G.; Yule, A. Temporary adhesion of the barnacle cyprid: The existence of an antennular adhesive secretion. J. Mar. Biol. Assoc. UK 1984, 64, 679–686. [Google Scholar] [CrossRef]
- Dreanno, C.; Kirby, R.R.; Clare, A.S. Smelly feet are not always a bad thing: The relationship between cyprid footprint protein and the barnacle settlement pheromone. Biol. Lett. 2006, 2, 423–425. [Google Scholar] [CrossRef] [PubMed]
- Phang, I.Y.; Aldred, N.; Clare, A.S.; Vancso, G.J. Towards a nanomechanical basis for temporary adhesion in barnacle cyprids (Semibalanus balanoides). J. R. Soc. Interface 2008, 5, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Phang, I.Y.; Aldred, N.; Ling, X.Y.; Huskens, J.; Clare, A.S.; Vancso, G.J. Atomic force microscopy of the morphology and mechanical behaviour of barnacle cyprid footprint proteins at the nanoscale. J. R. Soc. Interface 2010, 7, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Aldred, N.; Høeg, J.T.; Maruzzo, D.; Clare, A.S. Analysis of the behaviours mediating barnacle cyprid reversible adhesion. PLoS ONE 2013, 8, e68085. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Puniredd, S.R.; Jańczewski, D.; Lee, S.S.C.; Teo, S.L.M.; He, T.; Zhu, X.; Vancso, G.J. Barnacle larvae exploring surfaces with variable hydrophilicity: Influence of morphology and adhesion of “footprint” proteins by AFM. ACS Appl. Mater. Interfaces 2014, 6, 13667–13676. [Google Scholar] [CrossRef] [PubMed]
- Walker, G. A study of the cement apparatus of the cypris larva of the barnacle Balanus balanoides. Mar. Biol. 1971, 9, 205–212. [Google Scholar] [CrossRef]
- Okano, K.E.; Shimizu, K.A.; Satuito, C.; Fusetani, N.O. Visualization of cement exocytosis in the cypris cement gland of the barnacle Megabalanus rosa. J. Exp. Biol. 1996, 199, 2131–2137. [Google Scholar] [PubMed]
- Ödling, K.; Albertsson, C.; Russell, J.T.; Mårtensson, L.G. An in vivo study of exocytosis of cement proteins from barnacle Balanus improvisus (D.) cyprid larva. J. Exp. Biol. 2006, 209, 956–964. [Google Scholar] [CrossRef] [PubMed]
- Nott, J.A.; Foster, B.A. On the structure of the antennular attachment organ of the cypris larvae of Balanus balanoides (L.). Philos. Trans. R. Soc. B 1969, 256, 115–134. [Google Scholar]
- Yap, F.C.; Wong, W.L.; Maule, A.G.; Brennan, G.P.; Chong, V.C.; Lim, L.H.S. First evidence for temporary and permanent adhesive systems in the stalked barnacle cyprid, Octolasmis angulata. Sci. Rep. 2017, 7, 44980. [Google Scholar] [CrossRef] [PubMed]
- Belevich, I.; Joensuu, M.; Kumar, D.; Vihinen, H.; Jokitalo, E. Microscopy image browser: A platform for segmentation and analysis of multidimensional datasets. PLoS Biol. 2016, 14, e1002340. [Google Scholar] [CrossRef] [PubMed]
- Deerinck, T.J.; Bushong, E.A.; Lev-Ram, V.; Shu, X.; Tsien, R.Y.; Ellisman, M.H. Enhancing serial block-face scanning electron microscopy to enable high resolution 3-D nanohistology of cells and tissues. Microsc. Microanal. 2010, 16, 1138–1139. [Google Scholar] [CrossRef]
- Noirot, C.; Quennedey, A. Fine structure of insect epidermal glands. Annu. Rev. Entomol. 1974, 19, 61–80. [Google Scholar] [CrossRef]
- Talbot, P.; Demers, D. Tegumental glands of Crustacea. In Crustacean Integument. Morphology and Biochemistry Boca Raton; Horst, M.N., Freeman, J.A., Eds.; CRC Press: Boca Raton, FL, USA, 1993; pp. 151–191. [Google Scholar]
- Quennedey, A. Insect epidermal gland cells: Ultrastructure and morphogenesis. In Microscopic Anatomy of Invertebrates; Harrison, F., Locke, M., Eds.; Wiley-Liss: Hoboken, NJ, USA, 1998; pp. 177–207. [Google Scholar]
- Freeman, S.C.; Malik, A.; Basit, H. Physiology, Exocrine Gland; StatPearls Publishing: Treasure Island, FL, USA, 2020; StatPearls [Internet]; (Updated 24 August 2020). Available online: https://www.ncbi.nlm.nih.gov/books/NBK542322/ (accessed on 25 November 2020).
- Lagersson, N.; Høeg, J. Settlement behavior and antennulary biomechanics in cypris larvae of Balanus amphitrite (Crustacea: Thecostraca: Cirripedia). Mar. Biol. 2002, 141, 513–526. [Google Scholar]
- Pérez-Losada, M.; Høeg, J.T.; Crandall, K.A. Unravelling the evolutionary radiation of the thoracican barnacles using molecular and morphological evidence: A comparison of several divergence time estimation approaches. Syst. Biol. 2004, 53, 278–298. [Google Scholar] [CrossRef] [PubMed]
- Ewers-Saucedo, C.; Owen, C.L.; Pérez-Losada, M.; Høeg, J.T.; Glenner, H.; Chan, B.; Crandall, K.A. Towards a barnacle tree of life: Integrating diverse phylogenetic efforts into a comprehensive hypothesis of thecostracan evolution. Peer J. 2019, 7, e7387. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Magnification | Pixel Time | Section Thickness | Pixel Resolution |
|---|---|---|---|---|
| Balanus amphitrite | Series: 1290–1880× Stills: 898–4000× as required | Series: 20 µs Stills: 20 µs | Series: 100 nm Stills: 100 nm | Series: 20 nm, 2000 × 2000–3500 × 3500 Stills: 10 nm 4000 × 4000 |
| Megabalanus coccopoma | Series: 646× Stills: 157–6700× as required | Series: 20 µs Stills: 20 µs | Series: 100 nm Stills: 100 nm | Series: 25 nm, 3000 × 3000 Stills: 5.6 nm–478.5 nm, 2000 × 2000 |
| Species | TAV Diameter | TAD Diameter | TAG Cells | TAG | Cyprid Length | A1 Length | A2 Length | A3 Diameter |
|---|---|---|---|---|---|---|---|---|
| Balanus amphitrite | ~250 nm (±89) | ~0.5 µm (±0.11) | ~4.5 × 4.5 × 10 µm | ~10 × 10 × 40 µm | ~400 µm | ~90 µm | ~80 µm | ~20 µm |
| Megabalanus coccopoma | ~400 nm (±104) | ~0.8 µm (±0.26) | ~5 × 5 × 25 µm | ~25 × 25 × 50 µm | ~500 µm | ~150 µm | ~140 µm | ~35 µm |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raine, J.J.; Aldred, N.; Clare, A.S. Anatomy and Ultrastructure of the Cyprid Temporary Adhesive System in Two Species of Acorn Barnacle. J. Mar. Sci. Eng. 2020, 8, 968. https://doi.org/10.3390/jmse8120968
Raine JJ, Aldred N, Clare AS. Anatomy and Ultrastructure of the Cyprid Temporary Adhesive System in Two Species of Acorn Barnacle. Journal of Marine Science and Engineering. 2020; 8(12):968. https://doi.org/10.3390/jmse8120968
Chicago/Turabian StyleRaine, Joshua J., Nick Aldred, and Anthony S. Clare. 2020. "Anatomy and Ultrastructure of the Cyprid Temporary Adhesive System in Two Species of Acorn Barnacle" Journal of Marine Science and Engineering 8, no. 12: 968. https://doi.org/10.3390/jmse8120968
APA StyleRaine, J. J., Aldred, N., & Clare, A. S. (2020). Anatomy and Ultrastructure of the Cyprid Temporary Adhesive System in Two Species of Acorn Barnacle. Journal of Marine Science and Engineering, 8(12), 968. https://doi.org/10.3390/jmse8120968