Multidisciplinary Analysis of Cystoseira sensu lato (SE Spain) Suggest a Complex Colonization of the Mediterranean



 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Cartography of Cystoseira Sensu Lato
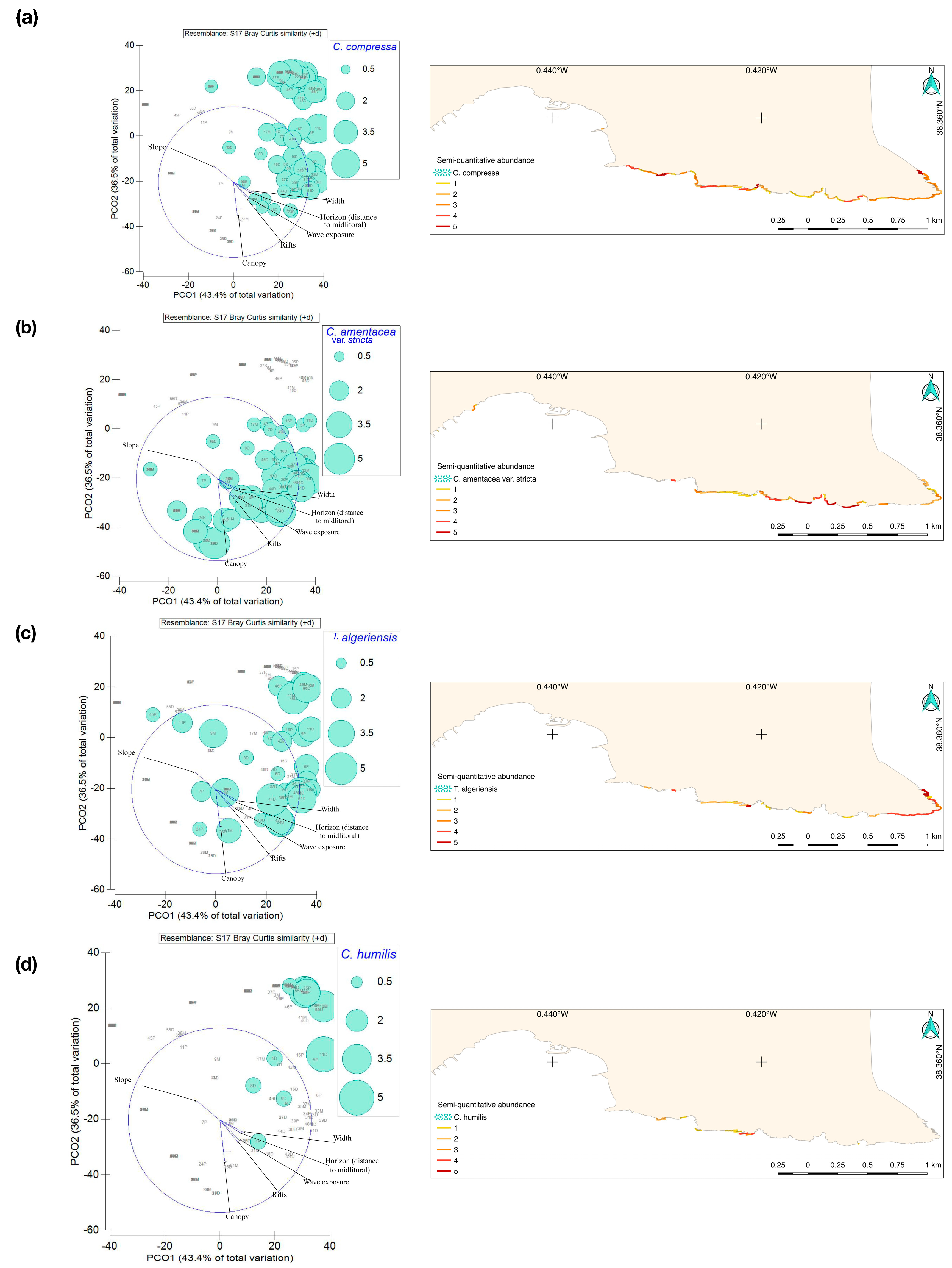
2.2. Abiotic Factors and Spatial Variability of Cystoseira Spp.
2.3. Morphological Characterization
2.4. DNA Extraction
2.5. Gene Amplification, Sequencing and Phylogenetic Analysis
3. Results
3.1. Cartography of Communities Associated with the Coastal Fringe
3.2. Distribution of Cystoseira Sensu Lato
3.3. Morphological Analysis
3.4. Phylogenetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cánovas, F.; Mota, C.; Serrão, E.; Pearson, G. Driving south: A multi-gene phylogeny of the brown algal family Fucaceae reveals relationships and recent drivers of a marine radiation. Evol. Biol. 2011, 11, 371. [Google Scholar] [CrossRef] [PubMed]
- Draisma, S.; Ballesteros, E.; Florence, R.; Thibaut, T. DNA Sequence Data Demonstrate the Polyphily of the Genus Cystoseira and Other Sargassaceae Genera (Phaeophyceae). J. Phycol. 2010, 46, 1329–1345. [Google Scholar] [CrossRef]
- García-Fernández, A.; Bárbara, I. Studies of Cystoseira assemblages in Northern Atlantic Iberia. An. Jard. Bot. Madr. 2016, 73, e0352016. [Google Scholar] [CrossRef]
- Guiry, M.D.; Guiry, G.M.; World-Wide Electronic Publication, National University of Ireland, Galway. AlgaeBase. Available online: https://www.algaebase.org (accessed on 6 June 2020).
- Thibaut, T.; Blanfuné, A.; Markovic, L.; Verlaque, M.; Boudouresque, C.F.; Perret-Boudouresque, M.; Macic, V.; Bottin, L. Unexpected abundance and long- term relative stability of the brown alga Cystoseira amentacea, hitherto regarded as a threatened species, in the north-western Mediterranean Sea. Mar. Pollut. Bull. 2014, 89, 305–323. [Google Scholar] [CrossRef]
- Hsü, K.J. The Mediterranean Was a Desert: A Voyage of the Glomar Challenger; Princeton University Press: Princeton, NJ, USA, 1982; p. 197. ISBN -10. [Google Scholar]
- Boudouresque, C. Marine biodiversity in the mediterranean status of species populations and communities. Sci. Rep. Port Cros Nat. Park 2004, 20, 97–146. [Google Scholar]
- Bermejo, R.; Chefaoui, R.M.; Engelen, A.H.; Buonomo, R.; Neiva, J.; Ferreira-Costa, J.; Pearson, G.A.; Marbà, N.; Duarte, C.M.; Airoldi, L.; et al. Marine forests of the Mediterranean-Atlantic Cystoseira tamariscifolia complex show a southern Iberian genetic hotspot and no reproductive isolation in parapatry. Sci. Rep. 2018, 8, 10427. [Google Scholar] [CrossRef]
- Orellana, S.; Hernández, M.; Sansón, M. Diversity of Cystoseira sensu lato (Fucales, Phaeophyceae) in the eastern Atlantic and Mediterranean based on morphological and DNA evidence, including Carpodesmia gen. emend. and Treptacantha gen. emend. Eur. J. Phycol. 2019, 54, 447–465. [Google Scholar] [CrossRef]
- Bruno de Sousa, C.; Cox, C.J.; Brito, L.; Pavão, M.M.; Pereira, H.; Ferreira, A.; Ginja, C.; Campino, L.; Bermejo, R.; Parente, M.; et al. Improved Phylogeny of Brown Algae Cystoseira (Fucales) from the Atlantic-Mediterranean Region Based on Mitochondrial Sequences. PLoS ONE 2019, 14, e0210143. [Google Scholar] [CrossRef]
- Cheminée, A.; Sala, E.; Pastor, J.; Bodilis, P.; Thiriet, P.; Mangialajo, L.; Cottalorda, J.M.; Francour, P. Nursery value of Cystoseira forests for Mediterranean rocky reef fishes. J. Exp. Mar. Biol. Ecol. 2013, 442, 70–79. [Google Scholar] [CrossRef]
- Bulleri, F.; Benedetti-Cecchi, L.; Acunto, S.; Cinelli, F.; Hawkins, S.J. The influence of canopy algae on vertical patterns of distribution of low-shore assemblages on rocky coasts in the northwest Mediterranean. J. Exp. Mar. Biol. Ecol. 2002, 267, 89–106. [Google Scholar] [CrossRef]
- Ballesteros, E.; Torras, X.; Pinedo, S.; García, M.; Mangialajo, L.; De Torres, M. A new methodology based on littoral community cartography dominated by macroalgae for the implementation of the European Water Framework Directive. Mar. Pollut. Bull. 2007, 55, 172–180. [Google Scholar] [CrossRef]
- Badreddine, A.; Abboud-Abi Saab, M.; Gianni, F.; Ballesteros, E.; Mangialajo, L. First assessment of the ecological status in the Levant Basin: Application of the CARLIT index along the Lebanese coastline. Ecol. Indic. 2018, 85, 37–47. [Google Scholar] [CrossRef]
- Oceana: Protecting the World’s Oceans. Available online: https://europe.oceana.org/en/home (accessed on 17 March 2019).
- Roberts, M. Active speciation in the taxonomy of the genus Cystoseira C. Agard. In Modern Approaches to the Taxonomy of Red and Brown Algae; Irvine, D.E.G., Price, J.H., Eds.; Academic Press: London, UK; New York, NY, USA, 1978; Volume Systematics. [Google Scholar]
- Ballesteros, E.; Catalán, J. Flora y vegetación marina y litoral del Cabo de Gata y el Puerto de Roquetas de Mar (Almería): Primera aproximación. An. Univ. Murcia 1981, 42, 237–277. [Google Scholar]
- Ballesteros, E.; Pinedo, S. Los bosques de algas pardas y rojas. In Praderas y Bosques Marinos de Andalucía; Luque, A.A., Templado, J.C., Eds.; Consejería de Medio Ambiente, Junta de Andalucía: Sevilla, Spain, 2004; Volume 336, pp. 199–222. [Google Scholar]
- Mangialajo, L.; Chiantore, M.; Cattaneo-Vietti, R. Loss of fucoid algae along a gradient of urbanization, and structure of benthic assemblages. Mar. Ecol. Prog. Ser. 2008, 358, 63–74. [Google Scholar] [CrossRef]
- Sales, M.; Cebrian, E.; Tomas, F.; Ballesteros, E. Pollution Impacts and Recovery Potential in Three Species of the Genus Cystoseira (Fucales, Heterokontophyta). Estuar. Coast. Shelf Sci. 2011, 92, 347–357. [Google Scholar] [CrossRef]
- Thibaut, T.; Blanfuné, A.; Boudouresque, C.F.; Verlaque, M. Decline and local extinction of Fucales in the French Riviera: The harbinger of future extinctions? Mediterr. Mar. Sci. 2015, 16, 206–224. [Google Scholar] [CrossRef]
- Susini, M.; Thibaut, T.; Meinesz, A.; Forcioli, D. A preliminary study of genetic diversity in Cystoseira amentacea (C. Agardh) Bory var. stricta Montagne (Fucales, Phaeophyceae) using random amplified polymorphic DNA. Phycologia 2007, 46, 605–611. [Google Scholar] [CrossRef][Green Version]
- Gianni, F.; Bartolini, F.; Airoldi, L.; Ballesteros, E.; Francour, P.; Guidetti, P.; Meinesz, A.; Thibaut, T.; Mangialajo, L. Conservation and restoration of marine forests in the Mediterranean Sea and the potential role of Marine Protected Areas. Adv. Oceanogr. Limnol. 2013, 4, 83–101. [Google Scholar] [CrossRef]
- De La Fuente, G.; Asnaghia, V.; Chiantorea, M.; Thrushb, S.; Poveroa, P.; Vassalloa, P.; Petrilloa, M.; Paoli, C. The effect of Cystoseira canopy on the value of midlittoral habitats in NW Mediterranean, an emergy assessment. Ecol. Model. 2019, 404, 1–11. [Google Scholar] [CrossRef]
- Zazo, C.; Goy, J.L.; Dabrio, C.J.; Bardají, T.; Hillaire-Marcel, C.; Ghaleb, B.; González-Delgado, J.A.; Soler, V. Pleistocene raised marine terraces of the Spanish Mediterranean and Atlantic coasts: Records of coastal uplift, sea-level high-stands and climate changes. Mar. Geol. 2003, 194, 103–133. [Google Scholar] [CrossRef]
- Terradas-Fernández, M.; Botana Gómez, C.; Valverde Urrea, M.; Zubcoff, J.; Ramos-Esplá, A.A. The dynamics of phytobenthos and its main drivers on abrasion platforms with vermetids (Alicante, Southeastern Iberian Peninsula). Mediterr. Mar. Sci. 2018, 0, 58–68. [Google Scholar] [CrossRef]
- Ballesteros, E. Structure and dynamics of the Cystoseira caespitosa Sauvageau (Fucales, Phaeophyceae) community in the North-Western Mediterranean. Sci. Mar. 1990, 54, 155–168. [Google Scholar]
- Pena-Martín, C.; Juan, A.; Crespo, J.M. Evaluación del estado de conservación de comunidades fitobentónicas en el litoral de Alicante (España). In Oceanos III Milenio, 2nd ed.; Editorial C.P.D.: Madrid, Spain, 2002; Volume 1, p. 142. [Google Scholar]
- Hereu, B.; Garcia-Rubies, A.; Linares, C.; Navarro, L.; Bonaviri, C.; Cebrian, E.; Diaz, D.; Garrabou, J.; Teixidó, N.; Zabala, M. Impact of the Sant Esteve’s storm (2008) on the algal cover in infralittoral rocky photophillic communities. In Assessment of the Ecological Impact of the Extreme Storm of Sant Esteve’s Day (26 December 2008) on the Littoral Ecosystems of the North Mediterranean Spanish Coasts; Mateo, M.A., Garcia-Rubies, T., Eds.; Centro de Estudios Avanzados de Blanes, Consejo Superior de Investigaciones Científicas: Blanes, Spain, 2012; pp. 123–143, Final Report (PIEC, 200430E599). [Google Scholar]
- Mariani, S.; Cefalì, M.E.; Chappuis, E.; Terradas-Fernández, M.; Pinedo, S.; Torras, X.; Jordana, E.; Medrano, A.; Verdura, J.; Ballesteros, E. Past and present of Fucales from shallow and sheltered shores in Catalonia. Reg. Stud. Mar. Sci. 2019, 32, 100824. [Google Scholar] [CrossRef]
- Sales, M.; Ballesteros, E. Shallow Cystoseira (Fucales: Ochrophyta) assemblages thriving in sheltered areas from Menorca (NW Mediterranean): Relationships with environmental factors and anthropogenic pressures. Est. Coast. Shelf Sci. 2009, 84, 476–482. [Google Scholar] [CrossRef]
- Medrano, A. Macroalgal Forests Ecology, Long-Term Monitoring, and Conservation in a Mediterranean Marine Protected Area. Ph.D. Thesis, Universitat de Barcelona, Barcelona, Spain, 2020. [Google Scholar]
- Terradas-Fernández, M. Caracterización de las Fitocenosis de las Plataformas de Abrasión con Vermétidos del Sureste Ibérico. Master’s Thesis, Universidad de Alicante-Universidad Miguel Hernández, Alicante, Spain, 2014. [Google Scholar]
- Ballesteros, E. Els Vegetals i la Zonació Litoral: Espècies, Comunitats i Factors que Influeixen en la Seva Distribució; Arxius Secció Ciències; Institud de Estudis Catalans: Barcelona, Spain, 1992; Volume 101, p. 1. [Google Scholar]
- Ballesteros, E. Production of seaweeds in Northwestern Mediterranean marine communities: Its relation with environmental factors. Sci. Mar. 1989, 53, 357–364. [Google Scholar]
- Menconi, M.; Benedetti-Cecchi, L.; Cinelli, F. Spatial and temporal variability in the distribution of algae and invertebrates on rocky shores in the northwest Mediterranean. J. Exp. Mar. Biol. Ecol. 1999, 233, 1–23. [Google Scholar] [CrossRef]
- Sangil, C.; Sansón, M.; Afonso-Carrillo, J. Spatial variation patterns of subtidal seaweed assemblages along a subtropical oceanic archipelago: Thermal gradient vs herbivore pressure. Est. Coast. Shelf Sci. 2011, 94, 322–333. [Google Scholar] [CrossRef]
- Davis, T.A.; Volesky, B.; Mucci, A. A review of the biochemistry of heavy metal bio-absorption by brown algae. Water Res. 2003, 37, 4311–4330. [Google Scholar] [CrossRef]
- Cho, G.Y.; Lee, S.H.; Boo, S.M. A new Brown algal order, Ishigeales (Phaeophyceae) established on the basis of plastid protein-coding rbcL, psaA, and psbA region comparisons. J. Phycol. 2004, 40, 921–936. [Google Scholar] [CrossRef]
- Coyer, A.; Hoarau, G.; Le-Secq, M.; Stam, W.; Olsen, J.L. A mtDNA-based phylogeny of the brown algal genus Fucus (Heterokontophyta; Phaeophyta). Mol. Phylogenet. Evol. 2006, 39, 209–222. [Google Scholar] [CrossRef]
- Ministerio Para la Transición Ecológica y el Reto Demográfico, Gobierno de España. Available online: https://www.miteco.gob.es/es/costas/temas/proteccion-medio-marino/biodiversidad-marina/espacios-marinos-protegidos/red-natura-2000-ambito-marino/bm_emprot_rednat2000_marino_LIC.aspx (accessed on 17 July 2020).
- European Commission: Natura 2000 Network. Available online: https://natura2000.eea.europa.eu/Natura2000/SDF.aspx?site=ESZZ16008#6 (accessed on 5 August 2020).
- Ministerio de Transportes, Movilidad y Agenda Urbana. Gobierno de España. Plan Nacional de Ortofotografía Aérea: Instituto Geográfico Nacional. Available online: http://www.ign.es/wms/pnoa-historico?request=GetCapabilities&service=WMS (accessed on 7 August 2019).
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; PRIMER-E: Plymouth, UK, 2008. [Google Scholar]
- Gómez-Garreta, A. Flora Phycologica Iberica. Vol. 1. Fucales; Gómez-Garreta, A., Ed.; Universidad de Murcia: Murcia, Spain, 2000. [Google Scholar]
- Cormaci, M.; Furnari, G.; Catra, M.; Alongi, G.; Giaccone, G. Flora marina bentonica del Mediterraneo: Phaeophyceae. Boll. Accad. Gioenia Sci. Nat. 2012, 45, 1–508. [Google Scholar]
- Lane, C.; Mayes, C.; Druehl, L.; Saunders, G. A multi-gene molecular investigation of the kelp (laminariales, Phaeophyceae) supports substantial taxonomic re-organization. J. Phycol. 2006, 42, 493–512. [Google Scholar] [CrossRef]
- Rogers, S.; Bendich, A. Extraction of DNA from plant tissues. In Plant Molecular Biology Manual.; Gelvin, S.B., Schilperoort, R.A., Verma, D.P.S., Eds.; Springer: Dordrecht, Netherlands, 1989. [Google Scholar]
- Stothard, P. The sequence manipulation suite: JavaScript programs for analyzing and formatting protein and DNA sequences. BioTechniques 2000, 28, 1102–1104. [Google Scholar] [CrossRef] [PubMed]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, 636–641. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Celis-Pla, P.S.M.; Bouzon, Z.L.; Hall-Spencer, J.M.; Schmidt, E.C.; Korbee, N.; Figueroa, F.L. Seasonal biochemical and photophysiological responses in the intertidal macroalga Cystoseira tamariscifolia (Ochrophyta). Mar. Environ. Res 2016, 115, 89–97. [Google Scholar] [CrossRef]
- Steneck, R.; Dethier, M. A Functional Group Approach to the Structure of Algal-Dominated Communities. Oikos 1994, 69, 476–498. [Google Scholar] [CrossRef]
- Falace, A.; Zanelli, E.; Bressan, G. Morphological and reproductive phenology of Cystoseira compressa (Esper) Gerloff & Nizzamuddin (Fucales, Fucophyceae) in the Gulf of Trieste (North Adriatic Sea). Ann. Hist. Sci. Soc. 2005, 15, 71–78. [Google Scholar]
- Mancuso, F.P.; Strain, E.M.A.; Piccioni, E.; De Clerck, O.; Sarà, G.; Airoldi, L. Status of vulnerable Cystoseira populations along the Italian infralittoral fringe, and relationships with environmental and anthropogenic variables. Mar. Pollut. Bullet. 2018, 129, 762–771. [Google Scholar] [CrossRef] [PubMed]
- Thibaut, T.; Pinedo, S.; Torras, X.; Ballesteros, E. Long-term decline of the populations of Fucales (Cystoseira spp. and Sargassum spp.) in the Albères coast (France, North-western Mediterranean). Mar. Pollut. Bullet. 2005, 50, 1472–1489. [Google Scholar] [CrossRef]
- Roberts, M. Studies on marine algae of the British Isles. 6. Cystoseira foeniculacea (Linnaeus) Greville. Br. Phycol. Bull. 1968, 3, 547–564. [Google Scholar] [CrossRef]
- INPN: Inventaire National du Patrimoine Culturel. Muséum National d’Histoire Naturelle. Available online: https://inpn.mnhn.fr (accessed on 5 August 2019).
- Devescovi, M. Effects of bottom topography and anthropogenic pressure on northern Adriatic Cystoseira spp. (Phaeophyceae, Fucales). Aquat. Bot. 2015, 121, 26–32. [Google Scholar] [CrossRef]
- Pizzuto, F. On the structure, typology and periodism of a Cystoseira brachycarpa J. Agardh emend. Giaccone community and of a duby community from the eastern coast of Sicily (Mediterranean Sea). Plant Biosyst. 1999, 133, 15–35. [Google Scholar] [CrossRef]
- Puertos del Estado. Predicción de viento y oleaje: Boyas y Mareógrafos. Available online: http://www.puertos.es/es-es/oceanografia/Paginas/portus.aspx (accessed on 22 August 2019).
- Jona Lasinio, G.; Tullio, M.A.; Ventura, D.; Ardizzone, G.; Abdelahad, N. Statistical analysis of the distribution of infralittoral Cystoseira populations on pristine coasts of four Tyrrhenian islands: Proposed adjustment to the CARLIT index. Ecol. Ind. 2017, 73, 293–301. [Google Scholar] [CrossRef]
- Milazzo, M.; Badalamenti, F.; Riggioa, S.; Chemello, R. Patterns of algal recovery and small-scale effects of canopy removal as a result of human trampling on a Mediterranean rocky shallow community. Biol. Cons. 2004, 117, 191–202. [Google Scholar] [CrossRef]
- Piattelli, M. Chemistry and taxonomy of Sicilian Cystoseira species. New J. Chem 1990, 14, 777–782. [Google Scholar]
- Valls, R.; Piovetti, L.; Praud, A. The use of diterpenoids as chemotaxonomic markers in the genus Cystoseira. Hydrobiol. 1993, 260, 549–556. [Google Scholar] [CrossRef]
- Gallardo, T.; Bárbara, I.; Afonso-Carrillo, J.; Bermejo, R.; Altamirano, M.; Gómez-Garreta, A.; Barceló Martí, C.; Rull-Lluch, J.; Ballesteros, E.; De la Rosa, J. Nueva lista crítica de las algas bentónicas marinas de España. Bol. Soc. Esp. Ficol. 2016, 51, 7–52. [Google Scholar]
- Bosc, E.; Bricaud, A.; Antoine, D. Seasonal and inter-annual variability in algal biomass and primary production in the Mediterranean Sea, as derived from 4 years of SeaWiFS observations. Global Biogeochem. Cycl. 2004, 18. [Google Scholar] [CrossRef]
- Amico, V.; Giaccone, G.; Colonna, P.; Mannino, A.M.; Randazzo, R. Un nuovo approccio allo studio della sistematica del genere Cystoseira C. Agardh (Phaeophyta, Fucales). Bol. Sed. Accad. Gioenia Sci. Nat. Catania 1985, 18, 887–985. [Google Scholar]
- Pla, M.; Gómez-Garreta, A. Chorology of the genus Cystoseira C. Agardh (Phaeophyceae, Fucales). An. Jard. Bot. Madr. 1989, 46, 89–97. [Google Scholar]
- Mangialajo, L.; Ruggieri, N.; Asnaghi, V.; Chiantore, M.; Povero, P.; Cattaneo-Vietti, R. Ecological status in the Ligurian Sea: The effect of coastline urbanization and the importance of proper reference sites. Mar. Pollut. Bull. 2007, 55, 30–41. [Google Scholar] [CrossRef]
- Díaz-Valdés, M.; Abellán, E.; Izquierdo, A.; Ramos-Esplá, A.A. Estudio preliminar de comunidades de macroalgas de la franja litoral rocosa de la Comunidad Valenciana dentro de la Directiva Marco del Agua. In Proceedings of the XIV Simposio Ibérico de Estudios de Biología Marina, Barcelona, Spain, 12–15 September 2006. [Google Scholar]
- Blanfuné, A.; Boudouresque, C.F.; Verlaque, M.; Thibaut, T. The fate of Cystoseira crinita, a forest-forming Fucale (Phaeophyceae, Stramenopiles): In France (North Western Mediterranean Sea). Estuar. Coast. Shelf Sci. 2016, 181, 196–208. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | df | SS | MS | Pseudo-F | P (perm) | Uniq. Perms |
|---|---|---|---|---|---|---|
| Width | 3 | 15,532 | 5177.3 | 3679 | 0.0002 | 4986 |
| Wave exposure | 2 | 14,298 | 7148.9 | 5.08 | 0.0002 | 4983 |
| Horizon | 2 | 10,849 | 5424.7 | 3.8548 | 0.0016 | 4985 |
| Width*WaveExp | 5 | 20,474 | 4094.9 | 2.9098 | 0.0002 | 4972 |
| Width*Horizon | 6 | 5638.4 | 939.73 | 0.66778 | 0.8234 | 4974 |
| Horizon*WaveExp | 4 | 6531.8 | 1632.9 | 1.1604 | 0.3108 | 4982 |
| Width*WaveExp*Horiz | 10 | 6083.7 | 608.37 | 0.43231 | 0.9948 | 4982 |
| Res | 150 | 2.1109 × 105 | 1407.3 |
| Width*Wave Exposure | |||
|---|---|---|---|
| Wave Exposure | t | P(perm) | perms |
| Width = 0 (No platform) | |||
| Med–Low | 1.9482 | 0.0438 | 2246 |
| Width = Narrow | |||
| Med–Low | 1.8362 | 0.0408 | 4981 |
| Med–High | 2.5084 | 0.0008 | 4990 |
| Low–High | 1.0787 | 0.3138 | 4983 |
| Width = Medium | |||
| Med–Low | 1.2407 | 0.2272 | 4994 |
| Med–High | 1.7067 | 0.0532 | 4989 |
| Low–High | 1.0416 | 0.3604 | 4992 |
| Width = Wide | |||
| Med–Low | 2.1585 | 0.0034 | 4985 |
| Med–High | 2.1066 | 0.0052 | 4993 |
| Low–High | 2.7538 | 0.0002 | 4992 |
| Horizon | |||
| Horizon | t | P(perm) | perms |
| Prox–Med | 1.4607 | 0.0978 | 4985 |
| Prox–Distal | 2783 | 0.0004 | 4986 |
| Med–Distal | 1314 | 0.1656 | 4987 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jódar-Pérez, A.B.; Terradas-Fernández, M.; López-Moya, F.; Asensio-Berbegal, L.; López-Llorca, L.V. Multidisciplinary Analysis of Cystoseira sensu lato (SE Spain) Suggest a Complex Colonization of the Mediterranean. J. Mar. Sci. Eng. 2020, 8, 961. https://doi.org/10.3390/jmse8120961
Jódar-Pérez AB, Terradas-Fernández M, López-Moya F, Asensio-Berbegal L, López-Llorca LV. Multidisciplinary Analysis of Cystoseira sensu lato (SE Spain) Suggest a Complex Colonization of the Mediterranean. Journal of Marine Science and Engineering. 2020; 8(12):961. https://doi.org/10.3390/jmse8120961
Chicago/Turabian StyleJódar-Pérez, Ana Belén, Marc Terradas-Fernández, Federico López-Moya, Leticia Asensio-Berbegal, and Luis Vicente López-Llorca. 2020. "Multidisciplinary Analysis of Cystoseira sensu lato (SE Spain) Suggest a Complex Colonization of the Mediterranean" Journal of Marine Science and Engineering 8, no. 12: 961. https://doi.org/10.3390/jmse8120961
APA StyleJódar-Pérez, A. B., Terradas-Fernández, M., López-Moya, F., Asensio-Berbegal, L., & López-Llorca, L. V. (2020). Multidisciplinary Analysis of Cystoseira sensu lato (SE Spain) Suggest a Complex Colonization of the Mediterranean. Journal of Marine Science and Engineering, 8(12), 961. https://doi.org/10.3390/jmse8120961