
Dietary Curcumin Supplementation Enhanced Ammonia Nitrogen Stress Tolerance in Greater Amberjack (Seriola dumerili): Growth, Serum Biochemistry and Expression of Stress-Related Genes



Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Fish and Feeding Experiment
2.2. Experimental Diet
2.3. Ammonia Nitrogen Challenge Assay
2.4. Sampling and Analysis
2.5. Gene Expression
2.6. Statistical Analyses
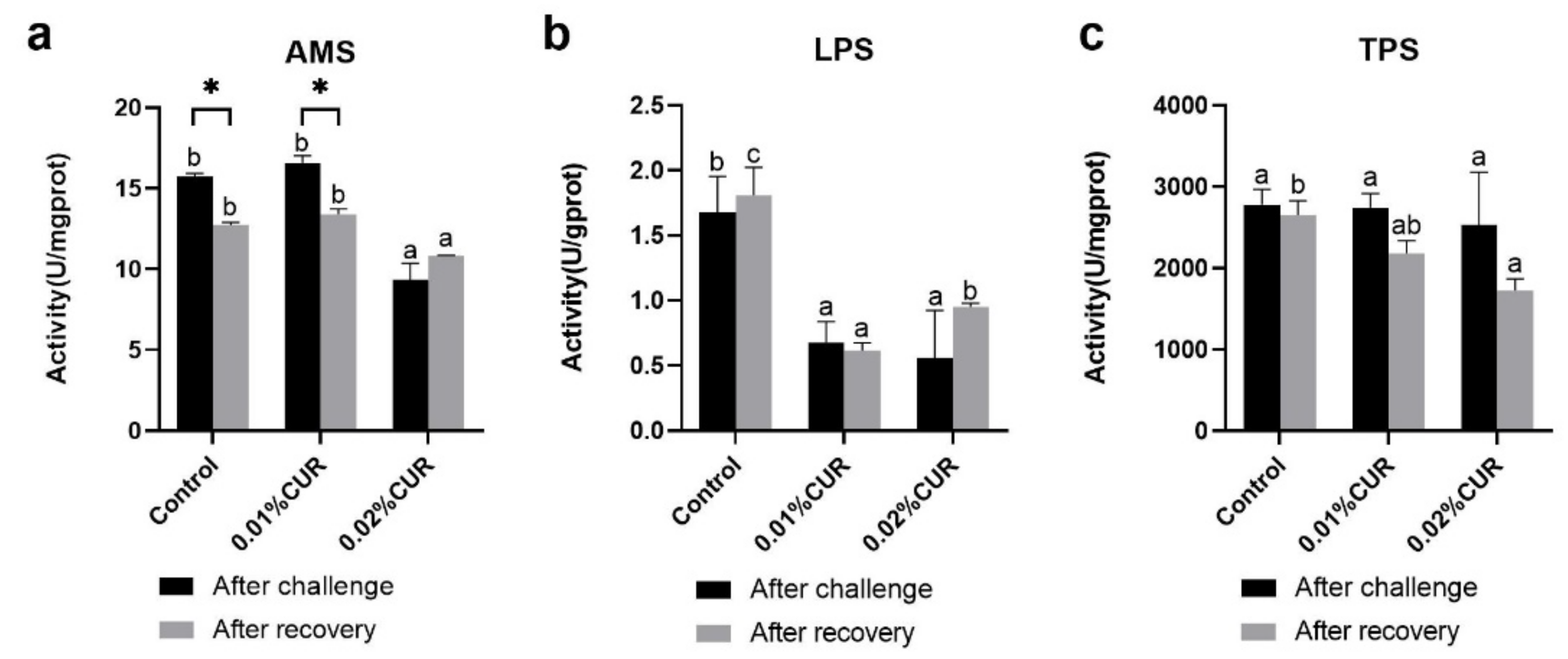
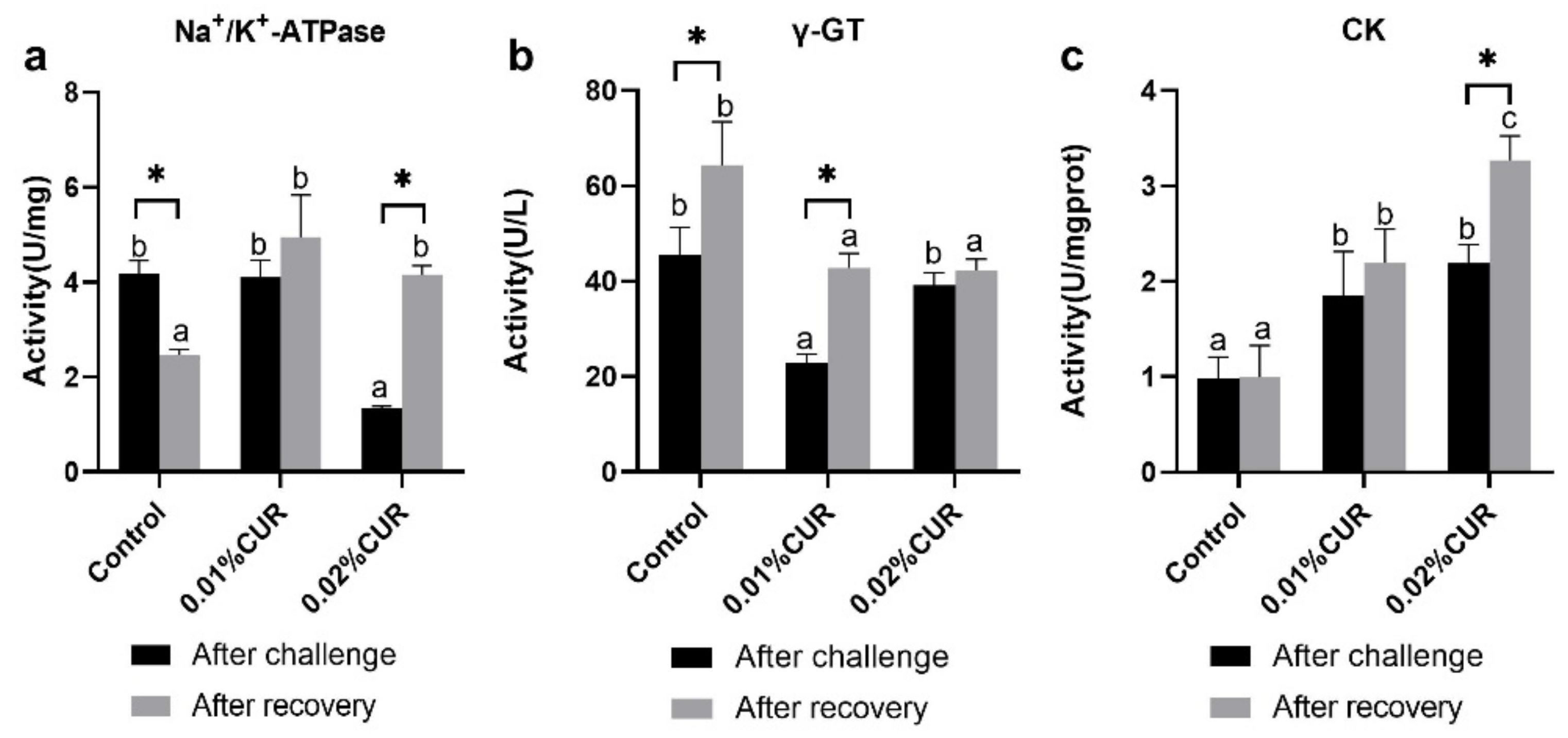
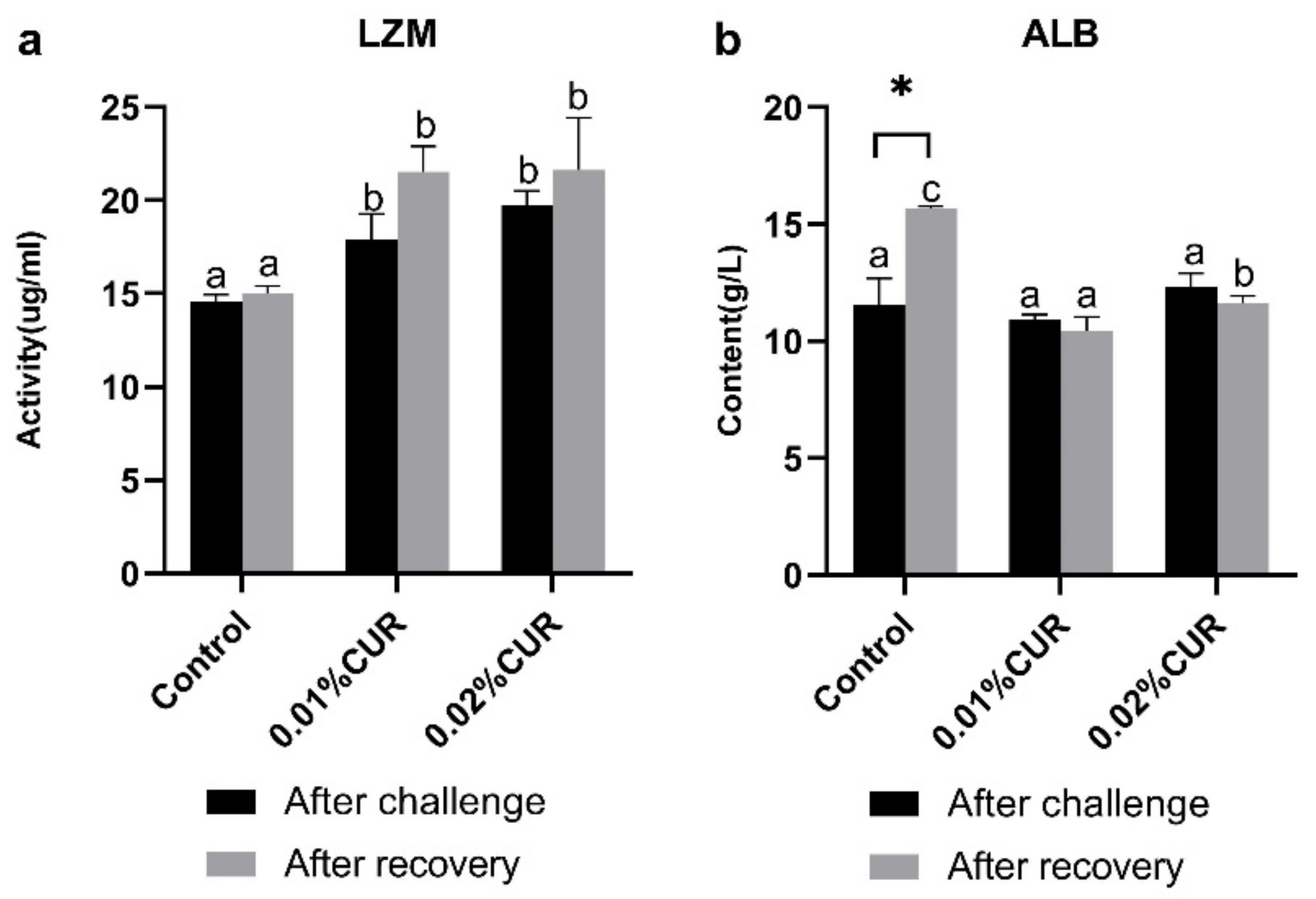
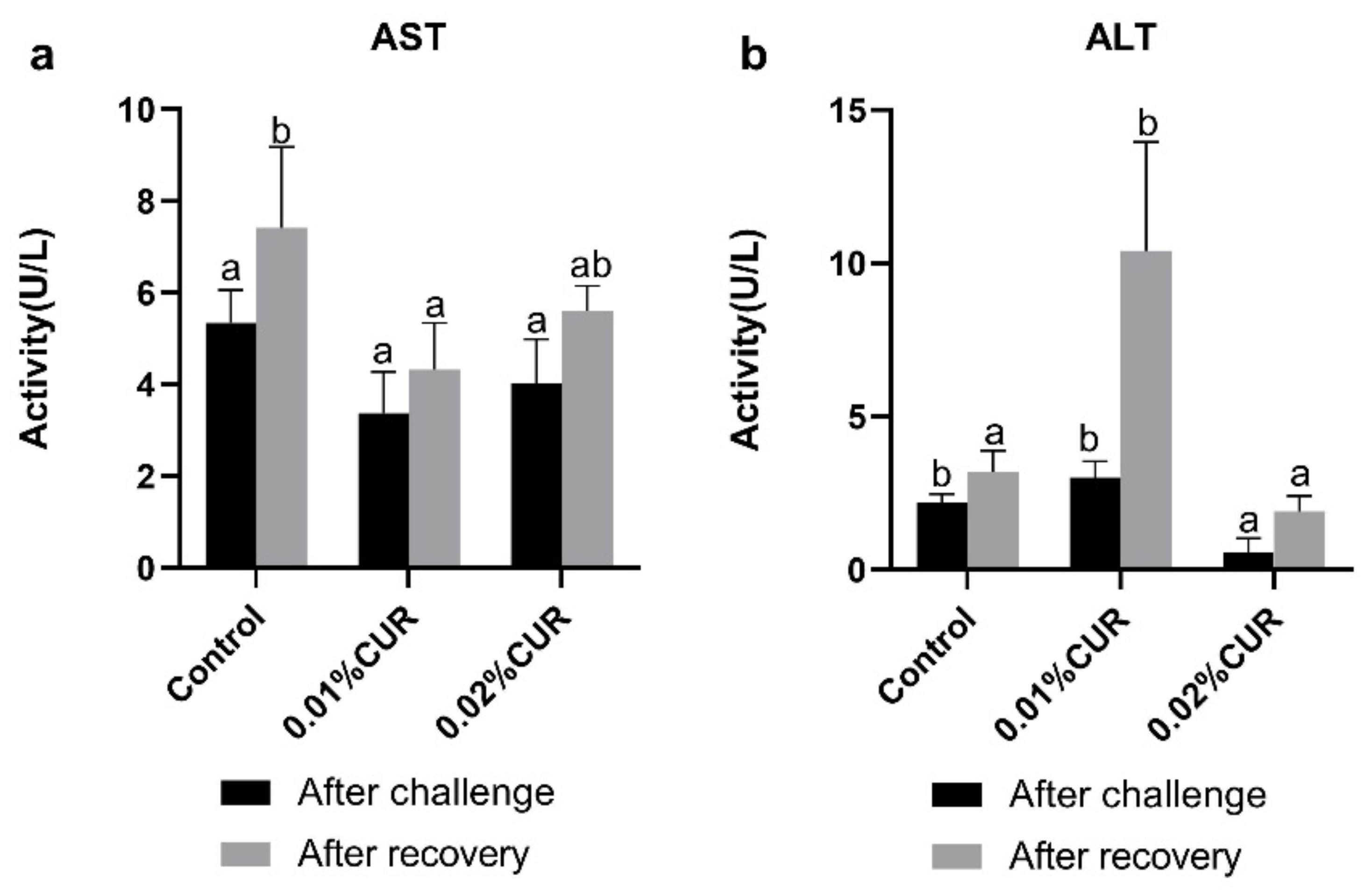
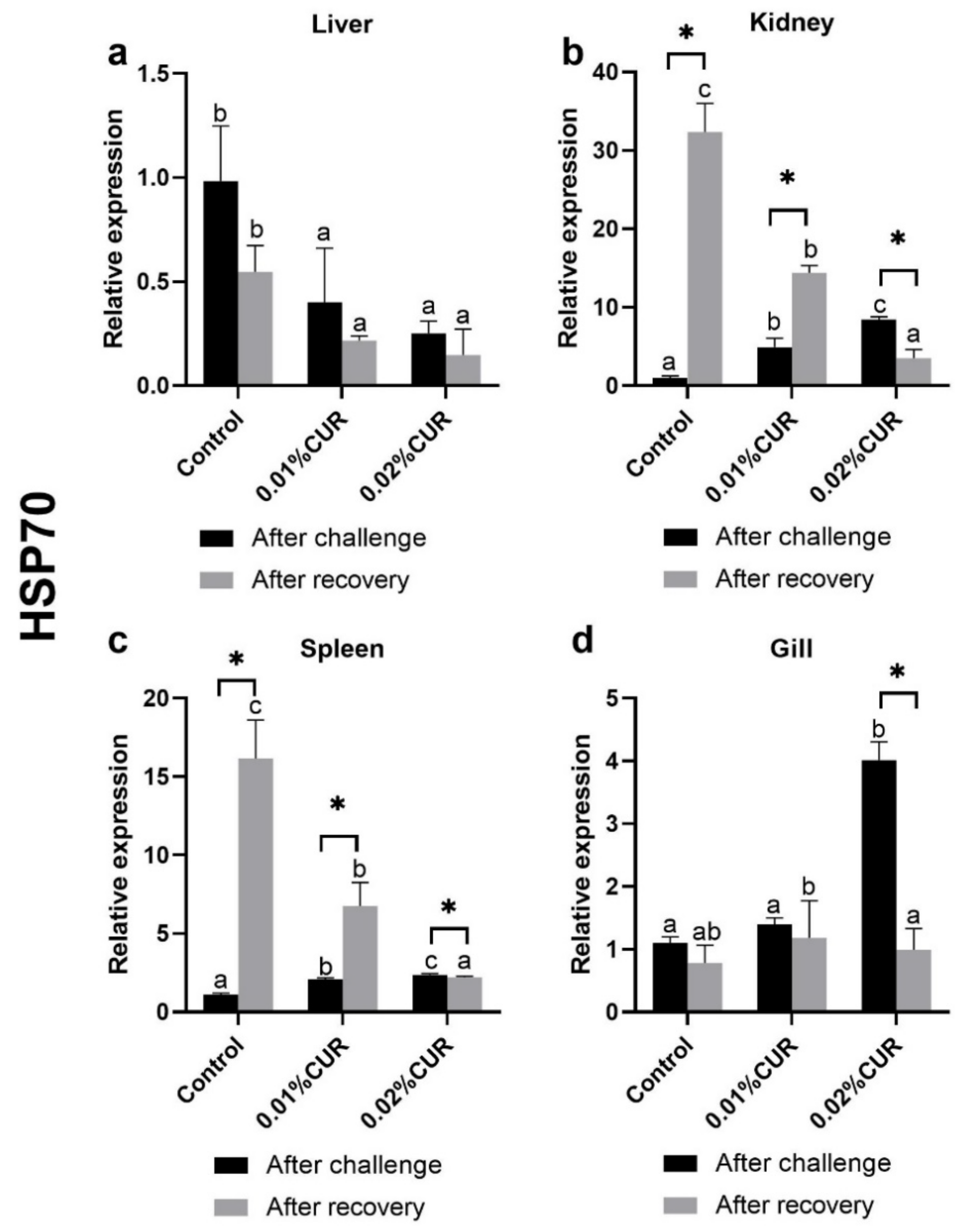
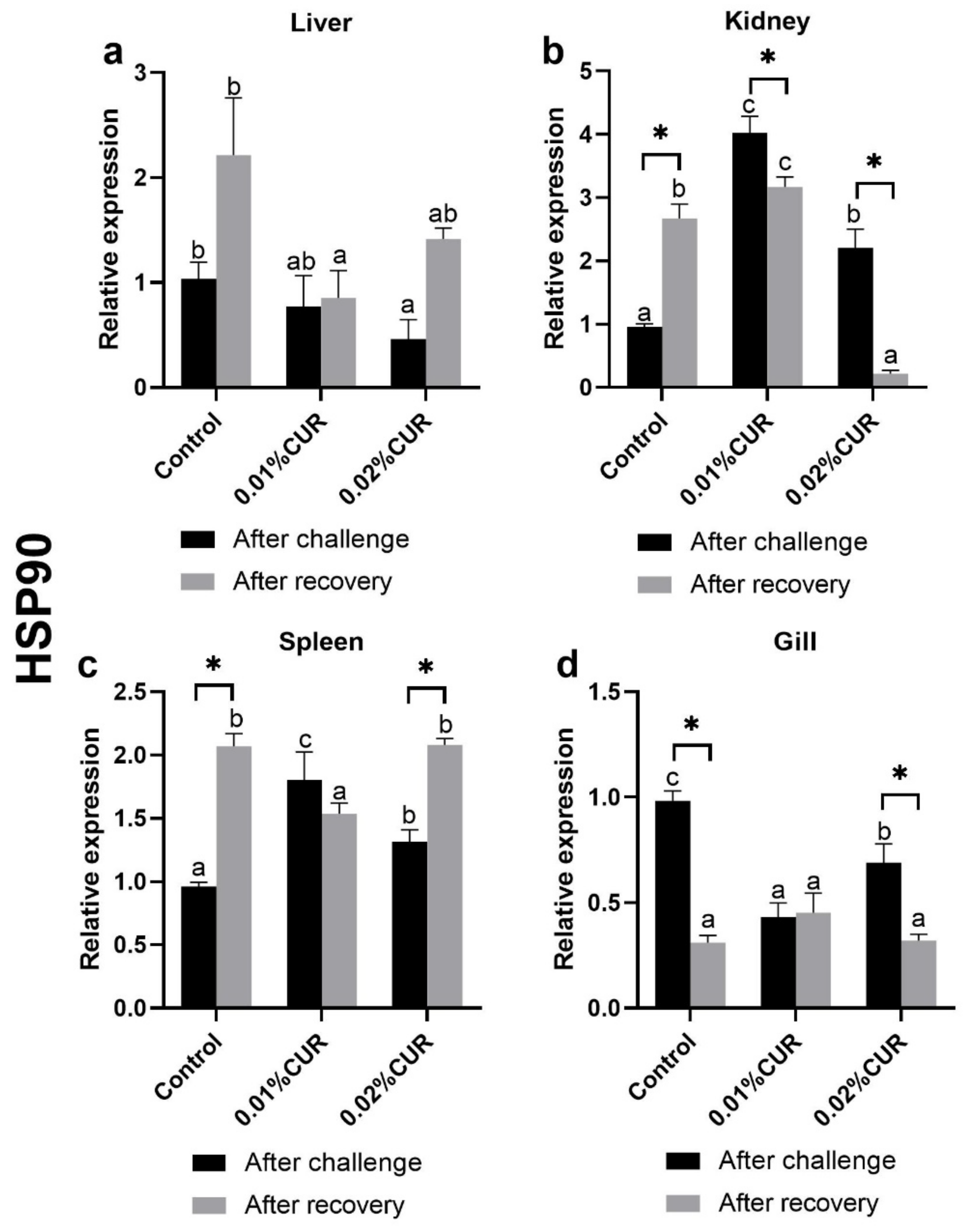
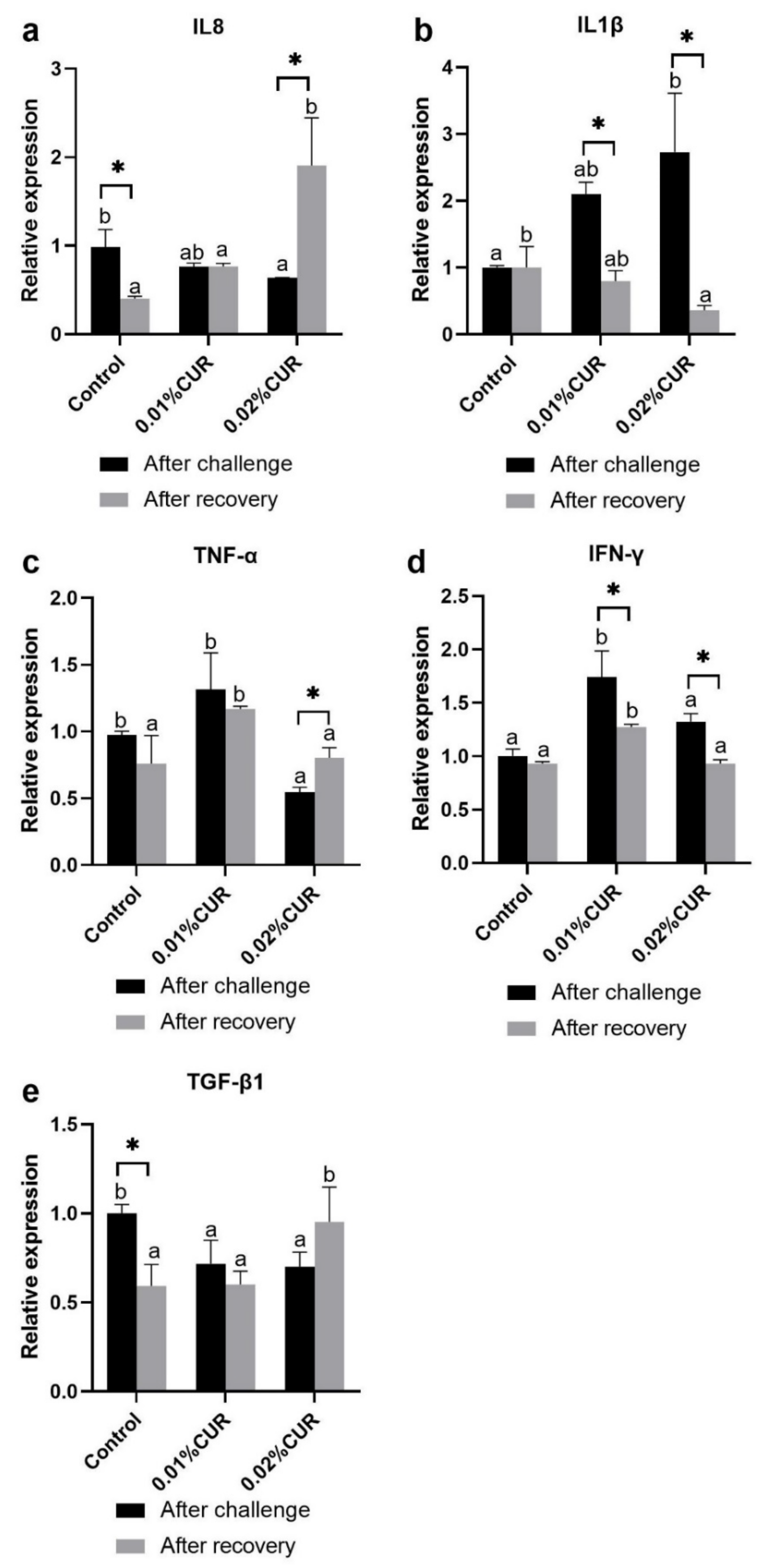
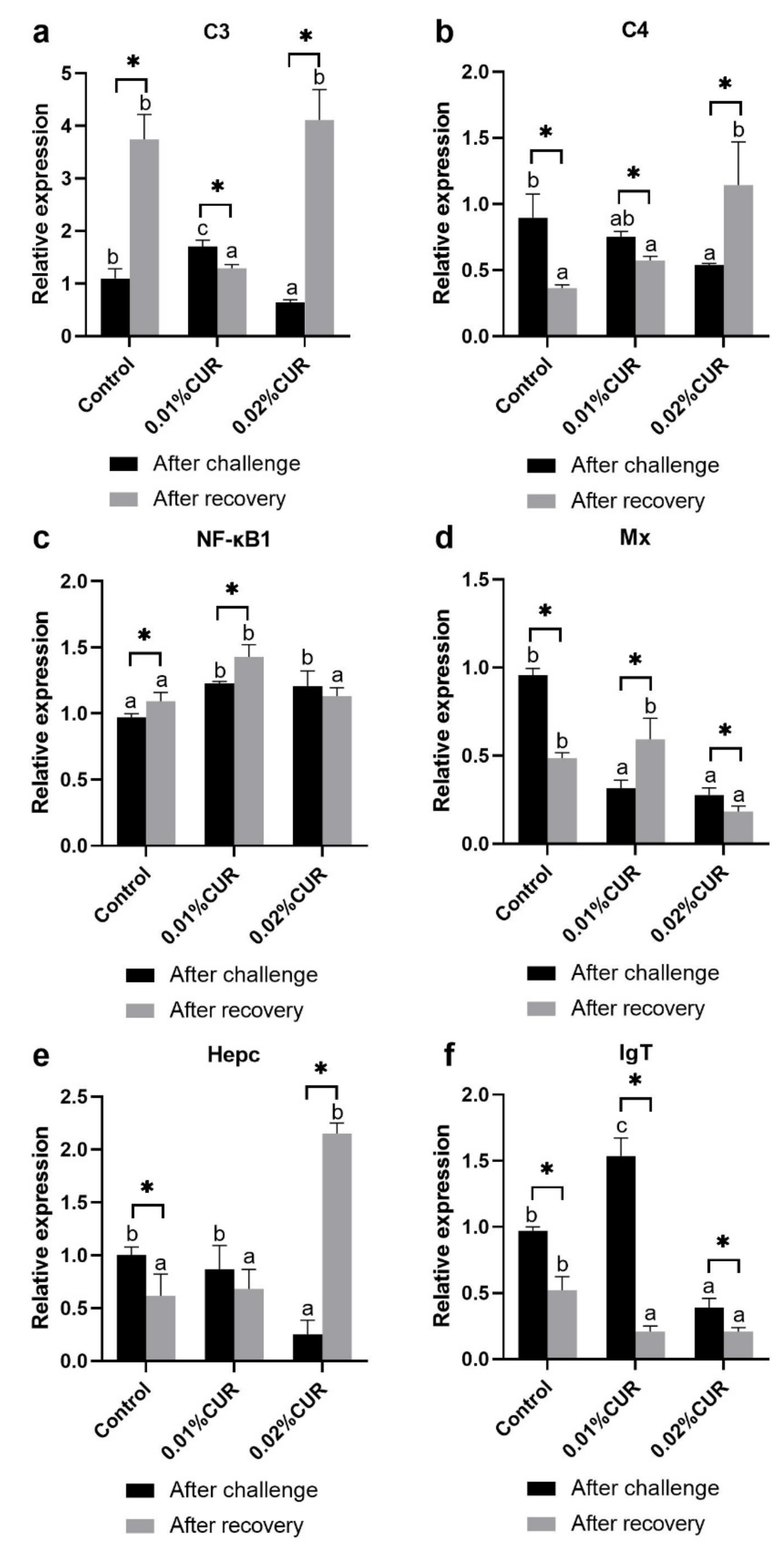
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Angel, D.L.; Eden, N.; Breitstein, S.; Yurman, A.; Katz, T.; Spanier, E. In situ biofiltration: A means to limit the dispersal of effluents from marine finfish cage aquaculture. Hydrobiologia 2002, 469, 1–10. [Google Scholar] [CrossRef]
- Luis, A.I.S.; Campos, E.V.R.; de Oliveira, J.L.; Fraceto, L.F. Trends in aquaculture sciences: From now to use of nanotechnology for disease control. Rev. Aquac. 2019, 11, 119–132. [Google Scholar] [CrossRef]
- Terlizzi, A.; Tedesco, P.; Patarnello, P. Spread of pathogens from marine cage aquaculture-a potential threat for wild fish assemblages under protection regimes. Health Environ. Aquac. 2012, 12, 403–414. [Google Scholar]
- Tovar, A.; Moreno, C.; Mánuel-Vez, M.P.; García-Vargas, M. Environmental impacts of intensive aquaculture in marine waters. Water Res. 2000, 34, 334–342. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, Y.; Zhang, Q.; Liu, P.; Guo, R.; Jin, S.; Liu, J.; Chen, L.; Ma, Z.; Liu, Y. Evaluation and analysis of water quality of marine aquaculture area. Int. J. Environ. Res. Public Health 2020, 17, 1446. [Google Scholar] [CrossRef]
- Wu, J.; Su, Y.; Deng, Y.; Guo, Z.; Cheng, C.; Ma, H.; Liu, G.; Xu, L.; Feng, J. Spatial and temporal variation of antibiotic resistance in marine fish cage-culture area of Guangdong, China. Environ. Pollut. 2019, 246, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, N.; Thompson, S.; Glaser, M. Global aquaculture productivity, environmental sustainability, and climate change adaptability. Environ. Manag. 2019, 63, 159–172. [Google Scholar] [CrossRef] [PubMed]
- Mazzola, A.; Favaloro, E.; Sarà, G. Cultivation of the Mediterranean amberjack, Seriola dumerili (Risso, 1810), in submerged cages in the Western Mediterranean Sea. Aquaculture 2000, 181, 257–268. [Google Scholar] [CrossRef]
- Rigos, G.; Katharios, P.; Kogiannou, D.; Cascarano, C.M. Infectious diseases and treatment solutions of farmed greater amberjack Seriola dumerili with particular emphasis in Mediterranean region. Rev. Aquac. 2021, 13, 301–323. [Google Scholar] [CrossRef]
- Fernández-Montero, A.; Torrecillas, S.; Tort, L.; Ginés, R.; Acosta, F.; Izquierdo, M.; Montero, D. Stress response and skin mucus production of greater amberjack (Seriola dumerili) under different rearing conditions. Aquaculture 2020, 520, 735005. [Google Scholar] [CrossRef]
- Mylonas, C.; Papandroulakis, N.; Corriero, A.; Montero, D.; Hernández-Cruz, C.; Banovic, M.; Tacken, G.; Robles, R. Rearing of greater amberjack (Seriola dumerili). Aquac. Eur. 2019, 44, 18–25. [Google Scholar]
- Yilmaz, E.; Şereflişan, H. Offshore Farming of the Mediterranean Amberjack (Seriola dumerili) in the Northeastern Mediterranean. Isr. J. Aquac. Bamidgeh 2011, 63, 7. [Google Scholar] [CrossRef]
- Zupa, R.; Rodríguez, C.; Mylonas, C.C.; Rosenfeld, H.; Fakriadis, I.; Papadaki, M.; Pérez, J.A.; Pousis, C.; Basilone, G.; Corriero, A. Comparative study of reproductive development in wild and captive-reared greater amberjack Seriola dumerili (Risso, 1810). PLoS ONE 2017, 12, e0169645. [Google Scholar] [CrossRef] [PubMed]
- Mazzora, A. Sistemi di maricoltura open-sea l’allevamento di Seriola dumerili (Pisces, Osteichthyes) nel Golfo di Catellammare (Sicilia Occidentale). Biol. Mar. Med. 1996, 3, 176–185. [Google Scholar]
- Porrello, S.; Andaloro, F.; Vivona, P.; Marino, G. Rearing trial of Seriola dumerili in a floating cage. In Production, Environment and Quality, Proceedings of the International Conference Bordeaux Aquaculture ’92, Bordeaux, France, 25–27 March 1992; EAS Special Publication: Bordeaux, France, 1993; pp. 299–307. [Google Scholar]
- Jover, M.; Garcıa-Gómez, A.; Tomas, A.; de la Gándara, F.; Pérez, L. Growth of Mediterranean yellowtail (Seriola dumerilii) fed extruded diets containing different levels of protein and lipid. Aquaculture 1999, 179, 25–33. [Google Scholar] [CrossRef]
- Wang, W.; Sun, J.; Liu, C.; Xue, Z. Application of immunostimulants in aquaculture: Current knowledge and future perspectives. Aquac. Res. 2017, 48, 1–23. [Google Scholar] [CrossRef]
- Khanna, D.; Sethi, G.; Ahn, K.S.; Pandey, M.K.; Kunnumakkara, A.B.; Sung, B.; Aggarwal, A.; Aggarwal, B.B. Natural products as a gold mine for arthritis treatment. Curr. Opin. Pharmacol. 2007, 7, 344–351. [Google Scholar] [CrossRef]
- Divyagnaneswari, M.; Christybapita, D.; Michael, R.D. Enhancement of nonspecific immunity and disease resistance in Oreochromis mossambicus by Solanum trilobatum leaf fractions. Fish Shellfish Immun. 2007, 23, 249–259. [Google Scholar] [CrossRef]
- Jian, J.; Wu, Z. Influences of traditional Chinese medicine on non-specific immunity of Jian carp (Cyprinus carpio var. Jian). Fish Shellfish Immun. 2004, 16, 185–191. [Google Scholar] [CrossRef]
- Ardó, L.; Yin, G.; Xu, P.; Váradi, L.; Szigeti, G.; Jeney, Z.; Jeney, G. Chinese herbs (Astragalus membranaceus and Lonicera japonica) and boron enhance the non-specific immune response of Nile tilapia (Oreochromis niloticus) and resistance against Aeromonas hydrophila. Aquaculture 2008, 275, 26–33. [Google Scholar] [CrossRef]
- Hwang, J.-H.; Lee, S.-W.; Rha, S.-J.; Yoon, H.-S.; Park, E.-S.; Han, K.-H.; Kim, S.-J. Dietary green tea extract improves growth performance, body composition, and stress recovery in the juvenile black rockfish, Sebastes schlegeli. Aquac. Int. 2013, 21, 525–538. [Google Scholar] [CrossRef]
- Thanigaivel, S.; Vijayakumar, S.; Gopinath, S.; Mukherjee, A.; Chandrasekaran, N.; Thomas, J. In vivo and in vitro antimicrobial activity of Azadirachta indica (Lin) against Citrobacter freundii isolated from naturally infected Tilapia (Oreochromis mossambicus). Aquaculture 2015, 437, 252–255. [Google Scholar] [CrossRef]
- Lestari, M.L.; Indrayanto, G. Curcumin. Profiles Drug Subst. Excip. Relat. Methodol. 2014, 39, 113–204. [Google Scholar] [PubMed]
- Singh, S.; Khar, A. Biological effects of curcumin and its role in cancer chemoprevention and therapy. Anti-Cancer Agents Med. Chem. 2006, 6, 259–270. [Google Scholar] [CrossRef]
- Brouk, B. Plants Consumed by Man; Academic Press: London, UK, 1975. [Google Scholar]
- WHO. Evaluation of Certain Food Additives and Contaminants: Sixty-First Report of the Joint FAO/WHO Expert Committee on Food Additives; World Health Organization: Geneva, Switzerland, 2004. [Google Scholar]
- Kunchandy, E.; Rao, M. Oxygen radical scavenging activity of curcumin. Int. J. Pharm. 1990, 58, 237–240. [Google Scholar] [CrossRef]
- Goel, A.; Boland, C.R.; Chauhan, D.P. Specific inhibition of cyclooxygenase-2 (COX-2) expression by dietary curcumin in HT-29 human colon cancer cells. Cancer Lett. 2001, 172, 111–118. [Google Scholar] [CrossRef]
- Ammon, H.P.; Wahl, M.A. Pharmacology of Curcuma longa. Planta Med. 1991, 57, 1–7. [Google Scholar] [CrossRef]
- Sharma, R.; Gescher, A.; Steward, W. Curcumin: The story so far. Eur. J. Cancer 2005, 41, 1955–1968. [Google Scholar] [CrossRef]
- Li, M.; Zhang, Z.; Hill, D.L.; Wang, H.; Zhang, R. Curcumin, a dietary component, has anticancer, chemosensitization, and radiosensitization effects by down-regulating the MDM2 oncogene through the PI3K/mTOR/ETS2 pathway. Cancer Res. 2007, 67, 1988–1996. [Google Scholar] [CrossRef]
- Abdel-Tawwab, M.; Abbass, F.E. Turmeric powder, Curcuma longa L., in common carp, Cyprinus carpio L., diets: Growth performance, innate immunity, and challenge against pathogenic Aeromonas hydrophila infection. J. World Aquac. Soc. 2017, 48, 303–312. [Google Scholar] [CrossRef]
- Adeshina, I.; Adewale, Y.; Tiamiyu, L. Growth performance and innate immune response of Clarias gariepinus infected with Aeromonas hydrophila fed diets fortified with Curcuma longa leaf. West Afr. J. Appl. Ecol. 2017, 25, 87–99. [Google Scholar]
- Jiang, J.; Wu, X.-Y.; Zhou, X.-Q.; Feng, L.; Liu, Y.; Jiang, W.-D.; Wu, P.; Zhao, Y. Effects of dietary curcumin supplementation on growth performance, intestinal digestive enzyme activities and antioxidant capacity of crucian carp Carassius auratus. Aquaculture 2016, 463, 174–180. [Google Scholar] [CrossRef]
- Ji, R.; Xiang, X.; Li, X.; Mai, K.; Ai, Q. Effects of dietary curcumin on growth, antioxidant capacity, fatty acid composition and expression of lipid metabolism-related genes of large yellow croaker fed a high-fat diet. Br. J. Nutr. 2021, 126, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, F.; Yao, F.; Ma, R.; Zhang, Y.; Mao, S.; Hu, B.; Ma, G.; Zhu, Y. Study of dietary curcumin on the restorative effect of liver injury induced by carbon tetrachloride in common carp Cyprinus carpio. Aquac. Rep. 2021, 21, 100825. [Google Scholar] [CrossRef]
- Mahmoud, H.K.; Al-Sagheer, A.A.; Reda, F.M.; Mahgoub, S.A.; Ayyat, M.S. Dietary curcumin supplement influence on growth, immunity, antioxidant status, and resistance to Aeromonas hydrophila in Oreochromis niloticus. Aquaculture 2017, 475, 16–23. [Google Scholar] [CrossRef]
- Garcia-Garrido, L.; Muñoz-Chapuli, R.; de Andres, A. Serum cholesterol and triglyceride levels in Scyliorhinus canicula (L.) during sexual maturation. J. Fish Biol. 1990, 36, 499–509. [Google Scholar] [CrossRef]
- Doumas, B.T.; Watson, W.A.; Biggs, H.G. Albumin standards and the measurement of serum albumin with bromcresol green. Clin. Chim. Acta 1971, 31, 87–96. [Google Scholar] [CrossRef]
- Reitman, S.; Frankel, S. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol. 1957, 28, 56–63. [Google Scholar] [CrossRef]
- Obach, A.; Quentel, C.; Laurencin, F.B. Dicen trarch us la brax. Dis. Aquat. Org. 1993, 15, 175–185. [Google Scholar] [CrossRef]
- Reimer, G. The influence of diet on the digestive enzymes of the Amazon fish Matrincha, Brycon cf. melanopterus. J. Fish Biol. 1982, 21, 637–642. [Google Scholar] [CrossRef]
- Ay, Ö.; Kalay, M.; Tamer, L.; Canli, M. Copper and lead accumulation in tissues of a freshwater fish Tilapia zillii and its effects on the branchial Na, K-ATPase activity. Bull. Environ. Contam. Toxicol. 1999, 62, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Weng, C.F.; Chiang, C.C.; Gong, H.Y.; Chen, M.H.; Lin, C.J.; Huang, W.T.; Cheng, C.Y.; Hwang, P.P.; Wu, J.L. Acute changes in gill Na+-K+-ATPase and creatine kinase in response to salinity changes in the euryhaline teleost, tilapia (Oreochromis mossambicus). Physiol Biochem. Zool 2002, 75, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Rosalki, S.; Rau, D.; Lehmann, D.; Prentice, M. Gamma-glutamyl transpeptidase in chronic alcoholism. Lancet 1970, 296, 1139. [Google Scholar] [CrossRef]
- Fernández-Montero, Á.; Torrecillas, S.; Izquierdo, M.; Caballero, M.J.; Milne, D.J.; Secombes, C.J.; Sweetman, J.; da Silva, P.; Acosta, F.; Montero, D. Increased parasite resistance of greater amberjack (Seriola dumerili Risso 1810) juveniles fed a cMOS supplemented diet is associated with upregulation of a discrete set of immune genes in mucosal tissues. Fish Shellfish Immun. 2019, 86, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Montero, Á.; Torrecillas, S.; Acosta, F.; Kalinowski, T.; Bravo, J.; Sweetman, J.; Roo, J.; Makol, A.; Docando, J.; Carvalho, M. Improving greater amberjack (Seriola dumerili) defenses against monogenean parasite Neobenedenia girellae infection through functional dietary additives. Aquaculture 2021, 534, 736317. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.P.; Zhou, X.Q.; Feng, L.; Jiang, J.; Liu, Y. Effect of dietary pantothenic acid supplement on growth, body composition and intestinal enzyme activities of juvenile Jian carp (Cyprinus carpio var. Jian). Aquac. Nutr. 2009, 15, 470–476. [Google Scholar] [CrossRef]
- Blier, P.; Pelletier, D.; Dutil, J.D. Does aerobic capacity set a limit on fish growth rate? Rev. Fish Sci. 1997, 5, 323–340. [Google Scholar] [CrossRef]
- Ling, J.; Feng, L.; Liu, Y.; Jiang, J.; Jiang, W.D.; Hu, K.; Li, S.H.; Zhou, X.Q. Effect of dietary iron levels on growth, body composition and intestinal enzyme activities of juvenile Jian carp (Cyprinus carpio var. Jian). Aquac. Nutr. 2010, 16, 616–624. [Google Scholar] [CrossRef]
- Li, G.; Zhou, X.; Jiang, W.; Wu, P.; Liu, Y.; Jiang, J.; Kuang, S.; Tang, L.; Shi, H.; Feng, L. Dietary curcumin supplementation enhanced growth performance, intestinal digestion, and absorption and amino acid transportation abilities in on-growing grass carp (Ctenopharyngodon idella). Aquac. Res. 2020, 51, 4863–4873. [Google Scholar] [CrossRef]
- Sruthi, M.; Nair, A.B.; Arun, D.; Thushara, V.; Sheeja, C.; Vijayasree, A.S.; Oommen, O.V.; Divya, L. Dietary curcumin influences leptin, growth hormone and hepatic growth factors in Tilapia (Oreochromis mossambicus). Aquaculture 2018, 496, 105–111. [Google Scholar] [CrossRef]
- Yonar, M.E.; Yonar, S.M.; İspir, Ü.; Ural, M.Ş. Effects of curcumin on haematological values, immunity, antioxidant status and resistance of rainbow trout (Oncorhynchus mykiss) against Aeromonas salmonicida subsp. achromogenes. Fish Shellfish Immun. 2019, 89, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Liu, B.; Ge, X.; XiE, J.; Xu, P.; Miao, L.; Sun, S.; Liao, Y.; Chen, R.; Ren, M. Effects of dietary curcumin on growth performance, biochemical parameters, HSP70 gene expression and resistance to Streptococcus iniae of juvenile Gift Tilapia, Oreochromis niloticus. Isr. J. Aquac. 2013, 66, 986–996. [Google Scholar]
- Manal, I. Detoxification and antioxidant effects of garlic and curcumin in Oreochromis niloticus injected with aflatoxin B 1 with reference to gene expression of glutathione peroxidase (GPx) by RT-PCR. Fish Physiol. Biochem. 2016, 42, 617–629. [Google Scholar]
- Manal, I. Impact of garlic and curcumin on the hepatic histology and cytochrome P450 gene expression of aflatoxicosis Oreochromis niloticus using RT-PCR. Turk. J. Fish. Aquat. Sci. 2018, 18, 405–415. [Google Scholar]
- Tort, L.; Balasch, J.; Mackenzie, S. Fish immune system. A crossroads between innate and adaptive responses. Inmunología 2003, 22, 277–286. [Google Scholar]
- Saurabh, S.; Sahoo, P. Lysozyme: An important defence molecule of fish innate immune system. Aquac. Res. 2008, 39, 223–239. [Google Scholar] [CrossRef]
- Leya, T.; Dar, S.A.; Kumar, G.; Ahmad, I. Curcumin supplement diet: Enhanced growth and down-regulated expression of pro-inflammatory cytokines in Labeo rohita fingerlings. Aquac. Res. 2020, 51, 4785–4792. [Google Scholar] [CrossRef]
- Misra, C.; Das, B.; Mukherjee, S.; Meher, P. The immunomodulatory effects of tuftsin on the non-specific immune system of Indian Major carp, Labeo rohita. Fish Shellfish Immun. 2006, 20, 728–738. [Google Scholar] [CrossRef]
- Wiegertjes, G.F. Immunogenetics of Disease Resistance in Fish. Doctoral Thesis, Wageningen Agricultural University, Wageningen, The Netherlands, 1995. [Google Scholar]
- Leya, T.; Raman, R.P.; Srivastava, P.P.; Kumar, K.; Ahmad, I.; Gora, A.H.; Poojary, N.; Kumar, S.; Dar, S.A. Effects of curcumin supplemented diet on growth and non-specific immune parameters of Cirrhinus mrigala against Edwardsiella tarda infection. Int J. Curr. Microbiol. Appl. Sci. 2017, 6, 1230–1243. [Google Scholar]
- Diab, A.M.; Saker, O.; Eldakroury, M.; Elseify, M. Effects of garlic (Allium sativum) and curcumin (Turmeric, Curcuma longa Linn) on Nile tilapia immunity. Vet. Med. J. 2014, 60, C1–C19. [Google Scholar]
- Kwo, P.Y.; Cohen, S.M.; Lim, J.K. ACG clinical guideline: Evaluation of abnormal liver chemistries. Off. J. Am. Coll. Gastroenterol. ACG 2017, 112, 18–35. [Google Scholar] [CrossRef] [PubMed]
- Bao, J.-W.; Qiang, J.; Tao, Y.-F.; Li, H.-X.; He, J.; Xu, P.; Chen, D.-J. Responses of blood biochemistry, fatty acid composition and expression of microRNAs to heat stress in genetically improved farmed tilapia (Oreochromis niloticus). J. Therm. Biol. 2018, 73, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Pakhira, C.; Nagesh, T.; Abraham, T.; Dash, G.; Behera, S. Stress responses in rohu, Labeo rohita transported at different densities. Aquac. Rep. 2015, 2, 39–45. [Google Scholar] [CrossRef]
- Hong, J.; Zhou, S.; Yu, G.; Qin, C.; Zuo, T.; Ma, Z. Effects of transporting stress on the immune responses of Asian seabass Lates calcarifer fry. Aquac. Res. 2021, 52, 2182–2193. [Google Scholar] [CrossRef]
- Xia, S.-L.; Ge, X.-P.; Liu, B.; Xie, J.; Miao, L.-H.; Ren, M.-C.; Zhou, Q.-L.; Zhang, W.-X.; Jiang, X.-J.; Chen, R.-L. Effects of supplemented dietary curcumin on growth and non-specific immune responses in juvenile wuchang bream (Megalobrama amblycephala). Isr. J. Aquac. Bamid. 2015, 67. [Google Scholar] [CrossRef]
- Yu, S.; Yu, Y.; Ai, T.; Li, Q. Study of the recovery effect of curcumin on liver injury in grass carp. Chin. J. Vet. Drug 2013, 47, 29–31. [Google Scholar]
- Shao, W.; Yu, Z.; Chiang, Y.; Yang, Y.; Chai, T.; Foltz, W.; Lu, H.; Fantus, I.G.; Jin, T. Curcumin prevents high fat diet induced insulin resistance and obesity via attenuating lipogenesis in liver and inflammatory pathway in adipocytes. PLoS ONE 2012, 7, e28784. [Google Scholar] [CrossRef]
- Yang, Y.S.; Su, Y.F.; Yang, H.W.; Lee, Y.H.; Chou, J.I.; Ueng, K.C. Lipid-lowering effects of curcumin in patients with metabolic syndrome: A randomized, double-blind, placebo-controlled trial. Phytother. Res. 2014, 28, 1770–1777. [Google Scholar] [CrossRef]
- Arafa, H.M. Curcumin attenuates diet-induced hypercholesterolemia in rats. Med. Sci. Monit. 2005, 11, BR228–BR234. [Google Scholar]
- Jang, E.-M.; Choi, M.-S.; Jung, U.J.; Kim, M.-J.; Kim, H.-J.; Jeon, S.-M.; Shin, S.-K.; Seong, C.-N.; Lee, M.-K. Beneficial effects of curcumin on hyperlipidemia and insulin resistance in high-fat–fed hamsters. Metabolism 2008, 57, 1576–1583. [Google Scholar] [CrossRef] [PubMed]
- Panahi, Y.; Kianpour, P.; Mohtashami, R.; Jafari, R.; Simental-Mendía, L.E.; Sahebkar, A. Curcumin lowers serum lipids and uric acid in subjects with nonalcoholic fatty liver disease: A randomized controlled trial. J. Cardiovasc. Pharm. 2016, 68, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Roberts, R.; Agius, C.; Saliba, C.; Bossier, P.; Sung, Y. Heat shock proteins (chaperones) in fish and shellfish and their potential role in relation to fish health: A review. J. Fish Dis. 2010, 33, 789–801. [Google Scholar] [CrossRef] [PubMed]
- Newton, J.; de Santis, C.; Jerry, D. The gene expression response of the catadromous perciform barramundi Lates calcarifer to an acute heat stress. J. Fish Biol. 2012, 81, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Stitt, B.C.; Burness, G.; Burgomaster, K.A.; Currie, S.; McDermid, J.L.; Wilson, C.C. Intraspecific variation in thermal tolerance and acclimation capacity in brook trout (Salvelinus fontinalis): Physiological implications for climate change. Physiol. Biochem. Zool 2014, 87, 15–29. [Google Scholar] [CrossRef] [PubMed]
- Templeman, N.M.; LeBlanc, S.; Perry, S.F.; Currie, S. Linking physiological and cellular responses to thermal stress: β-adrenergic blockade reduces the heat shock response in fish. J. Comp. Physiol. B 2014, 184, 719–728. [Google Scholar] [CrossRef]
- Currie, S. Temperature| Heat Shock Proteins and Temperature. Encycl. Fish Physiol. 2011, 3, 1732–1737. [Google Scholar]
- Savan, R.; Sakai, M. Genomics of fish cytokines. Comp. Biochem. Phys. D 2006, 1, 89–101. [Google Scholar] [CrossRef]
- Kohli, K.; Ali, J.; Ansari, M.; Raheman, Z. Curcumin: A natural antiinflammatory agent. Indian J. Pharm. 2005, 37, 141. [Google Scholar] [CrossRef]
- Zhou, H.; Beevers, C.S.; Huang, S. The targets of curcumin. Curr. Drug Targets 2011, 12, 332–347. [Google Scholar] [CrossRef]
- Cao, L.; Ding, W.; Du, J.; Jia, R.; Liu, Y.; Zhao, C.; Shen, Y.; Yin, G. Effects of curcumin on antioxidative activities and cytokine production in Jian carp (Cyprinus carpio var. Jian) with CCl4-induced liver damage. Fish Shellfish Immun. 2015, 43, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Alagawany, M.; Farag, M.R.; Abdelnour, S.A.; Dawood, M.A.; Elnesr, S.S.; Dhama, K. Curcumin and its different forms: A review on fish nutrition. Aquaculture 2021, 532, 736030. [Google Scholar] [CrossRef]
- Anthwal, A.; Thakur, B.K.; Rawat, M.; Rawat, D.; Tyagi, A.K.; Aggarwal, B.B. Synthesis, characterization and in vitro anticancer activity of C-5 curcumin analogues with potential to inhibit TNF-α-induced NF-κB activation. BioMed. Res. Int. 2014, 2014, 524161. [Google Scholar] [CrossRef] [PubMed]
- Akdemir, F.; Orhan, C.; Tuzcu, M.; Sahin, N.; Juturu, V.; Sahin, K. The efficacy of dietary curcumin on growth performance, lipid peroxidation and hepatic transcription factors in rainbow trout Oncorhynchus mykiss (Walbaum) reared under different stocking densities. Aquac. Res. 2017, 48, 4012–4021. [Google Scholar] [CrossRef]
- Giri, S.S.; Kim, M.J.; Kim, S.G.; Kim, S.W.; Kang, J.W.; Kwon, J.; Lee, S.B.; Jung, W.J.; Sukumaran, V.; Park, S.C. Role of dietary curcumin against waterborne lead toxicity in common carp Cyprinus carpio. Ecotoxicol. Environ. Saf. 2021, 219, 112318. [Google Scholar] [CrossRef] [PubMed]
- Holland, M.C.H.; Lambris, J.D. The complement system in teleosts. Fish Shellfish Immun. 2002, 12, 399–420. [Google Scholar] [CrossRef] [PubMed]
- Boshra, H.; Li, J.; Sunyer, J. Recent advances on the complement system of teleost fish. Fish Shellfish Immun. 2006, 20, 239–262. [Google Scholar] [CrossRef]
- Carroll, M.C. The role of complement and complement receptors in induction and regulation of immunity. Annu. Rev. Immunol. 1998, 16, 545–568. [Google Scholar] [CrossRef]
- Gasque, P. Complement: A unique innate immune sensor for danger signals. Mol. Immunol. 2004, 41, 1089–1098. [Google Scholar] [CrossRef]
- Chen, S.-L.; Li, W.; Meng, L.; Sha, Z.-X.; Wang, Z.-J.; Ren, G.-C. Molecular cloning and expression analysis of a hepcidin antimicrobial peptide gene from turbot (Scophthalmus maximus). Fish Shellfish Immun. 2007, 22, 172–181. [Google Scholar] [CrossRef]
- Cuesta, A.; Meseguer, J.; Esteban, M.A. The antimicrobial peptide hepcidin exerts an important role in the innate immunity against bacteria in the bony fish gilthead seabream. Mol. Immunol. 2008, 45, 2333–2342. [Google Scholar] [CrossRef] [PubMed]
- Lehrer, R.I.; Ganz, T. Antimicrobial peptides in mammalian and insect host defence. Currt. Opin. Immunol. 1999, 11, 23–27. [Google Scholar] [CrossRef]
- Haller, O.; Kochs, G. Interferon-induced mx proteins: Dynamin-like GTPases with antiviral activity. Traffic 2002, 3, 710–717. [Google Scholar] [CrossRef] [PubMed]
- Haller, O.; Kochs, G. Human MxA protein: An interferon-induced dynamin-like GTPase with broad antiviral activity. J. Interf. Cytok Res. 2011, 31, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-A.; Salinas, I.; Sunyer, J.O. Recent findings on the structure and function of teleost IgT. Fish Shellfish Immun. 2011, 31, 627–634. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-A.; Salinas, I.; Li, J.; Parra, D.; Bjork, S.; Xu, Z.; LaPatra, S.E.; Bartholomew, J.; Sunyer, J.O. IgT, a primitive immunoglobulin class specialized in mucosal immunity. Nat. Immunol. 2010, 11, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Giacomelli, S.; Buonocore, F.; Albanese, F.; Scapigliati, G.; Gerdol, M.; Oreste, U.; Coscia, M.R. New insights into evolution of IgT genes coming from Antarctic teleosts. Mar. Genom. 2015, 24, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Xavier, M.J.; Dardengo, G.M.; Navarro-Guillén, C.; Lopes, A.; Colen, R.; Valente, L.M.; Conceição, L.E.; Engrola, S. Dietary curcumin promotes gilthead seabream larvae digestive capacity and modulates oxidative status. Animals 2021, 11, 1667. [Google Scholar] [CrossRef]
- Xavier, M.J.; Navarro-Guillén, C.; Lopes, A.; Colen, R.; Teodosio, R.; Mendes, R.; Oliveira, B.; Valente, L.M.; Conceição, L.E.; Engrola, S. Effects of dietary curcumin in growth performance, oxidative status and gut morphometry and function of gilthead seabream postlarvae. Aquac. Rep. 2022, 24, 101128. [Google Scholar] [CrossRef]
- Kong, Y.; Li, M.; Guo, G.; Yu, L.; Sun, L.; Yin, Z.; Li, R.; Chen, X.; Wang, G. Effects of dietary curcumin inhibit deltamethrin-induced oxidative stress, inflammation and cell apoptosis in Channa argus via Nrf2 and NF-κB signaling pathways. Aquaculture 2021, 540, 736744. [Google Scholar] [CrossRef]
- Li, M.; Kong, Y.; Wu, X.; Guo, G.; Sun, L.; Lai, Y.; Zhang, J.; Niu, X.; Wang, G. Effects of dietary curcumin on growth performance, lipopolysaccharide-induced immune responses, oxidative stress and cell apoptosis in snakehead fish (Channa argus). Aquac. Rep. 2022, 22, 100981. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Proportion (g/kg) | ||
|---|---|---|---|
| Control | 0.01% CUR 1 | 0.02% CUR 1 | |
| Fish meal | 590 | 590 | 590 |
| Corn gluten meal | 70 | 70 | 70 |
| Soybean meal | 80 | 80 | 80 |
| Corn starch | 80 | 80 | 80 |
| Microcrystalline Cellulose | 50 | 50 | 50 |
| Fish oil | 70 | 70 | 70 |
| Phospholipids | 10 | 10 | 10 |
| 2 Vitamin premix | 5 | 5 | 5 |
| 3 Mineral premix | 5 | 5 | 5 |
| Choline chloride | 5 | 5 | 5 |
| Betaine | 5 | 5 | 5 |
| Carboxymethyl cellulose | 30 | 30 | 30 |
| Curcumin | 0 | 0.1 | 0.2 |
| Analyzed nutrients | |||
| Dry Matter | 878.98 | 878.98 | 878.98 |
| Crude protein | 497 | 497 | 497 |
| Crude lipids | 126.98 | 126.98 | 126.98 |
| Crude ash | 106.85 | 106.85 | 106.85 |
| Gene Abbreviation | Primer Sequence (5′-3′) | Amplicon Size (bp) | Accession Number |
|---|---|---|---|
| NF-κB1 | F:CACAGACAGTTCGCCATCG | 185 | XM_022761336.1 |
| R:AGCGTCTTCTGCCTCTTCC | |||
| 1TNF-α | F:GAAAACGCTTCATGCCTCTC | 212 | XM_022746377.1 |
| R:GTTGGTTTCCGTCCACAGTT | |||
| 1IL-1β | F:TGATGGAGAACATGGTGGAA | 205 | XM_022753745.1 |
| R:GTCGACATGGTCAGATGCAC | |||
| 1IL-8 | F:GAAGCCTGGGAGTAGAGCTG | 164 | XM_022758559.1 |
| R:GGGGTCTAGGCAGACCTCTT | |||
| TGF-β1 | F:CGGAGCTGCGGATGTTAA | 111 | XM_022738547.1 |
| R:TGGTGATGAAGCGGGAAG | |||
| C3 | F:CATCGTTCCGCATCATAGC | 81 | XM_022755728 |
| R:AGTCCTTGACATCCACCCA | XM_022755434 | ||
| C4 | F:ACATCGCAATGGAGGAGAAC | 170 | XM_022768450.01 |
| R:CAGTCCCGTGATAGGCTTTA | |||
| 2IgT | F:TGGACCAGTCGCCATCTGAG | 196 | XM_022756471.1 |
| R:GGGAAACGGCTTTGAAAGGA | |||
| 2Hepc | F:GATGATGCCGAATCCCGTCAGG | 99 | XM_022764299.1 |
| R:CAGAAACCGCAGCCCTTGTTGGC | |||
| IFN-γ | F:TCTGTCTGACCCTCTGGTTTTC | 136 | LC146385.1 |
| R:AAGATGGGCTTCCCGCTA | |||
| Mx | F:GACTTGGCTCTACCTGCTATCG | 177 | XM_022744797.1 |
| R:GCTTATCTTTCCGTACCACTCC | |||
| HSP70 | F:CACGTATTCTTGCGTTGGG | 146 | XM_022741879.1 |
| R:TCATGGCGACCTGGTTCT | |||
| HSP90 | F:GGCTACATGGCCGCTAAA | 187 | XM_022764360.01 |
| R:TGCGATTGGAGTGGGTCTG | |||
| 3EF1 α | F:ATCGTTGCCGCTGGTGTT | 134 | XM_022744048.1 |
| R:TCGGTGGAGTCCATCTTGTT | |||
| 1,3β-actin | F:TCTGGTGGGGCAATGATCTTGATCTT | 212 | XM_022757055.1 |
| R:CCTTCCTTCCTCGGTATGGAGTCC |
| Growth Performance | Group | ||
|---|---|---|---|
| Control Group | 0.01% Treatment Group | 0.02% Treatment Group | |
| Initial weight (g) | 149.02 ± 12.59 | 153.51 ± 4.09 | 151.78 ± 3.85 |
| Final weight (g) | 263.59 ± 83.29 | 327.38 ± 48.38 | 306.55 ± 23.95 |
| Weight gain percentage (%) | 73.89 ± 53.73 | 116.44 ± 34.07 | 102.30 ± 12.83 |
| Specific growth rate (%/d) | 1.26 ± 0.09 | 1.34 ± 0.25 | 1.25 ± 0.12 |
| Feed coefficient | 3.45 ± 0.33 a | 2.21 ± 0.21 b | 2.30 ± 0.31 b |
| Condition factor | 2.36 ± 0.05 b | 2.48 ± 0.06 a | 2.27 ± 0.05 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, J.; Fu, Z.; Hu, J.; Zhou, S.; Yu, G.; Ma, Z. Dietary Curcumin Supplementation Enhanced Ammonia Nitrogen Stress Tolerance in Greater Amberjack (Seriola dumerili): Growth, Serum Biochemistry and Expression of Stress-Related Genes. J. Mar. Sci. Eng. 2022, 10, 1796. https://doi.org/10.3390/jmse10111796
Hong J, Fu Z, Hu J, Zhou S, Yu G, Ma Z. Dietary Curcumin Supplementation Enhanced Ammonia Nitrogen Stress Tolerance in Greater Amberjack (Seriola dumerili): Growth, Serum Biochemistry and Expression of Stress-Related Genes. Journal of Marine Science and Engineering. 2022; 10(11):1796. https://doi.org/10.3390/jmse10111796
Chicago/Turabian StyleHong, Jiawei, Zhengyi Fu, Jing Hu, Shengjie Zhou, Gang Yu, and Zhenhua Ma. 2022. "Dietary Curcumin Supplementation Enhanced Ammonia Nitrogen Stress Tolerance in Greater Amberjack (Seriola dumerili): Growth, Serum Biochemistry and Expression of Stress-Related Genes" Journal of Marine Science and Engineering 10, no. 11: 1796. https://doi.org/10.3390/jmse10111796
APA StyleHong, J., Fu, Z., Hu, J., Zhou, S., Yu, G., & Ma, Z. (2022). Dietary Curcumin Supplementation Enhanced Ammonia Nitrogen Stress Tolerance in Greater Amberjack (Seriola dumerili): Growth, Serum Biochemistry and Expression of Stress-Related Genes. Journal of Marine Science and Engineering, 10(11), 1796. https://doi.org/10.3390/jmse10111796