
GIS Bioclimatic Profile and Seed Germination of the Endangered and Protected Cretan Endemic Plant Campanula cretica (A. DC.) D. Dietr. for Conservation and Sustainable Utilization

 , , , , and
, , , , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Value
2.2. GIS Ecological Profiling
2.3. Seed Collection and Storage
2.4. Seed Germination Experiment
2.5. Statistical Analysis
3. Results
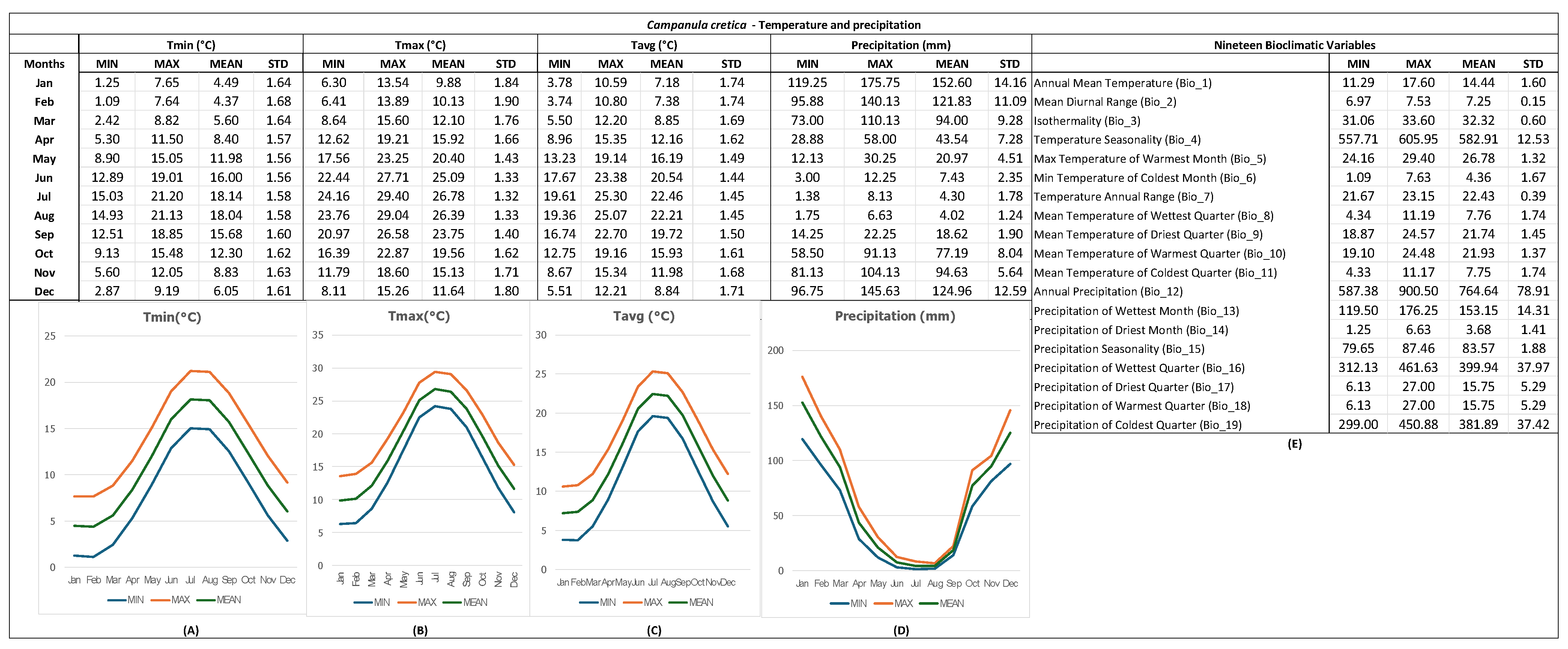
3.1. GIS-Derived Bioclimatic (Ecological) Profile
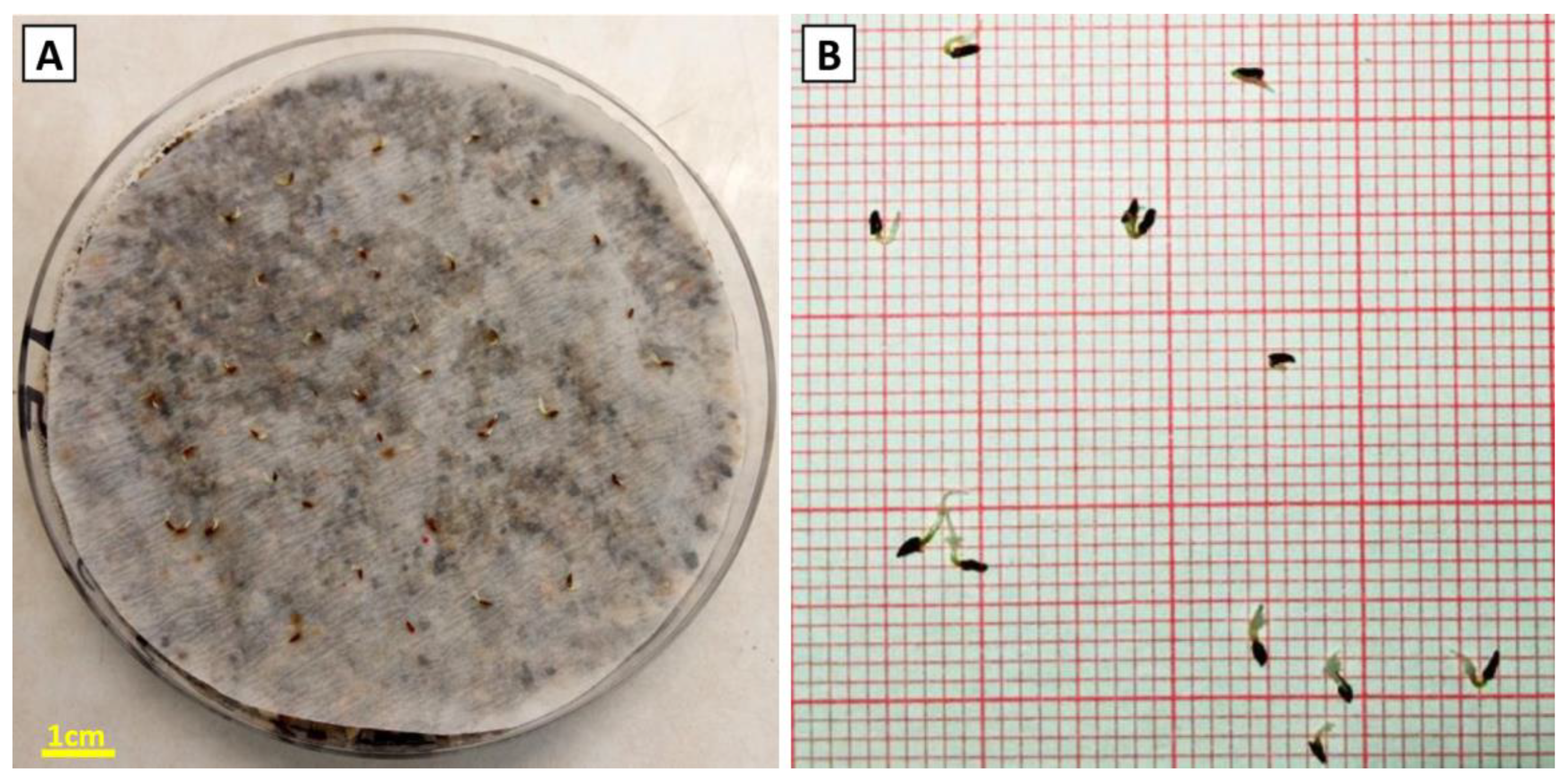
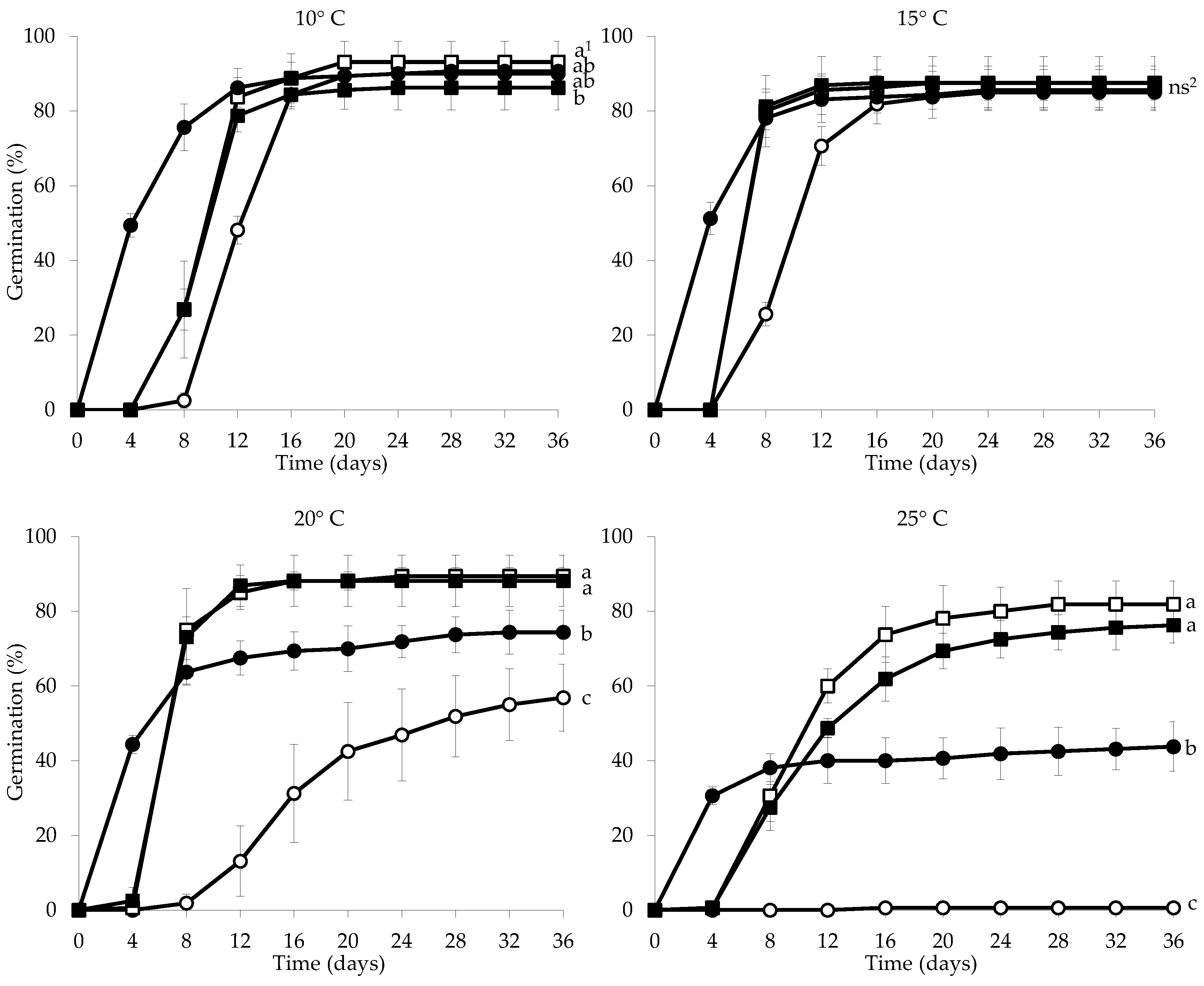
3.2. Seed Germination Experiment and Seed Germination Success
3.2.1. Effects of Incubation Temperature and Different Pre-Treatments
3.2.2. Effects of Light Treatment
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. 2024. Available online: https://powo.science.kew.org/ (accessed on 13 April 2025).
- Liveri, E.; Kalachanis, D.; Bareka, P.; Grammatikopoulos, G.; Kamari, G. Contribution to the seed morphology of some Greek Campanula species of Sect. Quinqueloculares (Campanulaceae). Flora Medit. 2020, 30, 347–363. [Google Scholar] [CrossRef]
- Flora of Greece Web. Available online: https://portal.cybertaxonomy.org/flora-greece/intro (accessed on 13 April 2025).
- Dimitriadis, K.M.; Karavergou, S.; Hadjipavlou-Litina, D.; Krigas, N.; Lazari, D. Phytochemical and antioxidant evaluation of the ex-situ cultivated species Petromarula pinnata (L.) A. DC. and Campanula cretica (A.DC.) Dietr. (Campanulaceae) from Crete (Greece). Planta Med. 2022, 88, 1523. [Google Scholar] [CrossRef]
- Tsiftsoglou, O.S.; Lagogiannis, G.; Psaroudaki, A.; Vantsioti, A.; Mitić, M.N.; Mrmošanin, J.M.; Lazari, D. Phytochemical analysis of the aerial parts of Campanula pelviformis Lam. (Campanulaceae): Documenting the dietary value of a local endemic plant of Crete (Greece) traditionally used as wild edible green. Sustainability 2023, 15, 7404. [Google Scholar] [CrossRef]
- Grigoriadou, K.; Krigas, N.; Maloupa, E. GIS-Facilitated ex situ conservation of the rare Greek endemic Campanula incurva Aucher: Seed germination requirements and effect of growth regulators on in vitro proliferation and rooting. Plant Biosyst. 2014, 148, 1169–1177. [Google Scholar] [CrossRef]
- Sarropoulou, V.; Krigas, N.; Tsoktouridis, G.; Maloupa, E.; Grigoriadou, K. Seed germination trials and ex situ conservation of local prioritized endemic plants of Crete (Greece) with commercial interest. Seeds 2022, 1, 279–302. [Google Scholar] [CrossRef]
- Anestis, I.; Pipinis, E.; Kostas, S.; Papaioannou, E.; Karapatzak, E.; Dariotis, E.; Tsoulpha, P.; Koundourakis, E.; Chatzileontari, E.; Tsoktouridis, G.; et al. GIS-Facilitated germination of stored seeds from five wild-growing populations of Campanula pelviformis Lam. and fertilization effects on growth, nutrients, phenol content and antioxidant potential. Horticulturae 2023, 9, 877. [Google Scholar] [CrossRef]
- Panagiotidou, T.-N.; Pipinis, E.; Anestis, I.; Kostas, S.; Tsoulpha, P.; Karapatzak, E.; Tsoktouridis, G.; Hatzilazarou, S.; Krigas, N. Integrated ex-situ conservation and ornamental evaluation of the Vulnerable and protected Greek endemic Campanula laciniata L.: A multifaceted approach. Agronomy 2024, 14, 1665. [Google Scholar] [CrossRef]
- Krigas, N.; Tsoktouridis, G.; Anestis, I.; Khabbach, A.; Libiad, M.; Megdiche-Ksouri, W.; Bourgou, S. Exploring the potential of neglected local endemic plants of three Mediterranean regions in the ornamental sector: Value chain feasibility and readiness timescale for their sustainable exploitation. Sustainability 2021, 13, 2539. [Google Scholar] [CrossRef]
- Menteli, V.; Krigas, N.; Avramakis, M.; Turland, N.; Vokou, D. Endemic plants of Crete in electronic trade and wildlife tourism: Current patterns and implications for conservation. J. Biol. Res. 2019, 26, 10. [Google Scholar] [CrossRef]
- Médail, F. Plant biogeography and vegetation patterns of the Mediterranean islands. Bot. Rev. 2022, 88, 63–129. [Google Scholar] [CrossRef]
- Kougioumoutzis, K.; Kokkoris, I.P.; Panitsa, M.; Kallimanis, A.; Strid, A.; Dimopoulos, P. Plant endemism centres and biodiversity hotspots in Greece. Biology 2021, 10, 72. [Google Scholar] [CrossRef]
- Kougioumoutzis, K.; Kokkoris, I.P.; Panitsa, M.; Trigas, P.; Strid, A.; Dimopoulos, P. Plant diversity patterns and conservation implications under climate-change scenarios in the Mediterranean: The case of Crete (Aegean, Greece). Diversity 2020, 12, 270. [Google Scholar] [CrossRef]
- Strid, A. Atlas of the Aegean Flora Part 1 (Text & Plates) & Part 2 (Maps), 1st ed.; Englera 33 (1 & 2); Botanic Garden and Botanical Museum; Freie Universität: Berlin, Germany, 2016. [Google Scholar]
- Fournaraki, C. Conservation of Threatened Plants of Crete—Seed Ecology, Operation and Management of a Gene Bank. Ph.D. Thesis, National and Kapodistrian University of Athens, Athens, Greece, 2010. (In Greek). [Google Scholar]
- Hatzilazarou, S.; Anestis, I.; Pipinis, E.; Kostas, S.; Avramakis, M.; Greveniotis, V.; Dariotis, E.; Tsoktrouridis, G.; Krigas, N. GIS-Facilitated seed germination of six local endemic plants of Crete (Greece) and multifaceted evaluation in three economic sectors. J. Biol. Res. 2023, 30, 5. [Google Scholar] [CrossRef]
- Geneve, R. Impact of temperature on seed dormancy. HortScience 2003, 38, 336–341. [Google Scholar] [CrossRef]
- Fenner, M.; Thompson, K. The Ecology of Seeds; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Koutsovoulou, A. Adaptation Strategies and Ecophysiology of Seed Germination in the Campanulaceae Family. Ph.D. Thesis, National and Kapodistrian University of Athens (EKPA), Athens, Greece, 2014. [Google Scholar]
- Kim, H.M.; Kim, J.H.; Lee, J.H.; Kim, G.M.; Lee, M.H.; Park, C.Y.; Kim, D.H.; Lee, D.H.; Kim, K.M.; Na, C.S. Dormancy-release and germination improvement of Korean bellflower (Campanula takesimana Nakai), a rare and endemic plant native to the Korean peninsula. PLoS ONE 2023, 18, e0292280. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and, Evolution of Dormancy and Germination, 2nd ed.; Elsevier: San Diego, CA, USA, 2014. [Google Scholar]
- Parra Quijano, M.; Iriondo, J.; Torres, E. Review. Applications of ecogeography and geographic information systems in conservation and utilization of plant genetic resources. Span. J. Agric. Res. 2012, 10, 419–429. [Google Scholar] [CrossRef]
- Wu, J.; Li, X.; Huang, L.; Meng, X.; Hu, H.; Luo, L.; Chen, S. A new GIS model for ecologically suitable distributions of medicinal plants. Chin. Med. 2019, 14, 4. [Google Scholar] [CrossRef] [PubMed]
- Anestis, I.; Pipinis, E.; Kostas, S.; Karapatzak, E.; Dariotis, E.; Paradeisopoulou, V.; Greveniotis, V.; Tsoktouridis, G.; Hatzilazarou, S.; Krigas, N. GIS-Facilitated germination of stored seeds from four wild-growing populations of Petromarula pinnata (L.) A. DC.—A valuable, yet Vulnerable local endemic Plant of Crete (Greece). Agronomy 2024, 14, 274. [Google Scholar] [CrossRef]
- Draper, D.; Rosselló-Graell, A.; Garcia, C.; Tauleigne Gomes, C.; Sérgio, C. Application of GIS in plant conservation programmes in Portugal. Biol. Conserv. 2003, 113, 337–349. [Google Scholar] [CrossRef]
- Cretan Flora. Available online: https://www.cretanflora.com/campanula_cretica.html (accessed on 13 April 2025).
- Lazarina, M.; Charalampopoulos, A.; Psaralexi, M.; Krigas, N.; Michailidou, D.-E.; Kallimanis, A.S.; Sgardelis, S.P. Diversity patterns of different life forms of plants along an elevational gradient in Crete, Greece. Diversity 2019, 11, 200. [Google Scholar] [CrossRef]
- WorldClim. Available online: https://www.worldclim.org/data/worldclim21.html (accessed on 13 January 2020).
- Ellis, R.H.; Roberts, E.H. The quantification of ageing and survival in orthodox seeds. Seed Sci. Technol. 1981, 9, 373–409. [Google Scholar]
- Gomez, K.A.; Gomez, A.A. Statistical Procedures in Agricultural Research, 2nd ed.; Wiley: New York, NY, USA, 1984; ISBN 0-471-87092-7. [Google Scholar]
- Klockars, A.; Sax, G. Multiple Comparisons; Sage Publications: Newbury Park, CA, USA, 1986; p. 87. ISBN 0-8039-2051-2. [Google Scholar]
- Sharrock, S.; Jones, M. Saving Europe’s threatened flora: Progress towards GSPC Target 8 in Europe. Biodivers. Conserv. 2009, 20, 325–333. [Google Scholar] [CrossRef]
- Turner, S.R.; Steadman, K.J.; Vlahos, S.; Koch, J.M.; Dixon, K.W. Seed treatment optimizes benefits of seed bank storage for restoration-ready seeds: The feasibility of prestorage dormancy alleviation for mine-site revegetation. Restor. Ecol. 2013, 21, 186–192. [Google Scholar] [CrossRef]
- Kildisheva, O.A.; Dixon, K.W.; Silveira, F.A.O.; Chapman, T.; Sacco, A.D.; Mondoni, A.; Turner, S.R.; Cross, A.T. Dormancy and germination: Making every seed count in restoration. Restor. Ecol. 2020, 28, S256–S265. [Google Scholar] [CrossRef]
- Rajpurohit, D.; Jhang, T. In situ and ex situ conservation of plant genetic resources and traditional knowledge. In Plant Genetic Resources and Traditional Knowledge for Food Security; Salgotra, R.K., Gupta, B.B., Eds.; Springer: Singapore, 2016; pp. 137–162. ISBN 978-981-10-0060-7. [Google Scholar]
- Di Martino, L.; Di Cecco, V.; Di Cecco, M.; Di Santo, M.; Ciaschetti, G.; Marcantonio, G. Use of native plants for ornamental purposes to conserve plant biodiversity: Case of study of Majella National Park. J. Nat. Conserv. 2020, 56, 125839. [Google Scholar] [CrossRef]
- Fernandes, M.P.; Pinto-Cruz, C.; Almeida, E.; Emídio, M.; Simões, M.P.; Gazarini, L.; Belo, A.D. Seed germination of six Iberian endemic species—A contribution to enhance plant conservation. Plant Biosyst. 2021, 155, 1146–1152. [Google Scholar] [CrossRef]
- Seglie, L.; Scariot, V.; Larcher, F.; Devecchi, M.; Chiavazza, P.M. In vitro seed germination and seedling propagation in Campanula spp. Plant Biosyst. 2012, 146, 15–23. [Google Scholar] [CrossRef]
- Scariot, V.; Gaino, W.; Devecchi, M. Propagation and cultivation protocols for wild creeping bellflowers (Campanula rapunculoides L.). In Proceedings of the International Symposium on Advanced Technologies and Management Towards Sustainable Greenhouse Ecosystems: Greensys 2011, Athens, Greece, 5–10 June 2011; Volume 952, pp. 265–272. [Google Scholar] [CrossRef]
- Scariot, V.; Seglie, L.; Gaino, W.; Devecchi, M. Evaluation of European native bluebells for sustainable floriculture. In Proceedings of the XXVIII International Horticultural Congress on Science and Horticulture for People (IHC2010): International Symposium on Advances in Ornamentals, Landscape and Urban Horticulture, Lisbon, Portugal, 22–27 August 2010; International Society for Horticultural Science (ISHS): Leuven, Belgium, 2012; Volume 937, pp. 273–279. [Google Scholar] [CrossRef]
- De Pascale, S.; Romano, D. Potential Use of Wild Plants in Floriculture. Acta Hortic. 2019, 1240, 87–98. [Google Scholar] [CrossRef]
- Facciuto, G.; Pannuzio, M.; Puerta, A.; Sanchez, M. Cut foliage: Potentiality of native Argentine ferns as new ornamental crops. Ornam. Hortic. 2021, 27, 566–574. [Google Scholar] [CrossRef]
- Luera, P.; Gabler, C.A. Combined Effects of scarification, phytohormones, stratification, and soil type on the germination and/or seedling performance of three tamaulipan thornscrub forest species. Plants 2022, 11, 2687. [Google Scholar] [CrossRef]
- Perrino, E.V.; Wagensommer, R.P. Crop Wild Relatives (CWR) Priority in Italy: Distribution, ecology, in situ and ex situ conservation and expected actions. Sustainability 2021, 13, 1682. [Google Scholar] [CrossRef]
- Medeiros, M.B.; Valls, J.F.M.; Abreu, A.G.; Heiden, G.; Ribeiro-Silva, S.; José, S.C.B.R.; Santos, I.R.I.; Passos, A.M.A.; Burle, M.L. Status of the ex situ and in situ conservation of brazilian crop wild relatives of rice, potato, sweet potato, and finger millet: Filling the gaps of germplasm collections. Agronomy 2021, 11, 638. [Google Scholar] [CrossRef]
- Gomes, A.; Pimpão, R.C.; Fortalezas, S.; Figueira, I.; Miguel, C.; Aguiar, C.; Salgueiro, L.; Cavaleiro, C.; Gonçalves, M.J.; Clemente, A.; et al. Chemical characterization and bioactivity of phytochemicals from Iberian endemic Santolina semidentata and strategies for ex situ propagation. Ind. Crop. Prod. 2015, 74, 505–513. [Google Scholar] [CrossRef]
- Ayuso, M.; García-Pérez, P.; Ramil-Rego, P.; Gallego, P.P.; Barreal, M.E. In vitro culture of the endangered plant Eryngium viviparum as dual strategy for its ex situ conservation and source of bioactive compounds. Plant Cell Tiss. Organ Cult. 2019, 138, 427–435. [Google Scholar] [CrossRef]
- Asgher, M.; Verma, S.; Khan, N.A.; Vyas, D.; Kumari, P.; Rashid, S.; Khan, S.; Qadir, S.; Ajmal Ali, M.; Ahmad, P. Physiological, biochemical and reproductive studies on Valeriana wallichii, a critically endangered medicinal plant of the Himalayan Region grown under in-situ and ex-situ conditions. Plants 2020, 9, 131. [Google Scholar] [CrossRef]
- Graikou, K.; Mpishinioti, A.; Tsafantakis, N.; Maloupa, E.; Grigoriadou, K.; Chinou, I. Comparative phytochemical analyses of flowers from Primula veris subsp. veris growing wild and from ex situ cultivation in Greece. Foods 2023, 12, 2623. [Google Scholar] [CrossRef]
- Sana; Aftab, T.; Khan, M.M.A.; Naeem, M. Chapter 7—Overexploitation and Conservation Strategies for Medicinal and Aromatic Plants. In Essential Oil-Bearing Plants; Naeem, M., Khan, M.M.A., Eds.; Academic Press: Cambridge, MA, USA, 2025; pp. 95–105. ISBN 978-0-443-24860-3. [Google Scholar]
- Bewley, J.D.; Bradford, K.J.; Hilhorst, H.W.M.; Nonogaki, H. Dormancy and the Control of Germination. In Seeds: Physiology of Development, Germination and Dormancy, 3rd ed.; Bewley, J.D., Bradford, K.J., Hilhorst, H.W.M., Nonogaki, H., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 247–297. [Google Scholar] [CrossRef]
- Willis, C.G.; Baskin, C.C.; Baskin, J.M.; Auld, J.R.; Venable, D.L.; Cavender-Bares, J.; Donohue, K.; de Casas, R.R.; The NESCent Germination Working Group. The evolution of seed dormancy: Environmental cues, evolutionary hubs, and diversification of the seed plants. New Phytol. 2014, 203, 300–309. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Y.; Sun, L.; Baskin, C.C.; Baskin, J.M.; Cao, M.; Yang, J. Seed dormancy in space and time: Global distribution, paleoclimatic and present climatic drivers, and evolutionary adaptations. New Phytol. 2022, 234, 1770–1781. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, Z.; Xiang, Y.; Tong, X.; Wojtyla, L.; Wang, Y. Editorial: Molecular basis of seed germination and dormancy. Front. Plant Sci. 2023, 14, 1242428. [Google Scholar] [CrossRef]
- Penfield, S. Seed Dormancy and Germination. Curr. Biol. 2017, 27, R874–R878. [Google Scholar] [CrossRef]
- Wagner, M.; Pywell, R.F.; Knopp, T.; Bullock, J.M.; Heard, M.S. The germination niches of grassland species targeted for restoration: Effects of seed pre-treatments. Seed Sci. Res. 2011, 21, 117–131. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M.; Yoshinaga, A.; Wolkis, D. Seed dormancy in Campanulaceae: Morphological and morphophysiological dormancy in six species of Hawaiian Lobelioids. Botany 2020, 98, 327–332. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C. A Classification system for seed dormancy. Seed Sci. Res. 2004, 14, 1–16. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Underdeveloped embryos in dwarf seeds and implications for assignment to dormancy class. Seed Sci. Res. 2005, 15, 357–360. [Google Scholar] [CrossRef]
- Baskin, C.; Baskin, J.; Yoshinaga, A. Morphophysiological dormancy in seeds of six endemic Lobelioid shrubs (Campanulaceae) from the Montane Zone in Hawaii. Can. J. Bot. 2011, 83, 1630–1637. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Sum of Squares | df | Mean Square | F | Sig. (p-Value) |
|---|---|---|---|---|---|
| Temperature | 8484.15 | 3 | 2828.05 | 132.26 | <0.001 |
| Pre-treatment | 4951.43 | 3 | 1650.48 | 77.19 | <0.001 |
| Temperature × pre-treatment | 6346.03 | 9 | 705.11 | 32.99 | <0.001 |
| Pre-Treatment | Incubation Temperature | |||
|---|---|---|---|---|
| 10 °C | 15 °C | 20 °C | 25 °C | |
| Control | 90.00 a 1 ± 2.04 | 85.00 a ± 4.08 | 56.88 b ± 8.98 | 0.63 c ± 1.25 |
| 0.5 g·L−1 GA3 | 93.13 a ± 5.54 | 87.50 ab ± 4.56 | 89.38 ab ± 2.39 | 81.88 b ± 6.25 |
| 1 g·L−1 GA3 | 86.25 a ± 5.95 | 87.50 a ± 7.07 | 88.13 a ± 6.88 | 76.25 b ± 4.79 |
| CS | 90.63 a ± 4.27 | 85.63 a ± 5.54 | 74.38 b ± 5.91 | 43.75 c ± 6.61 |
| Factor | Germination (%) | Mean Germination Time (Days) |
|---|---|---|
| Light/dark | 94.38 a 1 ± 1.25 | 9.70 a ± 0.80 |
| Dark | 69.38 b ± 6.88 | 15.34 b ± 1.21 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panagiotidou, T.-N.; Anestis, I.; Pipinis, E.; Kostas, S.; Tsoktouridis, G.; Hatzilazarou, S.; Krigas, N. GIS Bioclimatic Profile and Seed Germination of the Endangered and Protected Cretan Endemic Plant Campanula cretica (A. DC.) D. Dietr. for Conservation and Sustainable Utilization. Agriculture 2025, 15, 1161. https://doi.org/10.3390/agriculture15111161
Panagiotidou T-N, Anestis I, Pipinis E, Kostas S, Tsoktouridis G, Hatzilazarou S, Krigas N. GIS Bioclimatic Profile and Seed Germination of the Endangered and Protected Cretan Endemic Plant Campanula cretica (A. DC.) D. Dietr. for Conservation and Sustainable Utilization. Agriculture. 2025; 15(11):1161. https://doi.org/10.3390/agriculture15111161
Chicago/Turabian StylePanagiotidou, Theodora-Nafsika, Ioannis Anestis, Elias Pipinis, Stefanos Kostas, Georgios Tsoktouridis, Stefanos Hatzilazarou, and Nikos Krigas. 2025. "GIS Bioclimatic Profile and Seed Germination of the Endangered and Protected Cretan Endemic Plant Campanula cretica (A. DC.) D. Dietr. for Conservation and Sustainable Utilization" Agriculture 15, no. 11: 1161. https://doi.org/10.3390/agriculture15111161
APA StylePanagiotidou, T.-N., Anestis, I., Pipinis, E., Kostas, S., Tsoktouridis, G., Hatzilazarou, S., & Krigas, N. (2025). GIS Bioclimatic Profile and Seed Germination of the Endangered and Protected Cretan Endemic Plant Campanula cretica (A. DC.) D. Dietr. for Conservation and Sustainable Utilization. Agriculture, 15(11), 1161. https://doi.org/10.3390/agriculture15111161