Abstract
Proposed offshore windfarm sites could overlap with the brooding and spawning habitats of commercially important crustacea, including European lobster, Homarus gammarus and Edible crab, Cancer pagurus. Concerns have been raised on the biological effects of Electromagnetic Fields (EMFs) emitted from subsea power cables on the early life history of these species. In this study, ovigerous female H. gammarus and C. pagurus were exposed to static (Direct Current, DC) EMFs (2.8 mT) throughout embryonic development. Embryonic and larval parameters, deformities, and vertical swimming speed of freshly hatched stage I lobster and zoea I crab larvae were assessed. EMF did not alter embryonic development time, larval release time, or vertical swimming speed for either species. Chronic exposure to 2.8 mT EMF throughout embryonic development resulted in significant differences in stage-specific egg volume and resulted in stage I lobster and zoea I crab larvae exhibiting decreased carapace height, total length, and maximum eye diameter. An increased occurrence of larval deformities was observed in addition to reduced swimming test success rate amongst lobster larvae. These traits may ultimately affect larval mortality, recruitment and dispersal. This study increases our understanding on the effects of anthropogenic, static EMFs on crustacean developmental biology and suggests that EMF emissions from subsea power cables could have a measurable impact on the early life history and consequently the population dynamics of H. gammarus and C. pagurus.
1. Introduction
Marine species have adapted to naturally occurring magnetic and electric fields in the environment, with many having developed sensory abilities which enable them to detect and utilise these cues e.g.,: in geomagnetic navigation during migration [1]. With the expansion of marine renewable energy, the number of subsea power cables is rapidly increasing, introducing anthropogenic magnetic and electric fields to the marine environment. Marine Renewable Energy Devices (MREDs), are inter-connected via cable arrays through which electricity is transported to shore via High Voltage Direct Current (HVDC) export cables, following conversion of Alternating Current (AC) to Direct Current (DC). As with all current carrying cables, such power cables will emit static, electromagnetic fields of varying strengths. Whilst standard electric field leakage (E-field) can be successfully eliminated with insulation, there is no industry standard insulation at present to prevent magnetic field (B-field) leakage [2]. Consequently, a magnetic field leaking from individual cables will create an induced electric field (iE field) through the movement of charges in seawater, the strength of which will vary depend on current strength in cables, water current speed and directions and proximities of associated cables [3,4,5].
Previous studies have highlighted that there is insufficient in situ measurements to allow informed assessment of EMF emissions around MRED cable deployments [6]. EMF intensities can be modelled, but predicted values vary depending on cable properties, energy transmission, cable layout and local geomagnetic. Currents between 850 and 1600 Amperes (A) are commonly used in subsea power cables which would consequently produce an EMF of up to 3.20 millitesla (mT) (1600 A) on the cable surface in a perfect wire [7]. Previous studies have highlighted that there is insufficient in situ measurements to allow informed assessment of EMF emissions around MRED cable deployments [6]. However, results from a handful of studies, that used in situ methods to measure EMF report lower values on single cables [8,9,10]. Some windfarm projects use export corridors, with multiple HVDC laid in close proximity to each other. EMFs emitted from theses cables could potentially interact, resulting in higher EMF values than those obtained on single cables. EMF levels used in previous studies are highly variable, for example, 200 µT [11], 3.7 mT [3], 4.2 mT [12], 36 mT (DC) and 165 mT (AC) [13], 3 mT [14], 2.8 mT and 40 mT [15] levels have all been used to assess the biological effects of static EMFs.
Scientific data on EMF sensory abilities, thresholds and tolerance are only available for a few species. Anthropogenic EMFs, in contrast to natural magnetic fields, have increased biological activity, due to their ability to induce forced oscillations on critical, electrically charged or polar biomolecules [16]. Organisms’ defence mechanisms can mitigate the biological activity and potentially harmful effects of EMFs [17]. During early development, increased metabolic rates required for internal changes could cause these defence mechanisms to work less effectively and therefore early life stages could be more vulnerable to the effects of EMFs. There is scientific evidence, that magnetic fields of anthropogenic origin, can significantly decrease sperm motility of Mytilus galloprovincialis (Lmk.) [18]; delay onset of mitosis and cause developmental abnormalities in two species of sea urchins Lytechninus pictus (V.) and Strongylocentrotus purpurattus (S.) [19]; alter Xenopus laevis (D.) embryos cleavage planes [20]; increase egg-shell permeability of Atlantic salmon, Salmo salar (L.), Sea trout, Salmo trutta (L.), and Rainbow trout, Oncorhynchus mykiss (W.) [21]; accelerate rates of embryonic development of Daphnia magna (S.) [22] and delays the hatching period of zebrafish Danio rerio (H.) [23].
The effects of EMFs on crustacean physiology and behaviour, particularly their early life history is less understood. EMFs have been shown to affect the internal compass of Caribbean spiny lobster, Panulirus argus (L.) [24] and alter sheltering behaviour of Spinycheek crayfish, Orconectes limosus (R.) [25]. American lobster, Homarus americanus (H.M.E.) exhibited a greater range of large turns in their behaviour and had a significantly different spatial distribution in an EMF exposed enclosure [26]. When exposed to static magnetic fields, survival of North Sea prawn Crangon crangon (L.), round crab Rhithropanopeus harrisii (G.) and glacial relict isopod Saduria entomon (L.) were not affected [3]. Exposure to 0.23 mT had no adverse effects on juvenile European lobster Homarus gammarus behaviour [11]. Adult H. gammarus, and the Edible crab, Cancer pagurus have been shown to exhibit physiological and behavioural changes during exposure to elevated EMFs [15,27,28]. Whilst these studies reported reduced roaming activity in these species following EMF exposure, H. americanus has shown the opposite reaction to artificial EMFs with increased exploratory behaviour being exhibited [8]. Conversely, the impact on juvenile H. gammarus appears limited as EMF values up to 200 µT did not significantly impact daytime behaviour [11]. These findings highlight the potential variability in response to EMF between species and life stages.
H. gammarus and C. pagurus are high commercial value decapod crustaceans widely distributed throughout Western Europe [29,30,31]. Female H. gammarus can produce an egg mass of approximately 14,000 eggs, following mating in late summer to early autumn [32]. H. gammarus females carry eggs on their pleopods for a period of 9–11 months, sometimes longer [33]. Mature female C. pagurus have been shown to migrate considerable distances offshore to lay and incubate their eggs [34,35,36]. Once berried, females will spend 6–9 months half buried in pits they dig in fine substrate, with very little movement and significantly lower feeding rates [34,37,38,39].
Previous studies have suggested that both H. gammarus and C. pagurus, are found to inhabit areas around MREDs, particularly scour protection zones which can act as artificial reefs [40,41,42]. Both species have also exhibited attraction to artificial EMFs at predicted strengthsaround sub-sea power cables [27]. Given the reduction in movement of this species during the incubation period, combined with their attraction to artificial EMFs, it is likely that eggs could be exposed to elevated EMF levels throughout development.
The effects of anthropogenic EMF on early life history traits of both H. gammarus and C. pagurus are currently unknown. The purpose of this study is to determine whether chronic exposure to 2.8 mT EMF will affect embryonic development, egg parameters, embryonic development time, larval parameters and the occurrence of deformities. Potential effect of embryonic EMF exposure on stage I lobster and zoea I crab larvae fitness via vertical swimming test was also assessed. EMF values used in this study are environmentally relevant, correspond well with modelled values HVDC cables and are comparable to other studies.
2. Materials and Methods
2.1. Animal Collection and Husbandry
In autumn 2017, a total of 20 ovigerous female H. gammarus, and 25 adult female C. pagurus with sperm plugs were collected by local fishers in the Berwickshire Marine Reserve (St Abbs, Berwickshire, UK). All animals were immediately transported to holding tanks at St Abbs Marine Station. Lobsters were kept in 500–1000 L flow-through tanks with maximum stocking density of 3 lobsters per tanks. Female crabs were kept in large holding tanks (3000–10,000 L) with a maximum stocking density of 10 crabs per tank until experimentation began. All holding tanks were enriched with natural substrate (mixture of sand and small pebbles) and all animals were provided artificial shelters (ABS pipes). Tanks received a constant supply of raw, unfiltered, ambient temperature sea water and natural light conditions. Lobsters were fed live blue mussels twice a week. The crabs’ diet consisted of cooked mussels and raw squid and fish and were fed twice a week during holding and experimentation. Uneaten food was removed after 24 h from each tank.
Prior to experimentation all lobsters were categorised with condition index modified from [15], ensuring only those classed grades 1 or 2 were used for experimentation, with surplus lobsters being released. Lobsters utilised for experimentation were all above Minimum Landing Size (≥MLS), of comparable wet weight (584.71 ± 108.9 g) and carapace length, (92.33 ± 4.3 mm). Before experimentation, berried (ovigerous) females were visually inspected and only individuals with healthy eggs and similar egg mass size were used in this study. Experimentation was conducted between April 2018 and August 2018.
Crabs were visually inspected weekly for signs of egg laying. Once a female had finished laying eggs, their condition index was determined and only crabs with grades 1 or 2 were used for this study [15]. Female crabs’ carapace width, all above MLS (≥140 mm, 157.8 ± 8.8 mm) and wet weight (704.92 ± 211.5 g) were measured prior to the start of experimentation. Experiments took place between January 2018 and July 2018.
2.2. Experimental Setup
During experimentation, animals were placed in individual 30 L glass tanks, containing a black ABS (plastic) shelter, individual air stones and a sterilized pebble substrate (for lobsters) or fine sand (for crabs). All tanks were supplied with flow-through (flow rate 0.5 L min−1), raw, unfiltered seawater, at ambient temperature throughout the experimental period. Individual glass tanks were placed inside a 50 L black ABS water bath box allowing accurate temperature control over the experimental period and to limit stress caused by visual stimuli of conspecifics (Figure 1a). Each water bath was connected to a recirculating system consisting of a single 150 L sump tank and heater/chiller unit (Teco TK1000). Tank temperature (7.6–14.6 ± 0.1 °C), salinity (34 ± 0.20 ppt) and dissolved oxygen (>90%) were monitored throughout the experiment (YSI ProDSS multiprobe and HOBO UA-002-08 8k temp/light pendant). The transparent roof above the experimental arena ensured that all experimental animals received a natural photoperiod.
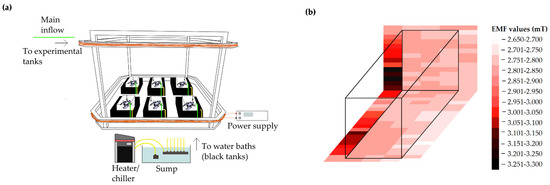
Figure 1.
Schematic illustrations of experimental set-up (a) Experimental arena and tanks within the Helmholtz coil; (b) Magnetic field distribution within an experimental tank. EMF generated was homogenous to ±5% of 2.8 mT within and between tanks.
An EMF was created using custom built Helmholtz coils. Coils were designed to create a uniform static (DC) electromagnetic field at 2.8 mT intensity in the 4 m3 experimental arena. This EMF strength was chosen based on modelled values expected around a single subsea power cable carrying current of I = 1600 A [7]. The EMF within the coil was mapped using a recently calibrated AlphaLab Inc, Gaussmeter Model GM-2 and was found to be spatially homogenous to approximately ±5% between experimental tanks placed in the test arena (Figure 1b). Helmholtz coils were powered using a variable DC power supply (Elektro-Automatik EA-PSI 8360-15) (Figure 1a).
In the EMF treatment, 6 berried female lobsters and 6 berried crabs were exposed to a constant uniform static DC EMF (2.8 mT) for the entire duration of egg incubation and larval release. In the control treatment, 6 berried female lobsters and 6 berried crabs were placed in an identical experimental arena described above, but the Helmholtz coils remained unpowered throughout the egg incubation and larval release period and only experienced the Earth’s local geomagnetic field, 49.96 µT total intensity (British Geological Survey, International Geomagnetic Reference Field (IGRF) Calculator).
2.3. Measured Parameters
2.3.1. Embryonic Development
Periodic sampling of lobster eggs (first two samples monthly, and weekly thereafter) took place throughout embryonic development. Female lobsters were removed from their tanks and a small portion of eggs were removed from the pleopods using a sterile spatula. Total egg sampling time was under 30 s to avoid inducing stress in the females. The average single egg weight was calculated based on the collective wet weight of 20 eggs. The eggs were analysed for each female under a microscope (Leica MZ125, camera Brunel Eyecam Plus) to determine the developmental percent-stages based on eye index [43], and to screen for deformities. Eggs were photographed and images were post-processed in ImageJ (Version 1.52A) [44]. Egg volume was calculated based on the formula of oblate spheroids: V = 1/6(πW2L), where V is the egg volume (mm3), W is the minimum diameter of the egg (mm) and L is the maximum diameter of the egg (mm) [45]. The same procedure described above for H. gammarus was utilised for egg sampling in C. pagurus. Crab eggs were sampled throughout embryonic development (first three sampling monthly thereafter every 10 days). Sampled eggs were photographed via a compound microscope (Leica DM750, camera DMC 5400), staged based on [46], and measured and screened for deformities. Egg volume was calculated using the same formula as described above.
Crabs were placed into the experimental tanks once they had finished laying eggs which varied between individuals by up to 4 weeks. All crabs used in experiments began trials in January 2018, with eggs in identical developmental stages (stage 1). In order to standardise the egg development between berried lobsters, experimentation began in April 2018, when all lobster eggs were at Stage 50.
2.3.2. Hatching
Individual hatching dates were recorded for each female lobster and crab. After hatching started each tank was put on a temperature-controlled recirculating system, with daily water change to avoid larval loss. As hatching occurs mainly at night, all freshly hatched stage I lobster larvae [47], and stage I zoea crab [48] larvae were collected at 09:00 am every day and counted throughout the hatching period [49]. Due to the large number of C. pagurus larvae hatching daily, 5 subsamples of larvae were carefully collected in a filter to avoid damage and counted to estimate density and subsequently the total number of larvae released. The larval release duration was recorded for each lobster and crab.
2.3.3. Larval Assessment
A total of 100 freshly hatched larvae were randomly selected from each female and were individually photographed under a dissection microscope (Leica MZ125, camera DMC 5400). Total length, carapace height, carapace length, and maximum eye diameter were measured in lobster larvae, while dorsal and rostral spine length, carapace height and length, total length and maximum eye diameter were measured on crab larvae using ImageJ (Version 1.52A) image analysing software. A total of 2651 EMF exposed and 3573 control stage I lobster larvae were screened for deformities such as curled or bent rostrum, swollen carapace, twisted abdomen, damage of tail fan, twisted pereopods, misshapen claws and chromatic aberrations in the eye [50]. In total, 2703 freshly hatched, EMF exposed larvae and 3701 control crab larvae were screened for the same deformities as above with the addition of rostral, dorsal, and lateral spine deformation.
2.3.4. Vertical Swimming Speed
Positive phototaxis is exhibited by freshy hatched lobster and crab larvae [51,52]. Larvae from each female were collected and were subjected to experiments to determine vertical swimming speed based on the method of Schmalenbach and Buchholz [53]. Before sunrise larvae were collected and transferred to a separate acclimating holding tank with ambient salinity, temperature, aeration and kept in a temperature controlled dark room prior to the trial. The experimental chamber consisted of 75 mL (lobster larvae) and 15 mL (crab larvae) black glass vertical cylinders with a clear window at the base and the top. The cylinders were filled with UV-sterilised and filtered (0.22 µm) seawater at the same temperature as the holding tank. The bottom window of the cylinder was lit using a handheld torch and the time taken for the individual larvae to sink down (sinking rate) was measured (Figure 2a). This light was then switched off and the light at the top of the cylinder was switched on. The time was measured for the individual larvae to reach the top of the cylinder (Figure 2b). The vertical swimming speed was calculated by the difference between the measured upward swimming speed and sinking rate in cm s−1 and mm s−1. Larvae were only used once, regardless of trial success. Trials where the larvae did not swim up after 5 min were classed as a failed trial.
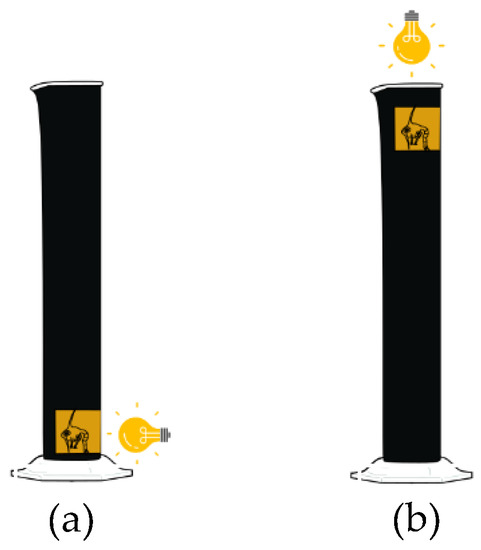
Figure 2.
Schematic illustration of vertical swimming test. (a) Sinking rates of freshly hatched lobster and crab larvae were determined by lighting up the window near the bottom of the test cylinder and measuring the time it took for the larvae to sink down; (b) Then the top window was lit and the time it took for the larvae to appear was measured to calculate vertical swimming speed.
2.4. Statistical Analysis
Data is presented as the mean and standard error (SEM) of replicates. The statistical analysis was performed with the computer programs SPSS (IBM SPSS Statistics v.23 SPSS Corp. Chicago, IL, USA) and STATISTICA version 7.1 (StatSoft Inc., Tulsa, OK, USA, 2005) with visualisation produced with R 4.1.2 [54,55]. The data were tested for parametric assumption by Kolmogorov-Smirnov test and Levene’s test. When data met these assumptions, they were subjected to one-way or two-way analysis of variance (ANOVA), multivariate analysis of variance (MANOVA) followed by post-hoc analysis via Tukey’s test. If data did not meet parametric assumptions, non-parametrical analysis of Mann-Whitney U-test was performed with Bonferroni correction where appropriate. To test difference of prevalence of larval deformities and mortality between treatments, pairwise comparisons by Chi-squared tests with Yates correction were performed. All statistics were tested at a probability of 0.05. Statistical differences of data sets are indicated on the presented graphs by single * (p < 0.05), double ** (p < 0.01) or triple *** (p < 0.001) asterisks.
3. Results
3.1. Embryonic Development
3.1.1. Homarus Gammarus
The total time of full embryonic development, from 50% development to hatching did not differ significantly between EMF exposed and control lobsters. The mean time taken from 50% to date of first hatch under control conditions was 82.8 ± 9.8 days (N = 5), whilst during EMF exposure the time taken to first hatch was 69.7 ± 7.3 days (N = 6) (Table 1). During experimentation one control lobster’s eggs reached 80% development then stopped advancing and were subsequently dropped by the female, therefore this data was omitted from further statistical analysis.

Table 1.
Embryonic development time (days) for each lobster and crab under control and EMF exposed conditions. H. gammarus N = 11, C. pagurus N = 11.
Egg weight between 50–100% development showed no significant differences between control and EMF exposed eggs. Egg weight increased from 4.05 ± 0.1 mg at 60% to 5.71 ± 0.25 mg at 100% with no significant deviation between treatments. Throughout the egg development trials, no embryonic deformations were observed.
In both treatments, egg volume increased by approx. 2 mm3 from experimental start (50%) to hatching (Figure 3). At 60% development, the volume of EMF exposed eggs was significantly greater (6.37%, t(514) = −5.87, p < 0.05, t-test) than control eggs (6.37%, t(514) = −5.87, p < 0.05, t-test). At 70% there were no differences in egg volume between the two treatments. At 80% and 90% development the egg volume of EMF exposed eggs stayed significantly under control volumes (13.5% and 15.5% respectively t(295) = 4.32, p < 0.05, t(39) = 4.90, p < 0.01, t-test). At 100% development, control and EMF exposed egg volumes showed no significant differences.
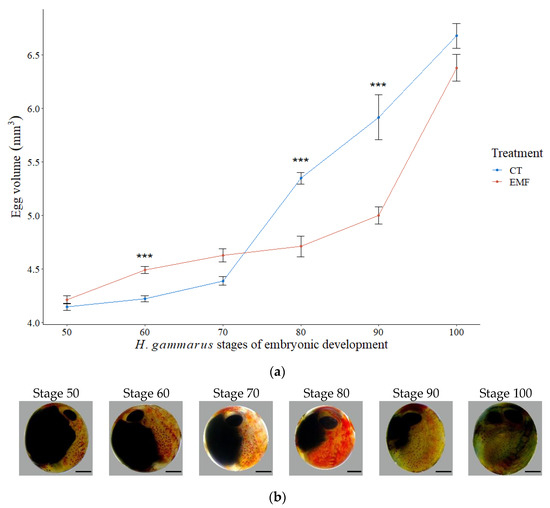
Figure 3.
Embryonic development of H. gammarus. (a) Egg volume (mm3) throughout development (50% to 100% development scale) in EMF exposed (red) and control groups (blue). Markers indicate mean values, vertical bars denote +/− standard error of means. Significance levels between two treatments at each stage indicated by asterisk, *** is the significance at p < 0.001. N = 2560; (b) Micrographs of H. gammarus embryonic stages, from stage 50 until pre-hatching (stage 100). Images were taken of unstained, unfixed specimens. Photo: St Abbs Marine Station. (Scale bar = 0.5 mm).
3.1.2. Cancer Pagurus
During experimentation eggs from one control crab reached stage 2 then stopped advancing, followed by the death of the female, therefore data obtained from this crab was omitted from statistical analysis. The total time of embryonic development, from stage 1 to hatching did not differ significantly between EMF exposed (124 ± 7 days) and control crabs (113 ± 5 days) (Table 1). Egg volume significantly increased throughout embryonic development by an average of 0.025 µm3 from stage 1 to stage 6 in both treatments (Figure 4) (F(5,1915) = 646.02, p < 0.001, one-way ANOVA). Between stage 2 and 4 egg volumes were significantly smaller in EMF exposed crabs compared to control.At stage 5 there were no differences between the two treatments, and stage 6 EMF exposed crab’s egg volume was significantly increased compared to control (F(1,438) = 19.39, p < 0.001, 2- way ANOVA, Tukey’s posthoc test).
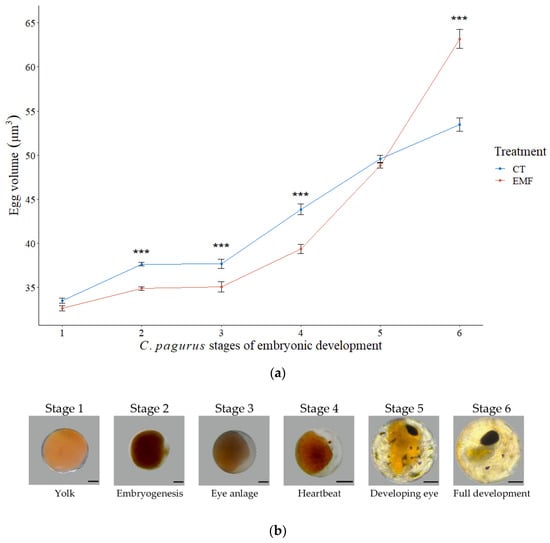
Figure 4.
Embryonic development C. pagurus. (a) Egg volume (µm3) throughout embryonic development (stage 1 to stage 6) in EMF exposed (red) and control groups (blue). Markers indicate mean values, vertical bars denote +/− standard error of means. Significance levels between two treatments at each stage indicated by asterisk, *** is the significance at p < 0.001. N = 1920; (b) Micrographs of C. pagurus embryonic stages from freshly laid eggs (stage 1) until pre-hatching (stage 6). Images were taken of unstained, unfixed specimens. Photos: St Abbs Marine Station. (Scale bar = 0.1 mm).
3.2. Hatching
3.2.1. Homarus Gammarus
The total number of hatched larvae per lobster ranged from 136–3812 under control conditions and 76–5062 under EMF conditions. Under control conditions a total of 3 (60%) lobsters successfully released over 2000 larvae whilst only 2 (33%) EMF exposed lobsters had a success rate of over 2000.
The mean total number of days taken from first hatch to final release of larvae was slightly higher for EMF exposed lobsters (18.2 ± 1.1 days) compared to under control conditions (14.4 ± 1.4 days), although this difference was not statistically significant.
3.2.2. Cancer Pagurus
The total larvae hatched ranged from 154,368 to 1,672,020 larvae under control conditions and between 94,985 and 673,670 larvae in EMF exposed crabs. The lower number of hatched larvae in EMF exposed crabs was not statistically significant. Under control conditions 50% of crabs successfully released over 500,000 larvae whilst only 20% of EMF exposed crabs had a similar success rate.
There were no significant differences between the mean total number of days taken from first hatch to final release of larvae in EMF exposed crabs (8.8 ± 1.4 days), compared to crabs kept in control conditions (8.3 ± 1.2 days).
3.3. Larval Assessment
3.3.1. Homarus Gammarus
Deformities were significantly higher in EMF exposed larvae compared to control (p < 0.001, Chi-square with Yates correction). Of the 2651 EMF exposed larvae assessed, 3% showed carapace, abdominal, rostral, and tail fan deformations, compared to 1% of the 3573 larvae assessed under control conditions (Figure 5). Mortality rate was significantly lower in EMF exposed larvae (0.01%), compared to the control group (0.24%) (p < 0.01, Chi-square with Yates correction).

Figure 5.
Deformities of H. gammarus larvae after exposure to 2.8 mT EMF throughout embryonic development. (a) Deformed rostrum; (b) Curled, puffy carapace; (c) Deformed chelae; (d) Chromatic aberration; (e) Abdominal and tail fan deformation. Photos: St Abbs Marine Station. (Deformities indicated by arrows, Scale bar = 1 mm).
Larvae exposed to EMF throughout their embryonic development had a significantly smaller total length (TL), carapace height (CH) and maximum eye diameter (ED) and a significantly longer carapace length (CL) (F(1,6222) = 3.9, 100.9, 118.98, 63.75, p < 0.001, MANOVA) (Figure 6a,c,e).
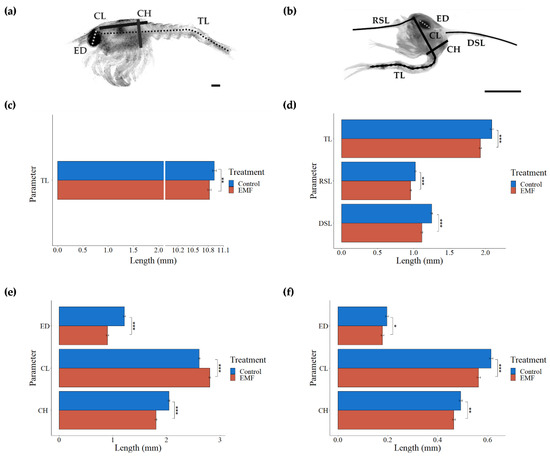
Figure 6.
Larval measurements (a) Stage I H. gammarus larvae length measurements: total length (TL), carapace length (CL), carapace height (CH), maximum eye diameter (ED) (b) Zoea I C. pagurus larvae length measurements: total length (TL), carapace length (CL), carapace height (CH), maximum eye diameter (ED), rostral spine length (RSL), dorsal spine length (DSL) Bar graphs show differences of freshly hatched H. gammarus larvae (c,e) and C. pagurus larvae (d,f) larval measurements between control and EMF exposed groups Significance levels indicated by * p < 0.05, ** p < 0.01, and *** p < 0.001. Photos: St Abbs Marine Station. (Scale bar = 0.5 mm).
3.3.2. Cancer Pagurus
A small percentage (<1%) of larval deformities were observed under both experimental conditions and did not differ significantly. The mortality rate of freshly hatched larvae was not affected by EMF exposure with 11.1% in EMF exposed crabs and 14.3% in control conditions.
EMF exposed larvae were significantly smaller in all the measured parameters compared to larvae from control treatment (RSL (F(1,6402) = 19.38, DSL F(1,6402) = 52.45, CH F(1,6402) = 9.9, CL F(1,6402) = 32.94, TL F(1,6402) = 42.05, p < 0.001, ED F(1,6402) = 42.05, p < 0.05, MANOVA) (Figure 6b,d,f).
3.4. Vertical Swimming Speed
3.4.1. Homarus Gammarus
A total of 151 EMF exposed larvae (and 106 control) were subjected to vertical swimming speed tests. There were significantly more failed trials (larvae failed to reach the top of the test chamber after 5 min) in the EMF exposed larvae (11%) than those reared under control conditions (4%) (p < 0.05, Chi-square with Yates correction). Swimming speed of successful trials, ranged from 0.23 to 18.9 cm s−1 in EMF larvae and 0.13 to 19.6 cm s−1 in control larvae. The mean vertical swimming speeds did not differ significantly between EMF exposed larvae (2.22 ± 0.3 cm s−1) and the control group (2.39 ± 0.4 cm s−1).
3.4.2. Cancer Pagurus
A total of 60 EMF exposed, and 129 control larvae were used for vertical swimming speed tests. The percentage of failed trials between EMF (55%) and control groups (64%) showed no significant difference. Swimming speed of freshly hatched larvae with an average of 3.3 ± 0.5 mm s−1 in the EMF exposed group and 1.6 ± 0.4 mm s−1 in the control group was not significantly affected by embryonic EMF exposure.
4. Discussion
This study has shown that anthropogenic EMFs can cause alteration in egg and larval parameters. Differences found in these traits suggests that artificial EMFs at 2.8 mT strength could have a measurable impact on early development of two commercially important crustaceans. Exposure to static 2.8 mT EMFs throughout embryonic development significantly altered egg volume and resulted in smaller larvae across most measured parameters in both H. gammarus and C. pagurus. A lower mortality rate, but higher number of deformities and a decrease in larval swimming performance of lobster larvae was also observed.
The average time to hatching, measured in number of days from 50% development in lobsters (82.8 ± 9.8 days in EMF, 69.7 ± 7.3 days in control) and number of days from stage 1 in crabs (124 ± 7 days EMF, 113 ± 5 days in control) coincides well with those found in literature. Wear found that C. pagurus embryonic development was approximately 80 days from end of diapause, which can be up to 60 days in duration [56]. Our results for H. gammarus aligned with previous published results which have reported 60–80 days for the average embryonic development time from 50% development, and 7 to 10 days hatching duration [57]. Our results showed slightly longer hatching duration of H. gammarus (18.2 ± 1.1 days in EMF, 14.4 ± 1.4 days in control). Hatching in C. pagurus in this study is in agreement with previous findings (8.8 ± 1.4 days EMF, 8.3 ± 1.2 days control) [37]. The timing of hatching may be critical to survival of larvae, as a mismatch between hatching time and food availability is a possible source of mortality [58]. Although mean time to hatching was slower in EMF exposed groups in both species, due to high individual variation between females these differences were not significantly different. To ensure these crucial parameters are unaffected by anthropogenic EMF exposure, it is important to re-assess these results in the future using larger numbers of replicates.
EMF exposure in this study did not affect the length of embryonic development. Similar results were reported in rainbow trout, Oncorhynchus mykiss, exposed to 10 mT static magnetic field [59], whereas, delays have been reported in sea urchin eggs [60] and in Japanese rice fish, Oryzias latipes (T. and S.) embryos exposed to a 60 Hz (AC) EMF [61]. The opposite result of accelerated hatching times was observed for Northern pike, Esox Lucius (L.), embryos developing in a 10 mT magnetic field [62]. The diverse findings in literature highlight the different sensitivity of species to EMF exposure. The highly variable nature of embryonic development time and hatching in individual females makes it difficult to standardise and may have contributed to differences observed in the results of this study. Further studies are required with a greater number of replicates to develop a full understanding of the effects of static EMFs on the egg development timescales of these species.
Stage specific differences of egg volumes were observed between treatments (control and 2.8 mT EMF exposure throughout egg incubation) in both H. gammarus and C. pagurus. Lobster egg volumes at stage 80 and 90 and crab egg volumes between stage 2 and 4 were significantly smaller, than control eggs, potentially indicating disruption of water and/or salt uptake. These differences in egg size however diminished prior to hatching in lobsters and EMF exposed crab eggs even exceeded the size of those kept in control conditions. Studies have shown that anthropogenic EMFs due to its polarized nature can create ionic forced oscillations in cells, causing irregular gating of membrane ion channels, which are responsible for maintaining cell osmosis. [16,63]. The disruption of membrane permeability could account for changes found in EMF exposed egg volumes in both species.
The eggs of marine invertebrates increase their volume significantly during embryonic development by a slow but steady osmotic uptake of water [64]. The egg volume of H. gammarus and C. pagurus in this study increased by approximately 50% during embryonic development which corresponds well with results found in the literature [43,56]. It has been reported that the rate of salt and water uptake drastically increases around the time the embryonic heartbeat is first observable, as the volume of the eggs increases more rapidly from this time [56]. As this increased water uptake has a crucial role in osmotic hatching, adjusting the larval body temperature and floating the hatched larva by reducing its specific gravity, disruption of this process could negatively affect hatching time and larval survival [65].
EMF exposure resulted in significant differences of all measured larval parameters. Stage I lobster larvae from EMF exposed females showed smaller total length, carapace height and eye diameter, but a longer carapace length. EMF exposed crab larvae had a decreased rostral and dorsal spine length, carapace length, total length, and maximum eye diameter. Smaller larval size could be attributed to disruption in metabolism, potentially through reduced yolk consumption and alteration in egg membrane gas and metabolite exchange. Larval size has been shown to be affected by several environmental factors such as salinity, temperature, and photoperiod [66,67]. Decreased carapace length and prolonged moulting cycle of larvae was found with increased pCO2 levels in the closely related H. americanus [68]. Previous studies on the effect of EMFs on hatching size show varying results, such as decreased weight of mice pups, increased size of chickens or no difference in American kestrel, Falco sparverius (L.), chicks [69,70,71]. Differences in literature could be attributed to exposure to different level and type of EMFs and species-specific sensitivity. To understand the underlying mechanism, anthropogenic EMF on metabolism and energetics of early life stages, it is important in the future to assess its effects on yolk consumption and composition, and egg membrane properties.
Whilst deformities of hatched larvae occurred in both species, a significantly higher rate of deformities was observed in H. gammarus exposed to EMF (3% developed deformities), but not C. pagurus. The mortality rate of zoea I crab larvae was also not affected by EMF exposure, whilst EMF exposed stage I lobster larvae showed a significantly lower mortality rate when compared to the control group. Deformities in marine invertebrate larvae have been reported to affect the mortality and egg production of individual adults [50]. The most frequent deformities in the stage I lobster larvae in EMF treatment were found in the carapace, tail fan and/or in the rostrum in this study. Similar deformities of embryonic and larval stages in other marine invertebrate species have been found, when exposed to elevated pCO2 levels [72,73,74]. Deformities generally affect carapace, walking legs, claws, abdomen, tail fan and even antenna with those affected usually developing multiple deformities [50]. Agnalt et al. observed that deformed tail fans of lobster larvae could not be repaired through subsequent moulting [50]. EMF has also been reported to have negative effects on ontogenesis in other species. Increased mutation rates, reduced pigmentation and axial anomalies have been reported in fruit flies, Drosophila melanogaster (M.) and African clawed toad, Xenopus laevis exposed to magnetic fields (2.5 mT and 1 T respectively) [75,76]. The ratio of lobster larvae with deformed exoskeletons indicates adverse effect of exposure to 2.8 mT EMF in H. gammarus. Some of the deformities may affect respiration (carapace), swimming ability (tail-fan damage) and consequently larval dispersal.
Stage I lobster larvae, exposed to 2.8 mT EMF during development were found to fail significantly more vertical swimming trials, with a total of 7% more failed trials. This could be due to a reduction in positive phototactic behaviour or a reduction in the ability to swim. However, EMF exposure did not affect the success of vertical swimming tests in C. pagurus zoea I larvae, nor the vertical swimming speed of C. pagurus or the successful H. gammarus larvae. The average vertical swimming speeds found in this study in both H. gammarus (2.39 ± 0.4 cm s−1) and C. pagurus (1.6 ± 0.4 mm s−1) correspond with values in the literature: 2.9 ± 0.5 cm s−1 in H. gammarus larvae; 1.79 cm s−1 in squat lobster, Galathea dispersa; and 0.15 ± 0.008 mm s−1 in Brachionus calyciflorus (p.) [54,77,78]. Newly released larvae show a strong positive phototaxis resulting in a preference for the uppermost water layers with high food abundance and surface currents, that carry the larvae to suitable areas of settlement [77,79]. Larval swimming ability plays an important role in maintaining position within currents [78]. When larvae stop swimming by beating of the exopodites, they sink towards the bottom, where currents are reduced [47]. Acute exposure to EMF has been observed to disrupt orientation and locomotory behaviour in D. rerio, D. melanogaster, Talorchestia martensii (W.) and H. americanus [26,80,81,82]. This study concludes that long-term exposure to EMF during embryonic development can affect phototactic behaviour and swimming ability of lobster larvae, which could adversely affect larval recruitment processes. Differences found in the vertical swimming test failure ratio of EMF treated H. gammarus could also be accounted for by the elevated prevalence of lobster larval deformities.
EMF sensitivity thresholds and tolerance can be species and life stage specific. Early results from EMF work have highlighted that closely related crustacean species may react differently to EMFs, with H. gammarus exhibiting an increased amount of time spent in EMF-exposed shelters [15] whilst H. americanus have shown increased exploratory behaviours around subsea cables [8], and juvenile lobsters showed no difference in shelter finding and exploratory behaviour [11]. Although different experimental design and EMF intensities preclude the ability for a direct comparison between these papers, it is important that sensory thresholds and tolerance are identified in specific species and not generalized across related taxa; a concern previously highlighted in elasmobranch EMF research [4]. Whilst some effects of anthropogenic EMFs may be deleterious to certain species and life stages (such as those in the present study), it is important to note that it may be benign or perhaps even beneficial to others. Field-based studies have demonstrated that the catchability of Dungeness crab, Metacarcinus magister (D.), was not affected by the presence of an energised subsea cable [83], and the responses of two species of rock crab, Metacarcinus anthonyi (R.) and Cancer productus (R.), did not differ when placed in boxes along energised versus non-energised subsea cables [84]. As such, it is necessary to understand if and how different organisms and life stages may be impacted by anthropogenic EMFs on a species-level scale.
It is important to consider how anthropogenic EMF transmitted to heterogenous marine environments will interact with the local geomagnetic field and how this may change the sensory environment and EMF reception of different species. To assess the impacts of MREDs on marine benthic fauna, accurate in situ measurements of EMF levels around different sites are imperative. Cumulative effects of MRED related stressors on marine invertebrates, such as current alterations around wind turbines, increased sea temperature around cables and substations, and operational underwater sound must be investigated in the future on all life stages of inhabiting organisms. The effect of temperature is particularly important, as ovigerous female lobsters move offshore to reduce thermal variability during nursing periods [85]. Previous studies showed, that ovigerous C. pagurus will likely be attracted to EMF emitted from subsea power cables [15]. Further studies on gene expression, larval physiology, growth, dispersal, settlement, survival and feeding rate are needed to fully understand the impact of marine renewable energy devices on the early life history of H. gammarus and C. pagurus.
5. Conclusions
The data obtained from this study provides further evidence on the impacts of anthropogenic static electromagnetic fields on early development of marine invertebrates. Our study demonstrates that 2.8 mT EMF exposure during egg development caused stage and species-specific alteration of egg volumes, likely to be attributed to irregular gating of membrane ion channels. We observed significantly smaller larval parameters in both EMF treated species and a higher occurrence of larval deformities and lower swimming test success rate amongst lobster larvae. These results suggest that EMF may negatively impact larval mortality, recruitment and dispersal. In contrast, EMF exposure did not appear to impact embryonic development time, larval release time, or vertical swimming speed for either species, or crab larval deformities and swimming test success. Differences found in this study on the effects of EMF on early development of H. gammarus and C. pagurus highlight the importance of species-specific sensitivity to anthropogenic EMFs.
As reproduction and early life stages represent a bottleneck of crustacean population, the vulnerability of such stages to anthropogenic EMFs expected around subsea power cables demonstrated in this study suggest that marine renewable energy developments could have a considerable impact on shellfish fisheries. To fully understand the population-level impacts of MREDs, further studies are required to assess the biological effects of EMFs along with other stressors and environmental changes expected around these sites on all life stages of H. gammarus and C. pagurus.
Author Contributions
Conceptualization, P.H., K.S. and A.R.L.; methodology, P.H. and K.S.; validation, P.H., K.S.; B.A.A.E. and G.d.l.C.O.; formal analysis, P.H.; investigation, P.H. and K.S.; B.A.A.E., G.d.l.C.O., E.C.N.C., A.J.R.P. and C.M.V.R.; resources, P.H. and K.S.; data curation, P.H., K.S.; B.A.A.E. and G.d.l.C.O.; writing—original draft preparation, P.H., K.S., A.R.L., B.A.A.E. and G.d.l.C.O.; writing—review and editing, P.H., K.S., B.A.A.E., E.C.N.C., A.J.R.P. and C.M.V.R.; visualization, P.H., A.J.R.P. and C.M.V.R.; supervision, A.R.L.; project administration, P.H. and K.S.; funding acquisition, P.H. and K.S. All authors have read and agreed to the published version of the manuscript.
Funding
All work was carried out at St Abbs Marine Station with funding obtained from the Scottish Fishermen’s Federation (via Scottish Fishermen’s Trust) and the Nesbitt-Cleland Trust.
Institutional Review Board Statement
The research adhered to the legal requirements of the country (U.K.) in which the work was carried out, and all institutional guidelines of St Abbs Marine Station and Heriot-Watt University. Informal ethical review and approval of this research was conducted by authors and trustees of St Abbs Marine Station using published literature and information obtained from Animals (Scientific Procedures) Act 1986 (ASPA). Animal collection was conducted with the local commercial fishing fleet and did not require collection permits.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data presented in this article are available on request from the corresponding author.
Acknowledgments
We would like to express our sincere gratitude to all our volunteers helping with sample collection and microscopic image analysis. We are grateful for anonymous reviewers for providing constructive comments that improved this manuscript. We would like to extend our appreciation to Adam Houghton for providing animal care and technical support. We are thankful for all experimental animals provided by fishers in St Abbs.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Kirschvink, J.L. Homing in on vertebrates. Nature 1997, 390, 339–340. [Google Scholar] [CrossRef]
- Gill, A.B. Offshore renewable energy: Ecological implications of generating electricity in the coastal zone. J. Appl. Ecol. 2005, 42, 605–615. [Google Scholar] [CrossRef]
- Bochert, R.; Zettler, M.L. Long-term exposure of several marine benthic animals to static magnetic fields. Bioelectromagnetics 2004, 25, 498–502. [Google Scholar] [CrossRef] [PubMed]
- Tricas, T.; Gill, A.B. Effects of EMFs from Undersea Power Cables on Elasmobranchs and Other Marine Species; OCS Study BOEMRE 2011-09; U.S. Department of the Interior: Washington, DC, USA, 2011. [Google Scholar]
- Taormina, B.; Bald, J.; Want, A.; Thouzeau, G.; Lejart, M.; Desroy, N.; Carlier, A. A review of potential impacts of submarine power cables on the marine environment: Knowledge gaps, recommendations and future directions. Renew. Sustain. Energy Rev. 2018, 96, 380–391. [Google Scholar] [CrossRef]
- Centre for Marine and Coastal Studies. A Baseline Assessment of Electromagnetic Fields Generated by Offshore Windarm Cables; University of Liverpool: Birkenhead, UK, 2003; p. 71. [Google Scholar]
- Bochert, R.; Zettler, M.L. Effect of Electromagnetic Fields on Marine Organisms. In Offshore Wind Energy: Research on Environmental Impacts; Köller, J., Köppel, J., Peters, W., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 223–234. [Google Scholar]
- Hutchison, Z.L.; Gill, A.B.; Sigray, P.; He, H.; King, J.W. Anthropogenic electromagnetic fields (EMF) influence the behaviour of bottom-dwelling marine species. Sci. Rep. 2020, 10, 4219. [Google Scholar] [CrossRef]
- Kavet, R.; Wyman, M.T.; Klimley, A.P. Modeling Magnetic Fields from a DC Power Cable Buried Beneath San Francisco Bay Based on Empirical Measurements. PLoS ONE 2016, 11, e0148543. [Google Scholar] [CrossRef]
- Otremba, Z.; Jakubowska, M.; Urban-Malinga, B.; Andrulewicz, E. Potential effects of electrical energy transmission–the case study from the Polish Marine Areas (southern Baltic Sea). Oceanol. Hydrobiol. Stud. 2019, 48, 196–208. [Google Scholar] [CrossRef]
- Taormina, B.; Di Poi, C.; Agnalt, A.-L.; Carlier, A.; Desroy, N.; Escobar-Lux, R.H.; D’eu, J.-F.; Freytet, F.; Durif, C.M.F. Impact of magnetic fields generated by AC/DC submarine power cables on the behavior of juvenile European lobster (Homarus gammarus). Aquat. Toxicol. 2020, 220, 105401. [Google Scholar] [CrossRef]
- Formicki, K.; Sadowski, M.; Tański, A.; Korzelecka-Orkisz, A.; Winnicki, A. Behaviour of trout (Salmo trutta L.) larvae and fry in a constant magnetic field. J. Appl. Ichthyol. 2004, 20, 290–294. [Google Scholar] [CrossRef]
- Cada, G.F.; Bevelhimer, M.S.; Riemer, K.P.; Turner, J.W. Effects on Freshwater Organisms of Magnetic Fields Associated with Hydrokinetic Turbines; Technical Report; U.S. Department of Energy: Washington, DC, USA, 2011. [Google Scholar]
- Woodruff, D.L.; Schultz, I.R.; Marshall, K.E.; Ward, J.A.; Cullinan, V.I. Effects of Electromagnetic Fields on Fish and Invertebrates; Pacific Northwest National Laboratory: Richland, WA, USA, 2012. [Google Scholar]
- Scott, K.; Harsanyi, P.; Lyndon, A.R. Understanding the effects of electromagnetic field emissions from Marine Renewable Energy Devices (MREDs) on the commercially important edible crab, Cancer pagurus (L.). Mar. Pollut. Bull. 2018, 131, 580–588. [Google Scholar] [CrossRef]
- Panagopoulos, D.J.; Johansson, O.; Carlo, G.L. Polarization: A Key Difference between Man-made and Natural Electromagnetic Fields, in regard to Biological Activity. Sci. Rep. 2015, 5, 14914. [Google Scholar] [CrossRef] [PubMed]
- Panagopoulos, D.J. Electromagnetic interaction between environmental fields and living systems determines health and well-being. In Electromagnetic Fields: Principles, Engineering Applications and Biophysical Effects; Kwang, M.-H., Yoon, S.-O., Eds.; Nova Science Publishers: New York, NY, USA, 2013; Volume 13. [Google Scholar]
- Pecoraro, R.; Pavone, S.C.; Scalisi, E.M.; Sica, C.; Ignoto, S.; Contino, M.; Salvaggio, A.; Sorbello, G.; Di Donato, L.; Brundo, M.V. Biological Effects of Non-ionizing Electromagnetic Fields to 27 GHz on Sperm Quality of Mytilus galloprovincialis. J. Mar. Sci. Eng. 2022, 10, 521. [Google Scholar] [CrossRef]
- Levin, M.; Ernst, S.G. Applied DC magnetic fields cause alterations in the time of cell divisions and developmental abnormalities in early sea urchin embryos. Bioelectromagnetics 1997, 18, 255–263. [Google Scholar] [CrossRef]
- Denegre James, M.; Valles James, M.; Lin, K.; Jordan, W.B.; Mowry Kimberly, L. Cleavage planes in frog eggs are altered by strong magnetic fields. Proc. Natl. Acad. Sci. USA 1998, 95, 14729–14732. [Google Scholar] [CrossRef]
- Sadowski, M.; Winnicki, A.; Formicki, K.; Sobocinski, A.; Tanski, A. The effect of magnetic field on permeability of egg shells of salmonid fishes. Acta Ichthyol. Piscat. 2007, 37, 129–135. [Google Scholar] [CrossRef]
- Krylov, V.V. Effects of electromagnetic fields on parthenogenic eggs of Daphnia magna Straus. Ecotoxicol. Environ. Saf. 2010, 73, 62–66. [Google Scholar] [CrossRef]
- Skauli, K.S.; Reitan, J.B.; Walther, B.T. Hatching in zebrafish (Danio rerio) embryos exposed to a 50 Hz magnetic field. Bioelectromagnetics 2000, 21, 407–410. [Google Scholar] [CrossRef]
- Ernst, D.A.; Lohmann, K.J. Effect of magnetic pulses on Caribbean spiny lobsters: Implications for magnetoreception. J. Exp. Biol. 2016, 219, 1827–1832. [Google Scholar] [CrossRef]
- Tański, A.; Formicki, K.; ŚMietana, P.; Sadowski, M.; Winnicki, A. Sheltering behaviour of Spinycheek crayfish (Orconectes limosus) in the presence of an artifical magnetic field. Bull. Fr. Pêche Piscic. 2005, 376–377, 787–793. [Google Scholar] [CrossRef]
- Hutchison, Z.; Sigray, P.; He, H.; Gill, A.; King, J.; Gibson, C. Electromagnetic Field (EMF) impacts on elasmobranch (shark, rays, and skates) and American lobster movement and migration from direct current cables. BOEM 2018, 3, 2018. [Google Scholar]
- Scott, K. Understanding the Biology of Two Commercially Important Crustaceans in Relation to Fisheries and Anthropogenic Impacts. Ph.D. Thesis, Heriot-Watt University, Edinburgh, UK, 2019. [Google Scholar]
- Scott, K.; Harsanyi, P.; Easton, B.A.A.; Piper, A.J.R.; Rochas, C.M.V.; Lyndon, A.R. Exposure to Electromagnetic Fields (EMF) from Submarine Power Cables Can Trigger Strength-Dependent Behavioural and Physiological Responses in Edible Crab, Cancer pagurus (L.). J. Mar. Sci. Eng. 2021, 9, 776. [Google Scholar] [CrossRef]
- Holthuis, L.B. Marine Lobsters of the World; Food and Agriculture Organization of the United Nations: Rome, Italy, 1991; Volume 13. [Google Scholar]
- Scott, K.; Harsanyi, P.; Lyndon, A.R. Baseline measurements of physiological and behavioural stress markers in the commercially important decapod Cancer pagurus (L.). J. Exp. Mar. Biol. Ecol. 2018, 507, 1–7. [Google Scholar] [CrossRef]
- Bridges, T. Crab and Lobster Stock Assessment. 2017. Available online: https://www.gov.uk/government/publications/crab-and-lobster-stock-assessment-2017 (accessed on 17 March 2022).
- Lizárraga-Cubedo, H.A.; Tuck, I.; Bailey, N.; Pierce, G.J.; Kinnear, J.A.M. Comparisons of size at maturity and fecundity of two Scottish populations of the European lobster, Homarus gammarus. Fish. Res. 2003, 65, 137–152. [Google Scholar] [CrossRef]
- Branford, J.R. Incubation period for the lobster Homarus gammarus at various temperatures. Mar. Biol. 1978, 47, 363–368. [Google Scholar] [CrossRef]
- Williamson, H.C. Contributions to the Life-Histories of the Edible Crab (Cancer Pagurus) and of Other Decapod Crustacea; Edinburgh, UK, 1904. [Google Scholar]
- Ungfors, A. Sexual maturity of the edible crab (Cancer pagurus) in the Skagerrak and the Kattegat, based on reproductive and morphometric characters. ICES J. Mar. Sci. 2007, 64, 318–327. [Google Scholar] [CrossRef][Green Version]
- Hunter, E.; Eaton, D.; Stewart, C.; Lawler, A.; Smith, M.T. Edible Crabs “Go West”: Migrations and Incubation Cycle of Cancer pagurus Revealed by Electronic Tags. PLoS ONE 2013, 8, e63991. [Google Scholar] [CrossRef] [PubMed]
- Edwards, E. The Edible Crab and Its Fishery in British Waters; Farnham (UK) Fishing News Books: Farnham, UK, 1979. [Google Scholar]
- Howard, A.E. The distribution and behaviour of ovigerous edible crabs (Cancer pagurus), and consequent sampling bias. ICES J. Mar. Sci. 1982, 40, 259–261. [Google Scholar] [CrossRef]
- Naylor, J.K.; Taylor, E.W.; Bennett, D.B. The oxygen uptake of ovigerous edible crabs (Cancer pagurus) (L.) and their eggs. Mar. Freshw. Behav. Physiol. 1997, 30, 29–44. [Google Scholar] [CrossRef]
- Langhamer, O.; Wilhelmsson, D. Colonisation of fish and crabs of wave energy foundations and the effects of manufactured holes—A field experiment. Mar. Environ. Res. 2009, 68, 151–157. [Google Scholar] [CrossRef]
- Hunter, W.R.; Sayer, M.D.J. The comparative effects of habitat complexity on faunal assemblages of northern temperate artificial and natural reefs. ICES J. Mar. Sci. 2009, 66, 691–698. [Google Scholar] [CrossRef]
- Hiscock, K.; Sharrock, S.; Highfield, J.; Snelling, D. Colonization of an artificial reef in south-west England—ex-HMS ‘Scylla’. J. Mar. Biol. Assoc. UK 2010, 90, 69–94. [Google Scholar] [CrossRef]
- Helluy, S.M.; Beltz, B.S. Embryonic Development of the American Lobster (Homarus americanus): Quantitative Staging and Characterization of an Embryonic Molt Cycle. Biol. Bull. 1991, 180, 355–371. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Lei, S.; Zhang, X.; Zhang, P.; Ikeda, Y. Biochemical composition of cuttlefish (Sepia esculenta) eggs during embryonic development. Molluscan Res. 2014, 34, 1–9. [Google Scholar] [CrossRef]
- Chung, J.S.; Webster, S.G. Expression and release patterns of neuropeptides during embryonic development and hatching of the green shore crab, Carcinus maenas. Development 2004, 131, 4751–4761. [Google Scholar] [CrossRef]
- Hadley, P.B. The behavior of the larval and adolescent stages of the American lobster (Homarus americanus). J. Comp. Neurol. Psychol. 1908, 18, 199–301. [Google Scholar] [CrossRef]
- Ingle, R. The larval and post-larval development of the edible crab, Cancer pagurus Linnaeus (Decapoda: Brachyura). Bull. Br. Mus. (Nat. Hist.) Zool. 1981, 40, 211–236. [Google Scholar]
- Ennis, G.P. Endogenous rhythmicity associated with larval hatching in the lobster Homarus gammarus. J. Mar. Biol. Assoc. United Kingd. 1973, 53, 531–538. [Google Scholar] [CrossRef]
- Agnalt, A.L.; Grefsrud, E.S.; Farestveit, E.; Larsen, M.; Keulder, F. Deformities in larvae and juvenile European lobster (Homarus gammarus) exposed to lower pH at two different temperatures. Biogeosciences 2013, 10, 7883–7895. [Google Scholar] [CrossRef]
- Botero, L.; Atema, J. Behavior and Substrate Selection During Larval Settling in the Lobster Homarus Americanus. J. Crustacean Biol. 1982, 2, 59–69. [Google Scholar] [CrossRef]
- Webley, J.A.C.; Connolly, R.M. Vertical movement of mud crab megalopae (Scylla serrata) in response to light: Doing it differently down under. J. Exp. Mar. Biol. Ecol. 2007, 341, 196–203. [Google Scholar] [CrossRef]
- Schmalenbach, I.; Buchholz, F. Vertical positioning and swimming performance of lobster larvae (Homarus gammarus) in an artificial water column at Helgoland, North Sea. Mar. Biol. Res. 2010, 6, 89–99. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing 4.0.3; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Yu, G.; Xu, S.; Chen, M.; Feng, T.; Zhan, L.; Zhou, L. Use ggbreak to effectively utilize plotting space to deal with large datasets and outliers. Front. Genet. 2021, 12, 2122. [Google Scholar] [CrossRef]
- Wear, R.G. Incubation in British Decapod Crustacea, and the Effects of Temperature on the Rate and Success of Embryonic Development. J. Mar. Biol. Assoc. UK 1974, 54, 745–762. [Google Scholar] [CrossRef]
- Charmantier, G.; Mounet-Guillaume, R. Temperature-specific rates of embryonic development of the European lobster Homarus gammarus (L.). J. Exp. Mar. Biol. Ecol. 1992, 160, 61–66. [Google Scholar] [CrossRef]
- Hjort, J. Fluctuations in the great fisheries of northern Europe viewed in the light of biological research. In Proceedings of the RAPPORTS ET PROCRS-VERBAUX, Copenhagen, Denmark, May 1914. [Google Scholar]
- Fey, D.P.; Jakubowska, M.; Greszkiewicz, M.; Andrulewicz, E.; Otremba, Z.; Urban-Malinga, B. Are magnetic and electromagnetic fields of anthropogenic origin potential threats to early life stages of fish? Aquat. Toxicol. 2019, 209, 150–158. [Google Scholar] [CrossRef]
- Zimmerman, S.; Zimmerman, A.M.; Winters, W.D.; Cameron, I.L. Influence of 60-Hz magnetic fields on sea urchin development. Bioelectromagnetics 1990, 11, 37–45. [Google Scholar] [CrossRef]
- Cameron, I.L.; Hunter, K.E.; Winters, W.D. Retardation of embryogenesis by extremely low frequency 60 Hz electromagnetic fields. Physiol. Chem. Phys. Med. NMR 1985, 17, 135–138. [Google Scholar]
- Fey, D.P.; Greszkiewicz, M.; Otremba, Z.; Andrulewicz, E. Effect of static magnetic field on the hatching success, growth, mortality, and yolk-sac absorption of larval Northern pike Esox lucius. Sci. Total Environ. 2019, 647, 1239–1244. [Google Scholar] [CrossRef]
- Petrov, E.; Martinac, B. Modulation of channel activity and gadolinium block of MscL by static magnetic fields. Eur. Biophys. J. 2006, 36, 95. [Google Scholar] [CrossRef]
- Davis, C.C. Mechanisms of hatching in aquatic invertebrate eggs. In Oceanography and Marine Biology: An Annual Review; Taylor & Francis: Aberdeen, Scotland, 1968; Volume 6, pp. 325–376. [Google Scholar]
- Pandian, T.J. Ecophysiological studies on the developing eggs and embryos of the European lobster Homarus gammarus. Mar. Biol. 1970, 5, 154–167. [Google Scholar] [CrossRef]
- Fonds, M. Laboratory Observations on the Influence of Temperature and Salinity on Development of the Eggs and Growth of the Larvae of Solea solea (Pisces). Mar. Ecol. Prog. Ser. 1979, 1, 91–99. [Google Scholar] [CrossRef]
- Gardner, C.; Maguire, G.B. Effect of photoperiod and light intensity on survival, development and cannibalism of larvae of the Australian giant crab Pseudocarcinus gigas (Lamarck). Aquaculture 1998, 165, 51–63. [Google Scholar] [CrossRef]
- Keppel, E.A.; Scrosati, R.A.; Courtenay, S.C. Ocean acidification decreases growth and development in American lobster (Homarus americanus) larvae. J. Northwest Atl. Fish. Sci. 2012, 44, 61–66. [Google Scholar] [CrossRef]
- Zahedi, Y.; Zaun, G.; Maderwald, S.; Orzada, S.; Pütter, C.; Scherag, A.; Winterhager, E.; Ladd, M.E.; Grümmer, R. Impact of repetitive exposure to strong static magnetic fields on pregnancy and embryonic development of mice. J. Magn. Reson. Imaging 2014, 39, 691–699. [Google Scholar] [CrossRef] [PubMed]
- Rooze, M.; Hinsenkamp, M. In vivo modifications induced by electromagnetic stimulation of chicken embryos. Reconstr. Surg. Traumatol. 1985, 19, 87–92. [Google Scholar]
- Fernie, K.J.; Bird, D.M.; Dawson, R.D.; Laguë, P.C. Effects of electromagnetic fields on the reproductive success of American kestrels. Physiol Biochem. Zool 2000, 73, 60–65. [Google Scholar] [CrossRef]
- Kurihara, H. Effects of CO2-driven ocean acidification on the early developmental stages of invertebrates. Mar. Ecol. Prog. Ser. 2008, 373, 275–284. [Google Scholar] [CrossRef]
- Kawaguchi, S.; Kurihara, H.; King, R.; Hale, L.; Berli, T.; Robinson, J.P.; Ishida, A.; Wakita, M.; Virtue, P.; Nicol, S. Will krill fare well under Southern Ocean acidification? Biol. Lett. 2011, 7, 288–291. [Google Scholar] [CrossRef]
- Byrne, M. Impact of ocean warming and ocean acidification on marine invertebrate life history stages: Vulnerabilities and potential for persistence in a changing ocean. In Oceanography and Marine Biology: Annual Review; Gibson, R.N., Atkinson, R.J.A., Gordon, J.D.M., Eds.; Taylor & Francis: London, UK, 2011; Volume 49, p. 42. [Google Scholar]
- Giorgi, G.; Guerra, D.; Pezzoli, C.; Cavicchi, S.; Bersani, F. Genetic effects of static magnetic fields. Body size increase and lethal mutations induced in populations of Drosophila melanogaster after chronic exposure. Genet. Sel. Evol. 1992, 24, 393–413. [Google Scholar] [CrossRef]
- Ueno, S.; Harada, K.; Shiokawa, K. The embryonic development of frogs under strong DC magnetic fields. IEEE Trans. Magn. 1984, 20, 1663–1665. [Google Scholar] [CrossRef]
- Nichols, J.H.; Thompson, B.M.; Cryer, M. Production, drift and mortality of the planktonic larvae of the edible crab (cancer pagurus) off the North-East coast of England. Neth. J. Sea Res. 1982, 16, 173–184. [Google Scholar] [CrossRef]
- Ennis, G. Swimming ability of larval American lobsters, Homarus americanus, in flowing water. Can. J. Fish. Aquat. Sci. 1986, 43, 2177–2183. [Google Scholar] [CrossRef]
- Scarratt, D.J. Abundance and Distribution of Lobster Larvae (Homarus americanus) in Northumberland Strait. J. Fish. Res. Board Can. 1964, 21, 661–680. [Google Scholar] [CrossRef]
- Ward, B.K.; Tan, G.X.; Roberts, D.C.; Della Santina, C.C.; Zee, D.S.; Carey, J.P. Strong static magnetic fields elicit swimming behaviors consistent with direct vestibular stimulation in adult zebrafish. PLoS ONE 2014, 9, e92109. [Google Scholar] [CrossRef]
- Fedele, G.; Green, E.W.; Rosato, E.; Kyriacou, C.P. An electromagnetic field disrupts negative geotaxis in Drosophila via a CRY-dependent pathway. Nat. Commun. 2014, 5, 4391. [Google Scholar] [CrossRef]
- Ugolini, A. Equatorial sandhoppers use body scans to detect the earth’s magnetic field. J. Comp. Physiol. A 2006, 192, 45–49. [Google Scholar] [CrossRef]
- Love, M.S.; Nishimoto, M.M.; Clark, S.; McCrea, M.; Bull, A.S. Assessing potential impacts of energized submarine power cables on crab harvests. Cont. Shelf Res. 2017, 151, 23–29. [Google Scholar] [CrossRef]
- Milton, S.L.; Mary, M.N.; Scott, C.; Ann Scarborough, B. Identical Response of Caged Rock Crabs (Genera Metacarcinus and Cancer) to Energized and Unenergized Undersea Power Cables in Southern California, USA. Bull. South. Calif. Acad. Sci. 2015, 114, 33–41. [Google Scholar] [CrossRef]
- Campbell, A.; Stasko, A.B. Movements of lobsters (Homarus americanus) tagged in the Bay of Fundy, Canada. Mar. Biol. 1986, 92, 393–404. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).