
Oxidative Stress Responses and Recovery of Marine Medaka (Oryzias melastigma) in Early-Life Stages After Acute Exposure to Crude Oil
 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation and Chemical Analysis of Exposure Solutions
2.2. Marine Medaka Maintenance and Embryo Collection
2.3. Experimental Design
2.4. Lethal and Sublethal Morphological Assessment
2.5. Antioxidant Defense Capacity Assessment
2.6. Lipid Peroxidation Assessment
2.7. Non-Specific Immune Response Assessment
2.8. Statistical Analysis
3. Results
3.1. Chemical Analysis in the Water-Accommodated Fractions (WAFs) of Oman Crude Oil (OCO) Stock Solution and Exposure Solution
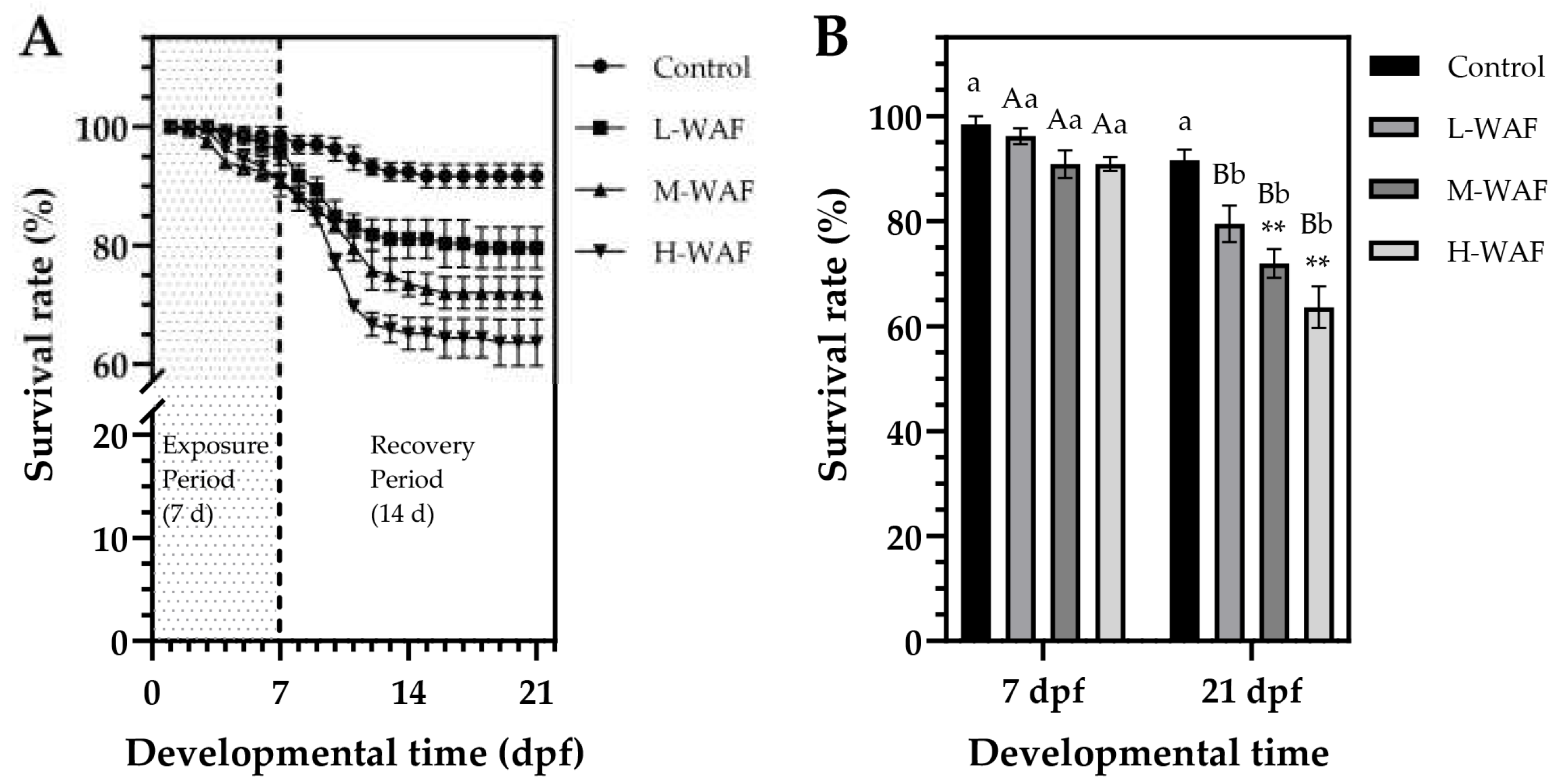
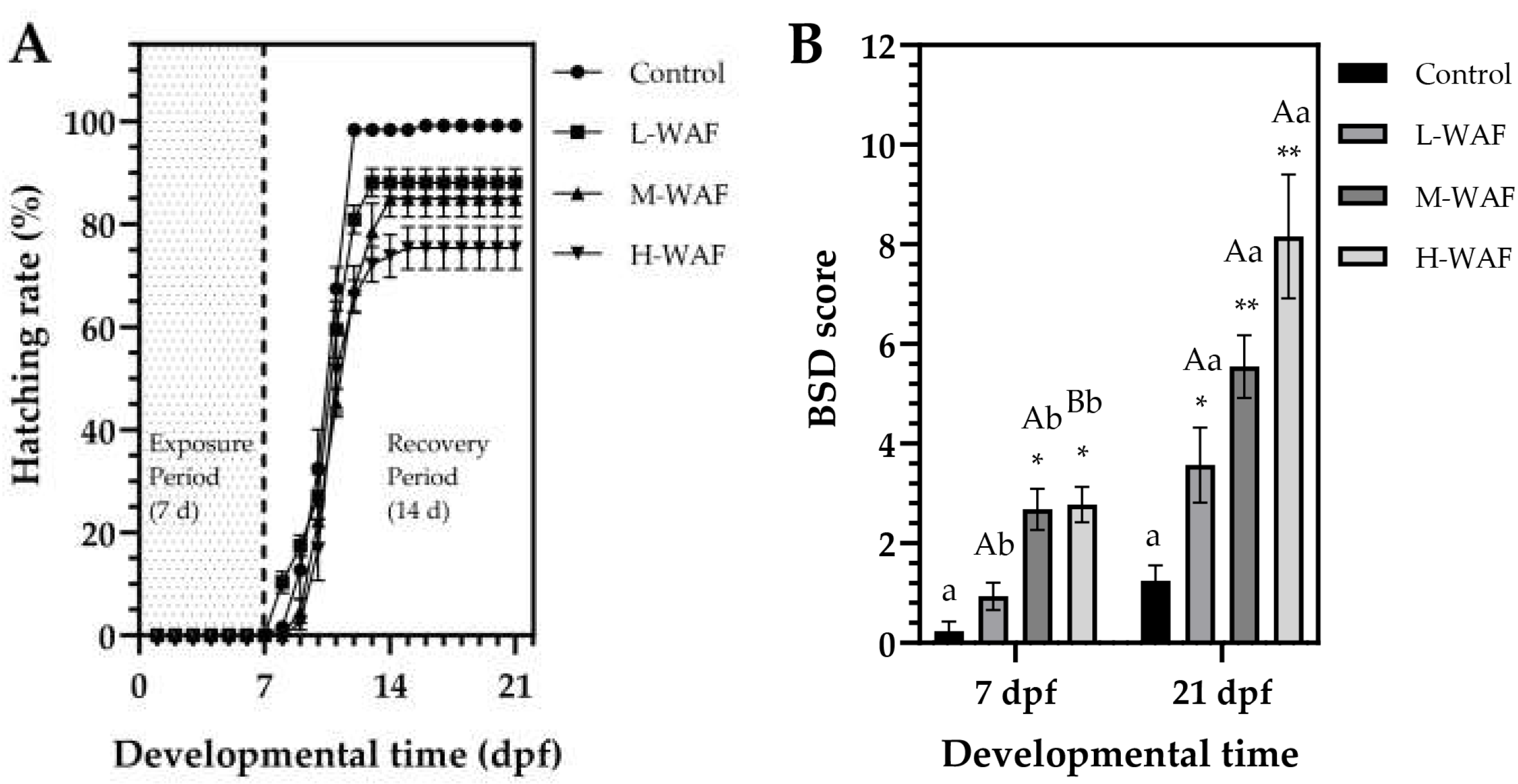
3.2. Lethal and Sublethal Morphological Effects of OCO WAF on Marine Medaka Early-Life Stages (ELS) During Exposure and Recovery Periods
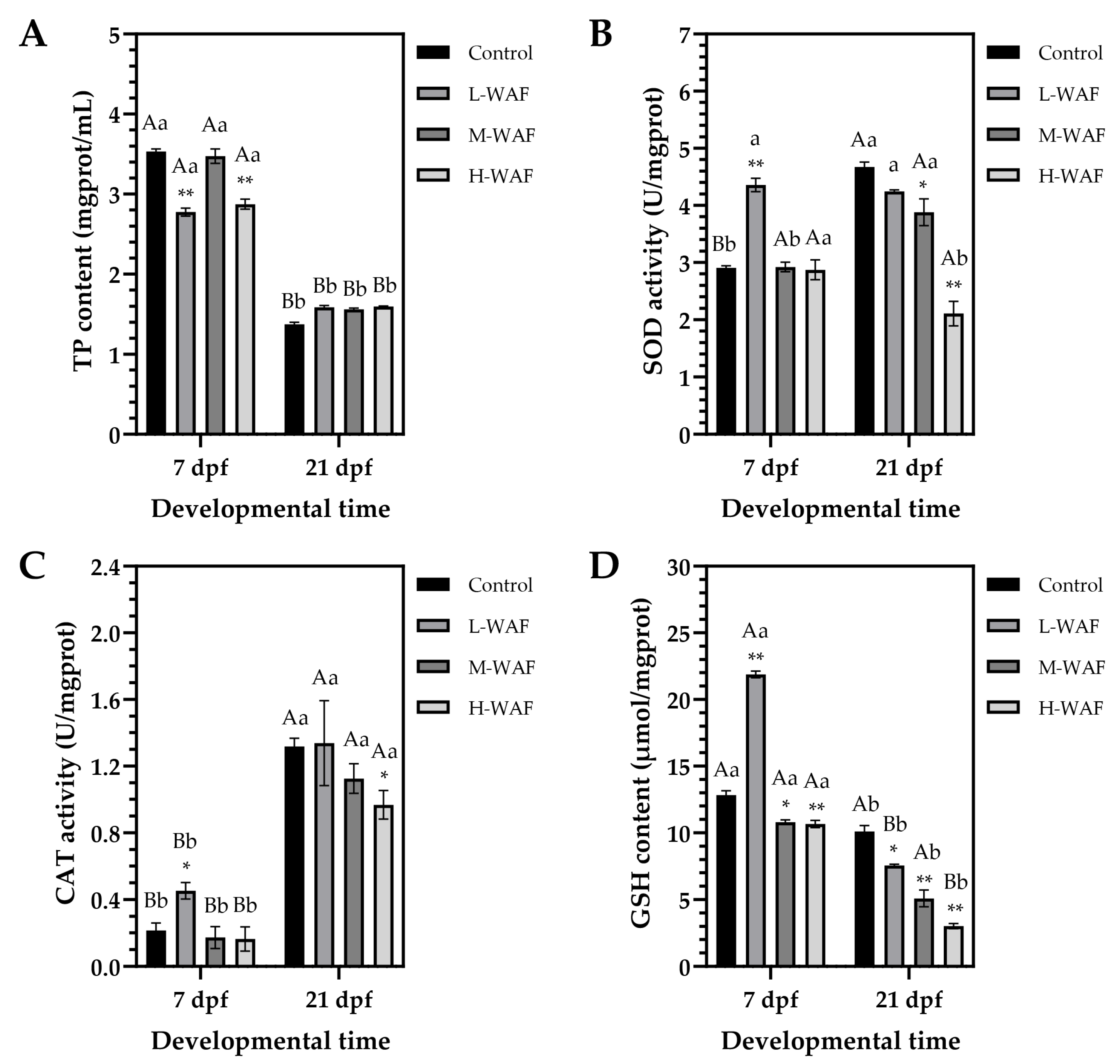
3.3. Effects of OCO WAF on the Antioxidant Defense Capacity of Marine Medaka on ELS During Exposure and Recovery Periods
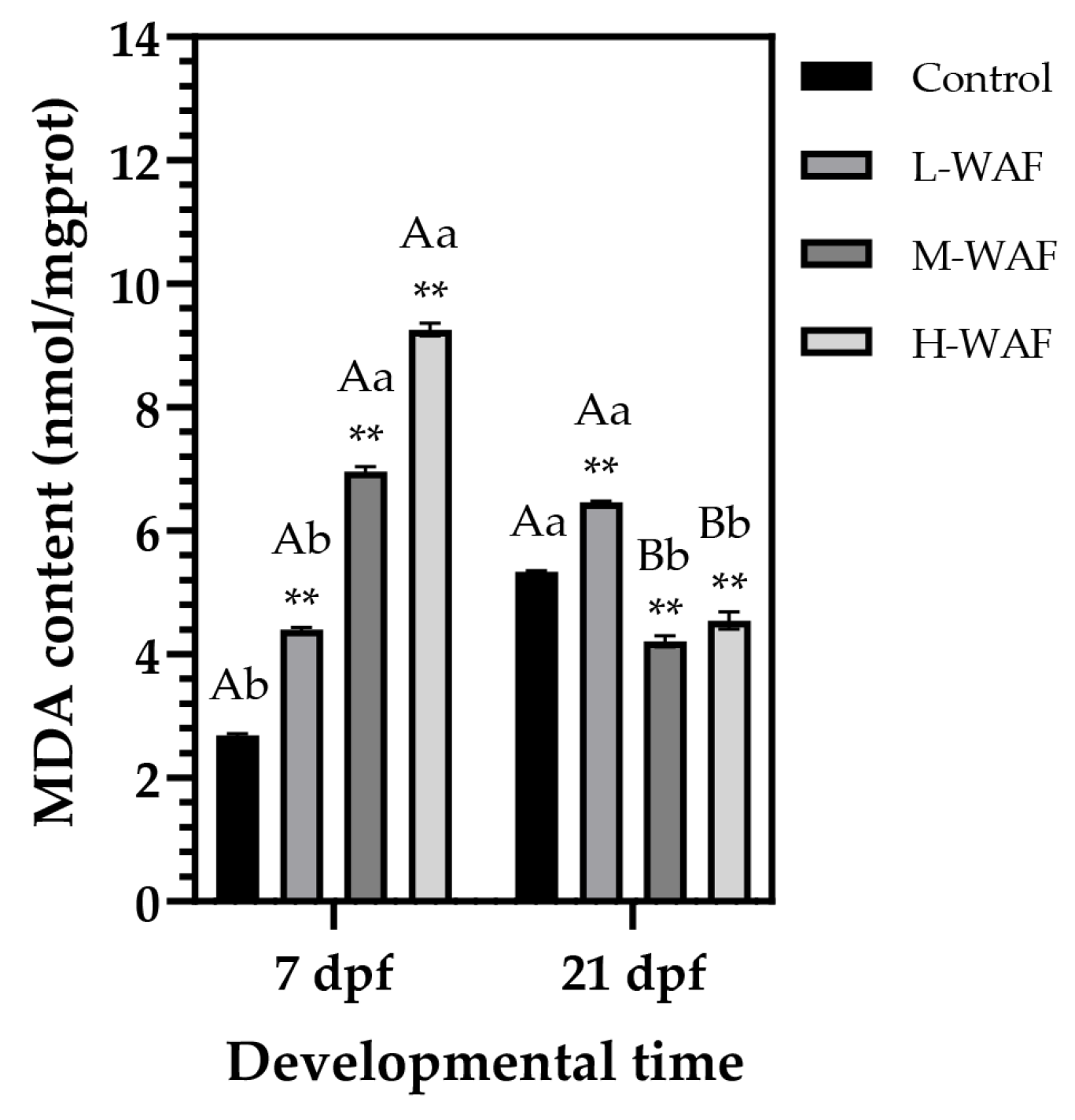
3.4. Effects of OCO WAF on the Lipid Peroxidation of Marine Medaka on ELS During Exposure and Recovery Periods
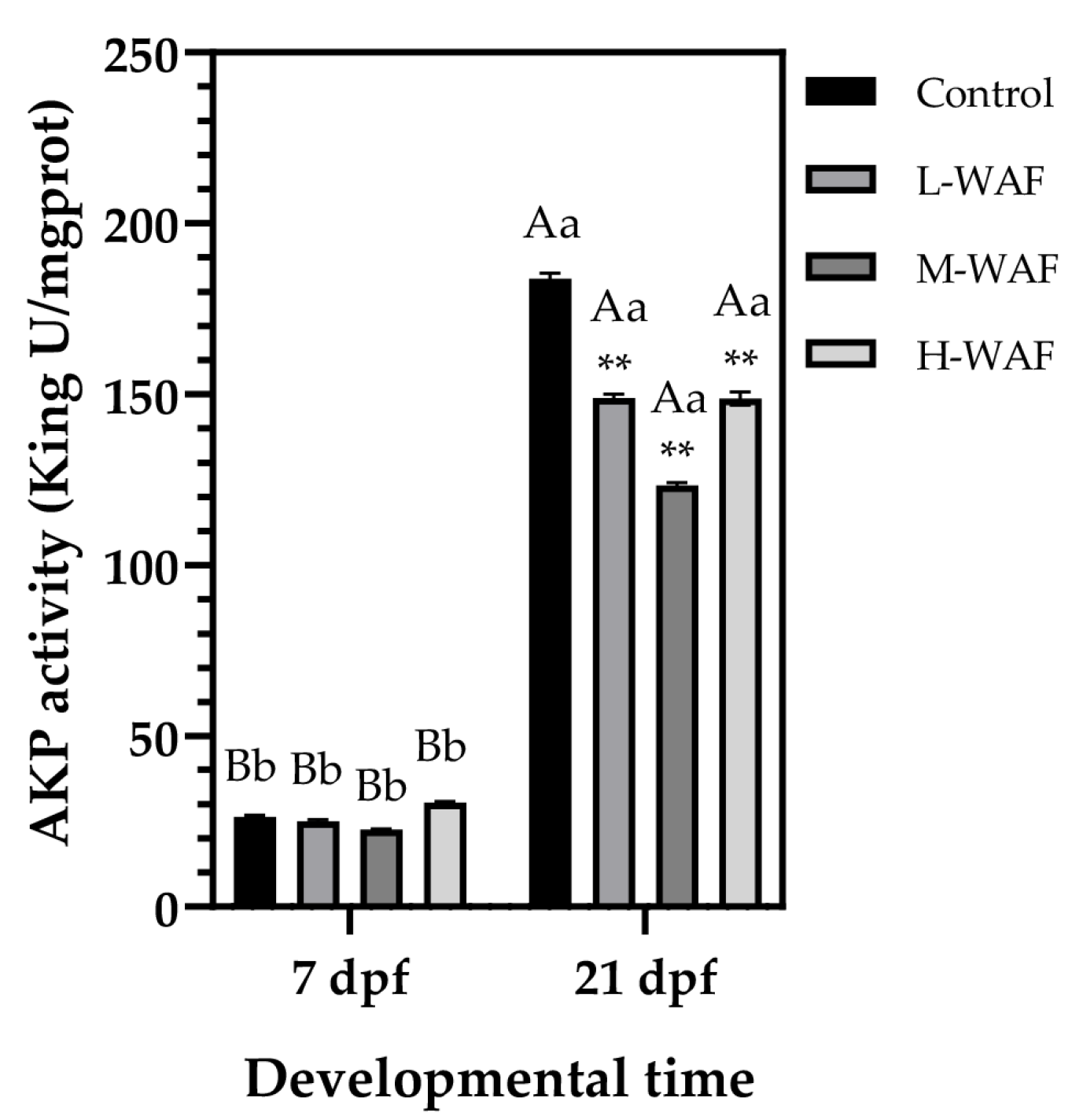
3.5. Effects of OCO WAF on the Immune Function of Marine Medaka on ELS During Exposure and Recovery Periods
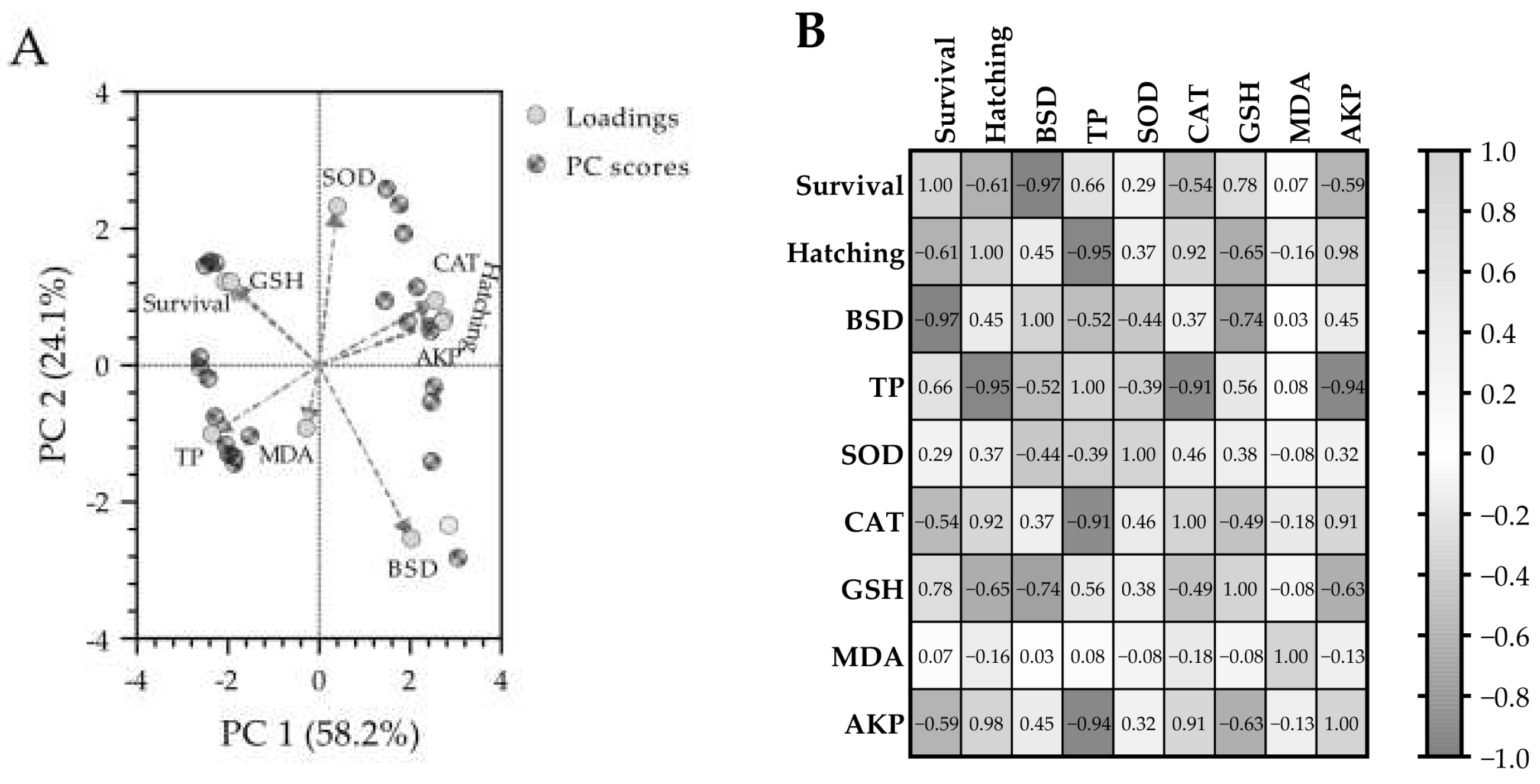
3.6. Principal Component Analysis (PCA) and Pearson Correlation Analysis
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, D.; Zhu, L. Assessing China’s legislation on compensation for marine ecological damage: A case study of the Bohai oil spill. Mar. Pol. 2014, 50, 18–26. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, W.; Wan, Z.; Li, S.; Huang, T.; Fei, Y. Oil spills from global tankers: Status review and future governance. J. Clean. Prod. 2019, 227, 20–32. [Google Scholar] [CrossRef]
- Wang, C.; Lou, Y.; Wang, T.; Li, R.; Peng, M.; Gao, D.; Lei, W. Embryonic exposure to water accommodated fraction of crude oil inhibits reproductive capability in adult female marine medaka (Oryzias melastigma). Chemosphere 2024, 362, 142616. [Google Scholar] [CrossRef]
- Yin, L.; Zhang, M.; Zhang, Y.; Qiao, F. The long-term prediction of the oil-contaminated water from the Sanchi collision in the East China Sea. Acta Oceanol. Sin. 2018, 37, 69–72. [Google Scholar] [CrossRef]
- Li, X.; Xiong, D.; Ding, G.; Fan, Y.; Ma, X.; Wang, C.; Xiong, Y.; Jiang, X. Exposure to water-accommodated fractions of two different crude oils alters morphology, cardiac function and swim bladder development in early-life stages of zebrafish. Chemosphere 2019, 235, 423–433. [Google Scholar] [CrossRef]
- Li, X.; Xiong, D.; Ju, Z.; Xiong, Y.; Ding, G.; Liao, G. Phenotypic and transcriptomic consequences in zebrafish early-life stages following exposure to crude oil and chemical dispersant at sublethal concentrations. Sci. Total Environ. 2021, 763, 143053. [Google Scholar] [CrossRef]
- Xu, Q.; Li, X.; Xu, Z.; Chen, S.; Xiong, D. Water-accommodated fractions of crude oil and its mixture with chemical dispersant impairs oxidase stress and energy metabolism disorders in Oryzias melastigma embryos. Chemosphere 2024, 363, 142912. [Google Scholar] [CrossRef]
- Meador, J.P.; Sommers, F.C.; Ylitalo, G.M.; Sloan, C.A. Altered growth and related physiological responses in juvenile Chinook salmon (Oncorhynchus tshawytscha) from dietary exposure to polycyclic aromatic hydrocarbons (PAHs). Can. J. Fish. Aquat. Sci. 2006, 63, 2364–2376. [Google Scholar] [CrossRef]
- Incardona, J.P.; Collier, T.K.; Scholz, N.L. Defects in cardiac function precede morphological abnormalities in fish embryos exposed to polycyclic aromatic hydrocarbons. Toxicol. Appl. Pharmacol. 2004, 196, 191–205. [Google Scholar] [CrossRef]
- Jin, F.; Wang, Y.; Yu, F.; Liu, X.; Zhang, M.; Li, Z.; Yao, Z.; Cong, Y.; Wang, J. Acute and chronic effects of crude oil water-accommodated fractions on the early life stages of marine medaka (Oryzias melastigma, McClelland, 1839). Toxics 2023, 11, 236. [Google Scholar] [CrossRef]
- Incardona, J.P.; Carls, M.G.; Holland, L.; Linbo, T.L.; Baldwin, D.H.; Myers, M.S.; Peck, K.A.; Tagal, M.; Rice, S.D.; Scholz, N.L. Very low embryonic crude oil exposures cause lasting cardiac defects in salmon and herring. Sci. Rep. 2015, 5, 13499. [Google Scholar] [CrossRef] [PubMed]
- Incardona, J.P.; Carls, M.G.; Teraoka, H.; Sloan, C.A.; Collier, T.K.; Scholz, N.L. Aryl hydrocarbon receptor–independent toxicity of weathered crude oil during fish development. Environ. Health Perspect. 2005, 113, 1755–1762. [Google Scholar] [CrossRef] [PubMed]
- Carls, M.G.; Holland, L.; Larsen, M.; Collier, T.K.; Scholz, N.L.; Incardona, J.P. Fish embryos are damaged by dissolved PAHs, not oil particles. Aquat. Toxicol. 2008, 88, 121–127. [Google Scholar] [CrossRef]
- Cherr, G.N.; Fairbairn, E.; Whitehead, A. Impacts of petroleum-derived pollutants on fish development. Annu. Rev. Anim. Biosci. 2017, 5, 185–203. [Google Scholar] [CrossRef] [PubMed]
- Hodson, P.V. The toxicity to fish embryos of PAH in crude and refined oils. Arch. Environ. Contam. Toxicol. 2017, 73, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Ding, G.; Li, X.; Xiong, D. Comparison of toxicity effects of fuel oil treated by different dispersants on marine medaka (Oryzias melastigma) embryo. Acta Oceanol. Sin. 2018, 37, 123–132. [Google Scholar] [CrossRef]
- Ju, Z.; Li, X.; Li, X.; Liang, C.; Xu, Z.; Chen, H.; Xiong, D. Stranded heavy fuel oil exposure causes deformities, cardiac dysfunction, and oxidative stress in marine medaka Oryzias melastigma using an oiled-gravel-column system. Fish Physiol. Biochem. 2025, 51, 29. [Google Scholar] [CrossRef]
- Mu, J.; Wang, J.; Jin, F.; Wang, X.; Hong, H. Comparative embryotoxicity of phenanthrene and alkyl-phenanthrene to marine medaka (Oryzias melastigma). Mar. Pollut. Bull. 2014, 85, 505–515. [Google Scholar] [CrossRef]
- Heintz, R.A.; Short, J.W.; Rice, S.D. Sensitivity of fish embryos to weathered crude oil: Part II. Increased mortality of pink salmon (Oncorhynchus gorbuscha) embryos incubating downstream from weathered Exxon Valdez crude oil. Environ. Toxicol. Chem. 1999, 18, 494–503. [Google Scholar] [CrossRef]
- Ju, Z.; Liao, G.; Zhang, Y.; Li, N.; Li, X.; Zou, Y.; Yang, W.; Xiong, D. Oxidative stress responses in the respiratory tree and the body wall of sea cucumber Apostichopus japonicus (Selenka) to high temperature. Environ. Sci. Pollut. Res. 2022, 30, 21288–21298. [Google Scholar] [CrossRef]
- Billiard, S.M.; Querbach, K.; Hodson, P.V. Toxicity of retene to early life stages of two freshwater fish species. Environ. Toxicol. Chem. 1999, 18, 2070–2077. [Google Scholar] [CrossRef]
- Incardona, J.P.; Carls, M.G.; Day, H.L.; Sloan, C.A.; Bolton, J.L.; Collier, T.K.; Scholz, N.L. Cardiac arrhythmia is the primary response of embryonic Pacific herring (Clupea pallasi) exposed to crude oil during weathering. Environ. Sci. Technol. 2009, 43, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Regoli, F.; Giuliani, M.E. Oxidative pathways of chemical toxicity and oxidative stress biomarkers in marine organisms. Mar. Environ. Res. 2014, 93, 106–117. [Google Scholar] [CrossRef]
- Lushchak, V.I. Environmentally induced oxidative stress in aquatic animals. Aquat. Toxicol. 2011, 101, 13–30. [Google Scholar] [CrossRef]
- Jiang, M.; Li, L.; Li, Y.; Shen, G.; Shen, X. Oxidative stress in shellfish Sinonovacula constricta exposed to the water accommodated fraction of zero sulfur diesel oil and Pinghu crude oil. Arch. Environ. Contam. Toxicol. 2017, 73, 294–300. [Google Scholar] [CrossRef]
- Corrales, J.; Fang, X.; Thornton, C.; Mei, W.; Barbazuk, W.B.; Duke, M.; Scheffler, B.E.; Willett, K.L. Effects on specific promoter DNA methylation in zebrafish embryos and larvae following benzo[a]pyrene exposure. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2014, 163, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Lushchak, V.I. Free radicals, reactive oxygen species, oxidative stress and its classification. Chem. Biol. Interact. 2014, 224, 164–175. [Google Scholar] [CrossRef]
- Freitas, J.S.; Pereira, T.S.B.; Boscolo, C.N.P.; Garcia, M.N.; de Oliveira Ribeiro, C.A.; de Almeida, E.A. Oxidative stress, biotransformation enzymes and histopathological alterations in Nile tilapia (Oreochromis niloticus) exposed to new and used automotive lubricant oil. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2020, 234, 108770. [Google Scholar] [CrossRef]
- Valavanidis, A.; Vlahogianni, T.; Dassenakis, M.; Scoullos, M. Molecular biomarkers of oxidative stress in aquatic organisms in relation to toxic environmental pollutants. Ecotoxicol. Environ. Saf. 2006, 64, 178–189. [Google Scholar] [CrossRef]
- Sadauskas-Henrique, H.; Braz-Mota, S.; Campos, D.F.; dos Santos Barroso, H.; Kochhann, D.; Luis Val, A.; Maria Fonseca de Almeida-Val, V. Oil spill in an amazon blackwater environment: Biochemical and physiological responses of local fish species. Environ. Res. 2024, 250, 118347. [Google Scholar] [CrossRef]
- Li, X.; Liao, G.; Ju, Z.; Wang, C.; Li, N.; Xiong, D.; Zhang, Y. Antioxidant response and oxidative stress in the respiratory tree of sea cucumber (Apostichopus japonicus) following exposure to crude oil and chemical dispersant. J. Mar. Sci. Eng. 2020, 8, 547. [Google Scholar] [CrossRef]
- Li, X.; Xiong, D.; Li, N.; Zou, Y.; Yang, W.; Ju, Z.; Liao, G. Effects of crude oil and chemically dispersed crude oil on the antioxidant response and apoptosis in the respiratory tree of sea cucumber (Apostichopus japonicus). In Proceedings of the 2021 6th Asia Conference on Environment and Sustainable Development, Phuket, Thailand, 6–8 November 2021; Springer: Singapore, 2022; pp. 375–383. [Google Scholar]
- Livingstone, D.R. Contaminant-stimulated reactive oxygen species production and oxidative damage in aquatic organisms. Mar. Pollut. Bull. 2001, 42, 656–666. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, D.B.; Mello, A.d.A.; Allodi, S.; de Barros, C.M. Acute exposure to water-soluble fractions of marine diesel oil: Evaluation of apoptosis and oxidative stress in an ascidian. Chemosphere 2018, 211, 308–315. [Google Scholar] [CrossRef]
- Sturve, J.; Hasselberg, L.; Fälth, H.; Celander, M.; Förlin, L. Effects of North Sea oil and alkylphenols on biomarker responses in juvenile Atlantic cod (Gadus morhua). Aquat. Toxicol. 2006, 78, S73–S78. [Google Scholar] [CrossRef] [PubMed]
- Reynaud, S.; Deschaux, P. The effects of polycyclic aromatic hydrocarbons on the immune system of fish: A review. Aquat. Toxicol. 2006, 77, 229–238. [Google Scholar] [CrossRef]
- Bado-Nilles, A.; Quentel, C.; Thomas-Guyon, H.; Le Floch, S. Effects of two oils and 16 pure polycyclic aromatic hydrocarbons on plasmatic immune parameters in the European sea bass, Dicentrarchus labrax (Linné). Toxicol. Vitr. 2009, 23, 235–241. [Google Scholar] [CrossRef]
- Pasparakis, C.; Esbaugh, A.J.; Burggren, W.; Grosell, M. Physiological impacts of Deepwater Horizon oil on fish. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2019, 224, 108558. [Google Scholar] [CrossRef]
- Mu, J.; Wang, X.; Jin, F.; Wang, J.; Hong, H. The role of cytochrome P4501A activity inhibition in three- to five-ringed polycyclic aromatic hydrocarbons embryotoxicity of marine medaka (Oryzias melastigma). Mar. Pollut. Bull. 2012, 64, 1445–1451. [Google Scholar] [CrossRef]
- Laurel, B.J.; Copeman, L.A.; Iseri, P.; Spencer, M.L.; Hutchinson, G.; Nordtug, T.; Donald, C.E.; Meier, S.; Allan, S.E.; Boyd, D.T.; et al. Embryonic crude oil exposure impairs growth and lipid allocation in a keystone Arctic forage fish. iScience 2019, 19, 1101–1113. [Google Scholar] [CrossRef]
- Horie, Y.; Takahashi, C. Development of an in vivo acute bioassay using the marine medaka Oryzias melastigma. Environ. Monit. Assess. 2021, 193, 10. [Google Scholar] [CrossRef]
- Kim, H.-S.; Lee, B.-Y.; Han, J.; Jeong, C.-B.; Hwang, D.-S.; Lee, M.-C.; Kang, H.-M.; Kim, D.-H.; Lee, D.; Kim, J.; et al. The genome of the marine medaka Oryzias melastigma. Mol. Ecol. Resour. 2018, 18, 656–665. [Google Scholar] [CrossRef]
- Bo, J.; Cai, L.; Xu, J.-H.; Wang, K.-J.; Au, D.W.T. The marine medaka Oryzias melastigma—A potential marine fish model for innate immune study. Mar. Pollut. Bull. 2011, 63, 267–276. [Google Scholar] [CrossRef]
- Hong, H.; Li, D.; Shen, R.; Wang, X.; Shi, D. Mechanisms of hexabromocyclododecanes induced developmental toxicity in marine medaka (Oryzias melastigma) embryos. Aquat. Toxicol. 2014, 152, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.M.; Kim, J.; Choi, I.Y.; Raisuddin, S.; Au, D.W.; Leung, K.M.; Wu, R.S.; Rhee, J.S.; Lee, J.S. Omics of the marine medaka (Oryzias melastigma) and its relevance to marine environmental research. Mar. Environ. Res. 2016, 113, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Ruan, J.; Lu, M.; Chen, M.; Dai, Z.; Zuo, Z. Combined effects of ocean acidification and crude oil pollution on tissue damage and lipid metabolism in embryo–larval development of marine medaka (Oryzias melastigma). Environ. Geochem. Health 2019, 41, 1847–1860. [Google Scholar] [CrossRef]
- Singer, M.M.; Aurand, D.; Bragin, G.E.; Clark, J.R.; Coelho, G.M.; Sowby, M.L.; Tjeerdema, R.S. Standardization of the preparation and quantitation of water-accommodated fractions of petroleum for toxicity testing. Mar. Pollut. Bull. 2000, 40, 1007–1016. [Google Scholar] [CrossRef]
- Li, X.; Zou, Y.; Xuan, H.; Yang, W.; Liao, G.; Wang, C.; Xiong, D. Impact of physically and chemically dispersed crude oil on the antioxidant defense capacities and non-specific immune responses in sea cucumber (Apostichopus japonicus). J. Mar. Sci. Eng. 2022, 10, 1544. [Google Scholar] [CrossRef]
- GB 17378.4-2007; The Specification for Marine Monitoring-Part 4: Seawater Analysis. Standards Press of China: Beijing, China, 2007; pp. 44–45.
- EPA. Method 8270D: Semivolatile Organic Compounds by Gas Chromatography/Mass Spectrometry, Part of Test Methods for Evaluating Solid Waste, Physical/Chemical Methods; U.S. Environmental Protection Agency: Washington, DC, USA, 2014; p. 71. [Google Scholar]
- ISO 28540:2011; Water Quality-Determination of 16 Polycyclic Aromatic Hydrocarbons (PAH) in Water-Method Using Gas Chromatography with Mass Spectrometric Detection (GC-MS). International Organization for Standardization: Geneva, Switzerland, 2011; p. 24.
- OECD. OECD Guidelines for the testing of chemicals. Section 2: Effects on biotic systems Test No. 236: Fish embryo acute toxicity (FET) test. In Organization for Economic Co-Operation and Development; OECD Publishing: Paris, France, 2013; p. 22. [Google Scholar]
- Wang, J.; Grisle, S.; Schlenk, D. Effects of salinity on aldicarb toxicity in Juvenile rainbow trout (Oncorhynchus mykiss) and striped bass (Morone saxatilis × chrysops). Toxicol. Sci. 2001, 64, 200–207. [Google Scholar] [CrossRef]
- Lavado, R.; Shi, D.; Schlenk, D. Effects of salinity on the toxicity and biotransformation of l-selenomethionine in Japanese medaka (Oryzias latipes) embryos: Mechanisms of oxidative stress. Aquat. Toxicol. 2012, 108, 18–22. [Google Scholar] [CrossRef]
- Duan, M.; Xiong, D.; Yang, M.; Xiong, Y.; Ding, G. Parental exposure to heavy fuel oil induces developmental toxicity in offspring of the sea urchin Strongylocentrotus intermedius. Ecotoxicol. Environ. Saf. 2018, 159, 109–119. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Peskin, A.V.; Winterbourn, C.C. Assay of superoxide dismutase activity in a plate assay using WST-1. Free Radic. Biol. Med. 2017, 103, 188–191. [Google Scholar] [CrossRef] [PubMed]
- Góth, L. A simple method for determination of serum catalase activity and revision of reference range. Clin. Chim. Acta 1991, 196, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Smith, I.K.; Vierheller, T.L.; Thorne, C.A. Assay of glutathione reductase in crude tissue homogenates using 5,5′-dithiobis(2-nitrobenzoic acid). Anal. Biochem. 1988, 175, 408–413. [Google Scholar] [CrossRef] [PubMed]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Powell, M.E.; Smith, M.J. The determination of serum acid and alkaline phosphatase activity with 4-aminoantipyrine (A.A.P.). J. Clin. Pathol. 1954, 7, 245–248. [Google Scholar] [CrossRef]
- Incardona, J.P.; Vines, C.A.; Anulacion, B.F.; Baldwin, D.H.; Day, H.L.; French, B.L.; Labenia, J.S.; Linbo, T.L.; Myers, M.S.; Olson, O.P.; et al. Unexpectedly high mortality in Pacific herring embryos exposed to the 2007 Cosco Busan oil spill in San Francisco Bay. Proc. Natl. Acad. Sci. USA 2012, 109, E51–E58. [Google Scholar] [CrossRef]
- Xu, E.G.; Mager, E.M.; Grosell, M.; Pasparakis, C.; Schlenker, L.S.; Stieglitz, J.D.; Benetti, D.; Hazard, E.S.; Courtney, S.M.; Diamante, G.; et al. Time- and oil-dependent transcriptomic and physiological responses to Deepwater Horizon oil in mahi-mahi (Coryphaena hippurus) embryos and larvae. Environ. Sci. Technol. 2016, 50, 7842–7851. [Google Scholar] [CrossRef]
- Mu, J.; Jin, F.; Ma, X.; Lin, Z.; Wang, J. Comparative effects of biological and chemical dispersants on the bioavailability and toxicity of crude oil to early life stages of marine medaka (Oryzias melastigma). Environ. Toxicol. Chem. 2014, 33, 2576–2583. [Google Scholar] [CrossRef]
- Pannetier, P.; Morin, B.; Clérandeau, C.; Lacroix, C.; Cabon, J.; Cachot, J.; Danion, M. Comparative biomarker responses in Japanese medaka (Oryzias latipes) exposed to benzo[a]pyrene and challenged with betanodavirus at three different life stages. Sci. Total Environ. 2019, 652, 964–976. [Google Scholar] [CrossRef] [PubMed]
- Bender, M.L.; Giebichenstein, J.; Teisrud, R.N.; Laurent, J.; Frantzen, M.; Meador, J.P.; Sorensen, L.; Hansen, B.H.; Reinardy, H.C.; Laurel, B.; et al. Combined effects of crude oil exposure and warming on eggs and larvae of an arctic forage fish. Sci. Rep. 2021, 11, 8410. [Google Scholar] [CrossRef] [PubMed]
- Philibert, D.A.; Lyons, D.D.; Tierney, K.B. Comparing the effects of unconventional and conventional crude oil exposures on zebrafish and their progeny using behavioral and genetic markers. Sci. Total Environ. 2021, 770, 144745. [Google Scholar] [CrossRef]
- Nahrgang, J.; Dubourg, P.; Frantzen, M.; Storch, D.; Dahlke, F.; Meador, J.P. Early life stages of an arctic keystone species (Boreogadus saida) show high sensitivity to a water-soluble fraction of crude oil. Environ. Pollut. 2016, 218, 605–614. [Google Scholar] [CrossRef]
- Mager, E.M.; Pasparakis, C.; Schlenker, L.S.; Yao, Z.; Bodinier, C.; Stieglitz, J.D.; Hoenig, R.; Morris, J.M.; Benetti, D.D.; Grosell, M. Assessment of early life stage mahi-mahi windows of sensitivity during acute exposures to Deepwater Horizon crude oil. Environ. Toxicol. Chem. 2017, 36, 1887–1895. [Google Scholar] [CrossRef]
- Jung, J.H.; Kim, M.; Yim, U.H.; Ha, S.Y.; Shim, W.J.; Chae, Y.S.; Kim, H.; Incardona, J.P.; Linbo, T.L.; Kwon, J.H. Differential toxicokinetics determines the sensitivity of two marine embryonic fish exposed to Iranian heavy crude oil. Environ. Sci. Technol. 2015, 49, 13639–13648. [Google Scholar] [CrossRef]
- Li, X.; Wang, C.; Li, N.; Gao, Y.; Ju, Z.; Liao, G.; Xiong, D. Combined effects of elevated temperature and crude oil pollution on oxidative stress and apoptosis in sea cucumber (Apostichopus japonicus, Selenka). Int. J. Environ. Res. Public Health 2021, 18, 801. [Google Scholar] [CrossRef]
- Sies, H. Oxidative stress: Concept and some practical aspects. Antioxidants 2020, 9, 852. [Google Scholar] [CrossRef] [PubMed]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef]
- He, F.; Li, X.; Huo, C.; Chu, S.; Cui, Z.; Li, Y.; Wan, J.; Liu, R. Evaluation of fluorene-caused ecotoxicological responses and the mechanism underlying its toxicity in Eisenia fetida: Multi-level analysis of biological organization. J. Hazard. Mater. 2022, 437, 129342. [Google Scholar] [CrossRef] [PubMed]
- Circu, M.L.; Aw, T.Y. Glutathione and modulation of cell apoptosis. Biochim. Biophys. Acta 2012, 1823, 1767–1777. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Tian, L.; Zhao, Q.; Tao, Z.; Yang, J.; Zhou, Y.; Cao, R.; Zhang, G.; Wu, W. Evaluation of the acute toxic effects of crude oil on intertidal mudskipper (Boleophthalmus pectinirostris) based on antioxidant enzyme activity and the integrated biomarker response. Environ. Pollut. 2022, 292, 118341. [Google Scholar] [CrossRef]
- Kerambrun, E.; Le Floch, S.; Sanchez, W.; Thomas Guyon, H.; Meziane, T.; Henry, F.; Amara, R. Responses of juvenile sea bass, Dicentrarchus labrax, exposed to acute concentrations of crude oil, as assessed by molecular and physiological biomarkers. Chemosphere 2012, 87, 692–702. [Google Scholar] [CrossRef]
- Milinkovitch, T.; Imbert, N.; Sanchez, W.; Le Floch, S.; Thomas-Guyon, H. Toxicological effects of crude oil and oil dispersant: Biomarkers in the heart of the juvenile golden grey mullet (Liza aurata). Ecotoxicol. Environ. Saf. 2013, 88, 1–8. [Google Scholar] [CrossRef]
- Helen, N.E.; Nathan, A.A.; Mathias, N.U.; Nwani, C.D. Genotoxicity and oxidative stress evaluations in juvenile African catfish Clarias gariepinus exposed to NPK fertilizer. J. Aquat. Anim. Health 2020, 32, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.-H.; Ko, J.; Lee, E.-H.; Choi, K.-M.; Kim, M.; Yim, U.H.; Lee, J.-S.; Shim, W.J. RNA seq- and DEG-based comparison of developmental toxicity in fish embryos of two species exposed to Iranian heavy crude oil. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2017, 196, 1–10. [Google Scholar] [CrossRef]
- Ozhan, K.; Zahraeifard, S.; Smith, A.P.; Bargu, S. Induction of reactive oxygen species in marine phytoplankton under crude oil exposure. Environ. Sci. Pollut. Res. 2015, 22, 18874–18884. [Google Scholar] [CrossRef]
- Wang, X.; Li, X.; Xiong, D.; Ren, H.; Chen, H.; Ju, Z. Exposure of adult sea urchin Strongylocentrotus intermedius to stranded heavy fuel oil causes developmental toxicity on larval offspring. PeerJ 2022, 10, e13298. [Google Scholar] [CrossRef]
- Wang, X.; Ren, H.; Li, X.; Chen, H.; Ju, Z.; Xiong, D. Sex-specific differences in the toxic effects of heavy fuel oil on sea urchin (Strongylocentrotus intermedius). Int. J. Environ. Res. Public Health 2021, 18, 499. [Google Scholar] [CrossRef]
- Sureda, A.; Box, A.; Tejada, S.; Blanco, A.; Caixach, J.; Deudero, S. Biochemical responses of Mytilus galloprovincialis as biomarkers of acute environmental pollution caused by the Don Pedro oil spill (Eivissa Island, Spain). Aquat. Toxicol. 2011, 101, 540–549. [Google Scholar] [CrossRef] [PubMed]
- Lallès, J.-P. Biology, environmental and nutritional modulation of skin mucus alkaline phosphatase in fish: A review. Fish Shellfish Immunol. 2019, 89, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Weng, Y.; Bo, J.; Zheng, R.; Zhang, Y.; Hong, F.; Xie, Q.; Fang, C.; Jiang, Y. Responses of lysosomal biomarkers from the clam Ruditapes philippinarum to water soluble fraction of diesel oil. J. Appl. Oceanogr. 2020, 39, 195–205. [Google Scholar] [CrossRef]
- Cao, J.; Feng, C.; Xie, L.; Li, L.; Chen, J.; Yun, S.; Guo, W.; Wang, T.; Wu, Y.; Meng, R.; et al. Sesamin attenuates histological alterations, oxidative stress and expressions of immune-related genes in liver of zebrafish (Danio rerio) exposed to fluoride. Fish Shellfish Immunol. 2020, 106, 715–723. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Abbreviation | Score Range | Scoring Criteria |
|---|---|---|---|
| Yolk sac edema | YE | 0–3 | 0 = no edema 1 = mild edema (slight membrane separation) 2 = moderate edema (pronounced membrane separation) 3 = severe edema (extreme separation impeding movement) |
| Pericardial edema | PE | 0–3 | 0 = no edema 1 = mild edema (slight membrane separation) 2 = moderate edema (pronounced membrane separation) 3 = severe edema (extreme separation impeding circulation) |
| Heart malformation | HM | 0–1 | 0 = normal 1 = deformed cardiac structure or function |
| Hemorrhage | HE | 0–1 | 0 = absent 1 = visible blood accumulation in tissues |
| Craniofacial malformation | CM | 0–1 | 0 = normal 1 = abnormal head/face development |
| Spinal deformity | SD | 0–1 | 0 = normal 1 = curved or abnormal spine |
| Decreased circulation | DC | 0–1 | 0 = normal blood flow 1 = reduced blood flow |
| Fin rot | FR | 0–1 | 0 = normal 1 = deterioration of fin tissue |
| Loss of mobility | LA | 0–1 | 0 = normal movement 1 = impaired movement capabilities |
| Death | DE | 13 | Maximum BSD score assigned to dead individuals |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Dai, Y.; Li, X.; Guo, H.; Dai, J.; Wang, H.; Xiong, D.; Liao, G. Oxidative Stress Responses and Recovery of Marine Medaka (Oryzias melastigma) in Early-Life Stages After Acute Exposure to Crude Oil. J. Mar. Sci. Eng. 2025, 13, 965. https://doi.org/10.3390/jmse13050965
Li X, Dai Y, Li X, Guo H, Dai J, Wang H, Xiong D, Liao G. Oxidative Stress Responses and Recovery of Marine Medaka (Oryzias melastigma) in Early-Life Stages After Acute Exposure to Crude Oil. Journal of Marine Science and Engineering. 2025; 13(5):965. https://doi.org/10.3390/jmse13050965
Chicago/Turabian StyleLi, Xishan, Yuekun Dai, Xin Li, Haiqiang Guo, Jiarui Dai, Haonan Wang, Deqi Xiong, and Guoxiang Liao. 2025. "Oxidative Stress Responses and Recovery of Marine Medaka (Oryzias melastigma) in Early-Life Stages After Acute Exposure to Crude Oil" Journal of Marine Science and Engineering 13, no. 5: 965. https://doi.org/10.3390/jmse13050965
APA StyleLi, X., Dai, Y., Li, X., Guo, H., Dai, J., Wang, H., Xiong, D., & Liao, G. (2025). Oxidative Stress Responses and Recovery of Marine Medaka (Oryzias melastigma) in Early-Life Stages After Acute Exposure to Crude Oil. Journal of Marine Science and Engineering, 13(5), 965. https://doi.org/10.3390/jmse13050965