

Spatio-Temporal Evolution of Net Ecosystem Productivity and Its Influencing Factors in Northwest China, 1982–2022


Abstract
1. Introduction
2. Study Area and Datasets
2.1. Study Area
2.2. Data Sources
3. Methodology
3.1. CASA Model
3.2. Drought Index
3.3. Quantification of Accumulative Effects of Drought
3.4. Trend Analysis
3.5. Geographical Detector
3.5.1. Parameter Optimization
3.5.2. Factor Detector
3.5.3. Interaction Detector
4. Results
4.1. Spatial and Temporal Distribution Pattern of NEP in Inner Mongolian Desert Grassland Ecosystems
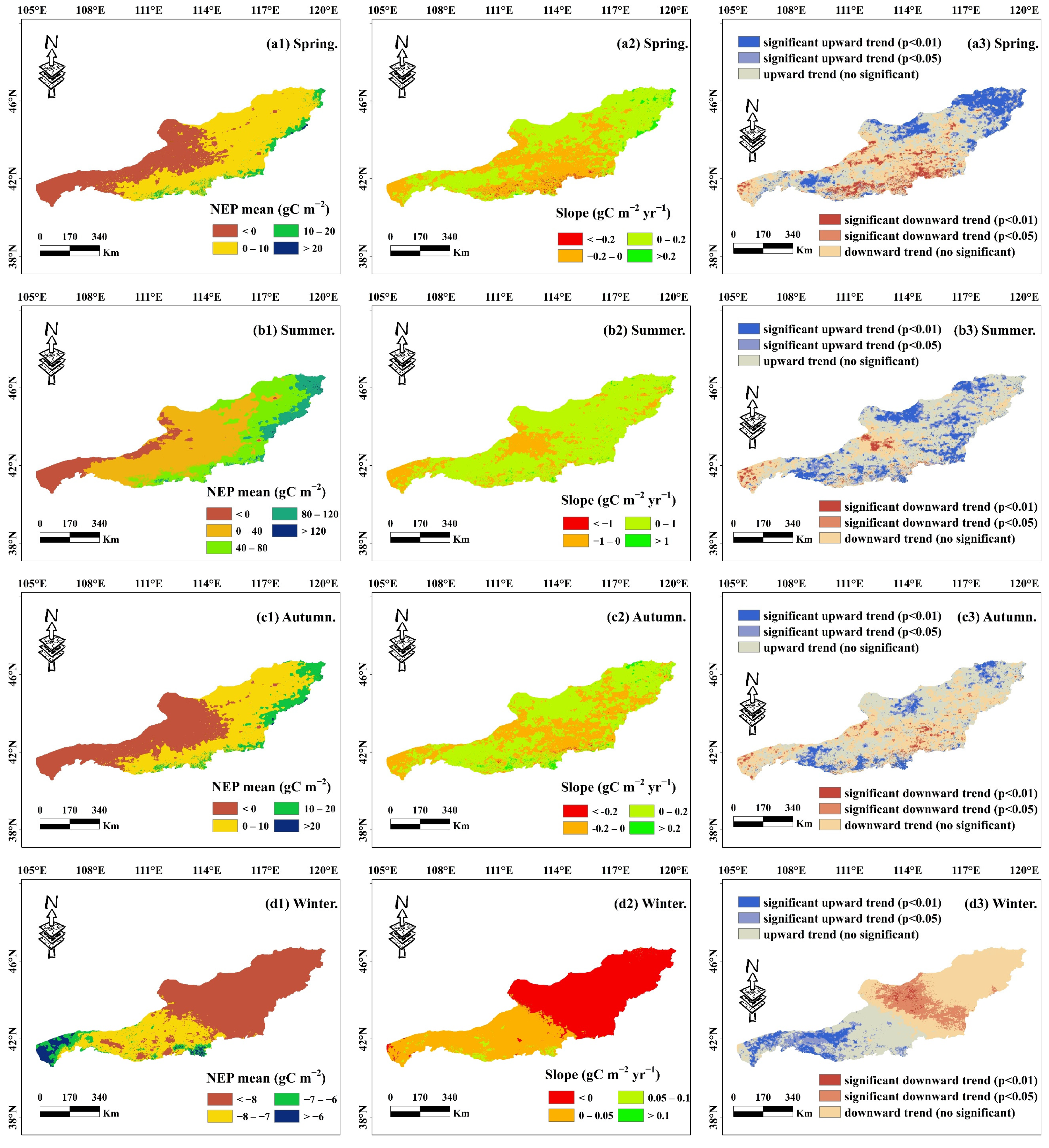
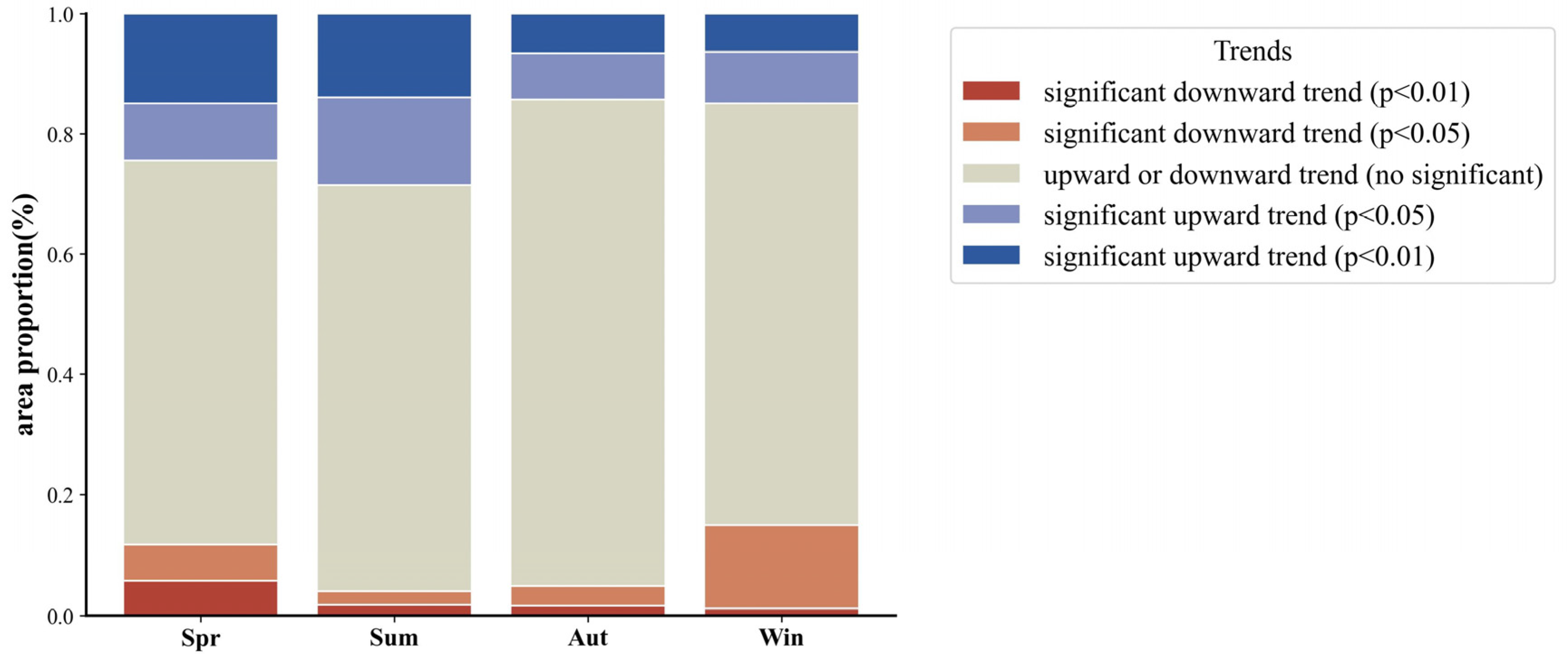
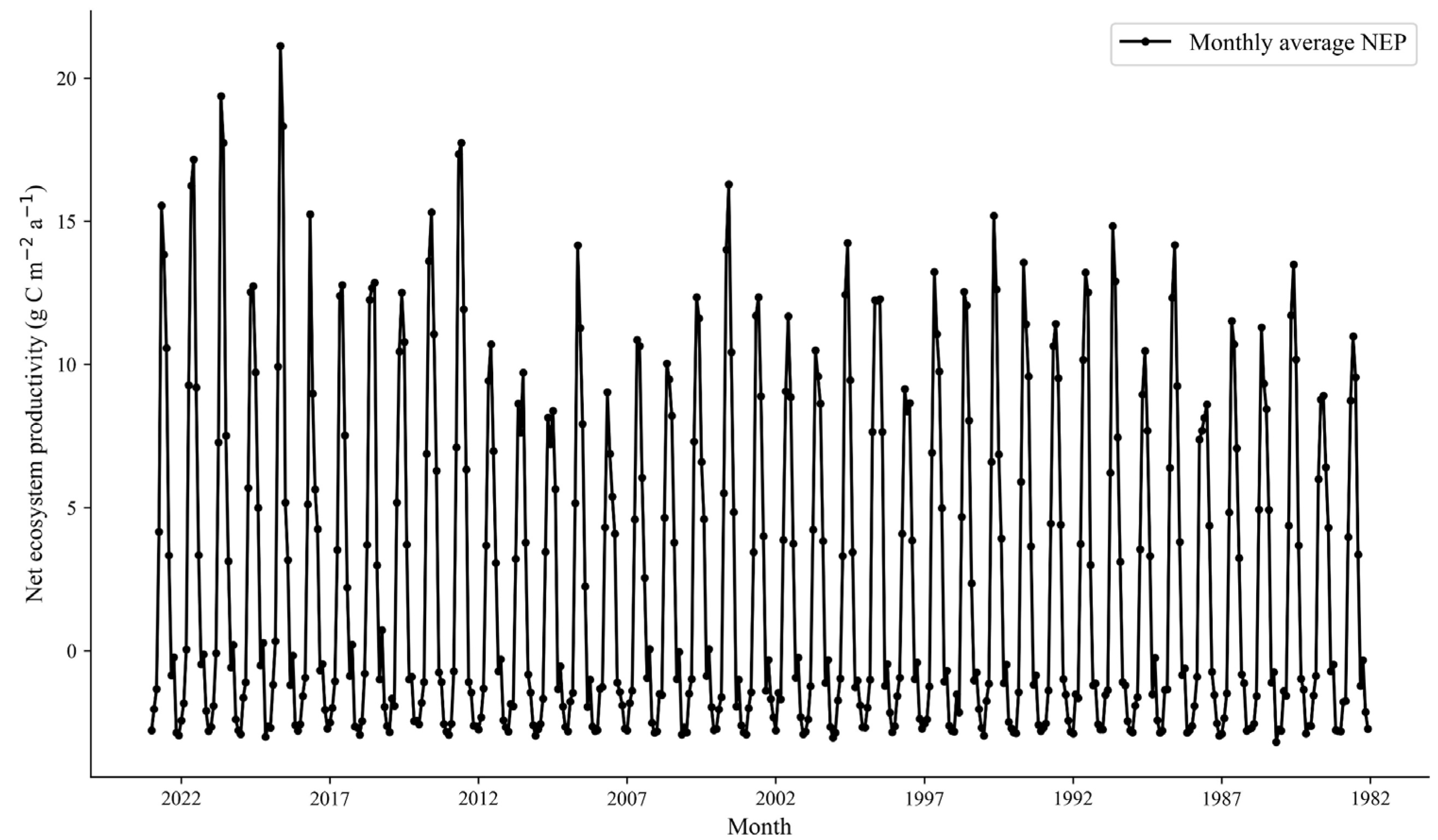
4.1.1. Spatial and Temporal Distribution of Annual and Seasonal NEP
4.1.2. Spatial and Temporal Distribution of Monthly NEP
4.2. Drought Assessment and Its Cumulative Effects on Vegetation Productivity
4.2.1. Drought Assessment
4.2.2. Cumulative Effects of Drought on Vegetation Productivity
4.3. Drivers of NEP Spatial Differentiation in Inner Mongolian Desert Grassland Ecosystems
4.3.1. Construction of Impact Factors
4.3.2. Factors Detection
4.3.3. Interaction Detector
5. Discussion
5.1. Cross-Validation of Data on Net Primary Productivity NPP
5.2. Analysis of NEP Trends and Driving Factors in Inner Mongolian Desert Grassland Ecosystems
5.3. Effects of Drought on Carbon Sinks in Desert Grassland Ecosystems of Inner Mongolia
5.4. Uncertainties and Limitations
6. Conclusions
- (1)
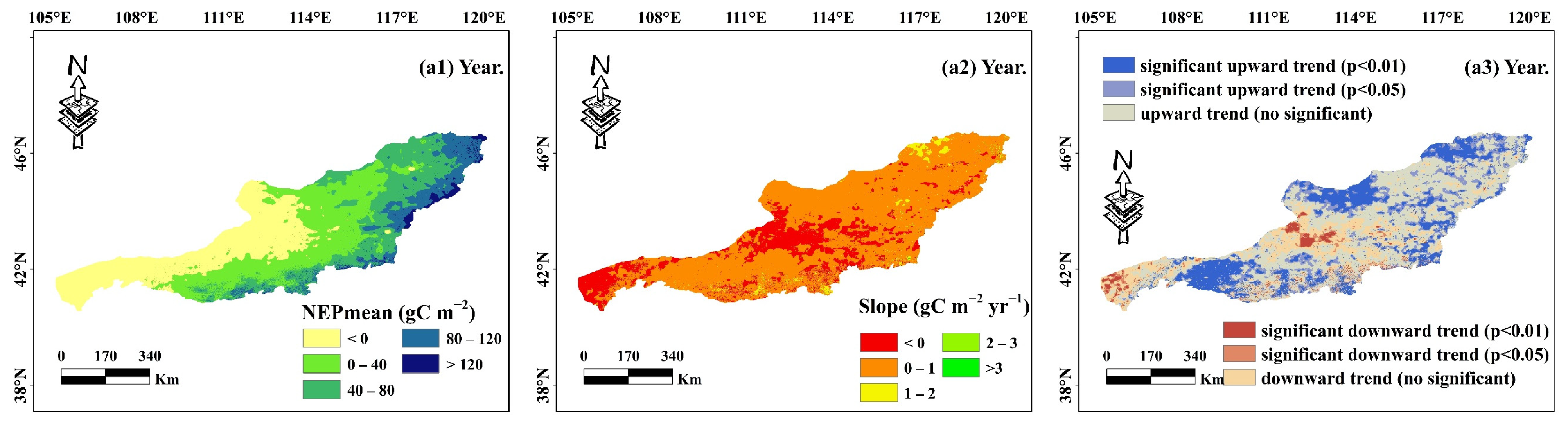
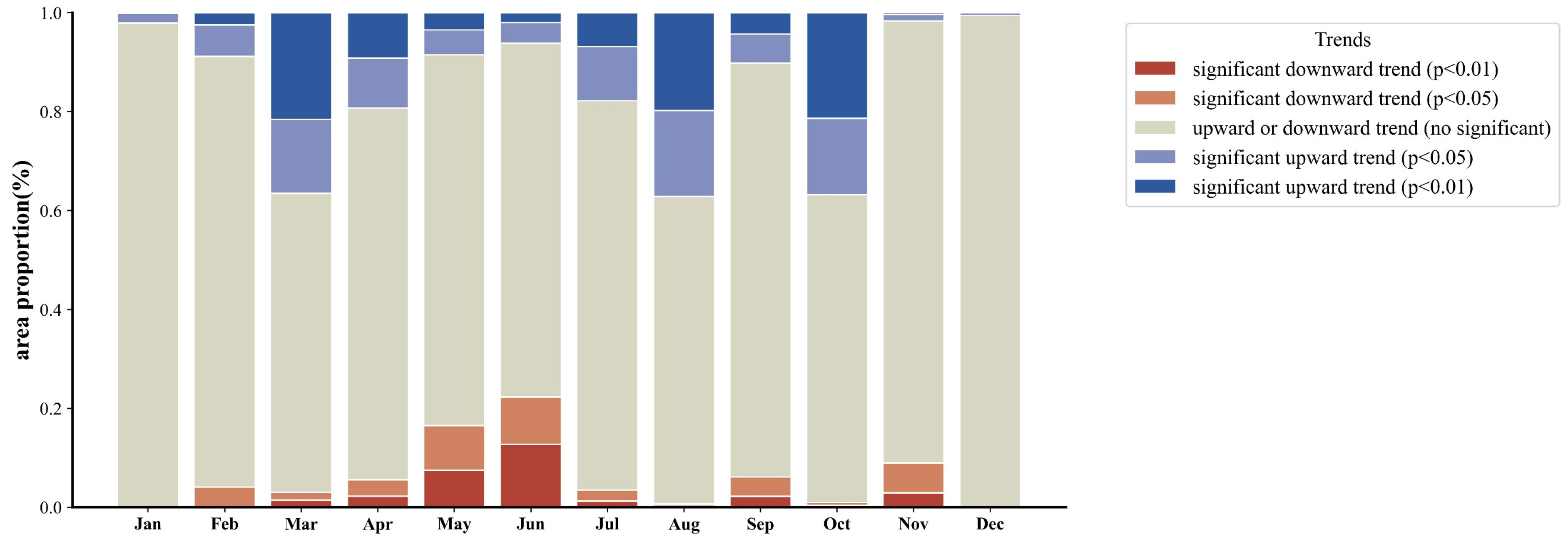
- The NEP distribution in the desert grassland ecosystem exhibited a gradual increase from southwest to northeast, with a multi-year average of 29.41 gC m−2. The carbon sink area (NEP > 0) covered 67.99% of the total area, while the carbon source area (NEP < 0) accounted for 32.01%. From 1982, the overall NEP showed a significant upward trend, with 35.40% of the area experiencing increased NEP, 59.59% showing no significant change, and only 5.01% exhibiting a significant decrease.
- (2)
- The Standardized Precipitation Evapotranspiration Index (SPEI) was employed to assess drought patterns in the region. The spatial distribution of moderate and severe droughts was similar in frequency, whereas exceptional droughts showed distinct regional variations. The duration of mild droughts was more evenly distributed compared to moderate and severe droughts. Overall, mild droughts had the most widespread impact.
- (3)
- Over the study period, drought had a cumulative effect on the desert grassland ecosystem, primarily in the short term (1–2 months), affecting 54.5% of the area. This indicates a swift ecosystem response, particularly in the central and eastern parts of the inland river region.
- (4)
- Geoprobes identified the key drivers of NEP, with mean annual precipitation exerting the strongest influence, followed by mean annual solar radiation, temperature, potential evapotranspiration, soil moisture, land use type, and elevation. Notably, combined effects of multiple factors generally surpassed the influence of individual factors, with two-factor interactions explaining a higher proportion of NEP variability.
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ballantyne, A.P.; Alden, C.B.; Miller, J.B.; Tans, P.P.; White, J.W.C. Increase in observed net carbon dioxide uptake by land and oceans during the past 50 years. Nature 2012, 488, 70–72. [Google Scholar] [CrossRef]
- Regnier, P.; Resplandy, L.; Najjar, R.G.; Ciais, P. The land-to-ocean loops of the global carbon cycle. Nature 2022, 603, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; He, Y.; Zhang, L.; Sun, Q.; Zhang, Y.; Li, H.; Wei, X.; Liu, Y. Spatiotemporal dynamics of vegetation net ecosystem productivity and its response to drought in Northwest China. GIScience Remote Sens. 2023, 60, 2194597. [Google Scholar] [CrossRef]
- Zhang, M.; Yuan, N.; Lin, H.; Liu, Y.; Zhang, H. Quantitative estimation of the factors impacting spatiotemporal variation in NPP in the Dongting Lake wetlands using Landsat time series data for the last two decades. Ecol. Indic. 2022, 135, 108544. [Google Scholar] [CrossRef]
- Wang, C.; Zhao, W.; Zhang, Y. The Change in Net Ecosystem Productivity and its Driving Mechanism in a Mountain Ecosystem of Arid Regions, Northwest China. Remote Sens. 2022, 14, 4046. [Google Scholar] [CrossRef]
- Fernández-Martínez, M.; Vicca, S.; Janssens, I.A.; Sardans, J.; Luyssaert, S.; Campioli, M.; Chapin Iii, F.S.; Ciais, P.; Malhi, Y.; Obersteiner, M.; et al. Nutrient availability as the key regulator of global forest carbon balance. Nat. Clim. Change 2014, 4, 471–476. [Google Scholar] [CrossRef]
- Dong, S.; Ren, W.; Dong, X.; Lei, F.; Wang, X.-C.; Xie, L.; Zhou, X. Decoupling the Impacts of Climate Change and Human Activities on Terrestrial Vegetation Carbon Sink. Remote Sens. 2024, 16, 4417. [Google Scholar] [CrossRef]
- Piao, S.; Zhang, X.; Chen, A.; Liu, Q.; Lian, X.; Wang, X.; Peng, S.; Wu, X. The impacts of climate extremes on the terrestrial carbon cycle: A review. Sci. China Earth Sci. 2019, 62, 1551–1563. [Google Scholar] [CrossRef]
- Wang, R.; Gamon, J.A.; Emmerton, C.A.; Springer, K.R.; Yu, R.; Hmimina, G. Detecting intra- and inter-annual variability in gross primary productivity of a North American grassland using MODIS MAIAC data. Agric. For. Meteorol. 2020, 281, 107859. [Google Scholar] [CrossRef]
- Faiz, M.A.; Zhang, Y.; Zhang, X.; Ma, N.; Aryal, S.K.; Ha, T.T.V.; Baig, F.; Naz, F. A composite drought index developed for detecting large-scale drought characteristics. J. Hydrol. 2022, 605, 127308. [Google Scholar] [CrossRef]
- Ciais, P.; Reichstein, M.; Viovy, N.; Granier, A.; Ogée, J.; Allard, V.; Aubinet, M.; Buchmann, N.; Bernhofer, C.; Carrara, A. Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature 2005, 437, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Zscheischler, J.; Mahecha, M.D.; Von Buttlar, J.; Harmeling, S.; Jung, M.; Rammig, A.; Randerson, J.T.; Schölkopf, B.; Seneviratne, S.I.; Tomelleri, E. A few extreme events dominate global interannual variability in gross primary production. Environ. Res. Lett. 2014, 9, 035001. [Google Scholar] [CrossRef]
- Zhao, M.; Running, S.W. Drought-Induced Reduction in Global Terrestrial Net Primary Production from 2000 Through 2009. Science 2010, 329, 940–943. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Zhen, Z.; Zhao, W.; Wang, X. Forest carbon sink in North China increased in recent two decades, but decreased in extreme drought years. GIScience Remote Sens. 2024, 61, 2383040. [Google Scholar] [CrossRef]
- Deng, Y.; Wang, X.; Wang, K.; Ciais, P.; Tang, S.; Jin, L.; Li, L.; Piao, S. Responses of vegetation greenness and carbon cycle to extreme droughts in China. Agric. For. Meteorol. 2021, 298, 108307. [Google Scholar] [CrossRef]
- Lindroth, A.; Holst, J.; Linderson, M.-L.; Aurela, M.; Biermann, T.; Heliasz, M.; Chi, J.; Ibrom, A.; Kolari, P.; Klemedtsson, L. Effects of drought and meteorological forcing on carbon and water fluxes in Nordic forests during the dry summer of 2018. Philos. Trans. R. Soc. B 2020, 375, 20190516. [Google Scholar] [CrossRef]
- McKee, T.B.; Doesken, N.J.; Kleist, J. The relationship of drought frequency and duration to time scales. In Proceedings of the 8th Conference on Applied Climatology, Anaheim, CA, USA, 17–22 January 1993; pp. 179–183. [Google Scholar]
- Palmer, W.C. Meteorological Drought; US Department of Commerce, Weather Bureau: Silver Spring, MD, USA, 1965; Volume 30. [Google Scholar]
- Shukla, S.; Wood, A.W. Use of a standardized runoff index for characterizing hydrologic drought. Geophys. Res. Lett. 2008, 35, L02405. [Google Scholar] [CrossRef]
- Hao, Z.; AghaKouchak, A. Multivariate Standardized Drought Index: A parametric multi-index model. Adv. Water Resour. 2013, 57, 12–18. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I. A Multiscalar Drought Index Sensitive to Global Warming: The Standardized Precipitation Evapotranspiration Index. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef]
- Ali, Z.; Hussain, I.; Faisal, M.; Almanjahie, I.M.; Ahmad, I.; Khan, D.M.; Grzegorczyk, M.; Qamar, S. A Probabilistic Weighted Joint Aggregative Drought Index (PWJADI) criterion for drought monitoring systems. Tellus A Dyn. Meteorol. Oceanogr. 2019, 71, 1588584. [Google Scholar] [CrossRef]
- Afshar, M.H.; Bulut, B.; Duzenli, E.; Amjad, M.; Yilmaz, M.T. Global spatiotemporal consistency between meteorological and soil moisture drought indices. Agric. For. Meteorol. 2022, 316, 108848. [Google Scholar] [CrossRef]
- Zhang, B.; He, C.; Burnham, M.; Zhang, L. Evaluating the coupling effects of climate aridity and vegetation restoration on soil erosion over the Loess Plateau in China. Sci. Total Environ. 2016, 539, 436–449. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Li, R.; Wu, Y.; Zhao, S. Effects of multi-temporal scale drought on vegetation dynamics in Inner Mongolia from 1982 to 2015, China. Ecol. Indic. 2022, 136, 108666. [Google Scholar] [CrossRef]
- Cai, Y.; Akiyama, H. Nitrogen loss factors of nitrogen trace gas emissions and leaching from excreta patches in grassland ecosystems: A summary of available data. Sci. Total Environ. 2016, 572, 185–195. [Google Scholar] [CrossRef]
- Gang, C.; Li, J.; Ju, W.; Chen, J. The comparisons of carbon sink/source of Chinese grassland ecosystem using different approaches. In Proceedings of the 2011 International Conference on Remote Sensing, Environment and Transportation Engineering, Nanjing, China, 24–26 June 2011; pp. 841–844. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, X.; Wang, Z.; Zhao, L.; Zhang, Q.-p.J.G.E. Conservation. Time scale selection and periodicity analysis of grassland drought monitoring index in Inner Mongolia. Glob. Ecol. Conserv. 2022, 36, e02138. [Google Scholar] [CrossRef]
- Wang, Q.; Yang, Y.; Liu, Y.; Tong, L.; Zhang, Q.-P.; Li, J. Assessing the Impacts of Drought on Grassland Net Primary Production at the Global Scale. Sci. Rep. 2019, 9, 14041. [Google Scholar] [CrossRef]
- Sun, J.; Du, W. Effects of precipitation and temperature on net primary productivity and precipitation use efficiency across China’s grasslands. GIScience Remote Sens. 2017, 54, 881–897. [Google Scholar] [CrossRef]
- Sun, C.-m.; Zhong, X.-c.; Chen, C.; Gu, T.; Chen, W. Evaluating the grassland net primary productivity of southern China from 2000 to 2011 using a new climate productivity model. J. Integr. Agric. 2016, 15, 1638–1644. [Google Scholar] [CrossRef]
- Chen, S.; Huang, Y.; Zou, J.; Shi, Y.; Lu, Y.; Zhang, W.; Hu, Z. Interannual variability in soil respiration from terrestrial ecosystems in China and its response to climate change. Sci. China Earth Sci. 2012, 55, 2091–2098. [Google Scholar] [CrossRef]
- Potter, C.S.; Randerson, J.T.; Field, C.B.; Matson, P.A.; Vitousek, P.M.; Mooney, H.A.; Klooster, S.A. Terrestrial ecosystem production: A process model based on global satellite and surface data. Glob. Biogeochem. Cycles 1993, 7, 811–841. [Google Scholar] [CrossRef]
- Zhu, W.-Q.; Pan, Y.-Z.; Zhang, J.-S. Estimation of net primary productivity of Chinese terrestrial vegetation based on remote sensing. Chin. J. Plant Ecol. 2007, 31, 413. [Google Scholar] [CrossRef]
- Pei, Z.-Y.; Ouyang, H.; Zhou, C.-P.; Xu, X.-L. Carbon Balance in an Alpine Steppe in the Qinghai-Tibet Plateau. J. Integr. Plant Biol. 2009, 51, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Yang, S.; Li, S.; Zhang, H.; Zhang, J.; Fan, H. The optimal applications of scPDSI and SPEI in characterizing meteorological drought, agricultural drought and terrestrial water availability on a global scale. Sci. Total Environ. 2024, 952, 175933. [Google Scholar] [CrossRef]
- Potopová, V.; Štěpánek, P.; Možný, M.; Türkott, L.; Soukup, J. Performance of the standardised precipitation evapotranspiration index at various lags for agricultural drought risk assessment in the Czech Republic. Agric. For. Meteorol. 2015, 202, 26–38. [Google Scholar] [CrossRef]
- Liu, X.; Yu, S.; Yang, Z.; Dong, J.; Peng, J. The first global multi-timescale daily SPEI dataset from 1982 to 2021. Sci. Data 2024, 11, 123. [Google Scholar] [CrossRef]
- Peng, J.; Wu, C.; Zhang, X.; Wang, X.; Gonsamo, A. Satellite detection of cumulative and lagged effects of drought on autumn leaf senescence over the Northern Hemisphere. Glob. Chang. Biol. 2019, 25, 2174–2188. [Google Scholar] [CrossRef]
- Gupta, N.; Mahato, P.K.; Patel, J.; Omar, P.J.; Tripathi, R.P. Chapter 30—Understanding trend and its variability of rainfall and temperature over Patna (Bihar). Curr. Dir. Water Scarcity Res. 2022, 7, 533–543. [Google Scholar] [CrossRef]
- Sen, P.K. Estimates of the regression coefficient based on Kendall’s tau. J. Am. Stat. Assoc. 1968, 63, 1379–1389. [Google Scholar] [CrossRef]
- Yin, Y.; Xu, C.-Y.; Chen, H.; Li, L.; Xu, H.; Li, H.; Jain, S.K. Trend and concentration characteristics of precipitation and related climatic teleconnections from 1982 to 2010 in the Beas River basin, India. Glob. Planet. Chang. 2016, 145, 116–129. [Google Scholar] [CrossRef]
- Song, Y.; Wang, J.; Ge, Y.; Xu, C. An optimal parameters-based geographical detector model enhances geographic characteristics of explanatory variables for spatial heterogeneity analysis: Cases with different types of spatial data. GIScience Remote Sens. 2020, 57, 593–610. [Google Scholar] [CrossRef]
- Luo, L.; Li, Y.; Ma, W.; Rong, J.; Wei, J.; Cui, Y.; Qu, T. Temporal–Spatial Dynamics and Collaborative Effects of Cropland Resilience in China. Land 2025, 14, 108. [Google Scholar] [CrossRef]
- Liu, C.; Li, W.; Wang, W.; Zhou, H.; Liang, T.; Hou, F.; Xu, J.; Xue, P. Quantitative spatial analysis of vegetation dynamics and potential driving factors in a typical alpine region on the northeastern Tibetan Plateau using the Google Earth Engine. CATENA 2021, 206, 105500. [Google Scholar] [CrossRef]
- Dai, E.F.; Huang, Y.; Wu, Z.; Zhao, D.S. Analysis of spatio-temporal features of a carbon source/sink and its relationship to climatic factors in the Inner Mongolia grassland ecosystem. J. Geogr. Sci. 2016, 26, 297–312. [Google Scholar] [CrossRef]
- Zhang, R.; Zhao, X.Y.; Zuo, X.A.; Qu, H.; Degen, A.A.; Luo, Y.Y.; Ma, X.J.; Chen, M.; Liu, L.X.; Chen, J.L. Impacts of Precipitation on Ecosystem Carbon Fluxes in Desert-Grasslands in Inner Mongolia, China. J. Geophys. Res.-Atmos. 2019, 124, 1266–1276. [Google Scholar] [CrossRef]
- Zarei, A.R.; Shabani, A.; Moghimi, M.M. Accuracy Assessment of the SPEI, RDI and SPI Drought Indices in Regions of Iran with Different Climate Conditions. Pure Appl. Geophys. 2021, 178, 1387–1403. [Google Scholar] [CrossRef]
- Schwalm, C.R.; Anderegg, W.R.L.; Michalak, A.M.; Fisher, J.B.; Biondi, F.; Koch, G.; Litvak, M.; Ogle, K.; Shaw, J.D.; Wolf, A.; et al. Global patterns of drought recovery. Nature 2017, 548, 202–205. [Google Scholar] [CrossRef]
- Jiao, T.; Williams, C.A.; De Kauwe, M.G.; Schwalm, C.R.; Medlyn, B.E. Patterns of post-drought recovery are strongly influenced by drought duration, frequency, post-drought wetness, and bioclimatic setting. Glob. Chang. Biol. 2021, 27, 4630–4643. [Google Scholar] [CrossRef]
- Zheng, L.; Lu, J.; Chen, X. Drought offsets the vegetation greenness-induced gross primary productivity from 1982 to 2018 in China. J. Hydrol. 2024, 632, 130881. [Google Scholar] [CrossRef]
- Sun, J.; Zhang, Q.; Xu, Y.J.; Liu, X.; Chen, L.; Zhang, G.; Wu, Y. Will drought evolution accelerate under future climate? J. Hydrol. 2025, 650, 132552. [Google Scholar] [CrossRef]
- Smith, T.; Traxl, D.; Boers, N. Empirical evidence for recent global shifts in vegetation resilience. Nat. Clim. Chang. 2022, 12, 477–484. [Google Scholar] [CrossRef]
- Lu, M.; Sun, H.; Yang, Y.; Xue, J.; Ling, H.; Zhang, H.; Zhang, W. Assessing recovery time of ecosystems in China: Insights into flash drought impacts on gross primary productivity. Hydrol. Earth Syst. Sci. 2025, 29, 613–625. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, X.; Jiao, W.; Wu, X.; Zeng, X.; Zhao, L.; Wang, L.; Guo, J.; Xing, X.; Hong, Y. Spatial Heterogeneity of Vegetation Resilience Changes to Different Drought Types. Earth’s Future 2023, 11, e2022EF003108. [Google Scholar] [CrossRef]
- Wu, T.; Xu, L.; Chen, N. Spatial pattern and attribution of ecosystem drought recovery in China. J. Hydrol. 2024, 638, 131578. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data | Acquired Time | Resolution | Data Source |
|---|---|---|---|
| CLCD | 1985–2023 | 0.00027° × 0.00027° | Annual China Land Cover Dataset (https://zenodo.org) (accessed on 7 August 2024) |
| DEM | / | 0.009° × 0.009° | National Cryosphere Desert Data Center (https://data.tpdc.ac.cn) (accessed on 6 June 2024) |
| NDVI | 1981–2022 | 0.0833° × 0.0833° | EARTHDATA (https://daac.ornl.gov) (accessed on 12 August 2024) |
| Precipitation and Temperature | 1901–2022 | 0.009° × 0.009° | National Cryosphere Desert Data Center (https://data.tpdc.ac.cn) (accessed on 10 August 2024) |
| Solar Radiation | 1950–2023 | 0.1° × 0.1° | (https://cds.climate.copernicus.eu) (accessed on 22 August 2024) |
| NPP | 2000–2022 | 0.0045° × 0.0045° | (https://lpdaac.usgs.gov/products/mod17a3hgfv061/) (accessed on 8 April 2024) |
| Drought Grade | Drought Type | SPEI Value |
|---|---|---|
| 1 | Normal | SPEI > −0.5 |
| 2 | Mild drought | −1.0 < SPEI ≤ −0.5 |
| 3 | Moderate drought | −1.5 < SPEI ≤ −1.0 |
| 4 | Severe drought | −2.0 < SPEI ≤ −1.5 |
| 5 | Extreme drought | SPEI ≤ −2.0 |
| Interaction | Base of Assessment |
|---|---|
| Weaken, nonlinear | q(X1∩X2) < Min[q(X1),q(X2)] |
| Weaken, univariate | Min[q(X1),q(X1)] < q(X1∩X2) < Max[q(X1),q(X2)] |
| Enhance, bivariate | q(X1∩X2) > Max[q(X1),q(X2)] |
| Independent | q(X1∩X2) = q(X1) + q(X2) |
| Enhance, nonlinear | q(X1∩X2) > q(X1) + q(X2) |
| Month | MIN (gCm−2) | MEAN (gCm−2) | MAX (gCm−2) | Percentage of Carbon Sink Areas (NEP > 0) |
|---|---|---|---|---|
| January | −3.10 | −2.75 | 1.85 | 0 |
| February | −3.22 | −2.45 | 0.13 | 0 |
| March | −3.74 | −0.59 | 4.00 | 16.30% |
| April | −4.69 | −1.02 | 3.86 | 8.45% |
| May | −5.10 | 3.93 | 25.89 | 89.18% |
| June | −6.70 | 8.76 | 49.70 | 86.84% |
| July | −7.68 | 11.97 | 53.31 | 83.05% |
| August | −7.08 | 12.17 | 47.87 | 89.86% |
| September | −5.63 | 5.27 | 26.51 | 89.01% |
| October | −4.67 | −1.23 | 4.16 | 9.19% |
| November | −3.47 | −1.89 | 0.56 | 0 |
| December | −3.31 | −2.76 | −1.62 | 0 |
| Driver Factors | Codes | Units |
|---|---|---|
| Land use type | X1 | / |
| Elevation | X2 | m |
| Potential evapotranspiration | X3 | mm |
| Average annual precipitation | X4 | mm |
| Soil moisture | X5 | m3·m−3 |
| Average annual solar radiation | X6 | W/m2 |
| Average annual temperature | X7 | ℃ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Xu, Z.; Yuan, H.; Wang, Y.; Feng, K.; Li, Y.; Wang, F.; Zhang, Z. Spatio-Temporal Evolution of Net Ecosystem Productivity and Its Influencing Factors in Northwest China, 1982–2022. Agriculture 2025, 15, 613. https://doi.org/10.3390/agriculture15060613
Zhang W, Xu Z, Yuan H, Wang Y, Feng K, Li Y, Wang F, Zhang Z. Spatio-Temporal Evolution of Net Ecosystem Productivity and Its Influencing Factors in Northwest China, 1982–2022. Agriculture. 2025; 15(6):613. https://doi.org/10.3390/agriculture15060613
Chicago/Turabian StyleZhang, Weijie, Zhichao Xu, Haobo Yuan, Yingying Wang, Kai Feng, Yanbin Li, Fei Wang, and Zezhong Zhang. 2025. "Spatio-Temporal Evolution of Net Ecosystem Productivity and Its Influencing Factors in Northwest China, 1982–2022" Agriculture 15, no. 6: 613. https://doi.org/10.3390/agriculture15060613
APA StyleZhang, W., Xu, Z., Yuan, H., Wang, Y., Feng, K., Li, Y., Wang, F., & Zhang, Z. (2025). Spatio-Temporal Evolution of Net Ecosystem Productivity and Its Influencing Factors in Northwest China, 1982–2022. Agriculture, 15(6), 613. https://doi.org/10.3390/agriculture15060613