
Addressing Biological Invasions in Agriculture with Big Data in an Informatics Age

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
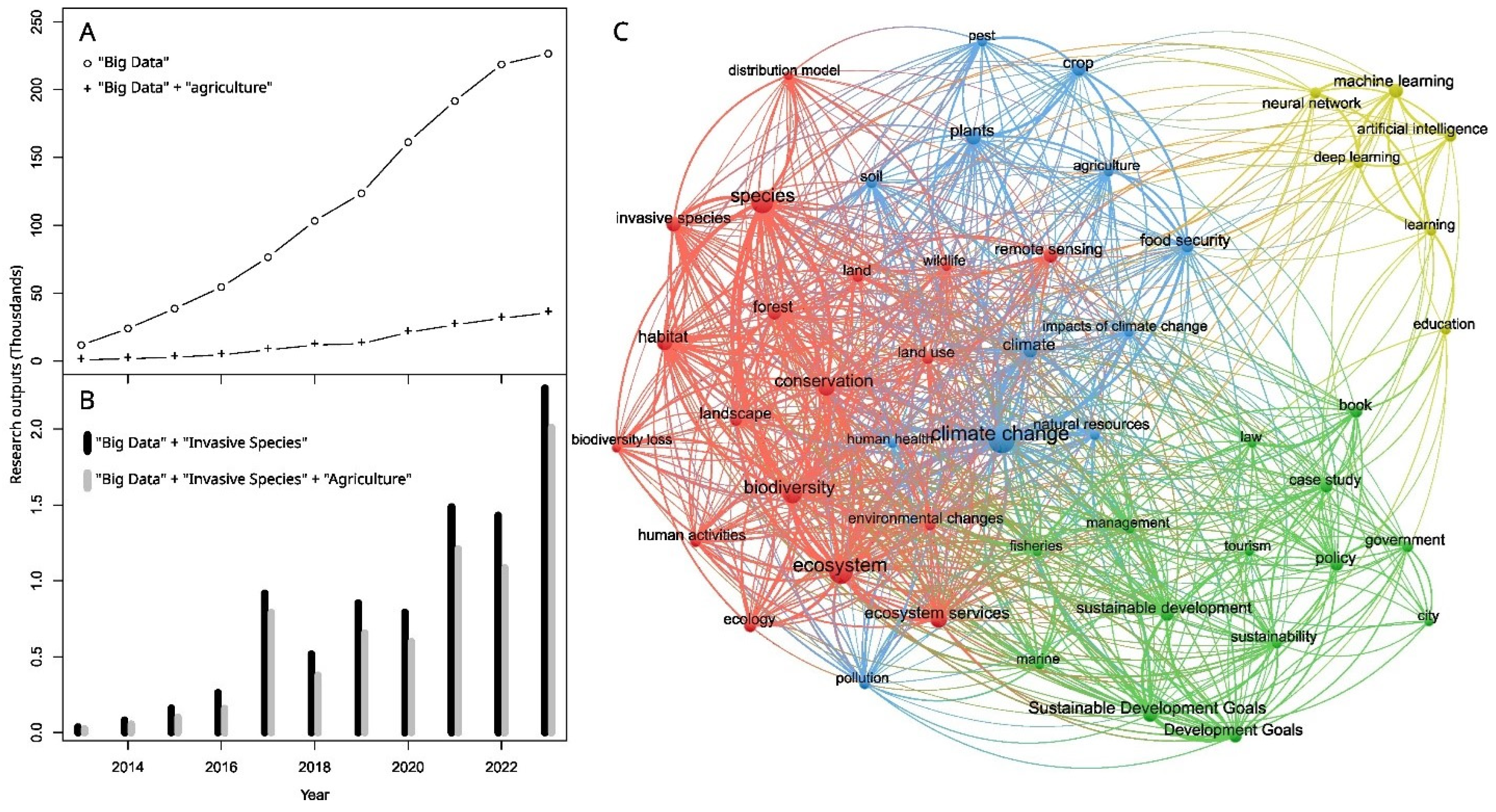
1. Introduction
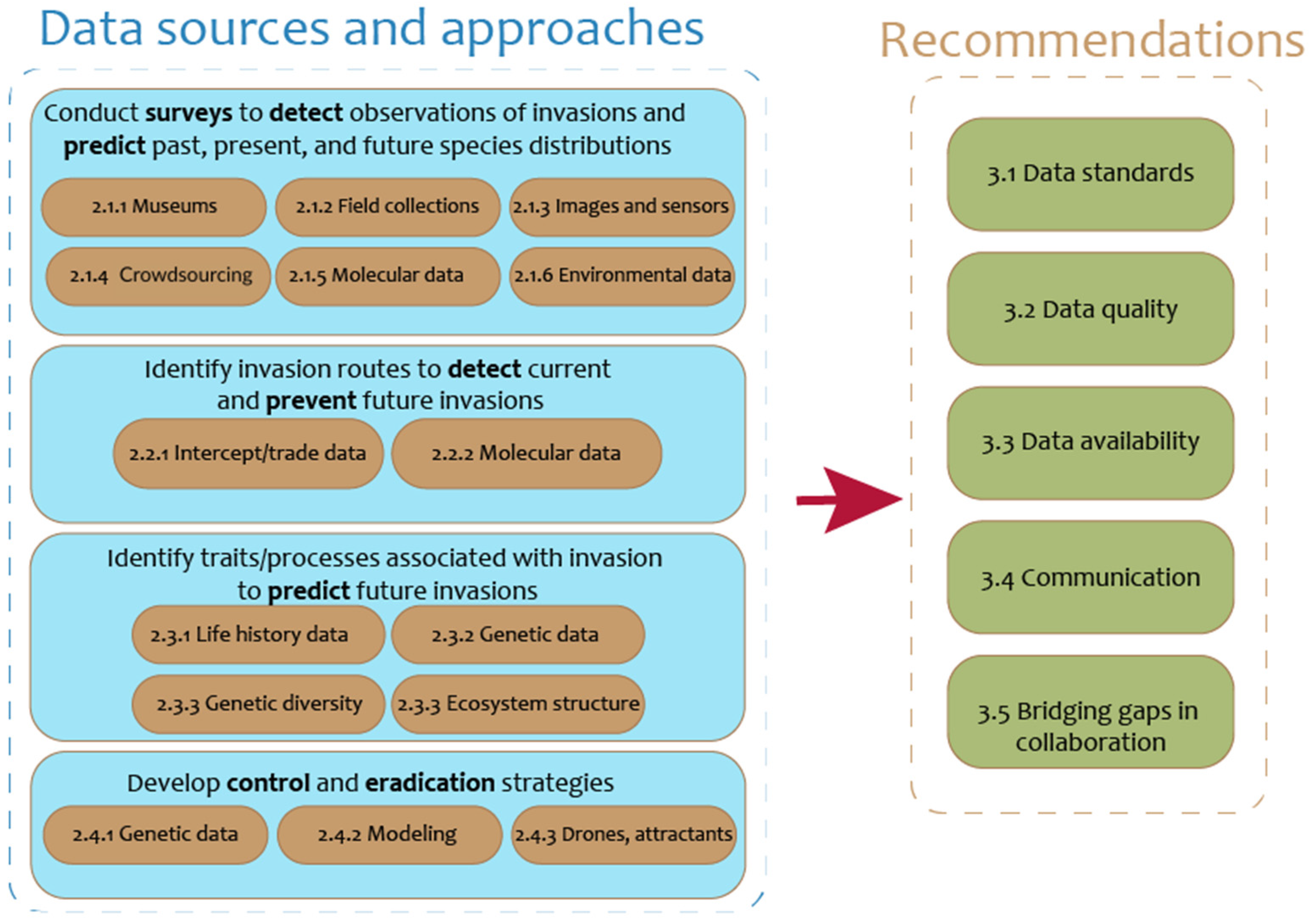
2. Data Are Used to Predict, Detect, Prevent, Control, and Eradicate Biological Invasions in Agriculture
2.1. Reconstructing Species Distributions with Occurrence Data
2.1.1. Museum Collections Are Key to Taxonomy and Occurrence Data
2.1.2. Monitoring with Field Sampling, Surveys, and Traps
2.1.3. Occurrence Data and Taxonomy from Image and Sensor Data
2.1.4. Crowdsourcing Observation Data
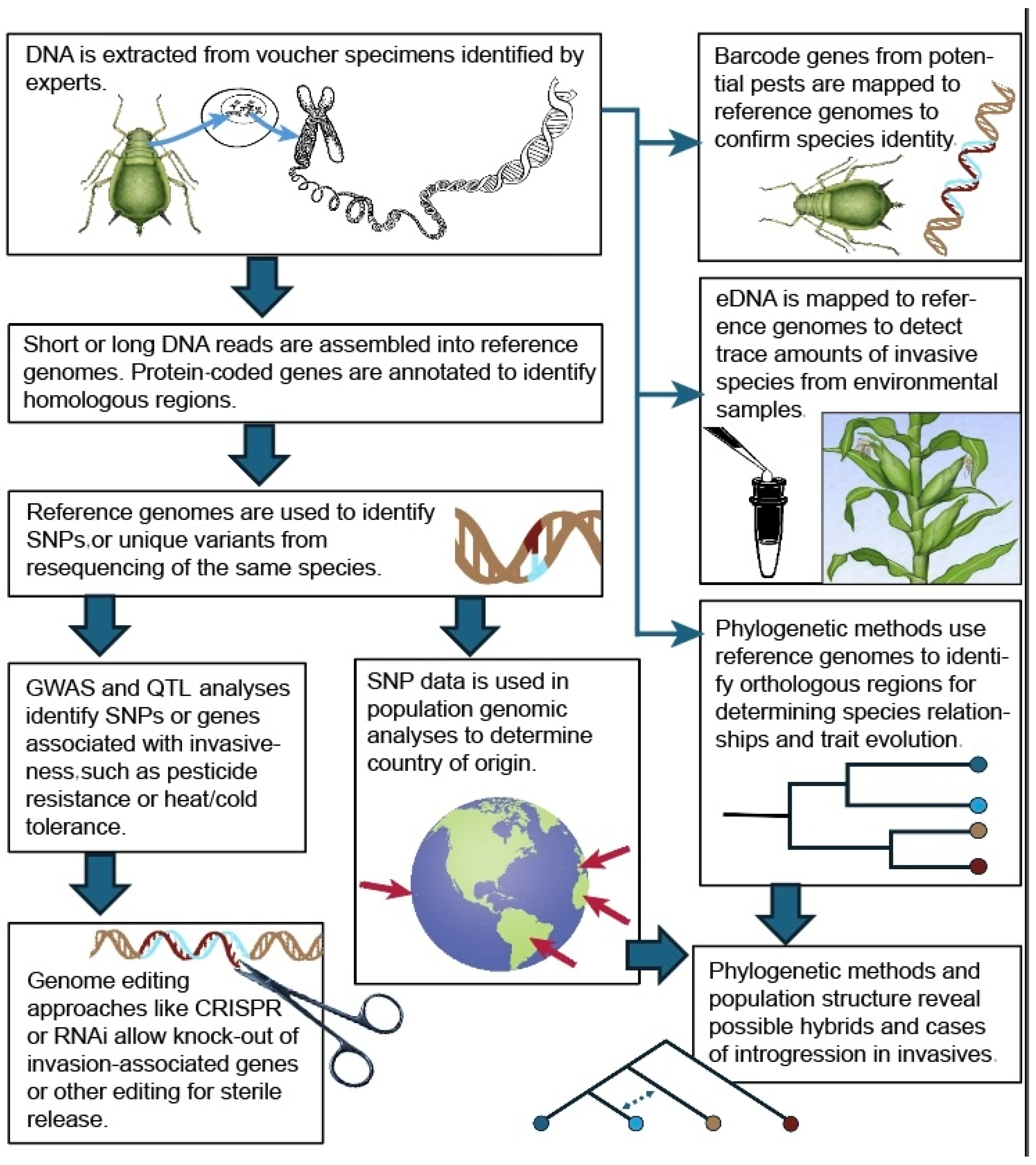
2.1.5. Observation Through Molecular Taxonomy and Sampling eDNA
2.1.6. Combining Occurrence and Environmental Data to Predict Species Distributions and Abundance
2.2. Identifying Invasion Routes
2.2.1. Intercept/Trade Route Data
2.2.2. DNA Data for Population Genetics
2.3. Traits and Processes Associated with Invasion
2.3.1. Life History Data
2.3.2. Genetic Traits from DNA Data
2.3.3. Importance of Genetic Diversity in Invaders
2.3.4. Ecosystem Structure and Function
2.4. Developing and Evaluating Control and Eradication Methods
2.4.1. Genetic Data for Control Methods
2.4.2. Modeling for Control Methods
2.4.3. Use of Drones and AI for Detecting and Controlling Invasive Species
3. Moving Forward: Key Tools, Obstacles, and Challenges
3.1. Data Standards
3.2. Data Quality, Taxonomic Accuracy, and Awareness of Data Bias
3.3. Proper Data Storage and Data Availability
3.4. Communication About and Between Data Sources
3.5. Bridging Gaps in Collaboration
4. Conclusions
5. Glossary Terms
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Laney, D. 3D Data Management: Controlling Data Volume, Velocity and Variety. META Group Res. Note 2001, 6, 1. [Google Scholar]
- Dash, S.; Shakyawar, S.K.; Sharma, M.; Kaushik, S. Big Data in Healthcare: Management, Analysis and Future Prospects. J. Big Data 2019, 6, 54. [Google Scholar] [CrossRef]
- Gopalkrishnan, V.; Steier, D.; Lewis, H.; Guszcza, J. Big Data, Big Business: Bridging the Gap. In Proceedings of the 1st International Workshop on Big Data, Streams and Heterogeneous Source Mining: Algorithms, Systems, Programming Models and Applications, Beijing, China, 12 August 2012; ACM: Beijing, China, 2012; pp. 7–11. [Google Scholar]
- Hasan, M.M.; Popp, J.; Oláh, J. Current Landscape and Influence of Big Data on Finance. J. Big Data 2020, 7, 21. [Google Scholar] [CrossRef]
- Allam, Z.; Dhunny, Z.A. On Big Data, Artificial Intelligence and Smart Cities. Cities 2019, 89, 80–91. [Google Scholar] [CrossRef]
- Neilson, A.; Indratmo; Daniel, B.; Tjandra, S. Systematic Review of the Literature on Big Data in the Transportation Domain: Concepts and Applications. Big Data Res. 2019, 17, 35–44. [Google Scholar] [CrossRef]
- Bunge, J. Big Data Comes to the Farm, Sowing Mistrust. Wall Str. J. 2014. Available online: https://www.wsj.com/articles/SB10001424052702304450904579369283869192124 (accessed on 21 May 2025).
- Lokers, R.; Knapen, R.; Janssen, S.; van Randen, Y.; Jansen, J. Analysis of Big Data Technologies for Use in Agro-Environmental Science. Environ. Model. Softw. 2016, 84, 494–504. [Google Scholar] [CrossRef]
- Kamilaris, A.; Kartakoullis, A.; Prenafeta-Boldú, F.X. A Review on the Practice of Big Data Analysis in Agriculture. Comput. Electron. Agric. 2017, 143, 23–37. [Google Scholar] [CrossRef]
- Carruthers, R.I. Invasive Species Research in the United States Department of Agriculture–Agricultural Research Service. Pest. Manag. Sci. 2003, 59, 827–834. [Google Scholar] [CrossRef]
- Cuthbert, R.N.; Diagne, C.; Haubrock, P.J.; Turbelin, A.J.; Courchamp, F. Are the “100 of the World’s Worst” Invasive Species Also the Costliest? Biol. Invasions 2022, 24, 1895–1904. [Google Scholar] [CrossRef]
- Diagne, C.; Leroy, B.; Vaissière, A.-C.; Gozlan, R.E.; Roiz, D.; Jarić, I.; Salles, J.-M.; Bradshaw, C.J.A.; Courchamp, F. High and Rising Economic Costs of Biological Invasions Worldwide. Nature 2021, 592, 571–576. [Google Scholar] [CrossRef] [PubMed]
- Guillemaud, T.; Ciosi, M.; Lombaert, É.; Estoup, A. Biological Invasions in Agricultural Settings: Insights from Evolutionary Biology and Population Genetics. Comptes Rendus Biol. 2011, 334, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Grandy, A.S.; Wieder, W.R.; Wickings, K.; Kyker-Snowman, E. Beyond Microbes: Are Fauna the next Frontier in Soil Biogeochemical Models? Soil Biol. Biochem. 2016, 102, 40–44. [Google Scholar] [CrossRef]
- Paini, D.R.; Sheppard, A.W.; Cook, D.C.; De Barro, P.J.; Worner, S.P.; Thomas, M.B. Global Threat to Agriculture from Invasive Species. Proc. Natl. Acad. Sci. USA 2016, 113, 7575–7579. [Google Scholar] [CrossRef]
- Ziska, L.H.; Blumenthal, D.M.; Runion, G.B.; Hunt, E.R.; Diaz-Soltero, H. Invasive Species and Climate Change: An Agronomic Perspective. Clim. Change 2011, 105, 13–42. [Google Scholar] [CrossRef]
- Dedryver, C.-A.; Le Ralec, A.; Fabre, F. The Conflicting Relationships between Aphids and Men: A Review of Aphid Damage and Control Strategies. Comptes Rendus Biol. 2010, 333, 539–553. [Google Scholar] [CrossRef]
- Clubley, C.H.; Firth, L.B.; Wood, L.E.; Bilton, D.T.; Silva, T.A.M.; Knights, A.M. Science Paper or Big Data? Assessing Invasion Dynamics Using Observational Data. Sci. Total Environ. 2023, 877, 162754. [Google Scholar] [CrossRef]
- Ahmed, I.; Ahmad, M.; Jeon, G.; Piccialli, F. A Framework for Pandemic Prediction Using Big Data Analytics. Big Data Res. 2021, 25, 100190. [Google Scholar] [CrossRef]
- Kamenova, S.; Bartley, T.J.; Bohan, D.A.; Boutain, J.R.; Colautti, R.I.; Domaizon, I.; Fontaine, C.; Lemainque, A.; Le Viol, I.; Mollot, G.; et al. Chapter Three—Invasions Toolkit: Current Methods for Tracking the Spread and Impact of Invasive Species. In Advances in Ecological Research; Bohan, D.A., Dumbrell, A.J., Massol, F., Eds.; Networks of Invasion: A Synthesis of Concepts; Academic Press: Cambridge, MA, USA, 2017; Volume 56, pp. 85–182. [Google Scholar]
- Robertson, T.; Döring, M.; Guralnick, R.; Bloom, D.; Wieczorek, J.; Braak, K.; Otegui, J.; Russell, L.; Desmet, P. The GBIF Integrated Publishing Toolkit: Facilitating the Efficient Publishing of Biodiversity Data on the Internet. PLoS ONE 2014, 9, e102623. [Google Scholar] [CrossRef]
- Wallace, R.D.; Bargeron, C.T.; Reaser, J.K. Enabling Decisions That Make a Difference: Guidance for Improving Access to and Analysis of Invasive Species Information. Biol. Invasions 2020, 22, 37–45. [Google Scholar] [CrossRef]
- Short, A.E.Z.; Dikow, T.; Moreau, C.S. Entomological Collections in the Age of Big Data. Annu. Rev. Entomol. 2018, 63, 513–530. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Cordero, V.; Martínez-Meyer, E. Museum Specimen Data Predict Crop Damage by Tropical Rodents. Proc. Natl. Acad. Sci. USA 2000, 97, 7074–7077. [Google Scholar] [CrossRef] [PubMed]
- Guralnick, R.; Van Cleve, J. Strengths and Weaknesses of Museum and National Survey Data Sets for Predicting Regional Species Richness: Comparative and Combined Approaches. Divers. Distrib. 2005, 11, 349–359. [Google Scholar] [CrossRef]
- Suarez, A.V.; Tsutsui, N.D. The Value of Museum Collections for Research and Society. BioScience 2004, 54, 66. [Google Scholar] [CrossRef]
- Reaser, J.K.; Burgiel, S.W.; Kirkey, J.; Brantley, K.A.; Veatch, S.D.; Burgos-Rodríguez, J. The Early Detection of and Rapid Response (EDRR) to Invasive Species: A Conceptual Framework and Federal Capacities Assessment. Biol. Invasions 2020, 22, 1–19. [Google Scholar] [CrossRef]
- Juanes, F. Visual and Acoustic Sensors for Early Detection of Biological Invasions: Current Uses and Future Potential. J. Nat. Conserv. 2018, 42, 7–11. [Google Scholar] [CrossRef]
- Batz, P.; Will, T.; Thiel, S.; Ziesche, T.M.; Joachim, C. From Identification to Forecasting: The Potential of Image Recognition and Artificial Intelligence for Aphid Pest Monitoring. Front. Plant Sci. 2023, 14, 1150748. [Google Scholar] [CrossRef]
- Delisle, Z.J.; Flaherty, E.A.; Nobbe, M.R.; Wzientek, C.M.; Swihart, R.K. Next-Generation Camera Trapping: Systematic Review of Historic Trends Suggests Keys to Expanded Research Applications in Ecology and Conservation. Front. Ecol. Evol. 2021, 9, 617996. [Google Scholar] [CrossRef]
- Diller, Y.; Shamsian, A.; Shaked, B.; Altman, Y.; Danziger, B.-C.; Manrakhan, A.; Serfontein, L.; Bali, E.; Wernicke, M.; Egartner, A.; et al. A Real-Time Remote Surveillance System for Fruit Flies of Economic Importance: Sensitivity and Image Analysis. J. Pest. Sci. 2023, 96, 611–622. [Google Scholar] [CrossRef]
- Jiang, J.-A.; Tseng, C.-L.; Lu, F.-M.; Yang, E.-C.; Wu, Z.-S.; Chen, C.-P.; Lin, S.-H.; Lin, K.-C.; Liao, C.-S. A GSM-Based Remote Wireless Automatic Monitoring System for Field Information: A Case Study for Ecological Monitoring of the Oriental Fruit Fly, Bactrocera dorsalis (Hendel). Comput. Electron. Agric. 2008, 62, 243–259. [Google Scholar] [CrossRef]
- Jiang, J.-A.; Lin, T.-S.; Yang, E.-C.; Tseng, C.-L.; Chen, C.-P.; Yen, C.-W.; Zheng, X.-Y.; Liu, C.-Y.; Liu, R.-H.; Chen, Y.-F.; et al. Application of a Web-Based Remote Agro-Ecological Monitoring System for Observing Spatial Distribution and Dynamics of Bactrocera dorsalis in Fruit Orchards. Precis. Agric. 2013, 14, 323–342. [Google Scholar] [CrossRef]
- Liao, M.-S.; Chuang, C.-L.; Lin, T.-S.; Chen, C.-P.; Zheng, X.-Y.; Chen, P.-T.; Liao, K.-C.; Jiang, J.-A. Development of an Autonomous Early Warning System for Bactrocera dorsalis (Hendel) Outbreaks in Remote Fruit Orchards. Comput. Electron. Agric. 2012, 88, 1–12. [Google Scholar] [CrossRef]
- Goldshtein, E.; Cohen, Y.; Hetzroni, A.; Gazit, Y.; Timar, D.; Rosenfeld, L.; Grinshpon, Y.; Hoffman, A.; Mizrach, A. Development of an Automatic Monitoring Trap for Mediterranean Fruit Fly (Ceratitis capitata) to Optimize Control Applications Frequency. Comput. Electron. Agric. 2017, 139, 115–125. [Google Scholar] [CrossRef]
- Welsh, T.J.; Bentall, D.; Kwon, C.; Mas, F. Automated Surveillance of Lepidopteran Pests with Smart Optoelectronic Sensor Traps. Sustainability 2022, 14, 9577. [Google Scholar] [CrossRef]
- Chen, Y.; Why, A.; Batista, G.; Mafra-neto, A.; Keogh, E. Flying Insect Classification with Inexpensive Sensors. J. Insect Behav. 2014, 27, 657–677. [Google Scholar] [CrossRef]
- Rydhmer, K.; Eckberg, J.O.; Lundgren, J.G.; Jansson, S.; Still, L.; Quinn, J.E.; Jr, R.W.; Lemmich, J.; Nikolajsen, T.; Sheller, N.; et al. Automating an Insect Biodiversity Metric Using Distributed Optical Sensors: An Evaluation across Kansas, USA Cropping Systems. eLife 2024, 13, RP92227. [Google Scholar] [CrossRef]
- Rydhmer, K.; Bick, E.; Still, L.; Strand, A.; Luciano, R.; Helmreich, S.; Beck, B.D.; Grønne, C.; Malmros, L.; Poulsen, K.; et al. Automating Insect Monitoring Using Unsupervised Near-Infrared Sensors. Sci. Rep. 2022, 12, 2603. [Google Scholar] [CrossRef]
- Lello, F.; Dida, M.; Mkiramweni, M.; Matiko, J.; Akol, R.; Nsabagwa, M.; Katumba, A. Fruit Fly Automatic Detection and Monitoring Techniques: A Review. Smart Agric. Technol. 2023, 5, 100294. [Google Scholar] [CrossRef]
- Su, W.-H. Advanced Machine Learning in Point Spectroscopy, RGB- and Hyperspectral-Imaging for Automatic Discriminations of Crops and Weeds: A Review. Smart Cities 2020, 3, 767–792. [Google Scholar] [CrossRef]
- Klarica, J.; Bittner, L.; Pallua, J.; Pezzei, C.; Huck-Pezzei, V.; Dowell, F.; Schied, J.; Bonn, G.K.; Huck, C.; Schlick-Steiner, B.C.; et al. Near-Infrared Imaging Spectroscopy as a Tool to Discriminate Two Cryptic Tetramorium Ant Species. J. Chem. Ecol. 2011, 37, 549–552. [Google Scholar] [CrossRef]
- Nansen, C.; Coelho, A., Jr.; Vieira, J.M.; Parra, J.R.P. Reflectance-Based Identification of Parasitized Host Eggs and Adult Trichogramma Specimens. J. Exp. Biol. 2014, 217, 1187–1192. [Google Scholar] [CrossRef] [PubMed]
- Iost Filho, F.H.; Heldens, W.B.; Kong, Z.; de Lange, E.S. Drones: Innovative Technology for Use in Precision Pest Management. J. Econ. Entomol. 2020, 113, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Dao, P.; Liu, J.; He, Y.; Shang, J. Recent Advances of Hyperspectral Imaging Technology and Applications in Agriculture. Remote Sens. 2020, 12, 2659. [Google Scholar] [CrossRef]
- Kellenberger, B.; Tuia, D.; Morris, D. AIDE: Accelerating Image-Based Ecological Surveys with Interactive Machine Learning. Methods Ecol. Evol. 2020, 11, 1716–1727. [Google Scholar] [CrossRef]
- Wäldchen, J.; Mäder, P. Machine Learning for Image Based Species Identification. Methods Ecol. Evol. 2018, 9, 2216–2225. [Google Scholar] [CrossRef]
- Teixeira, A.C.; Ribeiro, J.; Morais, R.; Sousa, J.J.; Cunha, A. A Systematic Review on Automatic Insect Detection Using Deep Learning. Agriculture 2023, 13, 713. [Google Scholar] [CrossRef]
- Tuia, D.; Kellenberger, B.; Beery, S.; Costelloe, B.R.; Zuffi, S.; Risse, B.; Mathis, A.; Mathis, M.W.; van Langevelde, F.; Burghardt, T.; et al. Perspectives in Machine Learning for Wildlife Conservation. Nat. Commun. 2022, 13, 792. [Google Scholar] [CrossRef]
- Goodwin, Z.A.; Harris, D.J.; Filer, D.; Wood, J.R.I.; Scotland, R.W. Widespread Mistaken Identity in Tropical Plant Collections. Curr. Biol. 2015, 25, R1066–R1067. [Google Scholar] [CrossRef]
- Tian, J.; Wang, L.; Yin, D.; Li, X.; Diao, C.; Gong, H.; Shi, C.; Menenti, M.; Ge, Y.; Nie, S.; et al. Development of Spectral-Phenological Features for Deep Learning to Understand Spartina alterniflora Invasion. Remote Sens. Environ. 2020, 242, 111745. [Google Scholar] [CrossRef]
- August, T.; Fox, R.; Roy, D.B.; Pocock, M.J.O. Data-Derived Metrics Describing the Behaviour of Field-Based Citizen Scientists Provide Insights for Project Design and Modelling Bias. Sci. Rep. 2020, 10, 11009. [Google Scholar] [CrossRef]
- Müllerová, J.; Brundu, G.; Große-Stoltenberg, A.; Kattenborn, T.; Richardson, D.M. Pattern to Process, Research to Practice: Remote Sensing of Plant Invasions. Biol. Invasions 2023, 25, 3651–3676. [Google Scholar] [CrossRef]
- Callaghan, C.T.; Mesaglio, T.; Ascher, J.S.; Brooks, T.M.; Cabras, A.A.; Chandler, M.; Cornwell, W.K.; Ríos-Málaver, I.C.; Dankowicz, E.; Dhiya’ulhaq, N.U.; et al. The Benefits of Contributing to the Citizen Science Platform iNaturalist as an Identifier. PLoS Biol. 2022, 20, e3001843. [Google Scholar] [CrossRef] [PubMed]
- Putman, B.J.; Williams, R.; Li, E.; Pauly, G.B. The Power of Community Science to Quantify Ecological Interactions in Cities. Sci. Rep. 2021, 11, 3069. [Google Scholar] [CrossRef] [PubMed]
- Site Stats · iNaturalist. Available online: https://www.inaturalist.org/stats (accessed on 15 July 2024).
- Callaghan, C.T.; Ozeroff, I.; Hitchcock, C.; Chandler, M. Capitalizing on Opportunistic Citizen Science Data to Monitor Urban Biodiversity: A Multi-Taxa Framework. Biol. Conserv. 2020, 251, 108753. [Google Scholar] [CrossRef]
- Pocock, M.J.O.; Tweddle, J.C.; Savage, J.; Robinson, L.D.; Roy, H.E. The Diversity and Evolution of Ecological and Environmental Citizen Science. PLoS ONE 2017, 12, e0172579. [Google Scholar] [CrossRef]
- Botan, A.L.M.; Heringer, G.; de Matos, A.C.L.; Oliveira, D.L.; de Alvarenga, D.R.; Almeida, J.W.; Tavares, K.P.; Bueno, M.L.; Lopes, V.H.; Zenni, R.D. Use of a Citizen Science Tool for the Determination of Biological Invasions in Urban Areas. bioRxiv 2022. [Google Scholar] [CrossRef]
- Hulbert, J.M.; Hallett, R.A.; Roy, H.E.; Cleary, M. Citizen Science Can Enhance Strategies to Detect and Manage Invasive Forest Pests and Pathogens. Front. Ecol. Evol. 2023, 11, 1113978. [Google Scholar] [CrossRef]
- Kelling, S.; Johnston, A.; Bonn, A.; Fink, D.; Ruiz-Gutierrez, V.; Bonney, R.; Fernandez, M.; Hochachka, W.M.; Julliard, R.; Kraemer, R.; et al. Using Semistructured Surveys to Improve Citizen Science Data for Monitoring Biodiversity. BioScience 2019, 69, 170–179. [Google Scholar] [CrossRef]
- Tiago, P.; Evaristo, I.; Pinto, B. The Role of BioBlitzes in Citizen Science: Insights from Participants and Experts. Front. Environ. Sci. 2024, 12, 1347428. [Google Scholar] [CrossRef]
- Niemiller, K.D.K.; Davis, M.A.; Niemiller, M.L. Addressing ‘Biodiversity Naivety’ through Project-Based Learning Using iNaturalist. J. Nat. Conserv. 2021, 64, 126070. [Google Scholar] [CrossRef]
- Carney, R.M.; Mapes, C.; Low, R.D.; Long, A.; Bowser, A.; Durieux, D.; Rivera, K.; Dekramanjian, B.; Bartumeus, F.; Guerrero, D.; et al. Integrating Global Citizen Science Platforms to Enable Next-Generation Surveillance of Invasive and Vector Mosquitoes. Insects 2022, 13, 675. [Google Scholar] [CrossRef]
- Jones, E.F.; Frei, R.J.; Lee, R.M.; Maxwell, J.D.; Shoemaker, R.; Follett, A.P.; Lawson, G.M.; Malmfeldt, M.; Watts, R.; Aanderud, Z.T.; et al. Citizen Science Reveals Unexpected Solute Patterns in Semiarid River Networks. PLoS ONE 2021, 16, e0255411. [Google Scholar] [CrossRef] [PubMed]
- Daume, S. Mining Twitter to Monitor Invasive Alien Species—An Analytical Framework and Sample Information Topologies. Ecol. Inform. 2016, 31, 70–82. [Google Scholar] [CrossRef]
- Tateosian, L.G.; Saffer, A.; Walden-Schreiner, C.; Shukunobe, M. Plant Pest Invasions, as Seen through News and Social Media. Comput. Environ. Urban. Syst. 2023, 100, 101922. [Google Scholar] [CrossRef]
- Lee, S.-B.; Lee, H.; Song, J.; Jang, B.; Cho, S.M.; Yum, J.; Ahn, N.-H.; Kim, J.; Lee, H.; Choi, Y.-S.; et al. A Post in an Internet Forum Led to a Discovery of an Invasive Drywood Termite in Korea, Cryptotermes domesticus (Haviland) (Blattodea: Kalotermitidae). J. Integr. Pest. Manag. 2024, 15, 34. [Google Scholar] [CrossRef]
- Encarnação, J.; Baptista, V.; Teodósio, M.A.; Morais, P. Low-Cost Citizen Science Effectively Monitors the Rapid Expansion of a Marine Invasive Species. Front. Environ. Sci. 2021, 9, 752705. [Google Scholar] [CrossRef]
- Minet, J.; Curnel, Y.; Gobin, A.; Goffart, J.-P.; Mélard, F.; Tychon, B.; Wellens, J.; Defourny, P. Crowdsourcing for Agricultural Applications: A Review of Uses and Opportunities for a Farmsourcing Approach. Comput. Electron. Agric. 2017, 142, 126–138. [Google Scholar] [CrossRef]
- Paini, D.R.; Worner, S.P.; Cook, D.C.; De Barro, P.J.; Thomas, M.B. Threat of Invasive Pests from within National Borders. Nat. Commun. 2010, 1, 115. [Google Scholar] [CrossRef]
- Johnson, B.A.; Mader, A.D.; Dasgupta, R.; Kumar, P. Citizen Science and Invasive Alien Species: An Analysis of Citizen Science Initiatives Using Information and Communications Technology (ICT) to Collect Invasive Alien Species Observations. Glob. Ecol. Conserv. 2020, 21, e00812. [Google Scholar] [CrossRef]
- Ebach, M.C.; Valdecasas, A.G.; Wheeler, Q.D. Impediments to Taxonomy and Users of Taxonomy: Accessibility and Impact Evaluation. Cladistics 2011, 27, 550–557. [Google Scholar] [CrossRef]
- Pennisi, E. Shuttered Natural History Museums Fight for Survival. Science 2020, 368, 1042–1043. [Google Scholar] [CrossRef]
- Crall, A.W.; Jarnevich, C.S.; Young, N.E.; Panke, B.J.; Renz, M.; Stohlgren, T.J. Citizen Science Contributes to Our Knowledge of Invasive Plant Species Distributions. Biol. Invasions 2015, 17, 2415–2427. [Google Scholar] [CrossRef]
- Cappa, F.; Franco, S.; Rosso, F. Citizens and Cities: Leveraging Citizen Science and Big Data for Sustainable Urban Development. Bus. Strategy Environ. 2022, 31, 648–667. [Google Scholar] [CrossRef]
- Soriano-Redondo, A.; Correia, R.A.; Barve, V.; Brooks, T.M.; Butchart, S.H.M.; Jarić, I.; Kulkarni, R.; Ladle, R.J.; Vaz, A.S.; Minin, E.D. Harnessing Online Digital Data in Biodiversity Monitoring. PLoS Biol. 2024, 22, e3002497. [Google Scholar] [CrossRef]
- Osinga, S.A.; Paudel, D.; Mouzakitis, S.A.; Athanasiadis, I.N. Big Data in Agriculture: Between Opportunity and Solution. Agric. Syst. 2022, 195, 103298. [Google Scholar] [CrossRef]
- Di Cecco, G.J.; Barve, V.; Belitz, M.W.; Stucky, B.J.; Guralnick, R.P.; Hurlbert, A.H. Observing the Observers: How Participants Contribute Data to iNaturalist and Implications for Biodiversity Science. BioScience 2021, 71, 1179–1188. [Google Scholar] [CrossRef]
- McMullin, R.T.; Allen, J.L. An Assessment of Data Accuracy and Best Practice Recommendations for Observations of Lichens and Other Taxonomically Difficult Taxa on iNaturalist. Botany 2022, 100, 491–497. [Google Scholar] [CrossRef]
- iNaturalist Research-Grade Observations. 2024. Available online: https://www.gbif.org/dataset/50c9509d-22c7-4a22-a47d-8c48425ef4a7 (accessed on 21 May 2025).
- Hochmair, H.H.; Scheffrahn, R.H.; Basille, M.; Boone, M. Evaluating the Data Quality of iNaturalist Termite Records. PLoS ONE 2020, 15, e0226534. [Google Scholar] [CrossRef]
- Ackland, S.J.; Richardson, D.M.; Robinson, T.B. A Method for Conveying Confidence in iNaturalist Observations: A Case Study Using Non-Native Marine Species. Ecol. Evol. 2024, 14, e70376. [Google Scholar] [CrossRef]
- Cristescu, M.E. Genetic Reconstructions of Invasion History. Mol. Ecol. 2015, 24, 2212–2225. [Google Scholar] [CrossRef]
- Zink, F.A.; Tembrock, L.R.; Timm, A.E.; Farris, R.E.; Perera, O.P.; Gilligan, T.M. A Droplet Digital PCR (ddPCR) Assay to Detect Helicoverpa armigera (Lepidoptera: Noctuidae) in Bulk Trap Samples. PLoS ONE 2017, 12, e0178704. [Google Scholar] [CrossRef]
- Hebert, P.D.; Floyd, R.; Jafarpour, S.; Prosser, S.W. Barcode 100 K Specimens: In a Single Nanopore Run. Mol. Ecol. Resour. 2023, 25, e14028. [Google Scholar] [CrossRef] [PubMed]
- Shashank, P.R.; Parker, B.M.; Rananaware, S.R.; Plotkin, D.; Couch, C.; Yang, L.G.; Nguyen, L.T.; Prasannakumar, N.R.; Braswell, W.E.; Jain, P.K.; et al. CRISPR-Based Diagnostics Detects Invasive Insect Pests. Mol. Ecol. Resour. 2023, 24, e13881. [Google Scholar] [CrossRef] [PubMed]
- Villari, C.; Mahaffee, W.F.; Mitchell, T.K.; Pedley, K.F.; Pieck, M.L.; Hand, F.P. Early Detection of Airborne Inoculum of Magnaporthe oryzae in Turfgrass Fields Using a Quantitative LAMP Assay. Plant Dis. 2017, 101, 170–177. [Google Scholar] [CrossRef]
- Larson, E.R.; Graham, B.M.; Achury, R.; Coon, J.J.; Daniels, M.K.; Gambrell, D.K.; Jonasen, K.L.; King, G.D.; LaRacuente, N.; Perrin-Stowe, T.I.; et al. From eDNA to Citizen Science: Emerging Tools for the Early Detection of Invasive Species. Front. Ecol. Environ. 2020, 18, 194–202. [Google Scholar] [CrossRef]
- Greenbaum, D.; Luscombe, N.M.; Jansen, R.; Qian, J.; Gerstein, M. Interrelating Different Types of Genomic Data, from Proteome to Secretome: ’Oming in on Function. Genome Res. 2001, 11, 1463–1468. [Google Scholar] [CrossRef]
- Arita, M.; Karsch-Mizrachi, I.; Cochrane, G. The International Nucleotide Sequence Database Collaboration. Nucleic Acids Res. 2020, 49, D121–D124. [Google Scholar] [CrossRef]
- Meiklejohn, K.A.; Damaso, N.; Robertson, J.M. Assessment of BOLD and GenBank—Their Accuracy and Reliability for the Identification of Biological Materials. PLoS ONE 2019, 14, e0217084. [Google Scholar] [CrossRef]
- Baena-Bejarano, N.; Reina, C.; Martínez-Revelo, D.E.; Medina, C.A.; Tovar, E.; Uribe-Soto, S.; Neita-Moreno, J.C.; Gonzalez, M.A. Taxonomic Identification Accuracy from BOLD and GenBank Databases Using over a Thousand Insect DNA Barcodes from Colombia. PLoS ONE 2023, 18, e0277379. [Google Scholar] [CrossRef]
- Clarke, J.L.; Cooper, L.D.; Poelchau, M.F.; Berardini, T.Z.; Elser, J.; Farmer, A.D.; Ficklin, S.; Kumari, S.; Laporte, M.-A.; Nelson, R.T.; et al. Data Sharing and Ontology Use among Agricultural Genetics, Genomics, and Breeding Databases and Resources of the Agbiodata Consortium. Database 2023, 2023, baad076. [Google Scholar] [CrossRef]
- Huang, C.; Lang, K.; Qian, W.; Wang, S.; Cao, X.; He, R.; Zhan, A.; Chen, M.; Yang, N.; Li, F. InvasionDB: A Genome and Gene Database of Invasive Alien Species. J. Integr. Agric. 2021, 20, 191–200. [Google Scholar] [CrossRef]
- Childers, A.K.; Geib, S.M.; Sim, S.B.; Poelchau, M.F.; Coates, B.S.; Simmonds, T.J.; Scully, E.D.; Smith, T.P.L.; Childers, C.P.; Corpuz, R.L.; et al. The USDA-ARS Ag100Pest Initiative: High-Quality Genome Assemblies for Agricultural Pest Arthropod Research. Insects 2021, 12, 626. [Google Scholar] [CrossRef] [PubMed]
- Giraldo-Calderón, G.I.; Harb, O.S.; Kelly, S.A.; Rund, S.S.; Roos, D.S.; McDowell, M.A. VectorBase.Org Updates: Bioinformatic Resources for Invertebrate Vectors of Human Pathogens and Related Organisms. Curr. Opin. Insect Sci. 2022, 50, 100860. [Google Scholar] [CrossRef] [PubMed]
- Legeai, F.; Shigenobu, S.; Gauthier, J.-P.; Colbourne, J.; Rispe, C.; Collin, O.; Richards, S.; Wilson, A.C.C.; Tagu, D. AphidBase: A Centralized Bioinformatic Resource for Annotation of the Pea Aphid Genome. Insect Mol. Biol. 2010, 19, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Yin, C.; Shen, G.; Guo, D.; Wang, S.; Ma, X.; Xiao, H.; Liu, J.; Zhang, Z.; Liu, Y.; Zhang, Y.; et al. InsectBase: A Resource for Insect Genomes and Transcriptomes. Nucleic Acids Res. 2016, 44, D801–D807. [Google Scholar] [CrossRef]
- Chen, W.; Hasegawa, D.K.; Kaur, N.; Kliot, A.; Pinheiro, P.V.; Luan, J.; Stensmyr, M.C.; Zheng, Y.; Liu, W.; Sun, H.; et al. The Draft Genome of Whitefly Bemisia Tabaci MEAM1, a Global Crop Pest, Provides Novel Insights into Virus Transmission, Host Adaptation, and Insecticide Resistance. BMC Biol. 2016, 14, 110. [Google Scholar] [CrossRef]
- Duvick, J.; Fu, A.; Muppirala, U.; Sabharwal, M.; Wilkerson, M.D.; Lawrence, C.J.; Lushbough, C.; Brendel, V. PlantGDB: A Resource for Comparative Plant Genomics. Nucleic Acids Res. 2008, 36, D959–D965. [Google Scholar] [CrossRef]
- Matheson, P.; McGaughran, A. Genomic Data Is Missing for Many Highly Invasive Species, Restricting Our Preparedness for Escalating Incursion Rates. Sci. Rep. 2022, 12, 13987. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-Km Spatial Resolution Climate Surfaces for Global Land Areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Austin, M.P.; Van Niel, K.P. Improving Species Distribution Models for Climate Change Studies: Variable Selection and Scale. J. Biogeogr. 2011, 38, 1–8. [Google Scholar] [CrossRef]
- Auclair, A.; Perez De Leon, A.; Teel, P.; Manoukis, N.; Messenger, M.; Bonilla, D. Novel Hurricane Hypothesis Predicts U.S Cattle Fever Tick Outbreaks. 2020. Available online: https://www.ars.usda.gov/research/publications/publication/?seqNo115=375172 (accessed on 21 May 2025).
- Borer, E.T.; Grace, J.B.; Harpole, W.S.; MacDougall, A.S.; Seabloom, E.W. A Decade of Insights into Grassland Ecosystem Responses to Global Environmental Change. Nat. Ecol. Evol. 2017, 1, 0118. [Google Scholar] [CrossRef]
- Li, D.; Record, S.; Sokol, E.R.; Bitters, M.E.; Chen, M.Y.; Chung, Y.A.; Helmus, M.R.; Jaimes, R.; Jansen, L.; Jarzyna, M.A.; et al. Standardized NEON Organismal Data for Biodiversity Research. Ecosphere 2022, 13, e4141. [Google Scholar] [CrossRef]
- Colwell, R.K.; Coddington, J.A. Estimating Terrestrial Biodiversity through Extrapolation. Philos. Trans. Biol. Sci. 1994, 345, 101–118. [Google Scholar]
- Jagadish, H.V.; Gehrke, J.; Labrinidis, A.; Papakonstantinou, Y.; Patel, J.M.; Ramakrishnan, R.; Shahabi, C. Big Data and Its Technical Challenges. Commun. ACM 2014, 57, 86–94. [Google Scholar] [CrossRef]
- Ficetola, G.F.; Thuiller, W.; Miaud, C. Prediction and Validation of the Potential Global Distribution of a Problematic Alien Invasive Species—The American Bullfrog. Divers. Distrib. 2007, 13, 476–485. [Google Scholar] [CrossRef]
- Rochlin, I.; Egizi, A.; Narvaez, Z.; Bonilla, D.L.; Gallagher, M.; Williams, G.M.; Rainey, T.; Price, D.C.; Fonseca, D.M. Microhabitat Modeling of the Invasive Asian Longhorned Tick (Haemaphysalis longicornis) in New Jersey, USA. Ticks Tick-Borne Dis. 2023, 14, 102126. [Google Scholar] [CrossRef]
- Lee, W.-H.; Song, J.-W.; Yoon, S.-H.; Jung, J.-M. Spatial Evaluation of Machine Learning-Based Species Distribution Models for Prediction of Invasive Ant Species Distribution. Appl. Sci. 2022, 12, 10260. [Google Scholar] [CrossRef]
- Streito, J.-C.; Chartois, M.; Pierre, É.; Dusoulier, F.; Armand, J.-M.; Gaudin, J.; Rossi, J.-P. Citizen Science and Niche Modeling to Track and Forecast the Expansion of the Brown Marmorated Stinkbug Halyomorpha halys (Stål, 1855). Sci. Rep. 2021, 11, 11421. [Google Scholar] [CrossRef]
- Fraser, D.; Kumar, S.; Aguilar, G. Mapping the Potential Global Range of the Brown Marmorated Stink Bug, Halyomorpha halys, with Particular Reference to New Zealand. Climate 2017, 5, 75. [Google Scholar] [CrossRef]
- Rubenstein, M.A.; Weiskopf, S.R.; Bertrand, R.; Carter, S.L.; Comte, L.; Eaton, M.J.; Johnson, C.G.; Lenoir, J.; Lynch, A.J.; Miller, B.W.; et al. Climate Change and the Global Redistribution of Biodiversity: Substantial Variation in Empirical Support for Expected Range Shifts. Environ. Evid. 2023, 12, 7. [Google Scholar] [CrossRef]
- Barbet-Massin, M.; Rome, Q.; Villemant, C.; Courchamp, F. Can Species Distribution Models Really Predict the Expansion of Invasive Species? PLoS ONE 2018, 13, e0193085. [Google Scholar] [CrossRef]
- Sillero, N.; Arenas-Castro, S.; Enriquez-Urzelai, U.; Vale, C.G.; Sousa-Guedes, D.; Martínez-Freiría, F.; Real, R.; Barbosa, A.M. Want to Model a Species Niche? A Step-by-Step Guideline on Correlative Ecological Niche Modelling. Ecol. Model. 2021, 456, 109671. [Google Scholar] [CrossRef]
- Jetz, W.; McGeoch, M.A.; Guralnick, R.; Ferrier, S.; Beck, J.; Costello, M.J.; Fernandez, M.; Geller, G.N.; Keil, P.; Merow, C.; et al. Essential Biodiversity Variables for Mapping and Monitoring Species Populations. Nat. Ecol. Evol. 2019, 3, 539–551. [Google Scholar] [CrossRef]
- Elith, J.; Franklin, J. Species Distribution Modelling. Encycl. Biodivers. 2013, 692–705. [Google Scholar] [CrossRef]
- Munro, H.L.; Montes, C.R.; Gandhi, K.J.K.; Poisson, M.A. A Comparison of Presence-Only Analytical Techniques and Their Application in Forest Pest Modeling. Ecol. Inform. 2022, 68, 101525. [Google Scholar] [CrossRef]
- Valavi, R.; Guillera-Arroita, G.; Lahoz-Monfort, J.J.; Elith, J. Predictive Performance of Presence-Only Species Distribution Models: A Benchmark Study with Reproducible Code. Ecol. Monogr. 2022, 92, e01486. [Google Scholar] [CrossRef]
- Meineke, E.K.; Daru, B.H. Bias Assessments to Expand Research Harnessing Biological Collections. Trends Ecol. Evol. 2021, 36, 1071–1082. [Google Scholar] [CrossRef]
- Araújo, M.B.; Anderson, R.P.; Márcia Barbosa, A.; Beale, C.M.; Dormann, C.F.; Early, R.; Garcia, R.A.; Guisan, A.; Maiorano, L.; Naimi, B.; et al. Standards for Distribution Models in Biodiversity Assessments. Sci. Adv. 2019, 5, eaat4858. [Google Scholar] [CrossRef]
- Brown, J.L. SDMtoolbox: A Python-Based GIS Toolkit for Landscape Genetic, Biogeographic and Species Distribution Model Analyses. Methods Ecol. Evol. 2014, 5, 694–700. [Google Scholar] [CrossRef]
- Capinha, C.; Anastácio, P. Assessing the Environmental Requirements of Invaders Using Ensembles of Distribution Models. Divers. Distrib. 2011, 17, 13–24. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.J.; Silander, J.A., Jr. A Practical Guide to MaxEnt for Modeling Species’ Distributions: What It Does, and Why Inputs and Settings Matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Muscatello, A.; Elith, J.; Kujala, H. How Decisions about Fitting Species Distribution Models Affect Conservation Outcomes. Conserv. Biol. 2021, 35, 1309–1320. [Google Scholar] [CrossRef] [PubMed]
- Araújo, M.B.; New, M. Ensemble Forecasting of Species Distributions. Trends Ecol. Evol. 2007, 22, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Kou, D.-M.; Sun, Y.; Long, L.-G.; Wang, J.-G.; Wu, J.-W.; Long, T.; Li, W.-J. Predicting the Suitable Habitat of the Invasive Alien Plant Lactuca serriola Using Biomod2 Model with ArcGIS. Environ. Res. Commun. 2025, 7, 045029. [Google Scholar] [CrossRef]
- Hao, T.; Elith, J.; Guillera-Arroita, G.; Lahoz-Monfort, J.J. A Review of Evidence about Use and Performance of Species Distribution Modelling Ensembles like BIOMOD. Divers. Distrib. 2019, 25, 839–852. [Google Scholar] [CrossRef]
- Hao, T.; Elith, J.; Lahoz-Monfort, J.J.; Guillera-Arroita, G. Testing Whether Ensemble Modelling Is Advantageous for Maximising Predictive Performance of Species Distribution Models. Ecography 2020, 43, 549–558. [Google Scholar] [CrossRef]
- Carboneras, C.; Genovesi, P.; Vilà, M.; Blackburn, T.M.; Carrete, M.; Clavero, M.; D’hondt, B.; Orueta, J.F.; Gallardo, B.; Geraldes, P.; et al. A Prioritised List of Invasive Alien Species to Assist the Effective Implementation of EU Legislation. J. Appl. Ecol. 2018, 55, 539–547. [Google Scholar] [CrossRef]
- Roy, H.E.; Bacher, S.; Essl, F.; Adriaens, T.; Aldridge, D.C.; Bishop, J.D.D.; Blackburn, T.M.; Branquart, E.; Brodie, J.; Carboneras, C.; et al. Developing a List of Invasive Alien Species Likely to Threaten Biodiversity and Ecosystems in the European Union. Glob. Change Biol. 2019, 25, 1032–1048. [Google Scholar] [CrossRef]
- McGeoch, M.A.; Genovesi, P.; Bellingham, P.J.; Costello, M.J.; McGrannachan, C.; Sheppard, A. Prioritizing Species, Pathways, and Sites to Achieve Conservation Targets for Biological Invasion. Biol. Invasions 2016, 18, 299–314. [Google Scholar] [CrossRef]
- McCullough, D.G.; Work, T.T.; Cavey, J.F.; Liebhold, A.M.; Marshall, D. Interceptions of Nonindigenous Plant Pests at US Ports of Entry and Border Crossings over a 17-Year Period. Biol. Invasions 2006, 8, 611–630. [Google Scholar] [CrossRef]
- Venette, R.C.; Hutchison, W.D. Invasive Insect Species: Global Challenges, Strategies & Opportunities. Front. Insect Sci. 2021, 1, 650520. [Google Scholar] [CrossRef]
- Meurisse, N.; Rassati, D.; Hurley, B.P.; Brockerhoff, E.G.; Haack, R.A. Common Pathways by Which Non-Native Forest Insects Move Internationally and Domestically. J. Pest. Sci. 2019, 92, 13–27. [Google Scholar] [CrossRef]
- Tobin, P.C.; Kean, J.M.; Suckling, D.M.; McCullough, D.G.; Herms, D.A.; Stringer, L.D. Determinants of Successful Arthropod Eradication Programs. Biol. Invasions 2014, 16, 401–414. [Google Scholar] [CrossRef]
- Pluess, T.; Cannon, R.; Jarošík, V.; Pergl, J.; Pyšek, P.; Bacher, S. When Are Eradication Campaigns Successful? A Test of Common Assumptions. Biol. Invasions 2012, 14, 1365–1378. [Google Scholar] [CrossRef]
- Simberloff, D. The Role of Propagule Pressure in Biological Invasions. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 81–102. [Google Scholar] [CrossRef]
- Saccaggi, D.L.; Wilson, J.R.U.; Robinson, A.P.; Terblanche, J.S. Arthropods on Imported Plant Products: Volumes Predict General Trends While Contextual Details Enhance Predictive Power. Ecol. Appl. 2022, 32, e2554. [Google Scholar] [CrossRef]
- Santosuosso, U.; Cini, A.; Papini, A. Tracing Outliers in the Dataset of Drosophila Suzukii Records with the Isolation Forest Method. J. Big Data 2020, 7, 14. [Google Scholar] [CrossRef]
- Wang, S.; Wang, C.; Wang, S.; Ma, L. Big Data Analysis for Evaluating Bioinvasion Risk. BMC Bioinform. 2018, 19, 287. [Google Scholar] [CrossRef]
- Carnegie, A.J.; Eslick, H.; Barber, P.; Nagel, M.; Stone, C. Airborne Multispectral Imagery and Deep Learning for Biosecurity Surveillance of Invasive Forest Pests in Urban Landscapes. Urban For. Urban Green. 2023, 81, 127859. [Google Scholar] [CrossRef]
- Latombe, G.; Pyšek, P.; Jeschke, J.M.; Blackburn, T.M.; Bacher, S.; Capinha, C.; Costello, M.J.; Fernández, M.; Gregory, R.D.; Hobern, D.; et al. A Vision for Global Monitoring of Biological Invasions. Biol. Conserv. 2017, 213, 295–308. [Google Scholar] [CrossRef]
- Turner, R.M.; Brockerhoff, E.G.; Bertelsmeier, C.; Blake, R.E.; Caton, B.; James, A.; MacLeod, A.; Nahrung, H.F.; Pawson, S.M.; Plank, M.J.; et al. Worldwide Border Interceptions Provide a Window into Human-Mediated Global Insect Movement. Ecol. Appl. 2021, 31, e02412. [Google Scholar] [CrossRef]
- Peters, K.; Sink, K.; Robinson, T.B. Aliens Cruising in: Explaining Alien Fouling Macro-Invertebrate Species Numbers on Recreational Yachts. Ocean Coast. Manag. 2019, 182, 104986. [Google Scholar] [CrossRef]
- Jaspers, C.; Ehrlich, M.; Pujolar, J.M.; Künzel, S.; Bayer, T.; Limborg, M.T.; Lombard, F.; Browne, W.E.; Stefanova, K.; Reusch, T.B.H. Invasion Genomics Uncover Contrasting Scenarios of Genetic Diversity in a Widespread Marine Invader. Proc. Natl. Acad. Sci. USA 2021, 118, e2116211118. [Google Scholar] [CrossRef] [PubMed]
- Browett, S.S.; O’Meara, D.B.; McDevitt, A.D. Genetic Tools in the Management of Invasive Mammals: Recent Trends and Future Perspectives. Mammal. Rev. 2020, 50, 200–210. [Google Scholar] [CrossRef]
- North, H.L.; McGaughran, A.; Jiggins, C.D. Insights into Invasive Species from Whole-Genome Resequencing. Mol. Ecol. 2021, 30, 6289–6308. [Google Scholar] [CrossRef]
- Zepeda-Paulo, F.A.; Simon, J.-C.; Ramírez, C.C.; Fuentes-Contreras, E.; Margaritopoulos, J.T.; Wilson, A.C.C.; Sorenson, C.E.; Briones, L.M.; Azevedo, R.; Ohashi, D.V.; et al. The Invasion Route for an Insect Pest Species: The Tobacco Aphid in the New World. Mol. Ecol. 2010, 19, 4738–4752. [Google Scholar] [CrossRef]
- Rius, M.; Bourne, S.; Hornsby, H.G.; Chapman, M.A. Applications of Next-Generation Sequencing to the Study of Biological Invasions. Curr. Zool. 2015, 61, 488–504. [Google Scholar] [CrossRef]
- Pereira, H.; Ferrier, S.; Walters, M.; Geller, G.; Scholes, R.; Bruford, M.; Brummitt, N.; Butchart, S.; Cardoso, A.; Coops, N.; et al. Essential Biodiversity Variables. Science 2013, 339, 277–278. [Google Scholar] [CrossRef]
- Liu, C.; Comte, L.; Olden, J.D. Heads You Win, Tails You Lose: Life-History Traits Predict Invasion and Extinction Risk of the World’s Freshwater Fishes. Aquat. Conserv. Mar. Freshw. Ecosyst. 2017, 27, 773–779. [Google Scholar] [CrossRef]
- Capellini, I.; Baker, J.; Allen, W.L.; Street, S.E.; Venditti, C. The Role of Life History Traits in Mammalian Invasion Success. Ecol. Lett. 2015, 18, 1099–1107. [Google Scholar] [CrossRef]
- Fournier, A.; Penone, C.; Pennino, M.G.; Courchamp, F. Predicting Future Invaders and Future Invasions. Proc. Natl. Acad. Sci. USA 2019, 116, 7905–7910. [Google Scholar] [CrossRef]
- Philibert, A.; Desprez-Loustau, M.-L.; Fabre, B.; Frey, P.; Halkett, F.; Husson, C.; Lung-Escarmant, B.; Marçais, B.; Robin, C.; Vacher, C.; et al. Predicting Invasion Success of Forest Pathogenic Fungi from Species Traits. J. Appl. Ecol. 2011, 48, 1381–1390. [Google Scholar] [CrossRef]
- Desprez-Loustau, M.-L.; Robin, C.; Buee, M.; Courtecuisse, R.; Garbaye, J.; Suffert, F.; Sache, I.; Rizzo, D. The Fungal Dimension of Biological Invasions. Trends Ecol. Evol. 2007, 22, 472–480. [Google Scholar] [CrossRef] [PubMed]
- Jarnevich, C.S.; Young, N.E.; Talbert, M.; Talbert, C. Forecasting an Invasive Species’ Distribution with Global Distribution Data, Local Data, and Physiological Information. Ecosphere 2018, 9, e02279. [Google Scholar] [CrossRef]
- Agosti, D.; Catapano, T.; Sautter, G.; Egloff, W. The Plazi Workflow: The PDF Prison Break for Biodiversity Data. Biodivers. Inf. Sci. Stand. 2019, 3, e37046. [Google Scholar] [CrossRef]
- Schulz, A.N.; Mech, A.M.; Ayres, M.P.; Gandhi, K.J.K.; Havill, N.P.; Herms, D.A.; Hoover, A.M.; Hufbauer, R.A.; Liebhold, A.M.; Marsico, T.D.; et al. Predicting Non-Native Insect Impact: Focusing on the Trees to See the Forest. Biol. Invasions 2021, 23, 3921–3936. [Google Scholar] [CrossRef]
- Medzihorský, V.; Trombik, J.; Mally, R.; Turčáni, M.; Liebhold, A.M. Insect Invasions Track a Tree Invasion: Global Distribution of Black Locust Herbivores. J. Biogeogr. 2023, 50, 1285–1298. [Google Scholar] [CrossRef]
- Lieurance, D.; Canavan, S.; Behringer, D.C.; Kendig, A.E.; Minteer, C.R.; Reisinger, L.S.; Romagosa, C.M.; Flory, S.L.; Lockwood, J.L.; Anderson, P.J.; et al. Identifying Invasive Species Threats, Pathways, and Impacts to Improve Biosecurity. Ecosphere 2023, 14, e4711. [Google Scholar] [CrossRef]
- Kaushik, P.; Pati, P.K.; Khan, M.L.; Khare, P.K. Plant Functional Traits Best Explain Invasive Species’ Performance within a Dynamic Ecosystem—A Review. Trees For. People 2022, 8, 100260. [Google Scholar] [CrossRef]
- Miles, C.M.; Wayne, M. Quantitative Trait Locus (QTL) Analysis. Nat. Educ. 2008, 1, 208. [Google Scholar]
- Thurber, C.S.; Jia, M.H.; Jia, Y.; Caicedo, A.L. Similar Traits, Different Genes? Examining Convergent Evolution in Related Weedy Rice Populations. Mol. Ecol. 2013, 22, 685–698. [Google Scholar] [CrossRef]
- Widstrom, N.W.; Butron, A.; Guo, B.Z.; Wilson, D.M.; Snook, M.E.; Cleveland, T.E.; Lynch, R.E. Control of Preharvest Aflatoxin Contamination in Maize by Pyramiding QTL Involved in Resistance to Ear-Feeding Insects and Invasion by Aspergillus spp. Eur. J. Agron. 2003, 19, 563–572. [Google Scholar] [CrossRef]
- Olazcuaga, L.; Loiseau, A.; Parrinello, H.; Paris, M.; Fraimout, A.; Guedot, C.; Diepenbrock, L.M.; Kenis, M.; Zhang, J.; Chen, X.; et al. A Whole-Genome Scan for Association with Invasion Success in the Fruit Fly Drosophila suzukii Using Contrasts of Allele Frequencies Corrected for Population Structure. Mol. Biol. Evol. 2020, 37, 2369–2385. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, S.; De Meyer, M.; Liao, Z.; Zhao, Y.; Virgilio, M.; Feng, S.; Qin, Y.; Singh, S.; Wee, S.L.; et al. Genomes of the Cosmopolitan Fruit Pest Bactrocera dorsalis (Diptera: Tephritidae) Reveal Its Global Invasion History and Thermal Adaptation. J. Adv. Res. 2022, 53, 61–74. [Google Scholar] [CrossRef]
- You, M.; Ke, F.; You, S.; Wu, Z.; Liu, Q.; He, W.; Baxter, S.W.; Yuchi, Z.; Vasseur, L.; Gurr, G.M.; et al. Variation among 532 Genomes Unveils the Origin and Evolutionary History of a Global Insect Herbivore. Nat. Commun. 2020, 11, 2321. [Google Scholar] [CrossRef]
- Li, H.; Wang, J.; Peng, Y.; Guo, C.; Qu, W.; Yang, N.; Zhu, Y.; Jeong, I.; Li, X.; Ghanim, M.; et al. Invasion Genomics Uncover Complex Introduction Patterns of the Globally Invasive Whitefly, Bemisia tabaci MED. Divers. Distrib. 2023, 29, 1172–1189. [Google Scholar] [CrossRef]
- Uden, D.R.; Mech, A.M.; Havill, N.P.; Schulz, A.N.; Ayres, M.P.; Herms, D.A.; Hoover, A.M.; Gandhi, K.J.K.; Hufbauer, R.A.; Liebhold, A.M.; et al. Phylogenetic Risk Assessment Is Robust for Forecasting the Impact of Non-Native Insects on North American Trees. Ecol. Appl. 2023, 33, e2761. [Google Scholar] [CrossRef]
- Smith, S.D.; Pennell, M.W.; Dunn, C.W.; Edwards, S.V. Phylogenetics Is the New Genetics (for Most of Biodiversity). Trends Ecol. Evol. 2020, 35, 415–425. [Google Scholar] [CrossRef]
- Gemmell, P.; Sackton, T.B.; Edwards, S.V.; Liu, J.S. A Phylogenetic Method Linking Nucleotide Substitution Rates to Rates of Continuous Trait Evolution. PLoS Comput. Biol. 2024, 20, e1011995. [Google Scholar] [CrossRef]
- Estoup, A.; Ravigné, V.; Hufbauer, R.; Vitalis, R.; Gautier, M.; Facon, B. Is There a Genetic Paradox of Biological Invasion? Annu. Rev. Ecol. Evol. Syst. 2016, 47, 51–72. [Google Scholar] [CrossRef]
- Wang, J.; Gaughan, S.; Lamer, J.T.; Deng, C.; Hu, W.; Wachholtz, M.; Qin, S.; Nie, H.; Liao, X.; Ling, Q.; et al. Resolving the Genetic Paradox of Invasions: Preadapted Genomes and Postintroduction Hybridization of Bigheaded Carps in the Mississippi River Basin. Evol. Appl. 2020, 13, 263–277. [Google Scholar] [CrossRef]
- Sax, D.F.; Brown, J.H. The Paradox of Invasion. Glob. Ecol. Biogeogr. 2000, 9, 363–371. [Google Scholar] [CrossRef]
- Chown, S.L.; Hodgins, K.A.; Griffin, P.C.; Oakeshott, J.G.; Byrne, M.; Hoffmann, A.A. Biological Invasions, Climate Change and Genomics. Evol. Appl. 2015, 8, 23–46. [Google Scholar] [CrossRef] [PubMed]
- Hessenauer, P.; Fijarczyk, A.; Martin, H.; Prunier, J.; Charron, G.; Chapuis, J.; Bernier, L.; Tanguay, P.; Hamelin, R.C.; Landry, C.R. Hybridization and Introgression Drive Genome Evolution of Dutch Elm Disease Pathogens. Nat. Ecol. Evol. 2020, 4, 626–638. [Google Scholar] [CrossRef]
- Wu, N.; Zhang, S.; Li, X.; Cao, Y.; Liu, X.; Wang, Q.; Liu, Q.; Liu, H.; Hu, X.; Zhou, X.J.; et al. Fall Webworm Genomes Yield Insights into Rapid Adaptation of Invasive Species. Nat. Ecol. Evol. 2019, 3, 105–115. [Google Scholar] [CrossRef]
- Bock, D.G.; Caseys, C.; Cousens, R.D.; Hahn, M.A.; Heredia, S.M.; Hübner, S.; Turner, K.G.; Whitney, K.D.; Rieseberg, L.H. What We Still Don’t Know about Invasion Genetics. Mol. Ecol. 2015, 24, 2277–2297. [Google Scholar] [CrossRef]
- Marin, P.; Genitoni, J.; Barloy, D.; Maury, S.; Gibert, P.; Ghalambor, C.K.; Vieira, C. Biological Invasion: The Influence of the Hidden Side of the (Epi)Genome. Funct. Ecol. 2020, 34, 385–400. [Google Scholar] [CrossRef]
- LaRue, E.A.; Hardiman, B.S.; Elliott, J.M.; Fei, S. Structural Diversity as a Predictor of Ecosystem Function. Environ. Res. Lett. 2019, 14, 114011. [Google Scholar] [CrossRef]
- Gholizadeh, H.; Rakotoarivony, M.N.A.; Hassani, K.; Johnson, K.G.; Hamilton, R.G.; Fuhlendorf, S.D.; Schneider, F.D.; Bachelot, B. Advancing Our Understanding of Plant Diversity-Biological Invasion Relationships Using Imaging Spectroscopy. Remote Sens. Environ. 2024, 304, 114028. [Google Scholar] [CrossRef]
- Gill, N.S.; Mahood, A.L.; Meier, C.L.; Muthukrishnan, R.; Nagy, R.C.; Stricker, E.; Duffy, K.A.; Petri, L.; Morisette, J.T. Six Central Questions about Biological Invasions to Which NEON Data Science Is Poised to Contribute. Ecosphere 2021, 12, e03728. [Google Scholar] [CrossRef]
- Knapp, A.K.; Smith, M.D.; Hobbie, S.E.; Collins, S.L.; Fahey, T.J.; Hansen, G.J.A.; Landis, D.A.; La Pierre, K.J.; Melillo, J.M.; Seastedt, T.R.; et al. Past, Present, and Future Roles of Long-Term Experiments in the LTER Network. BioScience 2012, 62, 377–389. [Google Scholar] [CrossRef]
- Haymer, D. Genetics and Insect Pest Management in Agriculture. CABI Rev. 2015, 2015, 1–8. [Google Scholar] [CrossRef]
- Moro, D.; Byrne, M.; Kennedy, M.; Campbell, S.; Tizard, M. Identifying Knowledge Gaps for Gene Drive Research to Control Invasive Animal Species: The next CRISPR Step. Glob. Ecol. Conserv. 2018, 13, e00363. [Google Scholar] [CrossRef]
- Medina, R.F. Gene Drives and the Management of Agricultural Pests. J. Responsible Innov. 2018, 5, S255–S262. [Google Scholar] [CrossRef]
- Whitten, M.M.A.; Facey, P.D.; Del Sol, R.; Fernández-Martínez, L.T.; Evans, M.C.; Mitchell, J.J.; Bodger, O.G.; Dyson, P.J. Symbiont-Mediated RNA Interference in Insects. Proc. R. Soc. B Biol. Sci. 2016, 283, 20160042. [Google Scholar] [CrossRef]
- Pampolini, F.; Rieske, L.K. Emerald Ash Borer Specific Gene Silencing Has No Effect on Non-Target Organisms. Front. Agron. 2020, 2, 608827. [Google Scholar] [CrossRef]
- Sjodin, B.M.F.; Irvine, R.L.; Russello, M.A. RapidRat: Development, Validation and Application of a Genotyping-by-Sequencing Panel for Rapid Biosecurity and Invasive Species Management. PLoS ONE 2020, 15, e0234694. [Google Scholar] [CrossRef]
- Lampert, A. Optimizing Strategies for Slowing the Spread of Invasive Species. PLoS Comput. Biol. 2024, 20, e1011996. [Google Scholar] [CrossRef]
- Taylor, B.A.; Tembrock, L.R.; Sankovitz, M.; Wilson, T.M.; Looney, C.; Takahashi, J.; Gilligan, T.M.; Smith-Pardo, A.H.; Harpur, B.A. Population Genomics of the Invasive Northern Giant Hornet Vespa mandarinia in North America and across Its Native Range. Sci. Rep. 2024, 14, 10803. [Google Scholar] [CrossRef]
- Baker, C.M.; Bode, M. Recent Advances of Quantitative Modeling to Support Invasive Species Eradication on Islands. Conserv. Sci. Pract. 2021, 3, e246. [Google Scholar] [CrossRef]
- Manoukis, N.C.; Hall, B.; Geib, S.M. A Computer Model of Insect Traps in a Landscape. Sci. Rep. 2014, 4, 7015. [Google Scholar] [CrossRef]
- Ivaskovic, P.; Ainseba, B.; Nicolas, Y.; Toupance, T.; Tardy, P.; Thiéry, D. Sensing of Airborne Infochemicals for Green Pest Management: What Is the Challenge? ACS Sens. 2021, 6, 3824–3840. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.M.; Jones, S.; Petrasova, A.; Petras, V.; Gaydos, D.; Skrip, M.M.; Takeuchi, Y.; Bigsby, K.; Meentemeyer, R.K. Iteratively Forecasting Biological Invasions with PoPS and a Little Help from Our Friends. Front. Ecol. Environ. 2021, 19, 411–418. [Google Scholar] [CrossRef] [PubMed]
- van Evert, F.K.; Fountas, S.; Jakovetic, D.; Crnojevic, V.; Travlos, I.; Kempenaar, C. Big Data for Weed Control and Crop Protection. Weed Res. 2017, 57, 218–233. [Google Scholar] [CrossRef]
- Kim, J.; Huebner, C.D.; Reardon, R.; Park, Y.-L. Spatially Targeted Biological Control of Mile-a-Minute Weed Using Rhinoncomimus latipes (Coleoptera: Curculionidae) and an Unmanned Aircraft System. J. Econ. Entomol. 2021, 114, 1889–1895. [Google Scholar] [CrossRef]
- Maran, S.P.M. Application of Artificial Intelligence (AI) Tools in Integrated Pest Management (IPM)—An Insect—Plant Interaction Perspective. Acta Sci. Agric. 2022, 6, 11–16. [Google Scholar] [CrossRef]
- Biasazin, T.D.; Larsson Herrera, S.; Kimbokota, F.; Dekker, T. Translating Olfactomes into Attractants: Shared Volatiles Provide Attractive Bridges for Polyphagy in Fruit Flies. Ecol. Lett. 2019, 22, 108–118. [Google Scholar] [CrossRef]
- Hampton, S.E.; Strasser, C.A.; Tewksbury, J.J.; Gram, W.K.; Budden, A.E.; Batcheller, A.L.; Duke, C.S.; Porter, J.H. Big Data and the Future of Ecology. Front. Ecol. Environ. 2013, 11, 156–162. [Google Scholar] [CrossRef]
- Farley, S.S.; Dawson, A.; Goring, S.J.; Williams, J.W. Situating Ecology as a Big-Data Science: Current Advances, Challenges, and Solutions. BioScience 2018, 68, 563–576. [Google Scholar] [CrossRef]
- Giovos, I.; Kleitou, P.; Poursanidis, D.; Batjakas, I.; Bernardi, G.; Crocetta, F.; Doumpas, N.; Kalogirou, S.; Kampouris, T.E.; Keramidas, I.; et al. Citizen-Science for Monitoring Marine Invasions and Stimulating Public Engagement: A Case Project from the Eastern Mediterranean. Biol Invasions 2019, 21, 3707–3721. [Google Scholar] [CrossRef]
- Kousteni, V.; Tsiamis, K.; Gervasini, E.; Zenetos, A.; Karachle, P.K.; Cardoso, A.C. Citizen Scientists Contributing to Alien Species Detection: The Case of Fishes and Mollusks in European Marine Waters. Ecosphere 2022, 13, e03875. [Google Scholar] [CrossRef]
- Ji, Y.; Ashton, L.; Pedley, S.M.; Edwards, D.P.; Tang, Y.; Nakamura, A.; Kitching, R.; Dolman, P.M.; Woodcock, P.; Edwards, F.A.; et al. Reliable, Verifiable and Efficient Monitoring of Biodiversity via Metabarcoding. Ecol. Lett. 2013, 16, 1245–1257. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Chen, L.; Zhang, X.; Chen, E. Improved Prototypical Network Model for Forest Species Classification in Complex Stand. Remote Sens. 2020, 12, 3839. [Google Scholar] [CrossRef]
- Ocean Biodiversity Information System; Intergovernmental Oceanographic Commission of UNESCO. The OBIS Manual. 2024. Available online: https://obis.org (accessed on 21 May 2025).
- ISO 19116:2019(En); Geographic Information—Positioning Services. ISO: Geneva, Switzerland, 2019. Available online: https://www.iso.org/obp/ui/en/#iso:std:iso:19116:ed-2:v1:en (accessed on 5 July 2024).
- ISO 19157-1:2023(En); Geographic Information—Data Quality—Part 1: General Requirements. ISO: Geneva, Switzerland, 2023. Available online: https://www.iso.org/obp/ui/en/#iso:std:iso:19157:-1:ed-1:v1:en (accessed on 5 July 2024).
- Zurell, D.; Franklin, J.; König, C.; Bouchet, P.J.; Dormann, C.F.; Elith, J.; Fandos, G.; Feng, X.; Guillera-Arroita, G.; Guisan, A.; et al. A Standard Protocol for Reporting Species Distribution Models. Ecography 2020, 43, 1261–1277. [Google Scholar] [CrossRef]
- Leray, M.; Knowlton, N.; Ho, S.-L.; Nguyen, B.N.; Machida, R.J. GenBank Is a Reliable Resource for 21st Century Biodiversity Research. Proc. Natl. Acad. Sci. USA 2019, 116, 22651–22656. [Google Scholar] [CrossRef]
- Maldonado, C.; Molina, C.I.; Zizka, A.; Persson, C.; Taylor, C.M.; Albán, J.; Chilquillo, E.; Rønsted, N.; Antonelli, A. Estimating Species Diversity and Distribution in the Era of Big Data: To What Extent Can We Trust Public Databases? Glob. Ecol. Biogeogr. 2015, 24, 973–984. [Google Scholar] [CrossRef]
- Zizka, A.; Silvestro, D.; Andermann, T.; Azevedo, J.; Duarte Ritter, C.; Edler, D.; Farooq, H.; Herdean, A.; Ariza, M.; Scharn, R.; et al. CoordinateCleaner: Standardized Cleaning of Occurrence Records from Biological Collection Databases. Methods Ecol. Evol. 2019, 10, 744–751. [Google Scholar] [CrossRef]
- Beck, J.; Böller, M.; Erhardt, A.; Schwanghart, W. Spatial Bias in the GBIF Database and Its Effect on Modeling Species’ Geographic Distributions. Ecol. Inform. 2014, 19, 10–15. [Google Scholar] [CrossRef]
- García-Roselló, E.; González-Dacosta, J.; Lobo, J.M. The Biased Distribution of Existing Information on Biodiversity Hinders Its Use in Conservation, and We Need an Integrative Approach to Act Urgently. Biol. Conserv. 2023, 283, 110118. [Google Scholar] [CrossRef]
- Wilkinson, M.D.; Dumontier, M.; Aalbersberg, I.J.; Appleton, G.; Axton, M.; Baak, A.; Blomberg, N.; Boiten, J.-W.; da Silva Santos, L.B.; Bourne, P.E.; et al. The FAIR Guiding Principles for Scientific Data Management and Stewardship. Sci. Data 2016, 3, 160018. [Google Scholar] [CrossRef]
- Shin, D.-H.; Choi, M.J. Ecological Views of Big Data: Perspectives and Issues. Telemat. Inform. 2015, 32, 311–320. [Google Scholar] [CrossRef]
- De Bona, S.D.; Barringer, L.; Kurtz, P.; Losiewicz, J.; Parra, G.R.; Helmus, M.R. Lydemapr: An R Package to Track the Spread of the Invasive Spotted Lanternfly (Lycorma delicatula, White 1845) (Hemiptera, Fulgoridae) in the United States. NeoBiota 2023, 86, 151–168. [Google Scholar] [CrossRef]
- Bronson, K.; Knezevic, I. Big Data in Food and Agriculture. Big Data Soc. 2016, 3, 2053951716648174. [Google Scholar] [CrossRef]
- Hackfort, S.; Marquis, S.; Bronson, K. Harvesting Value: Corporate Strategies of Data Assetization in Agriculture and Their Socio-Ecological Implications. Big Data Soc. 2024, 11, 20539517241234279. [Google Scholar] [CrossRef]
- Carbonell, I.M. The Ethics of Big Data in Big Agriculture. Internet Policy Rev. 2016, 5, 1–13. [Google Scholar] [CrossRef]
- Folger, P. The Geospatial Data Act of 2018; FGDC: Reston, VA, USA, 2018.
- Pagad, S.; Genovesi, P.; Carnevali, L.; Schigel, D.; McGeoch, M.A. Introducing the Global Register of Introduced and Invasive Species. Sci. Data 2018, 5, 170202. [Google Scholar] [CrossRef]
- Simpson, A.; Jarnevich, C.; Madsen, J.; Westbrooks, R.; Fournier, C.; Mehrhoff, L.; Browne, M.; Graham, J.; Sellers, E. Invasive Species Information Networks: Collaboration at Multiple Scales for Prevention, Early Detection, and Rapid Response to Invasive Alien Species. Biodiversity 2009, 10, 5–13. [Google Scholar] [CrossRef]
- Fegraus, E.H.; Andelman, S.; Jones, M.B.; Schildhauer, M. Maximizing the Value of Ecological Data with Structured Metadata: An Introduction to Ecological Metadata Language (EML) and Principles for Metadata Creation. Bull. Ecol. Soc. Am. 2005, 86, 158–168. [Google Scholar] [CrossRef]
- Gunia, B.; Sandusky, R.J. Designing Metadata for Long-Term Data Preservation: DataONE Case Study. Proc. Am. Soc. Inf. Sci. Technol. 2010, 47, 1–2. [Google Scholar] [CrossRef]
- Wieczorek, J.; Bloom, D.; Guralnick, R.; Blum, S.; Döring, M.; Giovanni, R.; Robertson, T.; Vieglais, D. Darwin Core: An Evolving Community-Developed Biodiversity Data Standard. PLoS ONE 2012, 7, e29715. [Google Scholar] [CrossRef]
- Simmons, M.; Tucker, A.; Chadderton, W.L.; Jerde, C.L.; Mahon, A.R. Active and Passive Environmental DNA Surveillance of Aquatic Invasive Species. Can. J. Fish. Aquat. Sci. 2015, 73, 76–83. [Google Scholar] [CrossRef]
- Brown, N.; Pérez-Sierra, A.; Crow, P.; Parnell, S. The Role of Passive Surveillance and Citizen Science in Plant Health. CABI Agric. Biosci. 2020, 1, 17. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, K.F.; Ball, S.L. DNA Barcodes for Biosecurity: Invasive Species Identification. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 1813–1823. [Google Scholar] [CrossRef] [PubMed]
- Taberlet, P.; Coissac, E.; Hajibabaei, M.; Rieseberg, L.H. Environmental DNA. Mol. Ecol. 2012, 21, 1789–1793. [Google Scholar] [CrossRef]
- Estoup, A.; Guillemaud, T. Reconstructing Routes of Invasion Using Genetic Data: Why, How and so What? Mol. Ecol. 2010, 19, 4113–4130. [Google Scholar] [CrossRef]
- Perry, G.L.W.; Seidl, R.; Bellvé, A.M.; Rammer, W. An Outlook for Deep Learning in Ecosystem Science. Ecosystems 2022, 25, 1700–1718. [Google Scholar] [CrossRef]
- Deiner, K.; Bik, H.M.; Mächler, E.; Seymour, M.; Lacoursière-Roussel, A.; Altermatt, F.; Creer, S.; Bista, I.; Lodge, D.M.; de Vere, N.; et al. Environmental DNA Metabarcoding: Transforming How We Survey Animal and Plant Communities. Mol. Ecol. 2017, 26, 5872–5895. [Google Scholar] [CrossRef]
- Data Quality Requirements: Occurrence Datasets. Available online: https://www.gbif.org/data-quality-requirements-occurrences (accessed on 15 July 2024).
- Potamitis, I.; Rigakis, I.; Fysarakis, K. The Electronic McPhail Trap. Sensors 2014, 14, 22285–22299. [Google Scholar] [CrossRef]
- Barker, N.; Cumming, S.; Darveau, M. Models to Predict the Distribution and Abundance of Breeding Ducks in Canada. Avian Conserv. Ecol. 2014, 9, 7. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R. Species Distribution Models: Ecological Explanation and Prediction Across Space and Time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Clement, R.A.; Lee, H.; Manoukis, N.C.; Pacheco, Y.M.; Ross, F.; Sisterson, M.S.; Owen, C.L. Addressing Biological Invasions in Agriculture with Big Data in an Informatics Age. Agriculture 2025, 15, 1157. https://doi.org/10.3390/agriculture15111157
Clement RA, Lee H, Manoukis NC, Pacheco YM, Ross F, Sisterson MS, Owen CL. Addressing Biological Invasions in Agriculture with Big Data in an Informatics Age. Agriculture. 2025; 15(11):1157. https://doi.org/10.3390/agriculture15111157
Chicago/Turabian StyleClement, Rebecca A., Hyoseok Lee, Nicholas C. Manoukis, Yelena M. Pacheco, Fallon Ross, Mark S. Sisterson, and Christopher L. Owen. 2025. "Addressing Biological Invasions in Agriculture with Big Data in an Informatics Age" Agriculture 15, no. 11: 1157. https://doi.org/10.3390/agriculture15111157
APA StyleClement, R. A., Lee, H., Manoukis, N. C., Pacheco, Y. M., Ross, F., Sisterson, M. S., & Owen, C. L. (2025). Addressing Biological Invasions in Agriculture with Big Data in an Informatics Age. Agriculture, 15(11), 1157. https://doi.org/10.3390/agriculture15111157