
Biomass, Physiological, and Antioxidant Activity Responses of Wheat Plants After Selenium Foliar Spray Under Water Deficit

 ,
,  , ,
, ,  ,
,  ,
,  , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
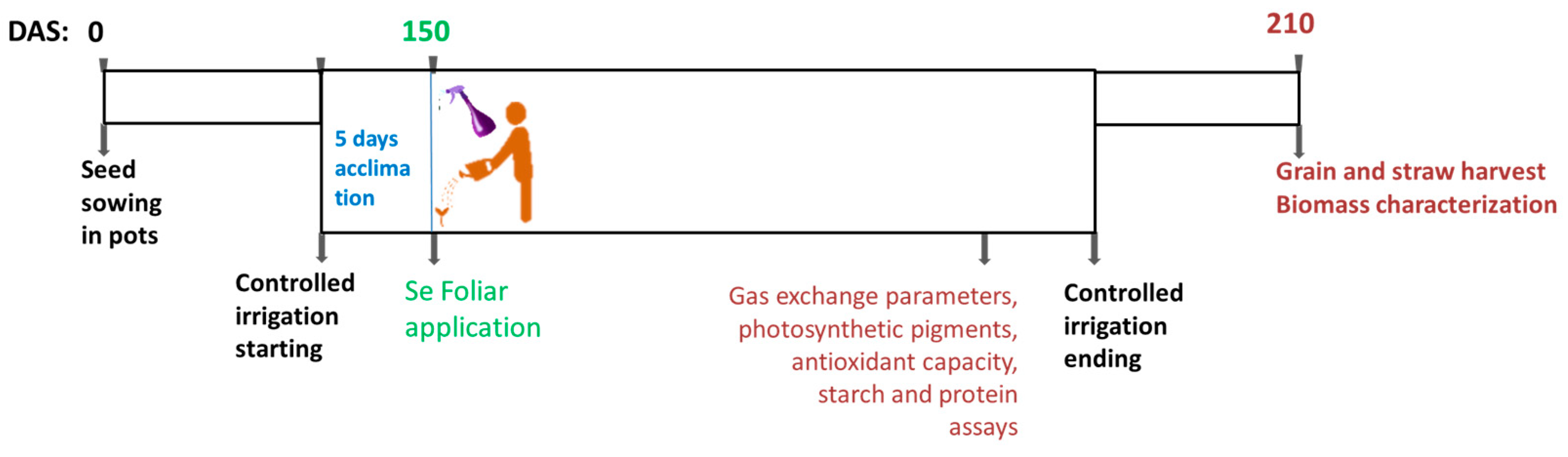
2.2. Pot Trial Description
2.3. Gas Exchange and Chlorophyll a Fluorescence Measurements
2.4. Pigments, Protein, Sugars, and Antioxidant Measurements
2.5. Yield and Biomass Measurements
2.6. Selenium Determination
2.7. Statistical Analysis
3. Results
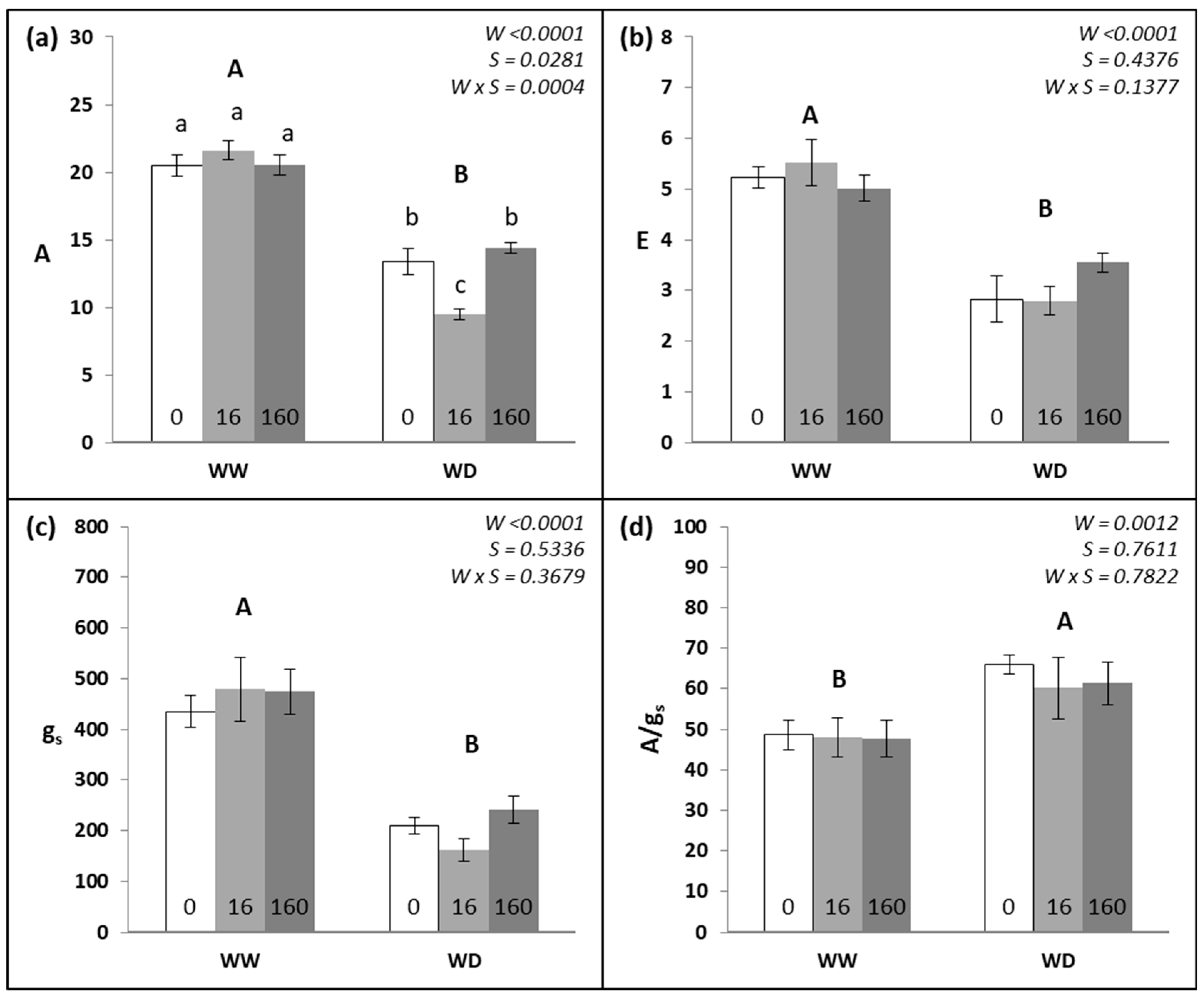
3.1. Leaf Gas Exchange and Chlorophyll a Fluorescence
3.2. Biochemical Analysis of Leaf Samples
3.3. Yield and Growth Parameters
3.4. Selenium Content
4. Discussion
4.1. Leaf Gas Exchange, Chlorophyll a Fluorescence Traits
4.2. Biochemical Analysis of Leaves
4.3. Straw and Grain Biomass Parameters
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roman, M.; Jitaru, P.; Barbante, C. Selenium biochemistry and its role for human health. Metallomics 2014, 6, 25–54. [Google Scholar] [CrossRef]
- Jiang, Y.; Zh, Z.; Bu, Y.; Cz, R.; Jz, L.; Jj, H.; Tao, C.; Zhang, K.; Xx, W.; Gx, L.; et al. Effects of selenium fertilizer on grain yield, Se uptake and distribution in common buckwheat (Fagopyrum esculentum Moench). Plant Soil Environ. 2016, 61, 371–377. [Google Scholar] [CrossRef]
- Fernandes, K.F.M.; Berton, R.; Coscione, A. Selenium biofortification of rice and radish: Effect of soil texture and efficiency of two extractants. Plant Soil Environ. 2014, 60, 105–110. [Google Scholar] [CrossRef]
- Graham, R.; Welch, R.; Saunders, D.; Ortiz-Monasterio, I.; Bouis, H.; Bonierbale, M.; Haan, S.; Burgos, G.; Thiele, G.; Liria, R.; et al. Nutritious Subsistence Food Systems. Adv. Agron. 2007, 92, 1–74. [Google Scholar]
- Puccinelli, M.; Pezzarossa, B.; Rosellini, I.; Malorgio, F. Selenium enrichment enhances the quality and shelf life of Basil leaves. Plants. 2020, 9, 801. [Google Scholar] [CrossRef]
- Mirlean, N.; Seus-Arrache, E.R.; Vlasova, O. Selenium deficiency in subtropical littoral pampas: Environmental and dietary aspects. Environ. Geochem. Health 2018, 40, 543–556. [Google Scholar] [CrossRef]
- Beyene, T.M. Application of Bio-fortification through Plant Breeding to Improve the Value of Staple Crops. J Biomed Biotechnol. 2015, 3, 11–19. [Google Scholar]
- da Silva, D.F.; Cipriano, P.E.; de Souza, R.R.; Siueia Júnior, M.; da Silva, R.F.; Faquin, V.; de Souza Silva, M.L.; Guimarães Guilherme, L.R. Anatomical and physiological characteristics of Raphanus sativus L. submitted to different selenium sources and forms application. Sci. Hortic. 2020, 260, 108839. [Google Scholar] [CrossRef]
- Shahrajabian, M.H.; Sun, W.; Cheng, Q. Foliar application of nutrients on medicinal and aromatic plants, the sustainable approaches for higher and better production. Beni-Suef Univ. J. Basic Appl. Sci. 2022, 11, 26. [Google Scholar] [CrossRef]
- Silva, M.A.; Sousa, G.F.; Van Opbergen, G.A.Z.; Van Opbergen, G.G.A.Z.; Corguinha, A.P.B.; Bueno, J.M.M.; Brunetto, G.; Leite, J.M.; Santos, A.A.; Lopes, G.; et al. Foliar Application of Selenium Associated with a Multi-Nutrient Fertilizer in Soybean: Yield, Grain Quality, and Critical Se Threshold. Plants 2023, 12, 2028. [Google Scholar] [CrossRef] [PubMed]
- Valença, A.W.; Bake, A.; Brouwer, I.D.; Giller, K.E. Agronomic biofortification of crops to fight hidden hunger in sub-Saharan Africa. Glob. Food Secur. 2017, 12, 8–14. [Google Scholar] [CrossRef]
- Mouta, E.R.; Melo, W.J.D.; Soares, M.R.; Alleoni, L.R.F.; Casagrande, J.C. Adsorção de selênio em latossolos. Rev. Bras. Ciência Solo 2008, 32, 1033–1041. [Google Scholar] [CrossRef]
- Araujo, A.M.; Lessa, J.H.D.L.; Ferreira, L.A.; Guilherme, L.R.G.; Lopes, G. Soil management and ionic strength on selenite retention in oxidic soils. Ciência e Agrotecnologia 2018, 42, 395–407. [Google Scholar] [CrossRef]
- Lessa, J.H.L.; Araujo, A.M.; Silva, G.N.T.; Guilherme, L.R.G.; Lopes, G. Adsorption-desorption reactions of selenium (VI) in tropical cultivated and uncultivated soils under Cerrado biome. Chemosphere 2016, 164, 271–277. [Google Scholar] [CrossRef]
- Zhang, Y.; Pan, G.; Chen, J.; Hu, Q. Uptake and transport of selenite and selenate by soybean seedlings of two genotypes. Plant Soil. 2003, 253, 437–443. [Google Scholar] [CrossRef]
- Jiang, C.; Zu, C.; Lu, D.; Zheng, Q.; Shen, J.; Wang, H.; Li, D. Effect of exogenous selenium supply on photosynthesis, Na+ accumulation and antioxidative capacity of maize (Zea mays L.) under salinity stress. Sci. Rep. 2017, 7, 42039. [Google Scholar] [CrossRef]
- Nawaz, F.; Ahmad, R.; Ashraf, M.Y.; Waraich, E.A.; Khan, S.Z. Effect of selenium foliar spray on physiological and biochemical processes and chemical constituents of wheat under drought stress. Ecotoxicol. Environ. Saf. 2015, 113, 191–200. [Google Scholar] [CrossRef]
- Zhang, M.; Tang, S.; Huang, X.; Zhang, F.; Pang, Y.; Huang, Q.; Yi, Q. Selenium uptake, dynamic changes in selenium content and its influence on photosynthesis and chlorophyll fluorescence in rice (Oryza sativa L.). Environ. Exp. Bot. 2014, 107, 39–45. [Google Scholar] [CrossRef]
- Hawrylak-Nowak, B. Comparative effects of selenite and selenate on growth and selenium accumulation in lettuce plants under hydroponic conditions. Plant Growth Regul. 2013, 70, 149–157. [Google Scholar] [CrossRef]
- Mostofa, M.G.; Hossain, M.A.; Siddiqui, M.N.; Fujita, M.; Tran, L.-S.P. Phenotypical, physiological and biochemical analyses provide insight into selenium-induced phytotoxicity in rice plants. Chemosphere 2017, 178, 212–223. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations, Statistical Database. Available online: http://www.fao.org/faostat/en/#data/ (accessed on 27 January 2025).
- Liu, J.; Feng, H.; He, J.; Chen, H.; Ding, D. The effects of nitrogen and water stresses on the nitrogen-to-protein conversion factor of winter wheat. Agric. Water Manag. 2018, 210, 217–223. [Google Scholar] [CrossRef]
- Zörb, C.; Ludewig, U.; Hawkesford, M.J. Perspective on wheat yield and quality with reduced nitrogen supply. Trends Plant Sci. 2018, 23, 1029–1037. [Google Scholar] [CrossRef]
- Ramkissoon, C.; Degryse, F.; da Silva, R.; Baird, R.; Young, S.; Bailey, E.; McLaughlin, M. Improving the efficacy of selenium fertilizers for wheat biofortification. Sci. Rep. 2019, 9, 19520. [Google Scholar] [CrossRef]
- Radawiec, A.; Szulc, W.; Rutkowska, B. Selenium biofortification of wheat as a strategy to improve Human nutrition. Agriculture 2021, 11, 144. [Google Scholar] [CrossRef]
- Galinha, C.; Pacheco, A.M.G.; Freitas, M.D.; Fikrle, M.; Kucera, J.; Coutinho, J.; Macas, B.; Almeida, A.S.; Wolterbeek, H.T. Selenium in bread and durum wheats grown under a soil-supplementation regime in actual field conditions, determined by cyclic and radiochemical neutron activation analysis. J. Radioanal. Nucl. Chem. 2015, 304, 139–143. [Google Scholar] [CrossRef]
- Manojlović, M.S.; Lončarić, Z.; Cabilovski, R.R.; Popović, B.; Karalić, K.; Ivezić, V.; Ademi, A.; Singh, B.R. Biofortification of wheat cultivars with selenium. Acta Agric. Scand. Sect. B-Soil Plant Sci. 2019, 69, 715–724. [Google Scholar] [CrossRef]
- Min, W.; Ali, F.; Wang, M.; Dinh, T.; Fei, Z.; Banuelos, G.; Liang, D. Understanding boosting selenium accumulation in wheat (Triticum aestivum L.) following foliar selenium application at different stages, forms, and doses. Environ. Sci. Pollut. Res. 2019, 27, 717–772. [Google Scholar]
- Ahmed, M.; Fayyaz ul, H. Response of spring wheat (Triticum aestivum L.) quality traits and yield to sowing date. PLoS ONE 2015, 10, e0126097. [Google Scholar] [CrossRef]
- Liu, X.; Pan, Y.; Zhu, X.; Yang, T.; Bai, J.; Sun, Z. Drought evolution and its impact on the crop yield in the North China Plain. J. Hydrol. 2018, 564, 984–996. [Google Scholar] [CrossRef]
- Kuwayama, Y.; Thompson, A.; Bernknopf, R.; Zaitchik, B.; Vail, P. Estimating the Impact of Drought on Agriculture Using the U.S. Drought Monitor. Am. J. Agric. Econ. 2019, 101, 193–210. [Google Scholar] [CrossRef]
- Sodani, R.; Singhal, R.K.; Gupta, S.; Gupta, N.; Chauhan, K.S.; Chauhan, J. Performance of Yield and Yield Attributes of Ten Indian Mustard (Brassica juncea L.) Genotypes under Drought Stress. Int. J. Pure Appl. Biosci. 2017, 5, 467–476. [Google Scholar]
- Rocha, L.; Silva, E.; Pavia, I.; Ferreira, H.; Matos, C.; Osca, J.M.; Moutinho-Pereira, J.; Lima-Brito, J. Seed Soaking with Sodium Selenate as a Biofortification Approach in Bread Wheat: Effects on Germination, Seedling Emergence, Biomass and Responses to Water Deficit. Agronomy 2022, 12, 1975. [Google Scholar] [CrossRef]
- DGAV. Catálogo nacional de variedades e espécies agrícolas e hortícolas 2020; Ministério da Agricultura, Florestas e Desenvolvimento Rural: Lisboa, Portugal, 2020; ISSN 0871-0295.
- Galinha, C.; Freitas, M.C.; Pacheco, A.M.G.; Coutinho, J.; Macas, B.; Almeida, A.S. Determination of selenium in bread-wheat samples grown under a Se-supplementation regime in actual field conditions. J. Radioanal. Nucl. Chem. 2012, 291, 231–235. [Google Scholar] [CrossRef]
- Almeida, A.; Maçãs, B.; Rodrigues, V.; Torrão, M. Wheat breeding: Country perspectives. In The History of Wheat Breeding in Portugal; Bonjean, A.P., Angus, W.J., Van Ginkel, M., Eds.; LAVOISIER S.A.S.: Lisbon, Portugal, 2016; pp. 93–125. [Google Scholar]
- Zadoks Growth Scale. 2025. Available online: https://www.agric.wa.gov.au/grains/zadoks-growth-scale (accessed on 22 February 2025).
- von Caemmerer, S.; Farquhar, G.D. Some relationships between the biochemistry of photosynthesis and the gas exchange of leaves. Planta 1981, 153, 376–387. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Irigoyen, J.J.; Einerich, D.W.; Sánchez-Díaz, M. Water stress induced changes in concentrations of proline and total soluble sugars in nodulated alfalfa (Medicago sativd) plants. Physiol. Plant. 1992, 84, 55–60. [Google Scholar] [CrossRef]
- Boldrin, P.F.; Faquin, V.; Ramos, S.J.; Boldrin, K.V.F.; Ávila, F.W.; Guilherme, L.R.G. Soil and foliar application of selenium in rice biofortification. J. Food Compos. Anal. 2013, 31, 238–244. [Google Scholar] [CrossRef]
- Oguz, M.C.; Aycan, M.; Oguz, E.; Poyraz, I.; Yildiz, M. Drought Stress Tolerance in Plants: Interplay of Molecular, Biochemical and Physiological Responses in Important Development Stages. Physiologia 2022, 2, 180–197. [Google Scholar] [CrossRef]
- Dias, M.C.; Correia, S.; Serôdio, J.; Silva, A.M.S.; Freitas, H.; Santos, C. Chlorophyll fluorescence and oxidative stress endpoints to discriminate olive cultivars tolerance to drought and heat episodes. Sci. Hortic. 2018, 231, 31–35. [Google Scholar] [CrossRef]
- Saha, D.; Choyal, P.; Mishra, U.N.; Dey, P.; Bose, B.; Md, P.; Gupta, N.K.; Mehta, B.K.; Kumar, P.; Pandey, S.; et al. Drought stress responses and inducing tolerance by seed priming approach in plants. Plant Stress 2022, 4, 100066. [Google Scholar] [CrossRef]
- Moualeu-Ngangue, D.P.; Chen, T.W.; Stützel, H. A new method to estimate photosynthetic parameters through net assimilation rate−intercellular space CO2 concentration (A–Ci) curve and chlorophyll fluor. New Phytol. 2017, 213, 1543–1554. [Google Scholar] [CrossRef]
- Wang, M.; Dinh, T.; Qi, M.; Min, W.; Yang, W.; Fei, Z.; Liang, D. Radicular and foliar uptake, and xylem- and phloem-mediated transport of selenium in maize (Zea mays L.): A comparison of five Se exogenous species. Plant Soil 2019, 446, 111–123. [Google Scholar] [CrossRef]
- Zhang, A.-l.; Su, L.; Jiang, Z.-q.; Kang, Y.-t.; Qiu, P. Cyclic loading tests of earthquake-resilient prefabricated steel cross joints with different FCP connections. Structures 2021, 32, 1–14. [Google Scholar] [CrossRef]
- Diao, M.; Ma, L.; Wang, J.; Cui, J.; Fu, A.; Liu, H.-Y. Selenium Promotes the Growth and Photosynthesis of Tomato Seedlings Under Salt Stress by Enhancing Chloroplast Antioxidant Defense System. J. Plant Growth Regul. 2014, 33, 671–682. [Google Scholar] [CrossRef]
- Proietti, P.; Nasini, L.; Del Buono, D.; D’Amato, R.; Tedeschini, E.; Businelli, D. Selenium protects olive (Olea europaea L.) from drought stress. Sci. Hortic. 2013, 164, 165–171. [Google Scholar] [CrossRef]
- Santos, L.C.d.; Martins, G.S.; Benevenute, P.A.N.; de Sousa Lima, J.; Santos, F.R.d.; Andrade, O.V.S.; de Oliveira, I.P.; Bispo, F.H.A.; Botelho, L.; Rabêlo, F.H.S.; et al. Soil Application of Selenium in Wheat (Triticum aestivum L.) Under Water Stress Improves Grain Quality and Reduces Production Losses. Plants 2024, 13, 3460. [Google Scholar] [CrossRef]
- Sousa, G.F.d.; Silva, M.A.; Carvalho, M.R.d.; Morais, E.G.d.; Benevenute, P.A.N.; Van Opbergen, G.A.Z.; Van Opbergen, G.G.A.Z.; Guilherme, L.R.G. Foliar Selenium Application to Reduce the Induced-Drought Stress Effects in Coffee Seedlings: Induced Priming or Alleviation Effect? Plants 2023, 12, 3026. [Google Scholar] [CrossRef]
- Hajiboland, R.; Sadeghzadeh, N.; Ebrahimi, N.; Sadeghzadeh, B.; Mohammadi, S. Influence of selenium in drought-stressed wheat plants under greenhouse and field conditions. Acta Agric. Slov. 2015, 105, 175–191. [Google Scholar] [CrossRef]
- Nawaz, F.; Naeem, M.; Ashraf, M.Y.; Tahir, M.N.; Zulficiar, B.; Salahuddin, M.; Shabbir, R.N.; Aslam, M. Selenium supplementation affects physiological and biochemical processes to improve fodder yield and quality of maize (Zea mays L.) under water deficit conditions. Front. Plant Sci. 2016, 7, 1438. [Google Scholar] [CrossRef]
- Liu, Y.N.; Xu, Q.Z.; Li, W.C.; Yang, X.H.; Zheng, Q.; Li, B.; Li, Z.S.; Li, H.W. Long-term high light stress induces leaf senescence in wheat (Triticum aestivum L.). Photosynthetica 2019, 57, 830–840. [Google Scholar] [CrossRef]
- Yao, X.; Chu, J.; Wang, G. Effects of Selenium on Wheat Seedlings Under Drought Stress. Biol. Trace Elem. Res. 2009, 130, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef]
- Guneidy, R.A.; Zaki, E.R.; Gad, A.A.M.; Saleh, N.S.E.; Shokeer, A. Evaluation of Phenolic Content Diversity along with Antioxidant/Pro-Oxidant, Glutathione Transferase Inhibition, and Cytotoxic Potential of Selected Commonly Used Plants. Prev. Nutr. Food Sci. 2022, 27, 282–298. [Google Scholar] [CrossRef] [PubMed]
- Begum, N.; Hasanuzzaman, M.; Li, Y.; Akhtar, K.; Zhang, C.; Zhao, T. Seed Germination Behavior, Growth, Physiology and Antioxidant Metabolism of Four Contrasting Cultivars under Combined Drought and Salinity in Soybean. Antioxidants 2022, 11, 498. [Google Scholar] [CrossRef]
- Hou, P.; Wang, F.; Luo, B.; Li, A.; Wang, C.; Shabala, L.; Ahmed, H.A.I.; Deng, S.; Zhang, H.; Song, P.; et al. Antioxidant Enzymatic Activity and Osmotic Adjustment as Components of the Drought Tolerance Mechanism in Carex duriuscula. Plants 2021, 10, 436. [Google Scholar] [CrossRef]
- Ozturk, M.; Turkyilmaz Unal, B.; García-Caparrós, P.; Khursheed, A.; Gul, A.; Hasanuzzaman, M. Osmoregulation and its actions during the drought stress in plants. Physiol. Plant. 2021, 172, 1321–1335. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, T.; Abdullah, M.; Ahmar, S.; Yasir, M.; Iqbal, M.S.; Yasir, M.; Ur Rehman, S.; Ahmed, S.; Rana, R.M.; Ghafoor, A.; et al. Incredible Role of Osmotic Adjustment in Grain Yield Sustainability under Water Scarcity Conditions in Wheat (Triticum aestivum L.). Plants 2020, 9, 1208. [Google Scholar] [CrossRef] [PubMed]
- Abobatta, W. Drought adaptive mechanisms of plants—A review. Adv. Agric. Environ. Sci. Open Access 2019, 2, 42–45. [Google Scholar] [CrossRef]
- Somayyeh, S.; Masouleh, S.; Sassine, Y.; Jamal Aldine, N. Ornamental Horticulture The role of organic solutes in the osmotic adjustment of chilling stressed plants (vegetable, ornamental and crop plants). Ornam. Hortic. 2019, 25, 434–442. [Google Scholar]
- Gautam, T.; Dutta, M.; Jaiswal, V.; Zinta, G.; Gahlaut, V.; Kumar, S. Emerging Roles of SWEET Sugar Transporters in Plant Development and Abiotic Stress Responses. Cells 2022, 11, 1303. [Google Scholar] [CrossRef]
- Brito, C.; Dinis, L.T.; Ferreira, H.; Coutinho, J.; Moutinho-Pereira, J.; Correia, C.M. Salicylic acid increases drought adaptability of young olive trees by changes on redox status and ionome. Plant Physiol. Biochem. 2019, 141, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Bacelar, E.A.; Santos, D.L.; Moutinho-Pereira, J.M.; Lopes, J.I.; Gonçalves, B.C.; Ferreira, T.C.; Correia, C.M. Physiological behaviour, oxidative damage and antioxidative protection of olive trees grown under different irrigation regimes. Plant Soil 2007, 292, 1–12. [Google Scholar] [CrossRef]
- Kabiri, R.; Nasibi, F.; Farahbakhsh, H. Effect of exogenous salicylic acid on some physiological parameters and alleviation of drought stress in Nigella sativa plant under hydroponic culture. Plant Prot. Sci. 2014, 50, 43–51. [Google Scholar] [CrossRef]
- Wang, Y.-D.; Wang, X.; Wong, Y.-s. Proteomics analysis reveals multiple regulatory mechanisms in response to selenium in rice. J. Proteom. 2012, 75, 1849–1866. [Google Scholar] [CrossRef]
- Andrade, F.R.; Da Silva, G.N.; Guimarães, K.C.; Barreto, H.B.F.; De Souza, K.R.D.; Guilherme, L.R.G.; Faquin, V.; Reis, A.R.D. Selenium protects rice plants from water deficit stress. Ecotoxicol. Environ. Saf. 2018, 164, 562–570. [Google Scholar] [CrossRef]
- Feng, T.; Chen, S.; Gao, D.; Liu, G.; Bai, H.; Li, A.; Peng, L.; Ren, Z. Selenium improves photosynthesis and protects photosystem II in pear (Pyrus bretschneideri), grape (Vitis vinifera), and peach (Prunus persica). Photosynthetica 2015, 53, 609–612. [Google Scholar] [CrossRef]
- Zahedi, S.M.; Moharrami, F.; Sarikhani, S.; Padervand, M. Selenium and silica nanostructure-based recovery of strawberry plants subjected to drought stress. Sci. Rep. 2020, 10, 17672. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Fv/Fm | ΦPSII | F’v/F’m | qP | NPQ | ETR | |
|---|---|---|---|---|---|---|
| Water Regime (W) | ||||||
| WW | 0.886 ± 0.005 a | 0.536 ± 0.016 | 0.728 ± 0.010 | 0.736 ± 0.018 | 1.209 ± 0.176 | 101 ± 2.1 a |
| WD | 0.857 ± 0.007 b | 0.512 ± 0.020 | 0.695 ± 0.019 | 0.735 ± 0.016 | 1.297 ± 0.171 | 95 ± 2.2 b |
| Se foliar spray (S) | ||||||
| 0 g ha−1 | 0.876 ± 0.012 | 0.513 ± 0.022 | 0.711 ± 0.021 | 0.720 ± 0.017 | 1.160 ± 0.272 | 95 ± 2.6 |
| 16 g ha−1 | 0.868 ± 0.006 | 0.530 ± 0.022 | 0.708 ± 0.018 | 0.748 ± 0.023 | 1.156 ± 0.120 | 100 ± 3.5 |
| 160 g ha−1 | 0.871 ± 0.009 | 0.529 ± 0.024 | 0.714 ± 0.019 | 0.739 ± 0.022 | 1.428 ± 0.215 | 98 ± 2.1 |
| W × S | ||||||
| WW × 0 g ha−1 | 0.906 ± 0.002 | 0.590 ± 0.008 | 0.770 ± 0.005 | 0.766 ± 0.016 | 0.473 ± 0.048 | 99 ± 1.4 |
| WW × 16 g ha−1 | 0.900 ± 0.003 | 0.548 ± 0.013 | 0.717 ± 0.006 | 0.764 ± 0.012 | 0.540 ± 0.185 | 92 ± 2.2 |
| WW × 160 g ha−1 | 0.908 ± 0.011 | 0.603 ± 0.017 | 0.765 ± 0.011 | 0.790 ± 0.033 | 0.539 ± 0.010 | 101 ± 2.9 |
| WD × 0 g ha−1 | 0.892 ± 0.009 | 0.529 ± 0.041 | 0.751 ± 0.016 | 0.702 ± 0.039 | 0.578 ± 0.038 | 89 ± 6.9 |
| WD × 16 g ha−1 | 0.875 ± 0.008 | 0.593 ± 0.014 | 0.767 ± 0.009 | 0.772 ± 0.009 | 1.122 ± 0.166 | 100 ± 2.3 |
| WD × 160 g ha−1 | 0.863 ± 0.010 | 0.601 ± 0.019 | 0.735 ± 0.000 | 0.818 ± 0.026 | 1.065 ± 0.007 | 101 ± 3.2 |
| ANOVA (p-values) | ||||||
| W | 0.0049 | 0.3539 | 0.1525 | 0.8873 | 0.7702 | 0.0372 |
| S | 0.8839 | 0.7975 | 0.9773 | 0.6253 | 0.5761 | 0.2681 |
| W × S | 0.2244 | 0.7468 | 0.9002 | 0.5610 | 0.3914 | 0.4524 |
| Chl a (mg g−1 DW) | Chl b (mg g−1 DW) | Car (mg g−1 DW) | Chl a + b (mg g−1 DW) | Chl a/Chl b | Chl/Car | |
|---|---|---|---|---|---|---|
| Water Regime (W) | ||||||
| WW | 13.8 ± 0.28 a | 7.71 ± 0.13 a | 3.47 ± 0.07 a | 20.4 ± 0.39 a | 1.79 ± 0.01 a | 5.87 ± 0.03 a |
| WD | 11.5 ± 0.36 b | 6.54 ± 0.14 b | 2.89 ± 0.09 b | 17.1 ± 0.47 b | 1.73 ± 0.02 b | 5.70 ± 0.03 b |
| Se foliar spray (S) | ||||||
| 0 g ha−1 | 13.6 ± 0.15 a | 7.53 ± 0.11 a | 3.45 ± 0.06 a | 20.0 ± 0.23 a | 1.78 ± 0.02 a | 5.85 ± 0.04 a |
| 16 g ha−1 | 13.0 ± 0.80 b | 7.32 ± 0.34 b | 3.13 ± 0.19 b | 19.2 ± 1.07 b | 1.74 ± 0.03 b | 5.77 ± 0.03 ab |
| 160 g ha−1 | 11.3 ± 0.11 c | 6.50 ± 0.06 c | 2.95 ± 0.03 c | 16.9 ± 0.16 c | 1.74 ± 0.01 b | 5.72 ± 0.05 b |
| W × S | ||||||
| WW × 0 g ha−1 | 14.2 ± 0.11 b | 7.95 ± 0.10 a | 3.53 ± 0.03 b | 20.9 ± 0.15 b | 1.75 ± 0.02 bc | 5.81 ± 0.04 a |
| WW × 16 g ha−1 | 15.3 ± 0.15 a | 8.30 ± 0.08 a | 3.86 ± 0.04 a | 22.4 ± 0.21 a | 1.85 ± 0.01 a | 5.76 ± 0.03 a |
| WW × 160 g ha−1 | 11.6 ± 0.11 d | 6.68 ± 0.07 c | 2.94 ± 0.04 d | 17.3 ± 0.15 d | 1.74 ± 0.02 c | 5.89 ± 0.05 a |
| WD × 0 g ha−1 | 13.1 ± 0.17 c | 7.12 ± 0.09 b | 3.35 ± 0.05 c | 19.1 ± 0.19 c | 1.80 ± 0.01 ab | 5.68 ± 0.04 a |
| WD × 16 g ha−1 | 8.7 ± 0.04 f | 5.52 ± 0.05 e | 2.24 ± 0.04 e | 13.4 ± 0.05 f | 1.59 ± 0.01 d | 5.75 ± 0.05 a |
| WD × 160 g ha−1 | 11.0 ± 0.14 e | 6.30 ± 0.06 d | 2.96 ± 0.05 d | 16.4 ± 0.18 e | 1.74 ± 0.01 c | 5.55 ± 0.04 b |
| ANOVA (p-values) | ||||||
| W | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| S | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | 0.0105 |
| W × S | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | 0.0056 |
| Stem Length (mm) | Spike Length (mm) | Tillers (Number per Plant−1) | Total Grain (Number per Plant−1) | Thousand-Grain Weight (g) | Grain Biomass (mg Plant−1) | Grain Harvest Index (%) | Straw Biomass (mg Plant−1) | Vegetative Biomass (mg Plant−1) | |
|---|---|---|---|---|---|---|---|---|---|
| Water Regime (W) | |||||||||
| WW | 568 ± 8.79 a | 77 ± 1.5 | 3.7 ± 0.22 | 93 ± 3.9 a | 30.9 ± 0.68 | 2855 ± 131 a | 49 ± 1.28 a | 2961 ± 45 a | 5770 ± 132 a |
| WD | 527 ± 8.86 b | 80 ± 1.4 | 3.3 ± 0.15 | 65 ± 3.4 b | 30.1 ± 0.68 | 1934 ± 97 b | 41 ± 1.22 b | 2718 ± 49 b | 4652 ± 112 b |
| Se foliar spray (S) | |||||||||
| 0 g ha−1 | 539 ± 10.5 | 78 ± 1.92 | 4.1 ± 0.28 | 83 ± 5.2 | 32.7 ± 0.70 a | 2715 ± 176 | 48 ± 1.67 | 2795 ± 52 | 5511 ± 194 |
| 16 g ha−1 | 554 ± 11.5 | 79 ± 1.34 | 3.5 ± 0.32 | 79 ± 5.9 | 29.6 ± 0.82 b | 2349 ± 180 | 44 ± 1.90 | 2811 ± 63 | 5160 ± 184 |
| 160 g ha−1 | 549 ± 11.7 | 79 ± 1.94 | 3.5 ± 0.25 | 78 ± 4.6 | 28.6 ± 0.69 b | 2220 ± 126 | 43 ± 1.44 | 2834 ± 64 | 5034 ± 140 |
| W × S | |||||||||
| WW × 0 g ha−1 | 555 ± 7.3 | 80 ± 1.90 ab | 3.8 ± 0.17 | 108 ± 3.3 | 32.8 ± 1.10 a | 3532 ± 153 a | 56 ± 1.12 a | 2887 ± 86 b | 6419 ± 183 |
| WW × 16 g ha−1 | 591 ± 17.8 | 77 ± 3.21 ab | 4.0 ± 0.54 | 100 ± 8.4 | 32.1 ± 0.93 a | 3156 ± 246 ab | 52 ± 1.70 ab | 2761 ± 52 b | 5917 ± 257 |
| WW × 160 g ha−1 | 594 ± 17.4 | 76 ± 2.81 ab | 4.3 ± 0.59 | 93 ± 11.2 | 27.9 ± 0.92 b | 2569 ± 303 bc | 44 ± 2.59 c | 3131 ± 61 a | 5700 ± 325 |
| WD × 0 g ha−1 | 518 ± 19.8 | 75 ± 2.97 b | 4.4 ± 0.49 | 61 ± 4.5 | 32.2 ± 0.95 a | 1929 ± 135 c | 41 ± 1.48 c | 2755 ± 58 b | 4685 ± 157 |
| WD × 16 g ha−1 | 534 ± 15.7 | 82 ± 1.27 a | 3.1 ± 0.29 | 71 ± 7.7 | 27.3 ± 1.01 b | 1903 ± 194 c | 39 ± 2.35 c | 2886 ± 100 b | 4789 ± 224 |
| WD × 160 g ha−1 | 5112 ± 9.2 | 82 ± 1.69 a | 3.3 ± 0.23 | 73 ± 5.6 | 30.4 ± 1.08 ab | 2165 ± 155 c | 45 ± 1.80 bc | 2548 ± 36 c | 4713 ± 162 |
| ANOVA (p-values) | |||||||||
| W | <0.0001 | 0.3381 | 0.2168 | <0.0001 | 0.2317 | <0.0001 | <0.0001 | 0.0010 | <0.0001 |
| S | 0.2860 | 0.6808 | 0.4086 | 0.9596 | 0.0028 | 0.2442 | 0.2125 | 0.9615 | 0.3446 |
| W × S | 0.3888 | 0.0368 | 0.0838 | 0.2122 | 0.0016 | 0.0187 | 0.0001 | <0.0001 | 0.2495 |
| Grain Se (µg Plant−1) | Straw Se (µg Plant−1) | |
|---|---|---|
| Water Regime (W) | ||
| WW | 892 ± 340 | 1964 ± 643 a |
| WD | 310 ± 160 | 1505 ± 749 b |
| Se foliar spray (S) | ||
| 0 g ha−1 | 27 ± 9.4 c | 290 ± 100 c |
| 16 g ha−1 | 207 ± 31 b | 798 ± 76.2 b |
| 160 g ha−1 | 1715 ± 406 a | 4253 ± 171 a |
| W × S | ||
| WW × 0 g ha−1 | 41 ± 6.65 | 452 ± 30.1 d |
| WW × 16 g ha−1 | 257 ± 7.26 | 919 ± 38.9 c |
| WW × 160 g ha−1 | 2165 ± 458 | 4521 ± 89.5 a |
| WD × 0 g ha−1 | 6 ± 0.31 | 48 ± 2.17 e |
| WD × 16 g ha−1 | 109 ± 8.60 | 617 ± 1.67 d |
| WD × 160 g ha−1 | 815 ± 3.05 | 3751 ± 46.9 b |
| ANOVA (p-values) | ||
| W | 0.0600 | <0.0001 |
| S | 0.0008 | <0.0001 |
| W × S | 0.0899 | 0.0240 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rocha, L.; Silva, E.; Gonçalves, A.; Brito, C.; Ferreira, H.; Matos, C.; Malheiro, A.C.; Araújo, S.; Lima-Brito, J.; Moutinho-Pereira, J. Biomass, Physiological, and Antioxidant Activity Responses of Wheat Plants After Selenium Foliar Spray Under Water Deficit. Agriculture 2025, 15, 1086. https://doi.org/10.3390/agriculture15101086
Rocha L, Silva E, Gonçalves A, Brito C, Ferreira H, Matos C, Malheiro AC, Araújo S, Lima-Brito J, Moutinho-Pereira J. Biomass, Physiological, and Antioxidant Activity Responses of Wheat Plants After Selenium Foliar Spray Under Water Deficit. Agriculture. 2025; 15(10):1086. https://doi.org/10.3390/agriculture15101086
Chicago/Turabian StyleRocha, Luís, Ermelinda Silva, Alexandre Gonçalves, Cátia Brito, Helena Ferreira, Carlos Matos, Aureliano C. Malheiro, Susana Araújo, José Lima-Brito, and José Moutinho-Pereira. 2025. "Biomass, Physiological, and Antioxidant Activity Responses of Wheat Plants After Selenium Foliar Spray Under Water Deficit" Agriculture 15, no. 10: 1086. https://doi.org/10.3390/agriculture15101086
APA StyleRocha, L., Silva, E., Gonçalves, A., Brito, C., Ferreira, H., Matos, C., Malheiro, A. C., Araújo, S., Lima-Brito, J., & Moutinho-Pereira, J. (2025). Biomass, Physiological, and Antioxidant Activity Responses of Wheat Plants After Selenium Foliar Spray Under Water Deficit. Agriculture, 15(10), 1086. https://doi.org/10.3390/agriculture15101086