How Does Water-Stressed Corn Respond to Potassium Nutrition? A Shoot-Root Scale Approach Study under Controlled Conditions

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Preparation
2.2. Experimental Design: K Fertilization and Watering Modalities
2.3. Leaf Parameters and Leaf Area Calculations
2.4. Continuous Shoot Fresh and Dry Matter Simulations
2.5. A Water Mass Balance to Calculate Soil Water Content and Transpiration
2.6. Long-Term Water Use Efficiency
2.7. Root Measurements
2.8. Analysis of the Effects of Water Stress, Potassium Deficiency, and Their Combined Effects
2.9. Statistical Analysis
3. Results
3.1. Plant K Status
3.2. Plant Water Status
3.3. Plant Development
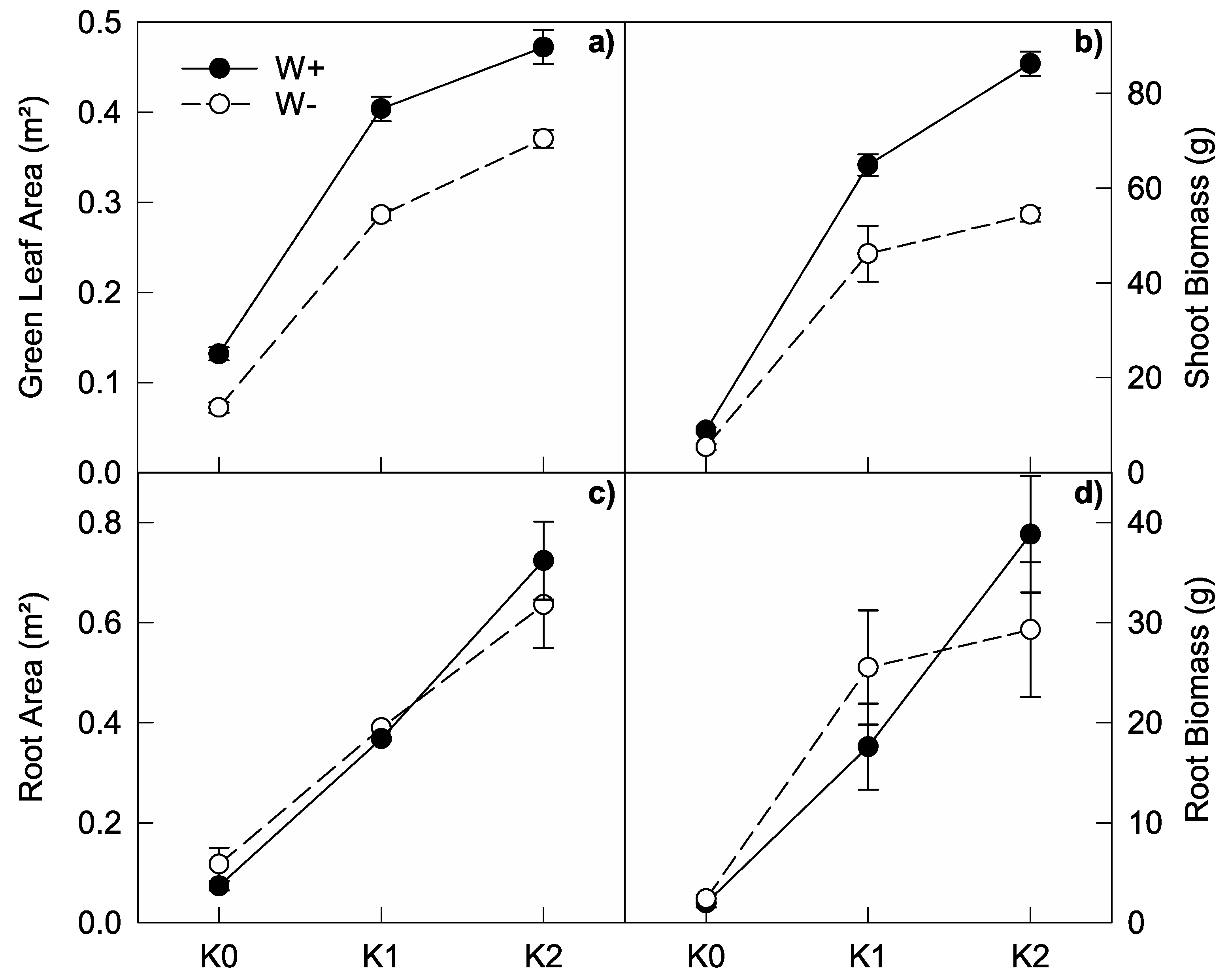
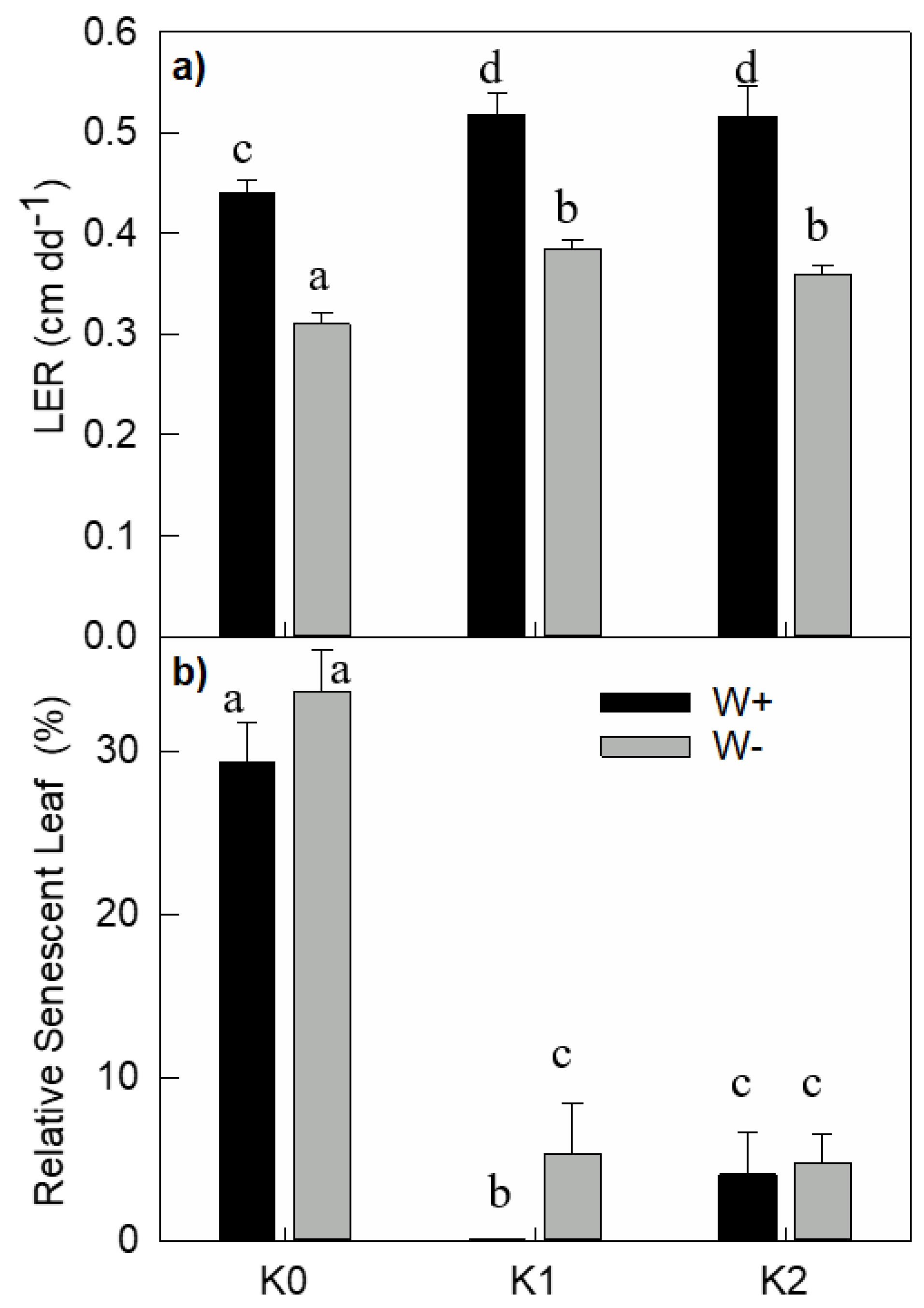
3.4. Leaf and Root Growth
3.5. Leaf Senescence
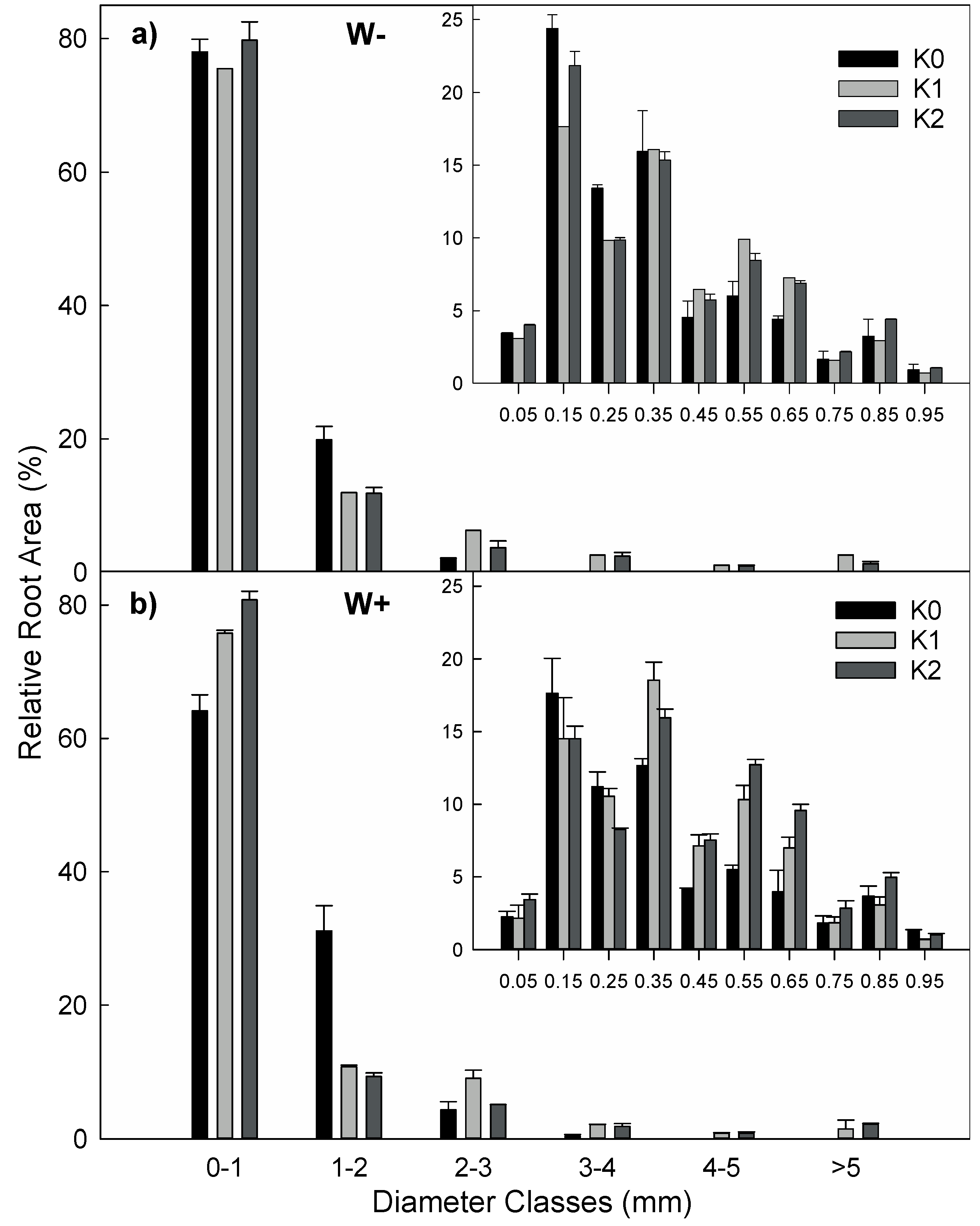
3.6. Morphology and Architecture at Organ and Plant Scales
3.7. Effects of Water and K Treatments on Water Flux and Use Efficiency
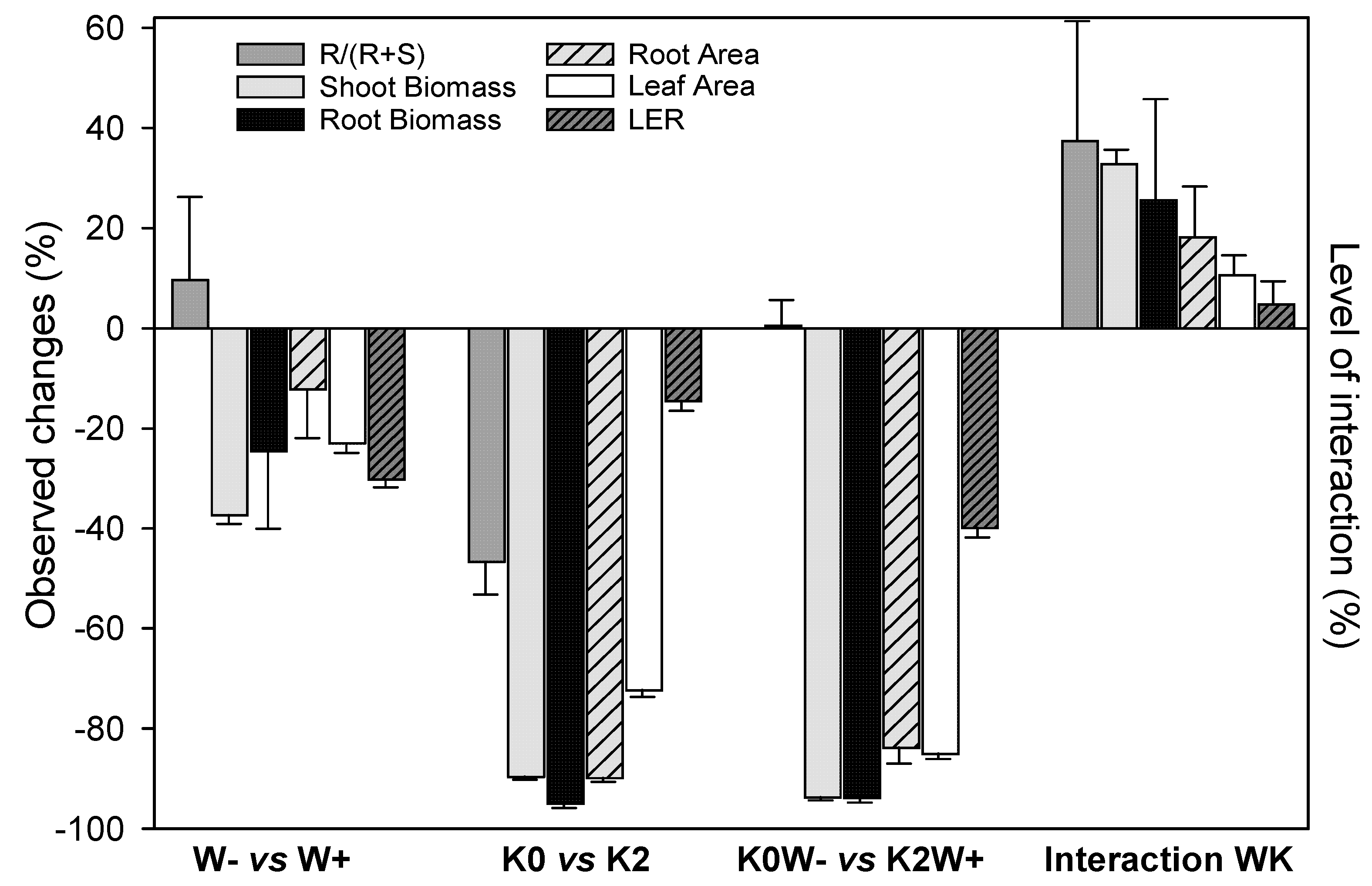
3.8. Water and Potassium Interactions on Morphological Traits
4. Discussion
4.1. Water and Potassium Status in Plants
4.2. Experimental Biases
4.3. Contrasted Contributions of Water and K on Corn Development, Growth, and Water Use
4.4. Finally, What Did We Learn about the Effects of K Nutrition on Drought Resistance?
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Çakir, R. Effect of water stress at different development stages on vegetative and reproductive growth of corn. Field Crop Res. 2004, 89, 1–16. [Google Scholar] [CrossRef]
- Martineau, E.; Domec, J.C.; Bosc, A.; Pascal, D.; Asensio-Fandino, V.; Lavres, J.; Jordan-Meille, L. The effects of potassium nutrition on water use in field-grown maize (Zea mays L.). Environ. Exp. Bot. 2017, 134, 62–71. [Google Scholar] [CrossRef]
- Ma, L.; Ahuja, L.R.; Islam, A.; Trout, T.J.; Saseendran, S.A.; Malone, R.W. Modeling yield and biomass responses of maize cultivars to climate change under full and deficit irrigation. Agric. Water Manag. 2017, 180, 88–98. [Google Scholar] [CrossRef]
- Lehner, B.; Döll, P.; Alcamo, J.; Henrichs, T.; Kaspar, F. Estimating the impact of global change on flood and drought risks in Europe: A continental, integrated analysis. Clim. Chang. 2006, 75, 273–299. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2014: Synthesis Report: Contribution of Working Groups I, II and III to the 5th 262 Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2014. [Google Scholar]
- Amigues, J.P.; Debaeke, P.; Itier, B.; Lemaire, G.; Seguin, B.; Tardieu, F.; Thomas, A. Sécheresse et Agriculture. Réduire la Vulnérabilité de L’agriculture à un Risque Accru de Manque D’eau; Expertise Scientifique Collective; INRA: Paris, France, 2006; pp. 37–45. [Google Scholar]
- Lobell, D.B.; Schlenker, W.; Costa-Roberts, J. Climate trends and global crop production since 1980. Science 2011, 333, 616–620. [Google Scholar] [CrossRef] [PubMed]
- Fanaei, H.R.; Galavi, M.; Kafi, M.; Ghanbari Bonjar, A. Amelioration of water stress by potassium fertilizer in two oilseed species. Int. J. Plant Prod. 2009, 3, 41–54. [Google Scholar]
- Aslam, M.; Zamir, I.; Afzal, I.; Yaseen, M.; Mubeen, M.; Shoaib, A. Drought stress, its effect on maize production and development of drought tolerance through potassium application. Cercetări Agronomice În Moldov 2013, 46, 99–114. [Google Scholar]
- Eyshi Rezaei, E.; Gaiser, T.; Siebert, S.; Sultan, B.; Ewert, F. Combined impacts of climate and nutrient fertilisation on yields of pearl millet in Niger. Eur. J. Agron. 2014, 55, 77–88. [Google Scholar] [CrossRef]
- Chaves, M.M.; Pereira, J.S.; Maroco, J.; Rodrigues, M.L.; Ricardo, C.P.; Osório, M.L.; Carvalho, I.; Faria, T.; Pinheiro, C. How plants cope with water stress in the field. Photosynthesis and growth. Ann. Bot. 2002, 89, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Carroll, M.J.; Slaughter, L.H.; Krouse, J.M. Turgor potential and osmotic constituents of Kentucky bluegrass leaves supplied with four levels of potassium. Agron. J. 1994, 86, 1079–1083. [Google Scholar] [CrossRef]
- Munné-Bosch, S.; Alegre, L. Die and let live: Leaf senescence contributes to plant survival under drought stress. Funct. Plant Biol. 2004, 31, 203–216. [Google Scholar] [CrossRef]
- Xu, W.Z.; Deng, X.P.; Xu, B.C. Effects of water stress and fertilization on leaf gas exchange and photosynthetic light-response curves of Bothriochloa ischaemum L. Photosynthetica 2013, 51, 603–612. [Google Scholar] [CrossRef]
- Benjamin, J.G.; Nielsen, D.C.; Vigil, M.F.; Mikha, M.M.; Calderon, F. Water deficit stress effects on corn (Zea Mays, L.) root:shoot ratio. Open J. Soil Sci. 2014, 4, 151–160. [Google Scholar] [CrossRef]
- Comas, L.H.; Becker, S.R.; Cruz, V.M.; Byrne, P.F.; Dierig, D.A. Root traits contributing to plant productivity under drought. Front. Plant Sci. 2013, 4, 442. [Google Scholar] [CrossRef] [PubMed]
- Warren, J.M.; Hanson, P.J.; Iversen, C.M.; Kumar, J.; Walker, A.P.; Wullschleger, S.D. Root structural and functional dynamics in terrestrial biosphere models—Evaluation and recommendations. New Phytol. 2015, 205, 59–78. [Google Scholar] [CrossRef] [PubMed]
- Zörb, C.; Senbayram, M.; Peiter, E. Potassium in agriculture—Status and perspectives. J. Plant Physiol. 2014, 171, 656–669. [Google Scholar] [CrossRef] [PubMed]
- Egilla, J.N.; Davies, F.T., Jr.; Drew, M.C. Effect of potassium on drought resistance of Hibiscus rosa-sinensis Cv. Leprechaun: Plant growth, leaf macro- and micronutrient content and root longevity. Plant Soil 2001, 229, 213–224. [Google Scholar] [CrossRef]
- Martineau, E.; Domec, J.C.; Bosc, A.; Dannoura, M.; Gibon, Y.; Bernard, C.; Jordan-Meille, L. The role of potassium on maize leaf carbon exportation under drought condition. Acta Physiol. Plant. 2017, 39, 219. [Google Scholar] [CrossRef]
- Mengel, K.; Arneke, W.W. Effect of potassium on the water potential, the pressure potential, the osmotic potential and cell elongation in leaves of Phaseolus vulgaris. Physiol. Plant 1982, 54, 402–408. [Google Scholar] [CrossRef]
- Hsiao, T.C.; Acevedo, E.; Fereres, E.; Henderson, D.W. Water stress, growth, and osmotic adjustment. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1976, 273, 479–500. [Google Scholar] [CrossRef]
- Hsiao, T.C.; Lauchli, A. Role of potassium in plant-water relations. In Advances in Plant Nutrition; Tinker, B., Lauchli, A., Eds.; Praeger Publishers: New York, NY, USA, 1986; Volume 2, pp. 281–312. [Google Scholar]
- Benlloch-González, M.; Arquero, O.J.; Fournier, M.; Barranco, D.; Benlloch, M. K+ starvation inhibits water-stress-induced stomatal closure. J. Plant Physiol. 2008, 165, 623–630. [Google Scholar] [CrossRef] [PubMed]
- Jákli, B.; Trankner, M.; Senbayram, M.; Dittert, K. Adequate supply of potassium improves plant water-use efficiency but not leaf water-use efficiency of spring wheat. J. Plant Nutr. Soil Sci. 2016, 179, 733–745. [Google Scholar] [CrossRef]
- Jákli, B.; Tavakol, E.; Trankner, M.; Senbayram, M.; Dittert, K. Quantitative limitations to photosynthesis in K deficient sunflower and their implications on water-use efficiency. J. Plant Physiol. 2017, 209, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Cakmak, I. The role of potassium in alleviating detrimental effects of abiotic stresses in plants. J. Plant Nutr. Soil Sci. 2005, 168, 521–530. [Google Scholar] [CrossRef]
- Battie-Laclau, P.; Laclau, J.P.; Beri, C.; Mietton, L.; Almeida Muniz, M.R.; Arenque, B.C.; Piccolo, M.C.; Jordan-Meille, L.; Bouillet, J.P.; Nouvellon, Y. Photosynthetic and anatomical responses of Eucalyptus grandis leaves to potassium and sodium supply in a field experiment. Plant Cell Environ. 2014, 37, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Battie-Laclau, P.; Laclau, J.P.; Domec, J.C.; Christina, M.; Bouillet, J.P.; Piccolo, M.C.; Moraes Gonçalves, J.L.; Moreira, R.M.; Krusche, A.V.; Bouvet, J.M.; et al. Effects of potassium and sodium supply on drought-adaptive mechanisms in Eucalyptus grandis plantations. New Phytol. 2014, 203, 401–413. [Google Scholar] [CrossRef] [PubMed]
- Cakmak, I.; Hengeler, C.; Marschner, H. Changes in phloem export of sucrose in leaves in response to phosphorus, potassium and magnesium deficiency in bean plants. J. Exp. Bot. 1994, 45, 1251–1257. [Google Scholar] [CrossRef]
- Triboulot, M.B.; Pritchard, J.; Levy, G. Effects of potassium deficiency on cell water relations and elongation of tap and lateral roots of maritime pine seedlings. New Phytol. 1997, 135, 183–190. [Google Scholar] [CrossRef]
- Alemán, F.; Nieves-Cordones, M.; Martínez, V.; Rubio, F. Root K+ acquisition in plants: The Arabidopsis thaliana model. Plant Cell Physiol. 2011, 52, 1603–1612. [Google Scholar] [CrossRef] [PubMed]
- Thaler, P.; Pagès, L. Root apical diameter and root elongation rate of rubber seedlings (Hevea brasiliensis) show parallel responses to photoassimilate availability. Physiol. Plant. 1996, 97, 365–371. [Google Scholar] [CrossRef]
- Hogh-Jensen, H.; Pedersen, M.B. Morphological plasticity by crop plants and their potassium use efficiency. J. Plant Nutr. 2003, 26, 969–984. [Google Scholar] [CrossRef]
- Song, W.; Liu, S.; Meng, L.; Xue, R.; Wang, C.; Liu, G.; Dong, C.; Wang, S.; Dong, J.; Zhang, Y. Potassium deficiency inhibits lateral root development in tobacco seedlings by changing auxin distribution. Plant Soil 2015, 396, 163–173. [Google Scholar] [CrossRef]
- Tatsumi, J.; Endo, N.; Kono, Y. Root growth and partitioning of 13C-labelled photosynthate in the seminal root of corn seedlings as affected by light intensity. Jpn. J. Crop Sci. 1992, 61, 271–278. [Google Scholar] [CrossRef]
- Zhao, X.H.; Yu, H.Q.; Wen, J.; Wang, X.G.; Du, Q.; Wang, J.; Wang, Q. Response of root morphology, physiology and endogeneous hormones in maize (Zea mays L.) to potassium deficiency. J. Integr. Agric. 2016, 15, 785–794. [Google Scholar] [CrossRef]
- Tavakol, E.; Jakli, B.; Cakmak, I.; Dittert, K.; Karlovsky, P.; Pfohl, K.; Senbayram, M. Optimized potassium nutrition improves plant-water-relations of barley under PEG-induced osmotic stress. Plant Soil 2018, 430, 23–35. [Google Scholar] [CrossRef]
- NFX31-108 Soil Quality—Determination of Ammonium Acetate Extractable Ca++, Mg++, K+ and Na+ Cations—Agitation Method; AFNOR: Paris, France, 2002.
- Jordan-Meille, L.; Pellerin, S. Leaf area establishment of a maize (Zea mays L.) field crop under potassium deficiency. Plant Soil 2004, 265, 75–92. [Google Scholar] [CrossRef]
- Plénet, D.; Mollier, A.; Pellerin, S. Growth analysis of maize field crops under phosphorus deficiency II. Radiation-use efficiency, biomass accumulation and yield components. Plant Soil 2000, 224, 259–272. [Google Scholar] [CrossRef]
- McMaster, G.S. Phytomers, phyllochrons, phenology and temperate cereal development. J. Agric. Sci. 2005, 143, 137–150. [Google Scholar] [CrossRef]
- Luo, Y.; Gerten, D.; Le Maire, G.; Parton, W.J.; Weng, E.; Zhou, X.; Keough, C.; Beier, C.; Ciais, P.; Cramer, W.; et al. Modeled interactive effects of precipitation, temperature, and [CO2] on ecosystem carbon and water dynamics in different climatic zones. Glob. Chang. Biol. 2008, 14, 1986–1999. [Google Scholar] [CrossRef]
- Christina, M.; Le Maire, G.; Battie-Laclau, P.; Nouvellon, Y.; Bouillet, J.P.; Jourdan, C.; Gonçalves, J.L.M.; Laclau, J.P. Measured and modeled interactive effects of potassium deficiency and water deficit on gross primary productivity and light-use efficiency in Eucalyptus grandis plantations. Glob. Chang. Biol. 2015, 21, 2022–2039. [Google Scholar] [CrossRef] [PubMed]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Öborn, I.; Edwards, A.C.; Hillier, S. Quantifying uptake rate of potassium from soil in a long-term grass rotation experiment. Plant Soil 2010, 335, 3–19. [Google Scholar] [CrossRef]
- Ge, T.D.; Sun, N.B.; Bai, L.P.; Tong, C.L.; Sui, F.G. Effects of drought stress on phosphorus and potassium uptake dynamics in summer maize (Zea mays) throughout the growth cycle. Acta Physiol. Plant. 2012, 34, 2179–2186. [Google Scholar] [CrossRef]
- Gerardeaux, E.; Jordan-Meille, L.; Constantin, J.; Pellerin, S.; Dingkuhn, M. Changes in plant morphology and dry matter partitioning caused by potassium deficiency in Gossypium hirsutum (L.). Environ. Exp. Bot. 2010, 67, 451–459. [Google Scholar] [CrossRef]
- Fournier, J.M.; Roldán, A.M.; Sánchez, C.; Alexandre, G.; Benlloch, M. K+ starvation increases water uptake in whole sunflower plants. Plant Sci. 2005, 168, 823–829. [Google Scholar] [CrossRef]
- Ericsson, T. Growth and shoot: Root ratio of seedlings in relation to nutrient availability. Plant Soil 1995, 168, 205–214. [Google Scholar] [CrossRef]
- Aguirrezabal, L.A.N.; Tardieu, F. An architectural analysis of the elongation of field-grown sunflower root systems. Elements for modeling in the effects of temperature and intercepted radiation. J. Exp. Bot. 1996, 47, 411–420. [Google Scholar] [CrossRef]
- Hermans, C.; Hammond, J.P.; White, P.J.; Verbruggen, N. How do plants respond to nutrient shortage by biomass allocation? Trends Plant Sci. 2006, 11, 610–617. [Google Scholar] [CrossRef] [PubMed]
- Bednarz, C.W.; Oosterhuis, D.M.; Evans, R.D. Leaf photosynthesis and carbon isotope discrimination of cotton in response to potassium deficiency. Environ. Exp. Bot. 1998, 39, 131–139. [Google Scholar] [CrossRef]
- Zhao, D.; Oosterhuis, D.M.; Bednarz, C.W. Influence of potassium deficiency on photosynthesis, chlorophyll content, and chloroplast ultrastructure of cotton plants. Photosynthetica 2001, 39, 103–109. [Google Scholar] [CrossRef]
- Gerardeaux, E.; Saur, E.; Constantin, J.; Porté, A.; Jordan-Meille, L. Effect of carbon assimilation on dry weight production and partitioning during vegetative growth. Plant Soil 2009, 324, 329–343. [Google Scholar] [CrossRef]
- Arquero, O.; Barranco, D.; Benlloch, M. Potassium starvation increases stomatal conductance in olive trees. HortScience 2006, 41, 433–436. [Google Scholar]
- Bahrani, A.; Pourreza, J.; Madani, A.; Amiri, F. Effect of PRD irrigation method and potassium fertilizer application on corn yield and water use efficiency. Bulg. J. Agric. Sci. 2012, 18, 616–625. [Google Scholar]
- Singh, A.K.; Madramootoo, C.A.; Smith, D.L. Impact of different water management scenarios on corn water use efficiency. Trans. ASABE 2014, 57, 1319–1328. [Google Scholar] [CrossRef]
- Zhang, C.Z.; Zhang, J.B.; Zhang, H.; Zhao, J.H.; Wu, Q.C.; Zhao, Z.H.; Cai, T.Y. Mechanisms for the relationships between water-use efficiency and carbon isotope composition and specific leaf area of maize (Zea mays L.) under water stress. Plant Growth Regul. 2015, 77, 233–243. [Google Scholar] [CrossRef]
- Grzebisz, W.; Gransee, A.; Szczepaniak, W.; Diatta, J. The effects of potassium fertilisation on water-use efficiency in crop plants. J. Plant Nutr. Soil Sci. 2013, 176, 355–374. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. Potassium: A neglected nutrient in global change. Glob. Ecol. Biogeogr. 2015, 24, 261–275. [Google Scholar] [CrossRef]
- Oosterhuis, D.M.; Loka, D.A.; Raper, T.B. Potassium and stress alleviation: Physiological functions and management of cotton. J. Plant Nutr. Soil Sci. 2013, 176, 331–343. [Google Scholar] [CrossRef]
- Zahoor, R.; Dong, H.; Abid, M.; Zhao, W.; Wang, Y.; Zhou, Z. Potassium fertilizer improves drought stress alleviation potential in cotton by enhancing photosynthesis and carbohydrate metabolism. Environ. Exp. Bot. 2017, 137, 73–83. [Google Scholar] [CrossRef]
- Jákli, B.; Hauer-Jákli, M.; Böttcher, F.; Meyer zur Müdehorst, J.; Senbayram, M.; Dittert, K. Leaf, canopy and agronomic water-use efficiency of field-grown sugar beet in response to potassium fertilisation. J. Agron. Crop Sci. 2018, 204, 99–110. [Google Scholar] [CrossRef]
- Christina, M.; le Maire, G.; Nouvellon, Y.; Vezy, R.; Bordon, B.; Battie-Laclau, P.; Goncalves, J.L.M.; Delgado-Rojas, J.S.; Bouillet, J.P.; Laclau, J.P. Simulating the effects of different potassium and water supply regimes on soil water content and water table depth over a rotation of a tropical Eucalyptus grandis plantation. For. Ecol. Manag. 2018, 148, 4–14. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Statistical Tests | Parameters | p Values | ||
|---|---|---|---|---|
| W | K | W × K | ||
| Parametric | Number of Leaves | * | *** | * |
| Leaf Area (m2) | *** | *** | * | |
| Leaf Biomass (g) | *** | *** | *** | |
| Root Biomass (g) | ns | *** | ns | |
| LER (cm dd−1) | *** | *** | ns | |
| Relative Senescent Leaf (%) | ns | *** | ns | |
| Phyllochron (dd) | *** | ** | ** | |
| SLA (m2 kg−1) | ns | ns | ns | |
| R/(R + S) | ** | . | ns | |
| Transp. (L plant−1) | *** | *** | *** | |
| Transp. per unit time and leaf surf. (L cm−2 day−1) | ** | ns | ns | |
| Water Use Efficiency (g L−1) | *** | ** | ** | |
| Non-parametric | Root Area (m2) | ns | ** | |
| Specific Root Area (m2 kg−1) | ns | ns | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jordan-Meille, L.; Martineau, E.; Bornot, Y.; Lavres, J.; Abreu-Junior, C.H.; Domec, J.-C. How Does Water-Stressed Corn Respond to Potassium Nutrition? A Shoot-Root Scale Approach Study under Controlled Conditions. Agriculture 2018, 8, 180. https://doi.org/10.3390/agriculture8110180
Jordan-Meille L, Martineau E, Bornot Y, Lavres J, Abreu-Junior CH, Domec J-C. How Does Water-Stressed Corn Respond to Potassium Nutrition? A Shoot-Root Scale Approach Study under Controlled Conditions. Agriculture. 2018; 8(11):180. https://doi.org/10.3390/agriculture8110180
Chicago/Turabian StyleJordan-Meille, Lionel, Elsa Martineau, Yoran Bornot, José Lavres, Cassio Hamilton Abreu-Junior, and Jean-Christophe Domec. 2018. "How Does Water-Stressed Corn Respond to Potassium Nutrition? A Shoot-Root Scale Approach Study under Controlled Conditions" Agriculture 8, no. 11: 180. https://doi.org/10.3390/agriculture8110180
APA StyleJordan-Meille, L., Martineau, E., Bornot, Y., Lavres, J., Abreu-Junior, C. H., & Domec, J.-C. (2018). How Does Water-Stressed Corn Respond to Potassium Nutrition? A Shoot-Root Scale Approach Study under Controlled Conditions. Agriculture, 8(11), 180. https://doi.org/10.3390/agriculture8110180