
Increased Temperature and Nitrogen Enrichment Inhibit the Growth of the Golden Tide Blooming Macroalgae Sargassum horneri in the Yellow Sea, China

Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Experimental Design
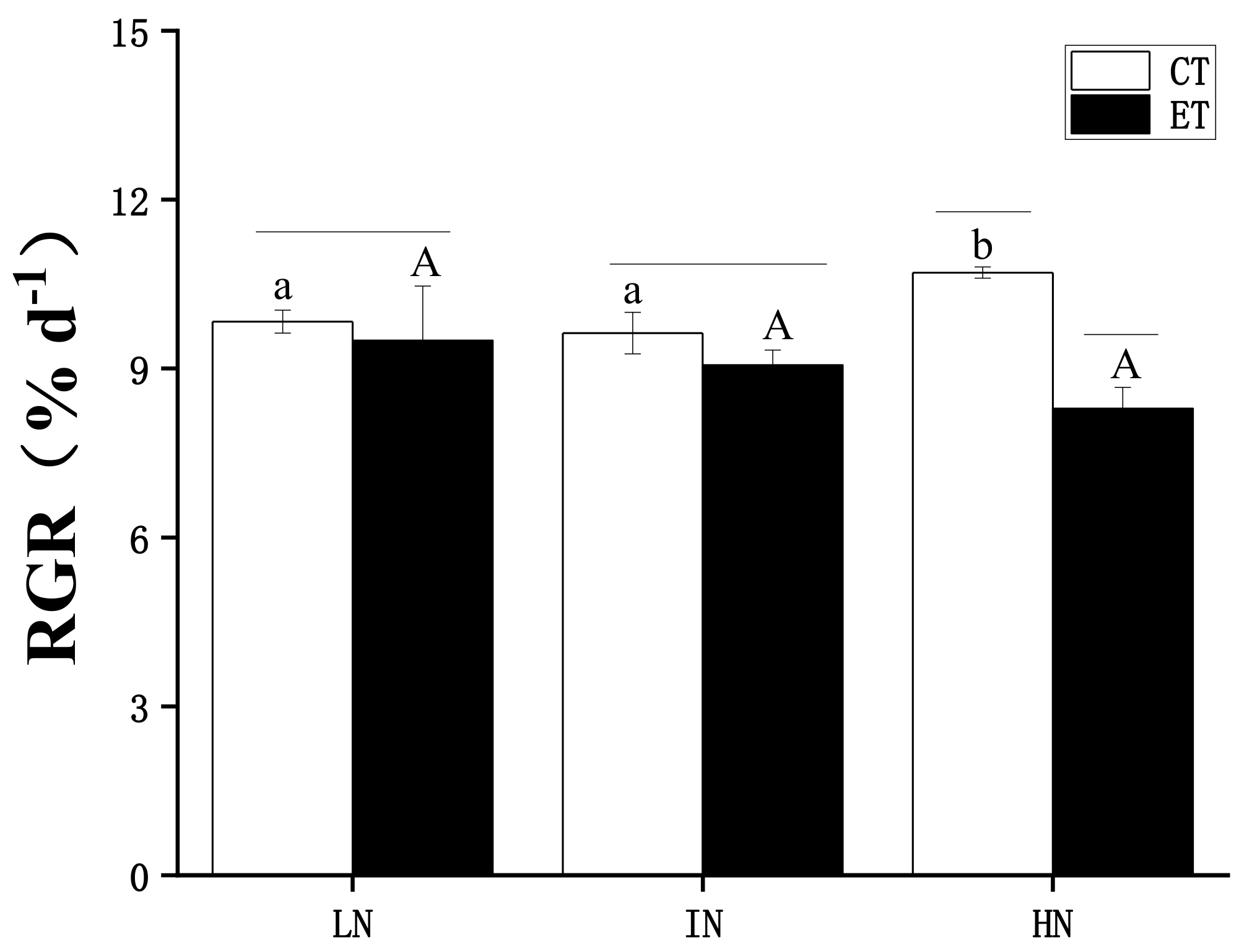
2.3. Measurement of Growth
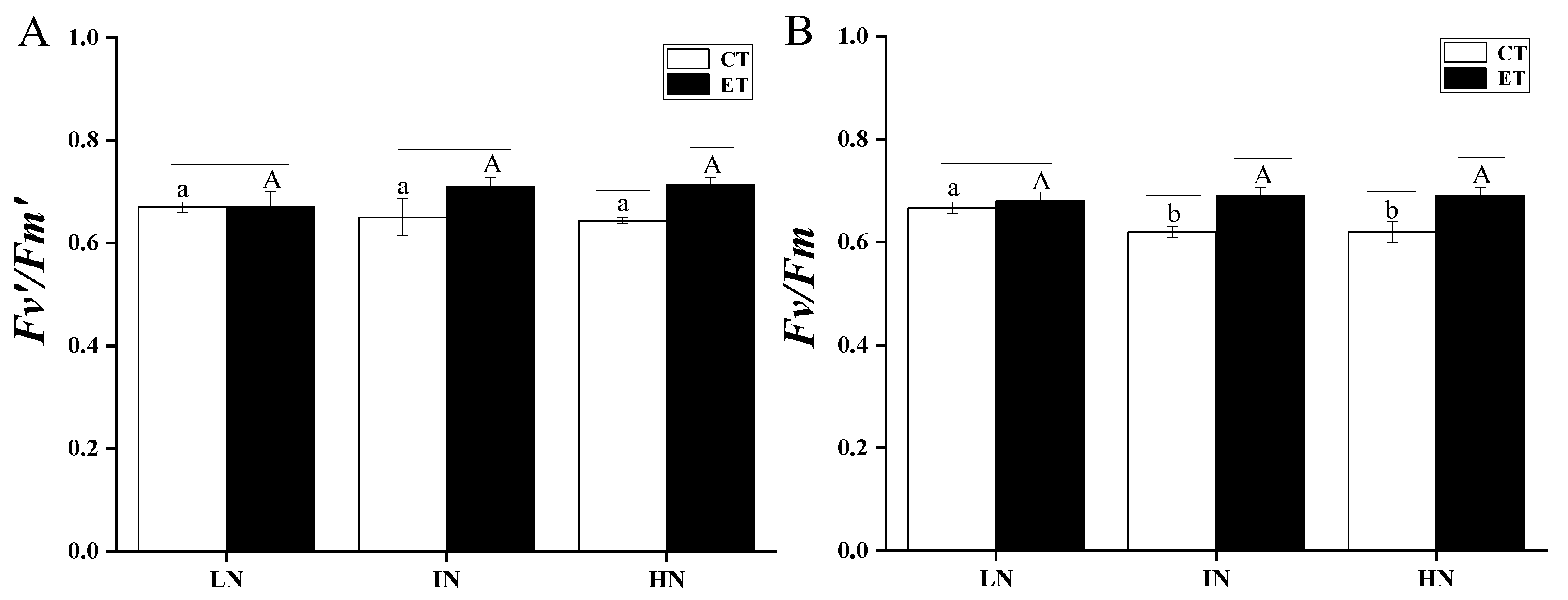
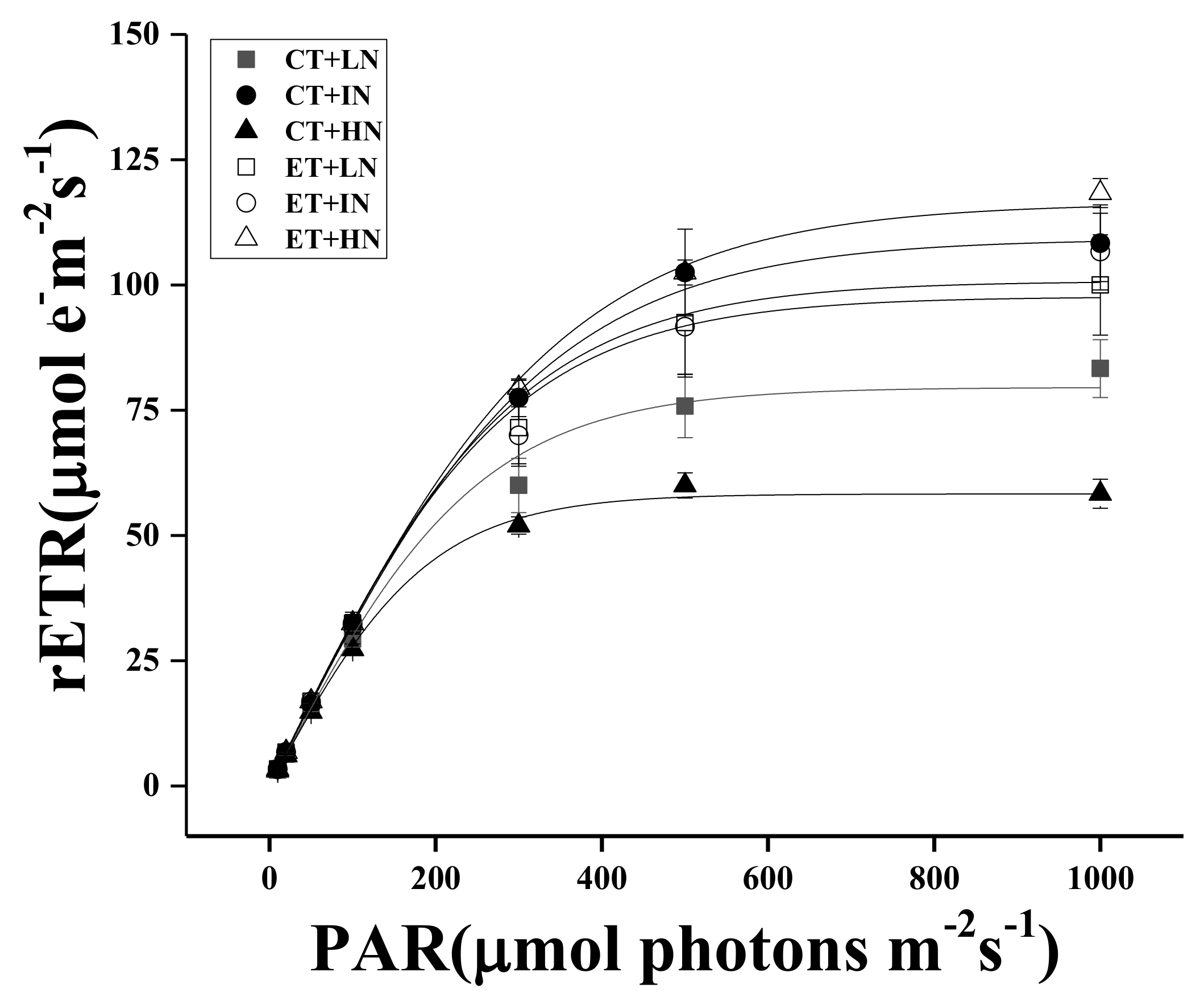
2.4. Chlorophyll Fluorescence Measurements
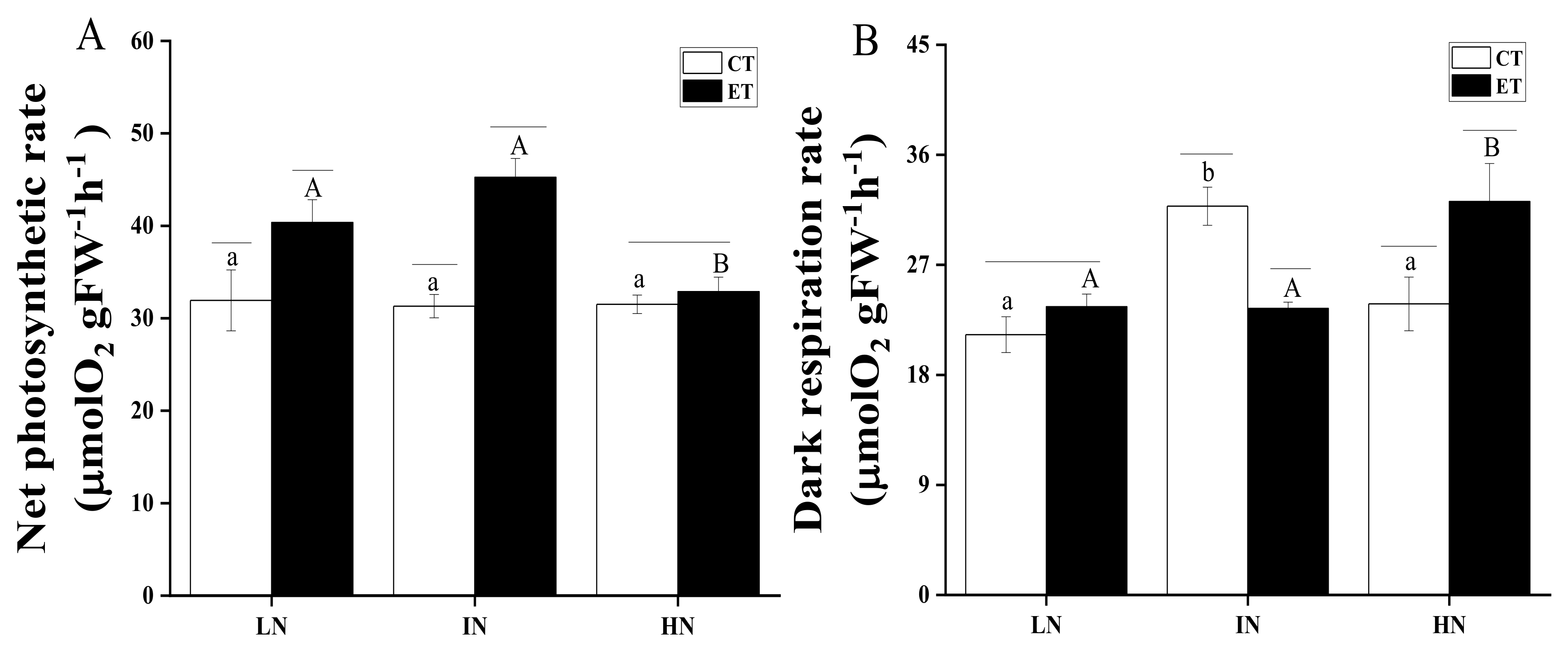
2.5. Measuring the Photosynthetic Rate
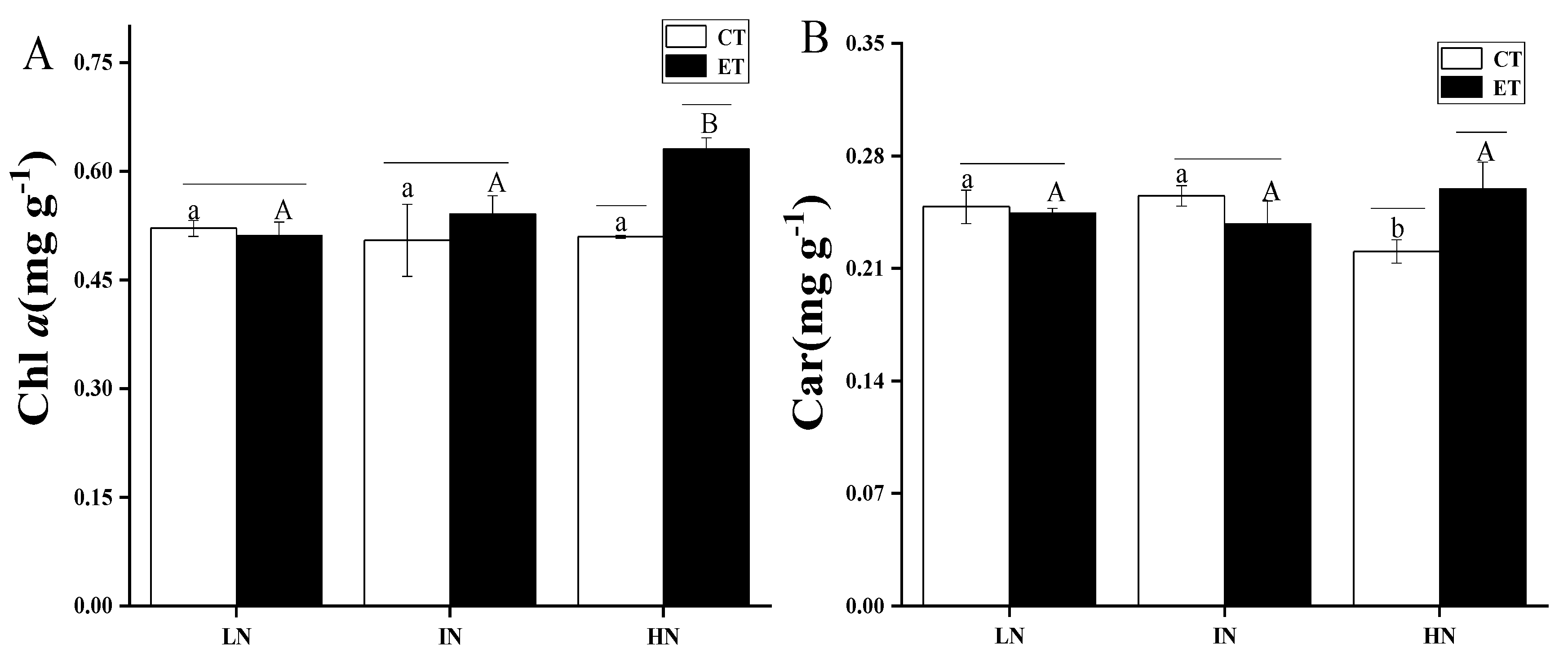
2.6. Pigments Measurement
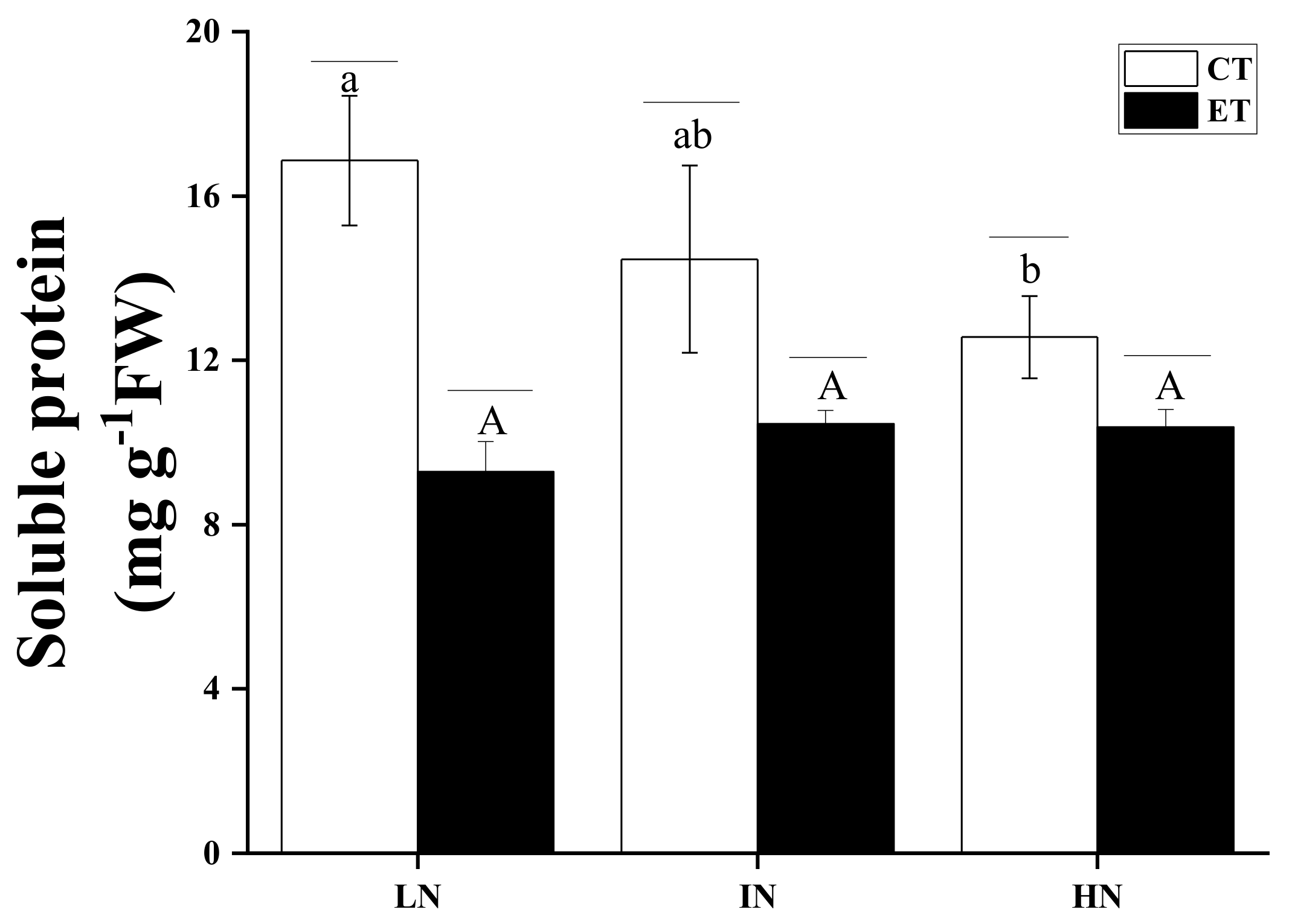
2.7. Soluble Protein Determination
2.8. Data Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fontela, M.; Velo, A.; Gilcoto, M.; Pérez, F. Anthropogenic CO2 and ocean acidification in Argentine Basin Water Masses over almost five decades of observations. Sci. Total Environ. 2021, 779, 146570. [Google Scholar] [CrossRef] [PubMed]
- Scheffer, M.; Schulz, K.G.; Bellerby, R.G.J.; Botros, M.; Fritsche, P.; Meyerhöfer, M.; Neill, C.; Nondal, G.; Oschlies, A.; Wohlers, J.; et al. Positive feedback between global warming and atmospheric CO2 concentration inferred from past climate change. Geophys. Res. Lett. 2006, 33, 229–237. [Google Scholar] [CrossRef]
- Riebesell, U.; Schulz, K.G.; Bellerby, R.G.J.; Botros, M.; Fritsche, P.; Meyerhoefer, M.; Neill, C.; Nondal, G.; Oschlies, A.; Wohlers, J. Enhanced biological carbon consumption in a high CO2 ocean. Nature 2007, 450, 545–548. [Google Scholar] [CrossRef] [PubMed]
- Feely, R.A.; Doney, S.C.; Cooley, S.R. Ocean acidification: Present conditions and future changes in a high-CO2 world. Oceanography 2009, 22, 36–47. [Google Scholar] [CrossRef]
- Gattuso, J.P.; Magnan, A.; Billé, R.; Cheung, W.W.; Howes, E.L.; Joos, F.; Allemand, D.; Bopp, L.; Cooley, S.R.; Eakin, C.M.; et al. Contrasting futures for ocean and society from different anthropogenic CO2 emissions scenarios. Science 2015, 349, aac4722. [Google Scholar] [CrossRef] [PubMed]
- Collins, M.; Knutti, R.; Arblaster, J.; Dufresne, J.L.; Fichefet, T.; Friedlingstein, P.; Gao, X.; Gutowski, W.J. Long-term climate change: Projections, commitments and irreversibility. Chapter 13; In Climate Change 2013: The Physical Science Basis; Cambridge University Press: London, UK, 2013; pp. 1029–1136. Available online: https://www.issuelab.org/resource/climate-change-2013-the-physical-science-basis.html (accessed on 15 June 2022).
- Wernberg, T.; Russell, B.D.; Moore, P.J.; Ling, S.D.; Smale, D.A.; Campbell, A.C.; Melinda, A.; Steinberg, P.D.; Kendrick, G.A.; Connell, S.D. Impacts of climate change in a global hotspot for temperate marine biodiversity and ocean warming. J. Exp. Mar. Biol. Ecol. 2011, 400, 7–16. [Google Scholar] [CrossRef]
- Kelaher, B.P.; Mamo, L.T.; Provost, E.; Litchfield, S.G.; Giles, A.; Butcherine, P. Influence of ocean warming and acidification on habitat-forming coralline algae and their associated molluscan assemblages. Glob. Ecol. Conserv. 2022, 35, e02081. [Google Scholar] [CrossRef]
- Barbier, E.B.; Hacker, S.D.; Kennedy, C.; Koch, E.W.; Stier, A.C.; Silliman, B.R. The value of estuarine and coastal ecosystem services. Ecol. Monogr. 2011, 81, 169–193. [Google Scholar] [CrossRef]
- Chung, I.K.; Beardall, J.; Mehta, S.; Sahoo, D.; Stojkovic, S. Using marine macroalgae for carbon sequestration: A critical appraisal. J. Appl. Phycol. 2010, 23, 877–886. [Google Scholar] [CrossRef]
- Arina, N.; Raynusha, C.; Hidayah, N.; Zainee, N.F.A.; Prathep, A.; Rozaimi, M. Coralline macroalgae contribution to ecological services of carbon storage in a disturbed seagrass meadow. Mar. Environ. Res. 2020, 162, 105156. [Google Scholar] [CrossRef]
- Krause-Jensen, D.; Duarte, C.M. Substantial role of macroalgae in marine carbon sequestration. Nat. Geosci. 2016, 9, 737–742. [Google Scholar] [CrossRef]
- Sondak, C.F.A.; Ang, P.O.; Beardall, J.; Bellgrove, A.; Boo, S.M.; Gerung, G.S.; Hepburn, C.D.; Hong, D.D.; Hu, Z.Y.; Kawai, H.; et al. Carbon dioxide mitigation potential of seaweed aquaculture beds (SABs). J. Appl. Phycol. 2016, 29, 2363–2373. [Google Scholar] [CrossRef]
- Yong, W.T.L.; Thien, V.Y.; Rupert, R.; Rodrigues, K.F. Seaweed: A potential climate change solution. Renew. Sust. Energ. Rev. 2022, 159, 112222. [Google Scholar] [CrossRef]
- Wu, H.L.; Chen, J.; Feng, J.C.; Liu, Y.H.; Li, X.B.; Chen, R.; Xu, J.T. Thermal fluctuations and nitrogen enrichment synergistically accelerate biomass yield of Pyropia haitanensis. Aquat. Bot. 2022, 179, 103501. [Google Scholar] [CrossRef]
- Wu, H.L.; Liu, Y.M.; Beardall, J.; Zhong, Z.H.; Gao, G.; Xu, J.T. Physiological acclimation of Ulva prolifera to seasonal environmental factors drives green tides in the Yellow Sea. Mar. Environ. Res. 2022, 179, 105695. [Google Scholar] [CrossRef]
- Boyce, D.G.; Lewis, M.R.; Worm, B. Global phytoplankton decline over the past century. Nature 2010, 466, 591–596. [Google Scholar] [CrossRef]
- Thomas, M.K.; Kremer, C.T.; Klausmeier, C.A.; Litchman, E. A global pattern of thermal adaptation in marine phytoplankton. Science 2012, 338, 1085–1088. [Google Scholar] [CrossRef]
- Bopp, L.; Monfray, P.; Aumont, O.; Belviso, S. Potential impact of climate change on marine export production. Global Biogeochem. Cycles 2001, 15, 81–99. [Google Scholar] [CrossRef]
- Steinacher, M.; Joos, F.; Frölicher, T.L.; Bopp, L. Projected 21st century decrease in marine productivity: A multi-model analysis. Biogeosciences 2009, 7, 979–1005. [Google Scholar] [CrossRef]
- Figueroa, F.L.; Barufi, J.B.; Malta, E.J.; Condeálvarez, R.; Nitschke, U.; Arenas, F.; Mata, M.; Connan, S.; Abreu, M.H.; Marquardt, R. Short-term effects of increasing CO2, nitrate and temperature on three Mediterranean macroalgae: Biochemical composition. Aquat. Biol. 2014, 22, 177–193. [Google Scholar] [CrossRef]
- Sobrino, C.; Neale, P.J. Short-term and long-term effects of temperature on photosynthesis in the diatom Thalassiosira pseudonana under UVR exposures. J. Phycol. 2007, 43, 426–436. [Google Scholar] [CrossRef]
- Thomas, M.K.; Aranguren-Gassis, M.; Kremer, C.T.; Gould, M.R.; Anderson, K.; Klausmeier, C.A.; Litchman, E. Temperature-nutrient interactions exacerbate sensitivity to warming in phytoplankton. Global Change Biol. 2017, 23, 3269–3280. [Google Scholar] [CrossRef] [PubMed]
- Schmid, M.; Guihéneuf, F.; Nitschke, U.; Stengel, D.B. Acclimation potential and biochemical response of four temperate macroalgae to light and future seasonal temperature scenario. Algal Res. 2021, 54, 102190. [Google Scholar] [CrossRef]
- Litchman, E.; Pinto, P.D.T.; Klausmeier, C.A.; Thomas, M.K.; Yoshiyama, K. Linking traits to species diversity and community structure in phytoplankton. Hydrobiologia 2010, 653, 15–28. [Google Scholar] [CrossRef]
- Xiong, Y.L.; Gao, L.; Qu, L.Y.; Xu, J.T.; Ma, Z.L.; Gao, G. The contribution of fish and seaweed mariculture to the coastal fluxes of biogenic elements in two important aquaculture areas, China. Sci. Total Environ. 2023, 856, 159056. [Google Scholar] [CrossRef] [PubMed]
- Rabalais, N.N.; Turner, R.E.; Díaz, R.J.; Justić, D. Global change and eutrophication of coastal waters. ICES J. Mar. Sci. 2009, 66, 1528–1537. [Google Scholar] [CrossRef]
- Shahar, B.; Guttman, L. Integrated biofilters with Ulva and periphyton to improve nitrogen removal from mariculture effluent. Aquaculture 2021, 532, 736011. [Google Scholar] [CrossRef]
- Zhang, C.; Lu, J.; Wu, J. Enhanced removal of phenolic endocrine disrupting chemicals from coastal waters by intertidal macroalgae. J. Hazard. Mater. 2021, 411, 125105. [Google Scholar] [CrossRef]
- Li, P.P.; Chen, H.H.; Zhang, J.Y.; Feng, X.Q.; Xiao, B.H.; Hu, Y.Y.; Sui, Z.H. Effects of nutrient deficiency on the branch phenotype of the macroalgae Gracilariopsis lemaneiformis (Rhodophyta). Aquaculture 2023, 562, 738794. [Google Scholar] [CrossRef]
- Zhou, W.; Wu, H.; Huang, J.J.; Wang, J.G.; Zhen, W.; Wang, J.W.; Ni, J.X.; Xu, J.T. Elevated-CO2 and nutrient limitation synergistically reduce the growth and photosynthetic performances of a commercial macroalga Gracilariopsis lemaneiformis. Aquaculture 2022, 550, 737878. [Google Scholar] [CrossRef]
- Luo, M.B.; Liu, F.; Xu, Z.L. Growth and nutrient uptake capacity of two co-occurring species, Ulva prolifera and Ulva linza. Aquat. Bot. 2012, 100, 18–24. [Google Scholar] [CrossRef]
- Fan, X.; Xu, D.; Wang, Y.T.; Zhang, X.W.; Cao, S.N.; Mou, S.L.; Ye, N.H. The effect of nutrient concentrations, nutrient ratios and temperature on photosynthesis and nutrient uptake by Ulva prolifera: Implications for the explosion in green tides. J. Appl. Phycol. 2013, 26, 537–544. [Google Scholar] [CrossRef]
- Li, H.M.; Zhang, Y.Y.; Han, X.R.; Shi, X.Y.; Rivkin, R.B.; Legendre, L. Growth responses of Ulva prolifera to inorganic and organic nutrients: Implications for macroalgal blooms in the southern Yellow Sea, China. Sci. Rep. 2016, 6, 26498. [Google Scholar] [CrossRef] [PubMed]
- Smetacek, V.; Zingone, A. Green and golden seaweed tides on the rise. Nature 2013, 504, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, M.M.; Liu, J.L.; Ding, X.W.; He, J.Z.; Zhao, S.; Wu, L.J.; Gao, S.; Zhao, C.Y.; Liu, D.Y.; Zhang, J.H.; et al. Sargassum blooms in the East China Sea and Yellow Sea: Formation and management. Mar. Pollut. Bull. 2021, 162, 111845. [Google Scholar] [CrossRef] [PubMed]
- Pang, S.J.; Liu, F.; Shan, T.F.; Gao, S.Q.; Zhang, Z.H. Cultivation of the brown alga Sargassum horneri: Sexual reproduction and seedling production in tank culture under reduced solar irradiance in ambient temperature. J. Appl. Phycol. 2009, 21, 413–422. [Google Scholar] [CrossRef]
- Yoon, J.T.; Sun, S.M.; Chung, G. Sargassum bed restoration by transplantation of germlings grown under protective mesh cage. J. Appl. Phycol. 2014, 26, 505–509. [Google Scholar] [CrossRef]
- Su, L.; Shan, T.F.; Pang, S.J.; Li, J. Analyses of the genetic structure of Sargassum horneri in the Yellow Sea: Implications of the temporal and spatial relations among floating and benthic populations. J. Appl. Phycol. 2018, 30, 1417–1424. [Google Scholar] [CrossRef]
- Lapointe, B.E. A comparison of nutrient-limited productivity in Sargassum natans from neritic vs. oceanic waters of the western North Atlantic Ocean. Limnol. Oceanogr. 1995, 40, 625–633. [Google Scholar] [CrossRef]
- Ohtake, M.; Natori, N.; Sugai, Y.; Tsuchiya, K.; Aketo, T.; Nishihara, G.N.; Toda, T. Growth and nutrient uptake characteristics of Sargassum macrocarpum cultivated with phosphorus-replete wastewater. Aquat. Bot. 2020, 163, 103208. [Google Scholar] [CrossRef]
- Bao, M.L.; Park, J.S.; Wu, H.L.; Lee, H.J.; Park, S.R.; Kim, T.H.; Son, Y.B.; Lee, T.H.; Yarish, C.; Kim, J.K. A comparison of physiological responses between attached and pelagic populations of Sargassum horneri under nutrient and light limitation. Mar. Environ. Res. 2022, 173, 105544. [Google Scholar] [CrossRef] [PubMed]
- Falkowski, P.G.; Barber, R.T.; Smetacek, V. Biogeochemical controls and feedbacks on ocean primary production. Science 1998, 281, 200–206. [Google Scholar] [CrossRef]
- Elser, J.J.; Bracken, M.E.S.; Cleland, E.E.; Gruner, D.S.; Harpole, W.S.; Hillebrand, H.; Ngai, J.T.; Seabloom, E.W.; Shurin, J.B.; Smith, J.E. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett. 2007, 10, 1135–1142. [Google Scholar] [CrossRef] [PubMed]
- Hee, Y.Y.; Suratman, S.; Weston, K. Nutrient cycling and primary production in Peninsular Malaysia waters; regional variation and its causes in the South China Sea. Estuar. Coast. Shelf Sci. 2020, 245, 106947. [Google Scholar] [CrossRef]
- Boyd, P.W.; Rynearson, T.A.; Armstrong, E.A.; Fu, F.X.; Hayashi, K.; Hu, Z.X.; Hutchins, D.A.; Kudela, R.M.; Litchman, E.; Mulholland, M.R.; et al. Marine phytoplankton temperature versus growth responses from polar to tropical waters—Outcome of a scientific community-wide study. PLoS ONE 2013, 8, e63091. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.K.; Kremer, C.T.; Litchman, E. Environment and evolutionary history determine the global biogeography of phytoplankton temperature traits. Global Ecol. Biogeogr. 2016, 25, 75–86. [Google Scholar] [CrossRef]
- IPCC. Climate Change, 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2014; p. 151. [Google Scholar]
- Genty, B.; Briantais, J.M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta Gen. Subj. 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Jassby, A.D.; Platt, T. Mathematical formulation of the relationship between photosynthesis and light for phytoplankton. Limnol. Oceanogr. 1976, 21, 540–547. [Google Scholar] [CrossRef]
- Xu, Z.G.; Gao, G.; Xu, J.T.; Wu, H.L. Physiological response of a golden tide alga (Sargassum muticum) to the interaction of ocean acidification and phosphorus enrichment. Biogeosciences 2017, 14, 671–681. [Google Scholar] [CrossRef]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Kochert, G. Protein Determination by Dye Binding; Cambridge University Press: London, UK, 1978; pp. 91–93. [Google Scholar] [CrossRef]
- Nishihara, G.N.; Noro, T.; Terada, R. Effect of temperature and light on the photosynthetic performance of two edible seaweeds: Meristotheca coacta Okamura and Meristotheca papulosa J. Agardh (Solieriaceae, Rhodophyta). Aquac. Sci. 2012, 60, 377–388. [Google Scholar] [CrossRef]
- Kim, J.K.; Yarish, C.; Pereira, R. Tolerances to hypo-osmotic and temperature stresses in native and invasive species of Gracilaria (Rhodophyta). Phycologia 2019, 55, 257–264. [Google Scholar] [CrossRef]
- Cui, J.J.; Zhang, J.H.; Huo, Y.Z.; Zhou, L.J.; Wu, Q.; Chen, L.P.; Yu, K.F.; He, P.M. Adaptability of free-floating green tide algae in the Yellow Sea to variable temperature and light intensity. Mar. Pollut. Bull. 2015, 101, 660–666. [Google Scholar] [CrossRef]
- Wu, H.L.; Zhang, J.H.; Yarish, C.; He, P.M.; Kim, J.K. Bioremediation and nutrient migration during blooms of Ulva in the Yellow Sea, China. Phycologia 2018, 57, 223–231. [Google Scholar] [CrossRef]
- Zhou, Z.X.; Yu, R.C.; Zhou, M.J. Evolution of harmful algal blooms in the East China Sea under eutrophication and warming scenarios. Water Res. 2022, 221, 118807. [Google Scholar] [CrossRef]
- Wang, Y.; Zhong, Z.H.; Qin, S.; Li, J.L.; Liu, Z.Y. Effects of temperature and light on growth rate and photosynthetic characteristics of Sargassum horneri. J. Ocean Univ. China 2021, 20, 101–110. [Google Scholar] [CrossRef]
- Zhang, P.; Wang, T.; Xie, Q.; Zhang, H.; Yan, X.; Chen, M. Effects of environment factors on growth of Sargassum horneri seedlings under indoor conditions. J. Shanghai Ocean Uni. 2014, 23, 200–207. Available online: https://www.shhydxxb.com/shhy/ch/reader/create_pdf.aspx?file_no=20130800833&year_id=2014&quarter_id=2&falg=1 (accessed on 21 September 2022).
- Wu, H.L.; Feng, J.C.; Li, X.S.; Zhao, C.Y.; Liu, Y.H.; Xu, J.T. Effects of increased CO2 and temperature on the physiological characteristics of the golden tide blooming macroalgae Sargassum horneri in the Yellow Sea, China. Mar. Pollut. Bull. 2019, 146, 639–644. [Google Scholar] [CrossRef] [PubMed]
- Padfield, D.; Lowe, C.; Buckling, A.; Ffrench-Constant, R.; Jennings, S.; Shelley, F.; Ólafsson, J.S.; Yvon-Durocher, G. Metabolic compensation constrains the temperature dependence of gross primary production. Ecol. Lett. 2017, 20, 1250–1260. [Google Scholar] [CrossRef] [PubMed]
- Marañón, E.; Lorenzo, M.P.; Cermeño, P.; Mouriño-Carballido, B. Nutrient limitation suppresses the temperature dependence of phytoplankton metabolic rates. ISME J. 2018, 12, 1836. [Google Scholar] [CrossRef]
- Deutsch, C.A.; Tewksbury, J.J.; Huey, R.B.; Sheldon, K.S.; Ghalambor, C.K.; Haak, D.C.; Martin, P.R. Impacts of climate warming on terrestrial ectotherms across latitude. Proc. Natl. Acad. Sci. USA 2008, 105, 6668–6672. [Google Scholar] [CrossRef] [PubMed]
- Martin, T.L.; Huey, R.B. Why “suboptimal” is optimal: Jensen's inequality and ectotherm thermal preferences. Am. Nat. 2008, 171, E102–E118. [Google Scholar] [CrossRef]
- Mandal, S.; Islam, M.S.; Biswas, M.H.; Akter, S. A mathematical model applied to investigate the potential impact of global warming on marine ecosystems. Appl. Math. Model. 2022, 101, 19–37. [Google Scholar] [CrossRef]
- Lobban, C.S.; Harrison, P.J.; Harrison, P.J. Seaweed Ecology and Physiology; Cambridge University Press: London, UK, 1994; p. 366. [Google Scholar] [CrossRef]
- Bews, E.; Booher, L.; Polizzi, T.; Long, C.; Kim, J.H.; Edwards, M.S. Effects of salinity and nutrients on metabolism and growth of Ulva lactuca: Implications for bioremediation of coastal watersheds. Mar. Pollut. Bull. 2021, 166, 112199. [Google Scholar] [CrossRef] [PubMed]
- Lapointe, B.E. Phosphorus-limited photosynthesis and growth of Sargassum natans and Sargassum fluitans (Phaeophyceae) in the western North Atlantic. Deep Sea Res. Part A Oceanogr. Res. Pap. 1986, 33, 391–399. [Google Scholar] [CrossRef]
- Leung, J.Y.S.; Russell, B.D.; Coleman, M.A.; Kelaher, B.K.; Connell, S.D. Long-term thermal acclimation drives adaptive physiological adjustments of a marine gastropod to reduce sensitivity to climate change. Sci. Total Environ. 2021, 771, 145208. [Google Scholar] [CrossRef]
- Zou, D.H.; Gao, K.S. The photosynthetic and respiratory responses to temperature and nitrogen supply in the marine green macroalga Ulva conglobata (Chlorophyta). Phycologia 2014, 53, 86–94. [Google Scholar] [CrossRef]
- Zou, D.H.; Gao, K.S. Temperature response of photosynthetic light- and carbon-use characteristics in the red seaweed Gracilariopsis lemaneiformis (Gracilariales, Rhodophyta). J. Phycol. 2014, 50, 366–375. [Google Scholar] [CrossRef]
- Dawes, C.J.; Koch, E.W. Physiological responses of the red algae Gracllarla verrucosa and G. tikvahiae before and after nutrient enrichment. Bull. Mar. Sci. 1990, 46, 335–344. Available online: https://www.ingentaconnect.com/content/umrsmas/bullmar/1990/00000046/00000002/art00007 (accessed on 10 August 2022).
- Crawford, N.M. Nitrate: Nutrient and signal for plant growth. Plant Cell 1995, 7, 859–868. [Google Scholar] [CrossRef][Green Version]
- Liu, L.; Zou, D.H.; Jiang, H.; Chen, B.B.; Zeng, X.P. Effects of increased CO2 and temperature on the growth and photosynthesis in the marine macroalga Gracilaria lemaneiformis from the coastal waters of South China. J. Appl. Phycol. 2017, 30, 1271–1280. [Google Scholar] [CrossRef]
- Alcoverro, T.; Cerbiān, E.; Ballesteros, E. The photosynthetic capacity of the seagrass Posidonia oceanica: Influence of nitrogen and light. J. Appl. Phycol. 2001, 30, 1271–1280. [Google Scholar] [CrossRef]
- Fork, D.C.; Murata, N.; Sato, N. Effect of growth temperature on the lipid and fatty acid composition, and the dependence on temperature of light-induced redox reactions of cytochrome f and of light energy redistribution in the thermophilic blue-green alga Synechococcus lividus. Plant Physiol. 1979, 63, 524–530. [Google Scholar] [CrossRef] [PubMed]
- Sendall, K.M.; Reich, P.B.; Zhao, C.; Hou, J.; Wei, X.; Stefanski, A.; Rice, K.; Rich, R.L.; Montqomery, R.A. Acclimation of photosynthetic temperature optima of temperate and boreal tree species in response to experimental forest warming. Global Change Biol. 2015, 21, 1342–1357. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, D.R.; Hamman, C.R.; Johnson, E.C.; Kremer, C.T.; Klausmeier, C.A.; Litchman, E. Rapid thermal adaptation in a marine diatom reveals constraints and trade-offs. Global Change Biol. 2018, 24, 4554–4565. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | RGR (% d−1) | ||
|---|---|---|---|
| DF | F-Value | p-Value | |
| Temperature | 1 | 24.68844 | <0.001 |
| N concentration | 2 | 0.69394 | 0.51858 |
| Temperature*N concentration | 2 | 8.72092 | 0.00458 |
| Source | DF | F-Value | p-Value |
|---|---|---|---|
| Fv’/Fm’ | |||
| Temperature | 1 | 17.68605 | 0.00122 |
| N concentration | 2 | 0.36047 | 0.70465 |
| Temperature*N concentration | 2 | 4.5 | 0.03482 |
| Fv/Fm | |||
| Temperature | 1 | 46 | <0.001 |
| N concentration | 2 | 2.63043 | 0.1129 |
| Temperature*N concentration | 2 | 6.28261 | 0.01359 |
| Treatments | rETRmax μmol e− m−2 s−1 | α | Ik μmol Photos m−2 s−1 |
|---|---|---|---|
| CT + LN | 82.19 ± 6.17 a | 0.27 ± 0.02 a | 299.58 ± 17.14 a |
| CT + IN | 110.17 ± 8.64 b | 0.33 ± 0.01 b | 331.61 ± 13.53 a* |
| CT + HN | 59.14 ± 2.49 c* | 0.29 ± 0.01 b* | 202.46 ± 9.30 b* |
| ET + LN | 100.28 ± 11.11 A | 0.31 ± 0.03 A | 318.82 ± 24.23 A |
| ET + IN | 106.03 ± 8.76 A | 0.29 ± 0.02 A | 362.15 ± 12.89 B* |
| ET + HN | 118.59 ± 2.96 A* | 0.32 ± 0.01 A* | 368.80 ± 3.42 B* |
| Source | DF | F-Value | p-Value |
|---|---|---|---|
| rETRmax | |||
| Temperature | 1 | 49.34006 | <0.001 |
| N concentration | 2 | 12.08173 | 0.00133 |
| Temperature*N concentration | 2 | 28.61007 | <0.001 |
| α | |||
| Temperature | 1 | 1.11486 | 0.31182 |
| N concentration | 2 | 1.18812 | 0.33823 |
| Temperature*N concentration | 2 | 6.75108 | 0.01085 |
| Ik | |||
| Temperature | 1 | 105.50349 | <0.001 |
| N concentration | 2 | 25.87745 | <0.001 |
| Temperature*N concentration | 2 | 45.41122 | <0.001 |
| Source | DF | F-Value | p-Value |
|---|---|---|---|
| NPR | |||
| Temperature | 1 | 64.89353 | <0.001 |
| N concentration | 2 | 13.19405 | <0.001 |
| Temperature*N concentration | 2 | 13.71753 | <0.001 |
| DRR | |||
| Temperature | 1 | 0.79293 | <0.001 |
| N concentration | 2 | 17.09959 | <0.001 |
| Temperature*N concentration | 2 | 31.733 | <0.001 |
| Source | DF | F-Value | p-Value |
|---|---|---|---|
| Chl α | |||
| Temperature | 1 | 17.01619 | 0.00141 |
| N concentration | 2 | 8.08644 | 0.00597 |
| Temperature*N concentration | 2 | 10.29292 | 0.00249 |
| Car | |||
| Temperature | 1 | 1.33014 | 0.27124 |
| N concentration | 2 | 0.71039 | 0.511 |
| Temperature*N concentration | 2 | 11.3529 | 0.00171 |
| Source | DF | F-Value | p-Value |
|---|---|---|---|
| SP | |||
| Temperature | 1 | 59.90949 | <0.001 |
| N concentration | 2 | 2.49521 | 0.12413 |
| Temperature*N concentration | 2 | 7.11245 | 0.00918 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, H.; Li, X.; Liu, Y.; Wang, C.; Ji, C.; Xu, J. Increased Temperature and Nitrogen Enrichment Inhibit the Growth of the Golden Tide Blooming Macroalgae Sargassum horneri in the Yellow Sea, China. J. Mar. Sci. Eng. 2022, 10, 1692. https://doi.org/10.3390/jmse10111692
Wu H, Li X, Liu Y, Wang C, Ji C, Xu J. Increased Temperature and Nitrogen Enrichment Inhibit the Growth of the Golden Tide Blooming Macroalgae Sargassum horneri in the Yellow Sea, China. Journal of Marine Science and Engineering. 2022; 10(11):1692. https://doi.org/10.3390/jmse10111692
Chicago/Turabian StyleWu, Hailong, Xuebing Li, Yanhong Liu, Chuchu Wang, Chenkai Ji, and Juntian Xu. 2022. "Increased Temperature and Nitrogen Enrichment Inhibit the Growth of the Golden Tide Blooming Macroalgae Sargassum horneri in the Yellow Sea, China" Journal of Marine Science and Engineering 10, no. 11: 1692. https://doi.org/10.3390/jmse10111692
APA StyleWu, H., Li, X., Liu, Y., Wang, C., Ji, C., & Xu, J. (2022). Increased Temperature and Nitrogen Enrichment Inhibit the Growth of the Golden Tide Blooming Macroalgae Sargassum horneri in the Yellow Sea, China. Journal of Marine Science and Engineering, 10(11), 1692. https://doi.org/10.3390/jmse10111692