
Scale-Up of an Aquaculture Plant for Reproduction and Conservation of the Sea Urchin Paracentrotus lividus: Development of Post-Larval Feeds

 , ,
, , 
 and
and 
Abstract
1. Introduction
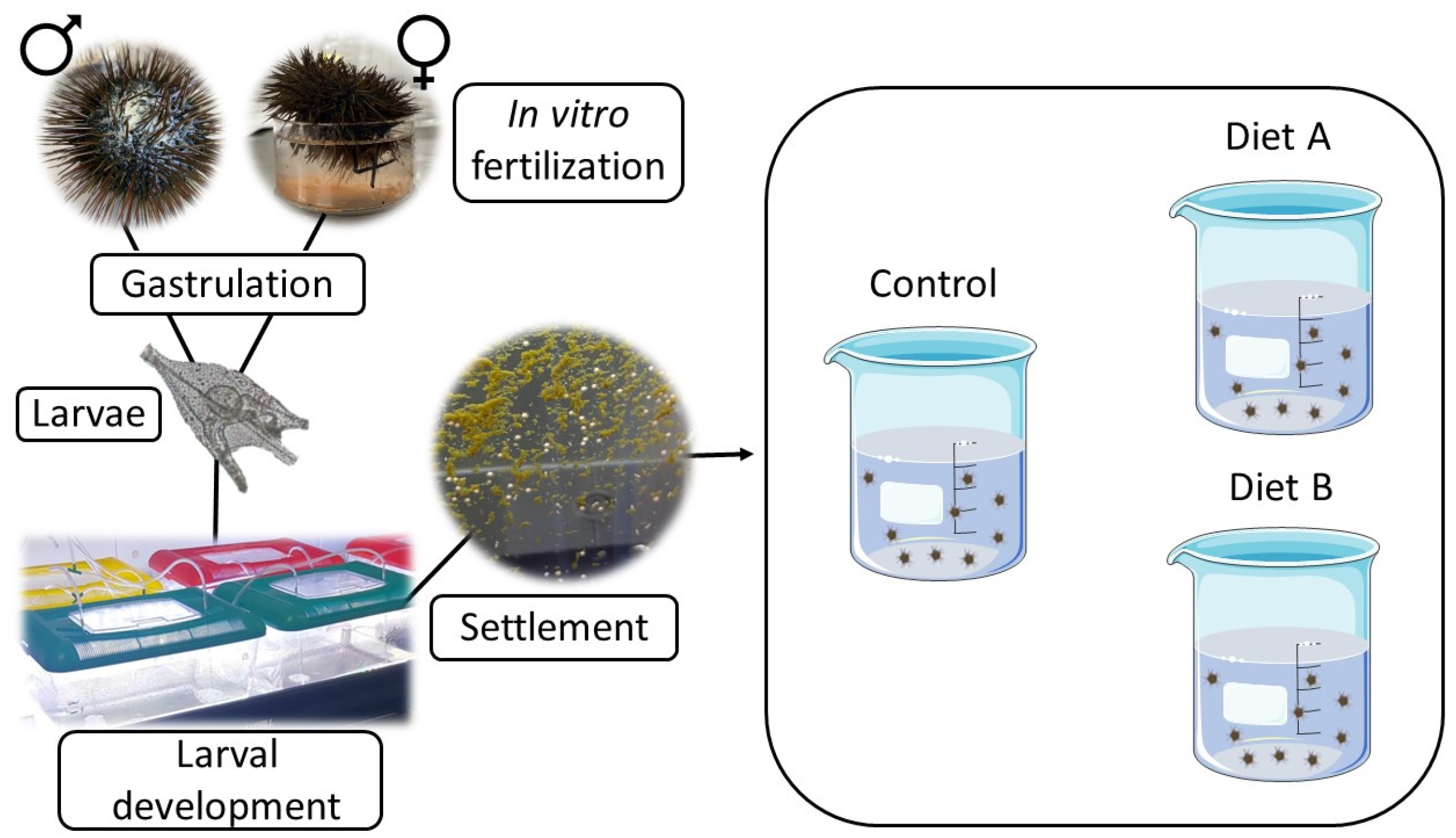
2. Materials and Methods
2.1. Gamete Collection and Larval Development
2.2. Feeding Experiments
2.3. Biotic and Abiotic Measurements
2.4. Statistical Analyses
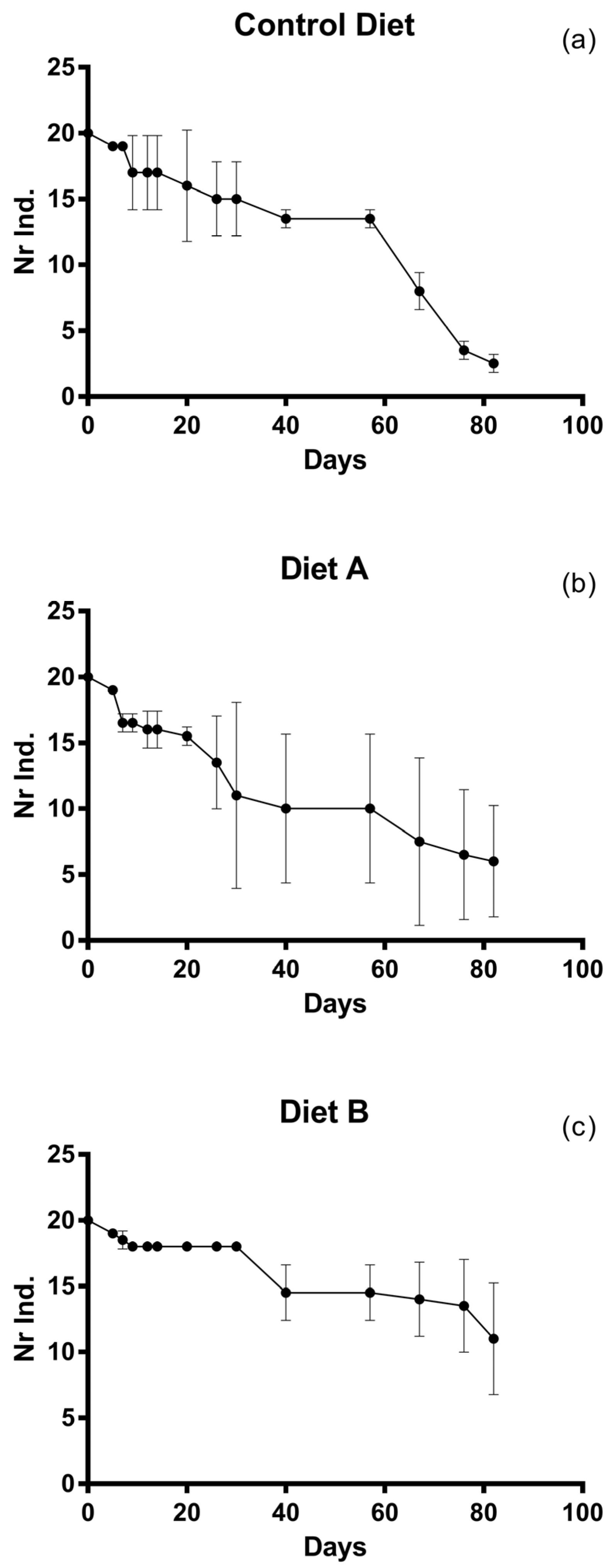
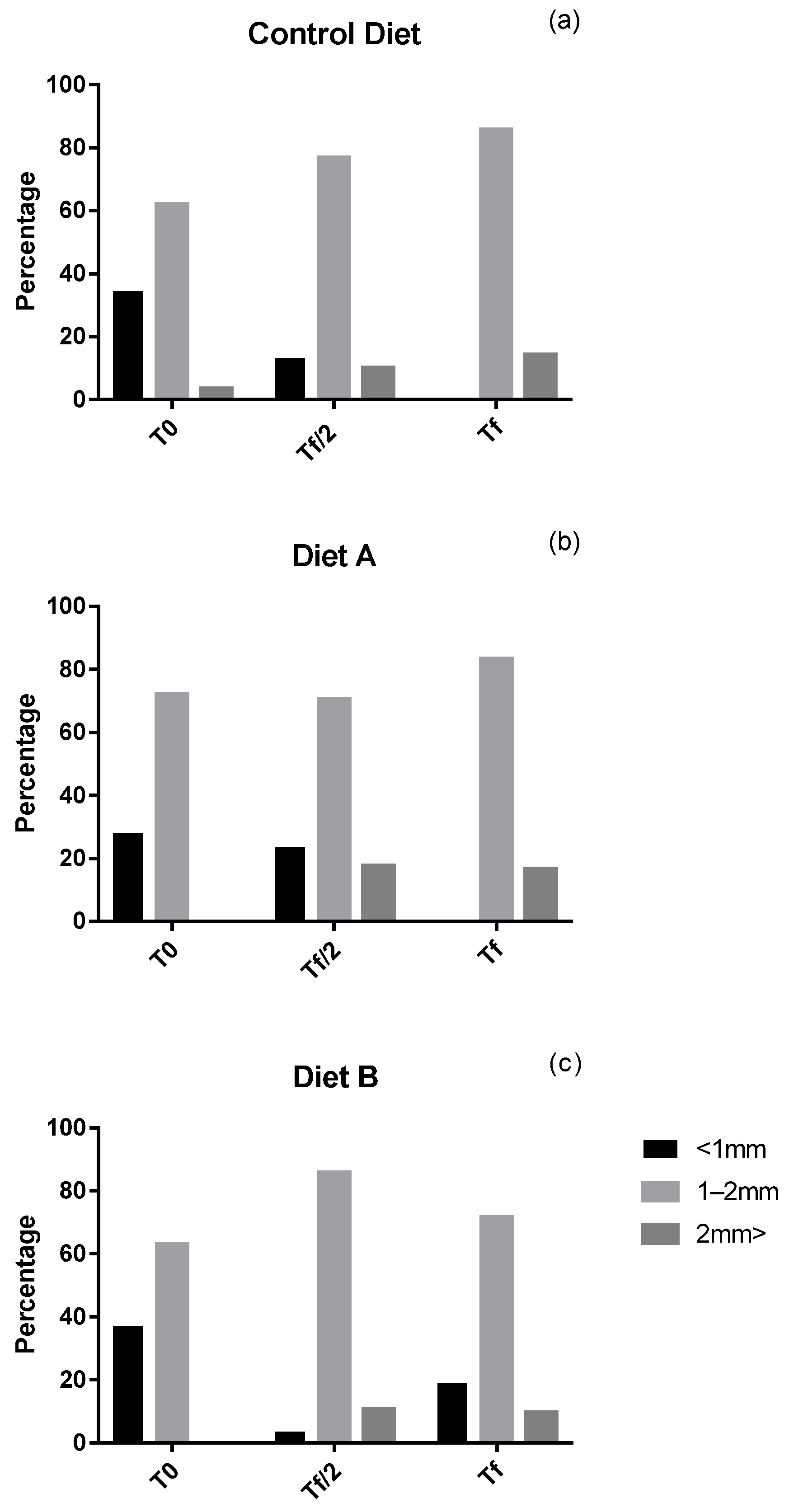
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boudouresque, C.F.; Verlaque, M. Ecology of Paracentrotus lividus. In Edible Sea Urchin: Biology and Ecology; Lawrence, J.M., Ed.; Elsevier Science B.V: Amsterdam, The Netherlands, 2007; pp. 243–285. [Google Scholar]
- Zupi, V.; Fresi, E. A study on the food web of the Posidonia oceanica ecosystem: Analysis of the gut contents of echinoderms. In International Workshop on Posidonia oceanica Beds 1; GIS Posidonie Publishing: Marseille, France, 1984; pp. 373–379. [Google Scholar]
- Andrew, N.L.; Agatsuma, Y.; Ballesteros, E.; Bazhin, A.G.; Creaser, E.P.; Barnes, D.K.A.; Xiaoqi, Z. Status and management of world sea urchin fisheries. Oceanogr. Mar. Biol. 2002, 40, 343–425. [Google Scholar]
- Boudouresque, C.F.; Verlaque, M. Paracentrotus lividus. In Developments in Aquaculture and Fisheries Science; Elsevier: Amsterdam, The Netherlands, 2013; pp. 297–327. [Google Scholar]
- McBride, S.C. Sea urchin aquaculture. Am. Fish. Soc. Symp. 2005, 46, 179. [Google Scholar]
- Unuma, T. Gonadal growth,and its relationship to aquaculture in sea urchins. In The Sea Urchin: From Basic Biology to Aquaculture; Swets & Zeitlinger: Lisse, The Netherlands, 2002; pp. 115–127. [Google Scholar]
- Addis, P.; Secci, M.; Manunza, A.; Corrias, S.; Niffoi, A.; Cau, A. A geostatistical approach for the stock assessment of the edible sea urchin, Paracentrotus lividus, in four coastal zones of Southern and West Sardinia (SW Italy, Mediterranean Sea). Fish. Res. 2009, 100, 215–221. [Google Scholar] [CrossRef]
- Pais, A.; Chessa, L.A.; Serra, S.; Ruiu, A.; Meloni, G.; Donno, Y. The impact of commercial and recreational harvesting for Paracentrotus lividus on shallow rocky reef sea urchin communities in North-western Sardinia, Italy. Estuarine. Coast. Shelf Sci. 2007, 73, 589–597. [Google Scholar] [CrossRef]
- Pearce, C. Sea urchin aquaculture. Bull. Aquac. Assoc. Can. 2010, 108, 1–2. [Google Scholar]
- Grosjean, P. Growth Model of the Reared Sea Urchin Paracentrotus lividus (Lamarck, 1816). Ph.D. Thesis, Universite Libre De Bruxelles, Brussels, Belgium, 2001. [Google Scholar]
- Carboni, S.; Hughes, A.D.; Atack, T.; Tocher, D.R.; Migaud, H. Influence of broodstock diet on somatic growth, fecundity, gonad carotenoids and larval survival of sea urchin. Aquac. Res. 2013, 46, 969–976. [Google Scholar] [CrossRef]
- Spirlet, C. Biologie de L’oursin Comestible (Paracentrotus lividus): Contrôle du cycle reproducteur et optimalisation de la phase de remplissage gonadique. Ph.D. Thesis, Universite Libre De Bruxelles, Brussels, Belgium, 1999. [Google Scholar]
- Spirlet, C.; Grosjean, P.; Jangoux, M. Reproductive cycle of the echinoid Paracentrotus lividus: Analysis by means of maturity index. Invertebr. Reprod. Dev. 1998, 1, 69–81. [Google Scholar] [CrossRef]
- Dhert, P.; Rombaut, G.; Suantika, G.; Sorgeloos, P. Advancement of rotifer culture and manipulation techniques in Europe. Aquaculture 2001, 200, 129–146. [Google Scholar] [CrossRef]
- Buitrago, E.; Lodeiros, C.; Lunar, K.; Alvarado, D.; Indorf, F.; Frontado, K.; Moreno, P.; Vasquez, Z. Mass production of competent larvae of the sea urchin Lytechinus variegatus (Echinodermata: Echinoidea). Aquac. Int. 2005, 4, 359–367. [Google Scholar] [CrossRef]
- Carboni, S.; Kelly, M.S.; Hughes, A.D.; Vignier, J.; Atack, T.; Migaud, H. Evaluation of flow through culture technique for commercial production of sea urchin (Paracentrotus lividus) larvae. Aquac. Res. 2014, 45, 768–772. [Google Scholar] [CrossRef]
- Carboni, S.; Vignier, J.; Chiantore, M.; Tocher, D.R.; Migaud, H. Effects of dietary microalgae on growth, survival and fatty acid composition of sea urchin Paracentrotus lividus throughout larval development. Aquaculture 2012, 324, 250–258. [Google Scholar] [CrossRef]
- Kelly, M.S.; Hunter, A.J.; Scholfield, C.L.; McKenzie, J.D. Morphology and survivorship of larval Psammechinus miliaris (Gmelin) (Echinodermata: Echinoidea) in response to varying food quantity and quality. Aquaculture 2000, 183, 223–240. [Google Scholar] [CrossRef]
- Liu, H.; Kelly, M.S.; Cook, E.J.; Black, K.; Orr, H.; Zhu, J.X.; Dong, S.L. The effect of diet type on growth and fatty-acid composition of sea urchin larvae, I. Paracentrotus lividus (Lamarck, 1816) (Echinodermata). Aquaculture 2007, 264, 247–262. [Google Scholar] [CrossRef]
- Jimmy, R.A.; Kelly, M.S.; Beaumont, A.R. The effect of diet type and quantity on the development of common sea urchin larvae Echinus esculentus. Aquaculture 2003, 220, 261–275. [Google Scholar] [CrossRef]
- Falugi, C.; Angelini, C.; Yokota, Y.; Matranga, V.; Smolenicka, Z. Sea urchin development from the egg to metamorphosis: An integrated model for cell-to-cell and environment interaction. In The Sea Urchin: From Basic Biology to Aquaculture; Swets & Zeitlinger: Lisse, The Netherlands, 2002; pp. 73–93. [Google Scholar]
- Lawrence, J.M.; Olave, S.; Otaiza, R.; Lawrence, A.L.; Bustos, E. Enhancement of gonad production in the sea urchin Loxechinus albus in Chile fed extruded feeds. J. World Aquac. Soc. 1977, 28, 91–96. [Google Scholar] [CrossRef]
- Taylor, A.M.; Powell, M.L.; Watts, S.A.; Lawrence, A.L. Formulated feed supports weight gain and survivorship in juvenile sea urchins Lytechinus variegatus. J. World Aquac. Soc. 2009, 40, 780–787. [Google Scholar] [CrossRef]
- Fernandez, C. Croissance et nutrition de Paracentrotus lividus dans le cadre d’un projet aquacole avec alimentation artificielle. Ph.D. Thesis, Ocean University Sri Lanka, Colombo, Sri Lanka, 1996; p. 279. [Google Scholar]
- Zhang, F.; Man, Y.B.; Mo, W.Y.; Wong, M.H. Application of Spirulina in aquaculture: A review on wastewater treatment and fish growth. Rev. Aquac. 2020, 12, 582–599. [Google Scholar] [CrossRef]
- Christaki, E.; Florou-Paneri, P.; Bonos, E. Microalgae: A novel ingredient in nutrition. Int. J. Food Sci. Nutr. 2011, 62, 794–799. [Google Scholar] [CrossRef]
- Priyadarshani, I.; Rath, B. Commercial and industrial applications of micro algae—A review. J. Algal Biomass Util. 2012, 3, 89–100. [Google Scholar]
- Brundu, G.; Cannavacciuolo, A.; Nannini, M.; Somma, E.; Munari, M.; Zupo, V.; Farina, S. Development of an efficient, noninvasive method for identifying gender year-round in the sea urchin Paracentrotus lividus. Aquaculture 2022, 564, 739082. [Google Scholar] [CrossRef]
- Varrella, S.; Romano, G.; Ianora, A.; Bentley, M.G.; Ruocco, N.; Costantini, M. Molecular response to toxic diatom-derived aldehydes in the sea urchin Paracentrotus lividus. Mar. Drugs 2014, 12, 2089–2113. [Google Scholar] [CrossRef] [PubMed]
- Federico, S.; Glaviano, F.; Esposito, R.; Pinto, B.; Gharbi, M.; Di Cosmo, A.; Costantini, M.; Zupo, V. Gene expression detects the factors influencing the reproductive success and the survival rates of Paracentrotus lividus offspring. Int. J. Mol. Sci. 2022, 23, 12790. [Google Scholar] [CrossRef] [PubMed]
- Zupo, V.; Glaviano, F.; Caramiello, D.; Mutalipassi, M. Effect of five benthic diatoms on the survival and development of Paracentrotus lividus post-larvae in the laboratory. Aquaculture 2018, 495, 13–20. [Google Scholar] [CrossRef]
- Fernandez, C.; Boudouresque, C.F. Evaluating artificial diets for small Paracentrotus lividus (Echinodermata: Echinoidea). Echinoderms San Fr. 1998, 651–657. [Google Scholar]
- George, S.B.; Lawrence, J.M.; Lawrence, A.L. Complete larval development of the sea urchin Lytechinus variegatus fed an artificial feed. Aquaculture 2004, 242, 217–228. [Google Scholar] [CrossRef]
- Jangoux, M. Land-based, closed-cycle echiniculture of Paracentrotus lividus (Lamarck) (Echinoidea: Echinodermata): A long-term experiment at a pilot scale. J. Shellfish. Res. 1998, 17, 152–153. [Google Scholar]
- Solari, P.; Pasquini, V.; Secci, M.; Giglioli, A.; Crnjar, R.; Addis, P. Chemosensitivity in the sea urchin Paracentrotus lividus (Echinodermata: Echinoidea) to food-related compounds: An innovative behavioral bioassay. Front. Ecol. Evol. 2021, 9, 723. [Google Scholar] [CrossRef]
- Luis, O.J.; Gago, J.M. Perception of Dissolved Food-Related Compounds by the Sea Urchin Paracentrotus lividus (Echinodermata: Echinoidea). Front. Mar. Sci. 2021, 8, 719670. [Google Scholar] [CrossRef]
- Pearce, C.M.; Daggett, T.L.; Robinson, S.M. Effect of protein source ratio and protein concentration in prepared diets on gonad yield and quality of the green sea urchin, Strongylocentrotus droebachiensis. Aquaculture 2002, 214, 307–332. [Google Scholar] [CrossRef]
- Mos, B.; Cowden, K.L.; Nielsen, S.J.; Dworjanyn, S.A. Do Cues Matter? Highly inductive settlement cues don’t ensure high post-settlement survival in sea urchin aquaculture. PLoS ONE 2011, 6, e28054. [Google Scholar] [CrossRef]
- Keesing, J.K.; Hall, K.C. Review of harvests and status of world sea urchin fisheries points to opportunities for aquaculture. J. Shellfish. Res. 1998, 17, 1597–1604. [Google Scholar]
- Fernandez, C.; Pergent, G. Effect of different formulated diets and rearing conditions on growth parameters in the sea urchin Paracentrotus lividus. J. Shellfish. Res. 1998, 17, 1571–1581. [Google Scholar]
- Woods, C.; James, P.J.; Moss, G.A.; Wright, J.; Siikavuopio, S. A comparison of the effect of urchin size and diet on gonad yield and quality in the sea urchin Evechinus chloroticus Valenciennes. Aquac. Int. 2008, 16, 49–68. [Google Scholar] [CrossRef]
- Lawrence, J.M. Sea urchin roe cuisine. In Developments in Aquaculture and Fisheries Science; Elsevier: Amsterdam, The Netherlands, 2007; Volume 37, pp. 521–523. [Google Scholar]
- Zupo, V. The use of feeding indices for the study of food webs: An application to a “Posidonia oceanica” ecosystem. Coenoses 1993, 8, 85–95. [Google Scholar]
- Sartori, D.; Gaion, A. Can sea urchins benefit from an artificial diet? Physiological and histological assessment for echinoculture feasibility evaluation. Aquac. Nutr. 2016, 22, 1214–1221. [Google Scholar] [CrossRef]
- Tomsic, S.; Conides, A.J.; Anicic, I. Growth and Gonad Changes in Stony Sea Urchin, Paracentrotus lividus (Lamark, 1816) Fed Artificially Formulated Feed and Benthic Macrophyte Diet. Nase More 2015, 62, 85–90. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trial |
|---|
| Control Diet Agarose 3% Ullva rigida 4% |
| Diet A Agarose 3% Ulva rigida 2% SHG® microperle 0.3% Biomarin Algamac Enrich® plant dry mix 1% |
| Diet B Agarose 3% Spirulina 2% Ulva rigida 2% |
| Spirulina | SHG® Microperle | Biomarin Algamac Enrich® |
|---|---|---|
| Protein 60.0% | Protein 50.0% | Protein 32.5% |
| Fat 4.0% | Fat 15.0% | Fat 34.7% |
| Fiber 7.0% | Fibers 2.0% | Carbohydrates 17.6% |
| Ash 7.0% | Ashes 20.0% | Ash 10.8% |
| Carotenoids 3.0% | Other 13.0% | Moisture 4.4% |
| Dunnett’s Multiple Comparisons Test | Mean Diff. | 95.00% CI of Diff. | Below Threshold | Summary | Adjusted p Value |
|---|---|---|---|---|---|
| Control Diet vs. Diet A | 0.8571 | −0.5974 to 2.312 | No | Ns | 0.2797 |
| Control Diet vs. Diet B | −2.643 | −4.798 to −0.4874 | Yes | * | 0.0176 |
| Dunnett’s Multiple Comparisons Test | Mean Diff. | 95.00% CI of Diff. | Below Threshold | Summary | Adjusted p Value |
|---|---|---|---|---|---|
| Control Diet vs. Diet A | −0.4286 | −0.9292 to 0.07204 | No | Ns | 0.0949 |
| Control Diet 1 vs. Diet B | −0.75 | −1.195 to −0.3048 | Yes | ** | 0.0021 |
| p Value | p Value Summary | Are Means Significantly Different? |
|---|---|---|
| 0.0179 | * | Yes |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gharbi, M.; Glaviano, F.; Federico, S.; Pinto, B.; Cosmo, A.D.; Costantini, M.; Zupo, V. Scale-Up of an Aquaculture Plant for Reproduction and Conservation of the Sea Urchin Paracentrotus lividus: Development of Post-Larval Feeds. J. Mar. Sci. Eng. 2023, 11, 154. https://doi.org/10.3390/jmse11010154
Gharbi M, Glaviano F, Federico S, Pinto B, Cosmo AD, Costantini M, Zupo V. Scale-Up of an Aquaculture Plant for Reproduction and Conservation of the Sea Urchin Paracentrotus lividus: Development of Post-Larval Feeds. Journal of Marine Science and Engineering. 2023; 11(1):154. https://doi.org/10.3390/jmse11010154
Chicago/Turabian StyleGharbi, Maissa, Francesca Glaviano, Serena Federico, Bruno Pinto, Anna Di Cosmo, Maria Costantini, and Valerio Zupo. 2023. "Scale-Up of an Aquaculture Plant for Reproduction and Conservation of the Sea Urchin Paracentrotus lividus: Development of Post-Larval Feeds" Journal of Marine Science and Engineering 11, no. 1: 154. https://doi.org/10.3390/jmse11010154
APA StyleGharbi, M., Glaviano, F., Federico, S., Pinto, B., Cosmo, A. D., Costantini, M., & Zupo, V. (2023). Scale-Up of an Aquaculture Plant for Reproduction and Conservation of the Sea Urchin Paracentrotus lividus: Development of Post-Larval Feeds. Journal of Marine Science and Engineering, 11(1), 154. https://doi.org/10.3390/jmse11010154