
Susceptibility Is New Resistance: Wheat Susceptibility Genes and Exploitation in Resistance Breeding

Abstract
:1. Introduction
2. Wheat S Genes Supporting Pathogen (Pre)Penetration
3. Wheat S Genes Suppressing Plant Immunity
4. Wheat S Genes Facilitating Pathogen Sustenance and Pest Feeding
5. Pathogen Effectors Targeting Wheat S Genes
6. Strategies and Challenges on Exploiting Wheat S Genes in Resistance Breeding
7. Concluding Remarks and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Levy, A.A.; Feldman, M. Evolution and origin of bread wheat. Plant Cell 2022, 34, 2549–2567. [Google Scholar] [CrossRef] [PubMed]
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Sinha, A.; Singh, L.; Rawat, N. Current understanding of atypical resistance against fungal pathogens in wheat. Curr. Opin. Plant Biol. 2022, 68, 102247. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.M.; Zhang, Y. Plant immunity: Danger perception and signaling. Cell 2020, 181, 978–989. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhu, X.; Chen, X.; Zhou, J.M. From plant immunity to crop disease resistance. J. Genet. Genom. 2022; in press. [Google Scholar] [CrossRef]
- Li, W.; Deng, Y.; Ning, Y.; He, Z.; Wang, G.L. Broad-spectrum disease resistance in crops: From molecular dissection to breeding. Annu. Rev. Plant Biol. 2020, 71, 575–603. [Google Scholar] [CrossRef]
- Fuchs, M. Pyramiding resistance-conferring gene sequences in crops. Curr. Opin. Virol. 2017, 26, 36–42. [Google Scholar] [CrossRef]
- Deng, Y.; Ning, Y.; Yang, D.L.; Zhai, K.; Wang, G.L.; He, Z. Molecular basis of disease resistance and perspectives on breeding strategies for resistance improvement in crops. Mol. Plant 2020, 13, 1402–1419. [Google Scholar] [CrossRef]
- van Schie, C.C.; Takken, F.L. Susceptibility genes 101: How to be a good host. Annu. Rev. Phytopathol. 2014, 52, 551–581. [Google Scholar] [CrossRef]
- Zaidi, S.S.; Mukhtar, M.S.; Mansoor, S. Editing: Targeting susceptibility genes for plant disease resistance. Trends Biotechnol. 2018, 36, 898–906. [Google Scholar] [CrossRef]
- Koseoglou, E.; van der Wolf, J.M.; Visser, R.; Bai, Y. Susceptibility reversed: Modified plant susceptibility genes for resistance to bacteria. Trends Plant Sci. 2022, 27, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Chang, C. Evolutionary insight of plant cuticle biosynthesis in bryophytes. Plant Signal. Behav. 2021, 16, 1943921. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wang, X.; Chang, C. Toward a smart skin: Harnessing cuticle biosynthesis for crop adaptation to drought, salinity, temperature, and ultraviolet stress. Front. Plant Sci. 2022, 13, 961829. [Google Scholar] [CrossRef]
- Wang, X.; Kong, L.; Zhi, P.; Chang, C. Cuticular wax biosynthesis and its roles in plant disease resistance. Int. J. Mol. Sci. 2020, 21, 5514. [Google Scholar] [CrossRef] [PubMed]
- Arya, G.C.; Sarkar, S.; Manasherova, E.; Aharoni, A.; Cohen, H. The plant cuticle: An ancient guardian barrier set against long-standing rivals. Front. Plant Sci. 2021, 12, 663165. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Chang, C. Suppression of wheat TaCDK8/TaWIN1 interaction negatively affects germination of Blumeria graminis f. sp. tritici by interfering with very-long-chain aldehyde biosynthesis. Plant Mol. Biol. 2018, 96, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhi, P.; Fan, Q.; Zhang, M.; Chang, C. Wheat CHD3 protein TaCHR729 regulates the cuticular wax biosynthesis required for stimulating germination of Blumeria graminis f. sp. tritici. J. Exp. Bot. 2019, 70, 701–713. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Zhi, P.; Liu, J.; Li, H.; Zhang, X.; Xu, J.; Zhou, J.; Wang, X.; Chang, C. Epigenetic activation of Enoyl-CoA Reductase by an acetyltransferase complex triggers wheat wax biosynthesis. Plant Physiol. 2020, 183, 1250–1267. [Google Scholar] [CrossRef]
- Büschges, R.; Hollricher, K.; Panstruga, R.; Simons, G.; Wolter, M.; Frijters, A.; van Daelen, R.; van der Lee, T.; Diergaarde, P.; Groenendijk, J.; et al. The barley Mlo gene: A novel control element of plant pathogen resistance. Cell 1997, 88, 695–705. [Google Scholar] [CrossRef]
- Bai, Y.; Pavan, S.; Zheng, Z.; Zappel, N.F.; Reinstädler, A.; Lotti, C.; De Giovanni, C.; Ricciardi, L.; Lindhout, P.; Visser, R.; et al. Naturally occurring broad-spectrum powdery mildew resistance in a Central American tomato accession is caused by loss of mlo function. Mol. Plant Microbe Interact. 2008, 21, 30–39. [Google Scholar] [CrossRef] [Green Version]
- Consonni, C.; Humphry, M.E.; Hartmann, H.A.; Livaja, M.; Durner, J.; Westphal, L.; Vogel, J.; Lipka, V.; Kemmerling, B.; Schulze-Lefert, P.; et al. Conserved requirement for a plant host cell protein in powdery mildew pathogenesis. Nat. Genet. 2006, 38, 716–720. [Google Scholar] [CrossRef] [PubMed]
- Humphry, M.; Reinstädler, A.; Ivanov, S.; Bisseling, T.; Panstruga, R. Durable broad-spectrum powdery mildew resistance in pea er1 plants is conferred by natural loss-of-function mutations in PsMLO1. Mol. Plant Pathol. 2011, 12, 866–878. [Google Scholar] [CrossRef] [PubMed]
- Jiwan, D.; Roalson, E.H.; Main, D.; Dhingra, A. Antisense expression of peach mildew resistance locus O (PpMlo1) gene confers cross-species resistance to powdery mildew in Fragaria x ananassa. Transgenic Res. 2013, 22, 1119–1131. [Google Scholar] [CrossRef] [PubMed]
- Miklis, M.; Consonni, C.; Bhat, R.A.; Lipka, V.; Schulze-Lefert, P.; Panstruga, R. Barley MLO modulates actin-dependent and actin-independent antifungal defense pathways at the cell periphery. Plant Physiol. 2007, 144, 1132–1143. [Google Scholar] [CrossRef]
- Várallyay, E.; Giczey, G.; Burgyán, J. Virus-induced gene silencing of MLO genes induces powdery mildew resistance in Triticum aestivum. Arch. Virol. 2012, 157, 1345–1350. [Google Scholar] [CrossRef]
- Acevedo-Garcia, J.; Spencer, D.; Thieron, H.; Reinstädler, A.; Hammond-Kosack, K.; Phillips, A.L.; Panstruga, R. mlo-based powdery mildew resistance in hexaploid bread wheat generated by a non-transgenic TILLING approach. Plant Biotechnol. J. 2017, 15, 367–378. [Google Scholar] [CrossRef]
- Wang, Y.; Cheng, X.; Shan, Q.; Zhang, Y.; Liu, J.; Gao, C.; Qiu, J.L. Simultaneous editing of three homoeoalleles in hexaploid bread wheat confers heritable resistance to powdery mildew. Nat. Biotechnol. 2014, 32, 947–951. [Google Scholar] [CrossRef]
- Li, S.; Lin, D.; Zhang, Y.; Deng, M.; Chen, Y.; Lv, B.; Li, B.; Lei, Y.; Wang, Y.; Zhao, L.; et al. Genome-edited powdery mildew resistance in wheat without growth penalties. Nature 2022, 602, 455–460. [Google Scholar] [CrossRef]
- Pieterse, C.M.; Van der Does, D.; Zamioudis, C.; Leon-Reyes, A.; Van Wees, S.C. Hormonal modulation of plant immunity. Annu. Rev. Cell Dev. Biol. 2012, 28, 489–521. [Google Scholar] [CrossRef]
- van der Burgh, A.M.; Joosten, M. Plant immunity: Thinking outside and inside the box. Trends Plant Sci. 2019, 24, 587–601. [Google Scholar] [CrossRef]
- Romero-Puertas, M.C.; Terrón-Camero, L.C.; Peláez-Vico, M.Á.; Molina-Moya, E.; Sandalio, L.M. An update on redox signals in plant responses to biotic and abiotic stress crosstalk: Insights from cadmium and fungal pathogen interactions. J. Exp. Bot. 2021, 72, 5857–5875. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.; Zheng, S.; Jiang, D.; Lu, J.; Huang, Z.; Liu, Z.; Zhou, H.; Zhuang, C.; Li, J. Initiation and execution of programmed cell death and regulation of reactive oxygen species in plants. Int. J. Mol. Sci. 2021, 22, 12942. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Liu, W.; Zhang, Q.; Wang, X.; Wang, X.; Duan, X.; Li, F.; Huang, L.; Kang, Z. TaMDHAR4, a monodehydroascorbate reductase gene participates in the interactions between wheat and Puccinia striiformis f. sp. tritici. Plant Physiol. Biochem. 2014, 76, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Han, L.; Huai, B.; Zheng, P.; Chang, Q.; Guan, T.; Li, D.; Huang, L.; Kang, Z. Down-regulation of a wheat alkaline/neutral invertase correlates with reduced host susceptibility to wheat stripe rust caused by Puccinia striiformis. J. Exp. Bot. 2015, 66, 7325–7338. [Google Scholar] [CrossRef]
- Xu, Q.; Tang, C.; Wang, X.; Sun, S.; Zhao, J.; Kang, Z.; Wang, X. An effector protein of the wheat stripe rust fungus targets chloroplasts and suppresses chloroplast function. Nat. Commun. 2019, 10, 5571. [Google Scholar] [CrossRef]
- Yang, Q.; Huai, B.; Lu, Y.; Cai, K.; Guo, J.; Zhu, X.; Kang, Z.; Guo, J. A stripe rust effector Pst18363 targets and stabilises TaNUDX23 that promotes stripe rust disease. New Phytol. 2020, 225, 880–895. [Google Scholar] [CrossRef]
- Guo, S.; Zhang, Y.; Li, M.; Zeng, P.; Zhang, Q.; Li, X.; Xu, Q.; Li, T.; Wang, X.; Kang, Z.; et al. TaBln1, a member of the Blufensin family, negatively regulates wheat resistance to stripe rust by reducing Ca2+ influx. Plant Physiol. 2022, 189, 1380–1396. [Google Scholar] [CrossRef]
- Hao, Y.; Wang, X.; Wang, K.; Li, H.; Duan, X.; Tang, C.; Kang, Z. TaMCA1, a regulator of cell death, is important for the interaction between wheat and Puccinia striiformis. Sci. Rep. 2016, 6, 26946. [Google Scholar] [CrossRef]
- Corredor-Moreno, P.; Minter, F.; Davey, P.E.; Wegel, E.; Kular, B.; Brett, P.; Lewis, C.M.; Morgan, Y.; Macías Pérez, L.A.; Korolev, A.V.; et al. The branched-chain amino acid aminotransferase TaBCAT1 modulates amino acid metabolism and positively regulates wheat rust susceptibility. Plant Cell 2021, 33, 1728–1747. [Google Scholar] [CrossRef]
- Frye, C.A.; Innes, R.W. An Arabidopsis mutant with enhanced resistance to powdery mildew. Plant Cell 1998, 10, 947–956. [Google Scholar] [CrossRef] [Green Version]
- Frye, C.A.; Tang, D.; Innes, R.W. Negative regulation of defense responses in plants by a conserved MAPKK kinase. Proc. Natl. Acad. Sci. USA 2001, 98, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Christiansen, K.M.; Innes, R.W. Regulation of plant disease resistance, stress responses, cell death, and ethylene signaling in Arabidopsis by the EDR1 protein kinase. Plant Physiol. 2005, 138, 1018–1026. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Nie, H.; Shen, Q.; Zhang, S.; Lukowitz, W.; Tang, D. EDR1 physically interacts with MKK4/MKK5 and negatively regulates a MAP kinase cascade to modulate plant innate immunity. PLoS Genet. 2014, 10, e1004389. [Google Scholar] [CrossRef] [PubMed]
- Serrano, I.; Gu, Y.; Qi, D.; Dubiella, U.; Innes, R.W. The Arabidopsis EDR1 protein kinase negatively regulates the ATL1 E3 ubiquitin ligase to suppress cell death. Plant Cell 2014, 26, 4532–4546. [Google Scholar] [CrossRef]
- Zhang, Y.; Bai, Y.; Wu, G.; Zou, S.; Chen, Y.; Gao, C.; Tang, D. Simultaneous modification of three homoeologs of TaEDR1 by genome editing enhances powdery mildew resistance in wheat. Plant J. 2017, 91, 714–724. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, X.; Giroux, M.J.; Huang, L. A wheat COP9 subunit 5-like gene is negatively involved in host response to leaf rust. Mol. Plant Pathol. 2017, 18, 125–133. [Google Scholar] [CrossRef]
- Yang, Q.; Islam, M.A.; Cai, K.; Tian, S.; Liu, Y.; Kang, Z.; Guo, J. TaClpS1, negatively regulates wheat resistance against Puccinia striiformis f. sp. tritici. BMC Plant Biol. 2020, 20, 555. [Google Scholar] [CrossRef]
- Shi, B.; Wang, J.; Gao, H.; Yang, Q.; Wang, Y.; Day, B.; Ma, Q. The small GTP-binding protein TaRop10 interacts with TaTrxh9 and functions as a negative regulator of wheat resistance against the stripe rust. Plant Sci. 2021, 309, 110937. [Google Scholar] [CrossRef]
- Adachi, H.; Tsuda, K. Convergence of cell-surface and intracellular immune receptor signalling. New Phytol. 2019, 221, 1676–1678. [Google Scholar] [CrossRef]
- Duan, X.; Wang, X.; Fu, Y.; Tang, C.; Li, X.; Cheng, Y.; Feng, H.; Huang, L.; Kang, Z. TaEIL1, a wheat homologue of AtEIN3, acts as a negative regulator in the wheat-stripe rust fungus interaction. Mol. Plant Pathol. 2013, 14, 728–739. [Google Scholar] [CrossRef]
- Feng, H.; Duan, X.; Zhang, Q.; Li, X.; Wang, B.; Huang, L.; Wang, X.; Kang, Z. The target gene of tae-miR164, a novel NAC transcription factor from the NAM subfamily, negatively regulates resistance of wheat to stripe rust. Mol. Plant Pathol. 2014, 15, 284–296. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Wei, J.; Song, N.; Wang, N.; Zhao, J.; Kang, Z. A novel wheat NAC transcription factor, TaNAC30, negatively regulates resistance of wheat to stripe rust. J. Integr. Plant Biol. 2018, 60, 432–443. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, T.; Jia, J.; Sun, J. The wheat mediator subunit TaMED25 interacts with the transcription factor TaEIL1 to negatively regulate disease resistance against powdery mildew. Plant Physiol. 2016, 170, 1799–1816. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Tang, C.; Fan, X.; He, M.; Gan, P.; Zhang, S.; Hu, Z.; Wang, X.; Yan, T.; Shu, W.; et al. Inactivation of a wheat protein kinase gene confers broad-spectrum resistance to rust fungi. Cell 2022, 185, 2961–2974.e19. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Fan, X.; He, M.; Hu, Z.; Tang, C.; Zhang, S.; Lin, D.; Gan, P.; Wang, J.; Huang, X.; et al. Transcriptional repression of TaNOX10 by TaWRKY19 compromises ROS generation and enhances wheat susceptibility to stripe rust. Plant Cell 2022, 34, 1784–1803. [Google Scholar] [CrossRef]
- Ramirez-Prado, J.S.; Abulfaraj, A.A.; Rayapuram, N.; Benhamed, M.; Hirt, H. Plant immunity: From signaling to epigenetic control of defense. Trends Plant Sci. 2018, 23, 833–844. [Google Scholar] [CrossRef]
- Zhi, P.; Chang, C. Exploiting epigenetic variations for crop disease resistance improvement. Front. Plant Sci. 2021, 12, 692328. [Google Scholar] [CrossRef]
- Yang, Z.; Zhi, P.; Chang, C. Priming seeds for the future: Plant immune memory and application in crop protection. Front. Plant Sci. 2022, 13, 961840. [Google Scholar] [CrossRef]
- Liu, J.; Zhi, P.; Wang, X.; Fan, Q.; Chang, C. Wheat WD40-repeat protein TaHOS15 functions in a histone deacetylase complex to fine-tune defense responses to Blumeria graminis f. sp. tritici. J. Exp. Bot. 2019, 70, 255–268. [Google Scholar] [CrossRef]
- Zhi, P.; Kong, L.; Liu, J.; Zhang, X.; Wang, X.; Li, H.; Sun, M.; Li, Y.; Chang, C. Histone deacetylase TaHDT701 functions in TaHDA6-TaHOS15 complex to regulate wheat defense responses to Blumeria graminis f. sp. tritici. Int. J. Mol. Sci. 2020, 21, 2640. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Zhao, J.; Duan, W.; Tian, S.; Wang, X.; Zhuang, H.; Fu, J.; Kang, Z. TaAMT2;3a, a wheat AMT2-type ammonium transporter, facilitates the infection of stripe rust fungus on wheat. BMC Plant Biol. 2019, 19, 239. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.W.; Herrera-Foessel, S.; Lan, C.; Schnippenkoetter, W.; Ayliffe, M.; Huerta-Espino, J.; Lillemo, M.; Viccars, L.; Milne, R.; Periyannan, S.; et al. A recently evolved hexose transporter variant confers resistance to multiple pathogens in wheat. Nat. Genet. 2015, 47, 1494–1498. [Google Scholar] [CrossRef] [PubMed]
- Huai, B.; Yang, Q.; Qian, Y.; Qian, W.; Kang, Z.; Liu, J. ABA-induced sugar transporter TaSTP6 promotes wheat susceptibility to stripe rust. Plant Physiol. 2019, 181, 1328–1343. [Google Scholar] [CrossRef] [PubMed]
- Huai, B.; Yang, Q.; Wei, X.; Pan, Q.; Kang, Z.; Liu, J. TaSTP13 contributes to wheat susceptibility to stripe rust possibly by increasing cytoplasmic hexose concentration. BMC Plant Biol. 2020, 20, 49. [Google Scholar] [CrossRef]
- Huai, B.; Yuan, P.; Ma, X.; Zhang, X.; Jiang, L.; Zheng, P.; Yao, M.; Chen, Z.; Chen, L.; Shen, Q.; et al. Sugar transporter TaSTP3 activation by TaWRKY19/61/82 enhances stripe rust susceptibility in wheat. New Phytol. 2022; in press. [Google Scholar] [CrossRef]
- Liu, X.; Khajuria, C.; Li, J.; Trick, H.N.; Huang, L.; Gill, B.S.; Reeck, G.R.; Antony, G.; White, F.F.; Chen, M.S. Wheat Mds-1 encodes a heat-shock protein and governs susceptibility towards the Hessian fly gall midge. Nat. Commun. 2013, 4, 2070. [Google Scholar] [CrossRef]
- Anderson, V.A.; Haley, S.D.; Peairs, F.B.; van Eck, L.; Leach, J.E.; Lapitan, N.L. Virus-induced gene silencing suggests (1,3;1,4)-β-glucanase is a susceptibility factor in the compatible russian wheat aphid-wheat interaction. Mol. Plant Microbe Interact. 2014, 27, 913–922. [Google Scholar] [CrossRef]
- Ceulemans, E.; Ibrahim, H.; De Coninck, B.; Goossens, A. Pathogen effectors: Exploiting the promiscuity of plant signaling hubs. Trends Plant Sci. 2021, 26, 780–795. [Google Scholar] [CrossRef]
- Zhang, S.; Li, C.; Si, J.; Han, Z.; Chen, D. Action mechanisms of effectors in plant-pathogen interaction. Int. J. Mol. Sci. 2022, 23, 6758. [Google Scholar] [CrossRef]
- Liu, X.; Ao, K.; Yao, J.; Zhang, Y.; Li, X. Engineering plant disease resistance against biotrophic pathogens. Curr. Opin. Plant Biol. 2021, 60, 101987. [Google Scholar] [CrossRef]
- Bigini, V.; Camerlengo, F.; Botticella, E.; Sestili, F.; Savatin, D.V. Biotechnological resources to increase disease-resistance by improving plant immunity: A sustainable approach to save cereal crop production. Plants 2021, 10, 1146. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Wang, B.; Yu, J.; Dou, D. Pathogen-informed breeding for crop disease resistance. J. Integr. Plant Biol. 2021, 63, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Wang, Y.; Zhang, R.; Zhang, H.; Gao, C. CRISPR/Cas genome editing and precision plant breeding in agriculture. Annu. Rev. Plant Biol. 2019, 70, 667–697. [Google Scholar] [CrossRef] [PubMed]
- Manghwar, H.; Lindsey, K.; Zhang, X.; Jin, S. CRISPR/Cas system: Recent advances and future prospects for genome editing. Trends Plant Sci. 2019, 24, 1102–1125. [Google Scholar] [CrossRef] [PubMed]
- Yin, K.; Qiu, J.L. Genome editing for plant disease resistance: Applications and perspectives. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2019, 374, 20180322. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Li, C.; Gao, C. Applications of CRISPR-Cas in agriculture and plant biotechnology. Nat. Rev. Mol. Cell Biol. 2020, 21, 661–677. [Google Scholar] [CrossRef]
- Schenke, D.; Cai, D. Applications of CRISPR/Cas to improve crop disease resistance: Beyond inactivation of susceptibility factors. iScience 2020, 23, 101478. [Google Scholar] [CrossRef]
- Gao, C. Genome engineering for crop improvement and future agriculture. Cell 2021, 184, 1621–1635. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Shi, L.; Liang, X.; Zhao, P.; Wang, W.; Liu, J.; Chang, Y.; Hiei, Y.; Yanagihara, C.; Du, L.; et al. The gene TaWOX5 overcomes genotype dependency in wheat genetic transformation. Nat. Plants 2022, 8, 110–117. [Google Scholar] [CrossRef]
- Debernardi, J.M.; Tricoli, D.M.; Ercoli, M.F.; Hayta, S.; Ronald, P.; Palatnik, J.F.; Dubcovsky, J. A GRF-GIF chimeric protein improves the regeneration efficiency of transgenic plants. Nat. Biotechnol. 2020, 38, 1274–1279. [Google Scholar] [CrossRef]
- Li, T.; Hu, J.; Sun, Y.; Li, B.; Zhang, D.; Li, W.; Liu, J.; Li, D.; Gao, C.; Zhang, Y.; et al. Highly efficient heritable genome editing in wheat using an RNA virus and bypassing tissue culture. Mol. Plant 2021, 14, 1787–1798. [Google Scholar] [CrossRef] [PubMed]
- McCallum, C.M.; Comai, L.; Greene, E.A.; Henikoff, S. Targeting induced local lesions IN genomes (TILLING) for plant functional genomics. Plant Physiol. 2000, 123, 439–442. [Google Scholar] [CrossRef] [PubMed]
- Kurowska, M.; Daszkowska-Golec, A.; Gruszka, D.; Marzec, M.; Szurman, M.; Szarejko, I.; Maluszynski, M. TILLING: A shortcut in functional genomics. J. Appl. Genet. 2011, 52, 371–390. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Hao, L.; Parry, M.A.; Phillips, A.L.; Hu, Y.G. Progress in TILLING as a tool for functional genomics and improvement of crops. J. Integr. Plant Biol. 2014, 56, 425–443. [Google Scholar] [CrossRef] [PubMed]
- Varshney, R.K.; Sinha, P.; Singh, V.K.; Kumar, A.; Zhang, Q.; Bennetzen, J.L. 5Gs for crop genetic improvement. Curr. Opin. Plant Biol. 2020, 56, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Tian, D.; Zhang, Z.; Hu, S.; Yu, J. Rice genomics: Over the past two decades and into the future. Genom. Proteom. Bioinform. 2018, 16, 397–404. [Google Scholar] [CrossRef]
- Bevan, M.W.; Uauy, C.; Wulff, B.B.; Zhou, J.; Krasileva, K.; Clark, M.D. Genomic innovation for crop improvement. Nature 2017, 543, 346–354. [Google Scholar] [CrossRef]
- Arif, M.A.R.; Waheed, M.Q.; Lohwasser, U.; Shokat, S.; Alqudah, A.M.; Volkmar, C.; Börner, A. Genetic insight into the insect resistance in bread wheat exploiting the untapped natural diversity. Front. Genet. 2022, 13, 898905. [Google Scholar] [CrossRef]
- Jia, H.; Zhang, Y.; Orbović, V.; Xu, J.; White, F.F.; Jones, J.B.; Wang, N. Genome editing of the disease susceptibility gene CsLOB1 in citrus confers resistance to citrus canker. Plant Biotechnol. J. 2017, 15, 817–823. [Google Scholar] [CrossRef]
- Zhang, J.; Huguet-Tapia, J.C.; Hu, Y.; Jones, J.; Wang, N.; Liu, S.; White, F.F. Homologues of CsLOB1 in citrus function as disease susceptibility genes in citrus canker. Mol. Plant Pathol. 2017, 18, 798–810. [Google Scholar] [CrossRef]
- Li, P.; Zhang, L.; Mo, X.; Ji, H.; Bian, H.; Hu, Y.; Majid, T.; Long, J.; Pang, H.; Tao, Y.; et al. Rice aquaporin PIP1;3 and harpin Hpa1 of bacterial blight pathogen cooperate in a type III effector translocation. J. Exp. Bot. 2019, 70, 3057–3073. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, L.; Ji, H.; Mo, X.; Li, P.; Wang, J.; Dong, H. Hpa1 is a type III translocator in Xanthomonas oryzae pv. oryzae. BMC Microbiol. 2018, 18, 105. [Google Scholar] [CrossRef] [PubMed]
- Miedaner, T.; Juroszek, P. Climate change will influence disease resistance breeding in wheat in Northwestern Europe. Theor. Appl. Genet. 2021, 134, 1771–1785. [Google Scholar] [CrossRef]
- Desaint, H.; Aoun, N.; Deslandes, L.; Vailleau, F.; Roux, F.; Berthomé, R. Fight hard or die trying: When plants face pathogens under heat stress. New Phytol. 2021, 229, 712–734. [Google Scholar] [CrossRef] [PubMed]
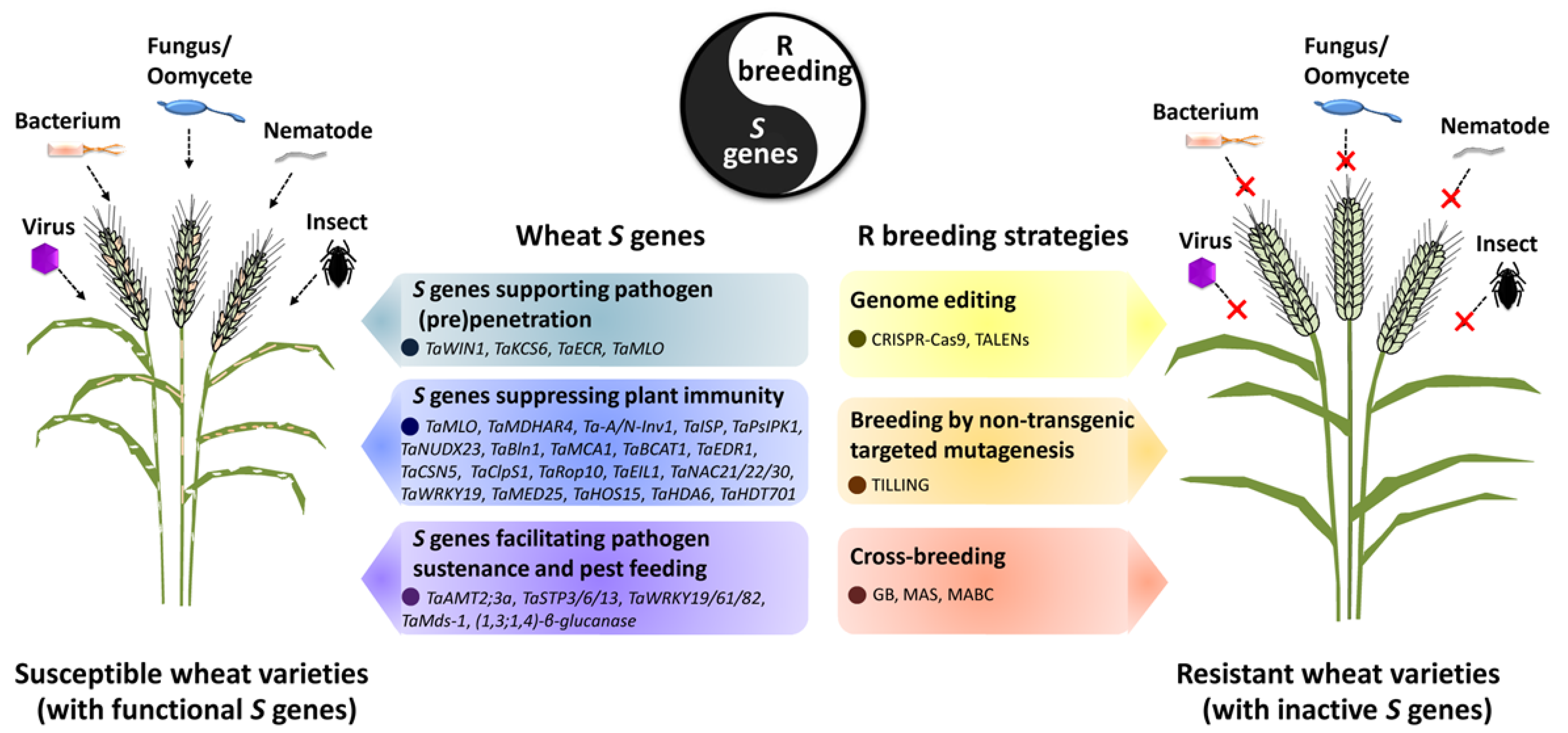

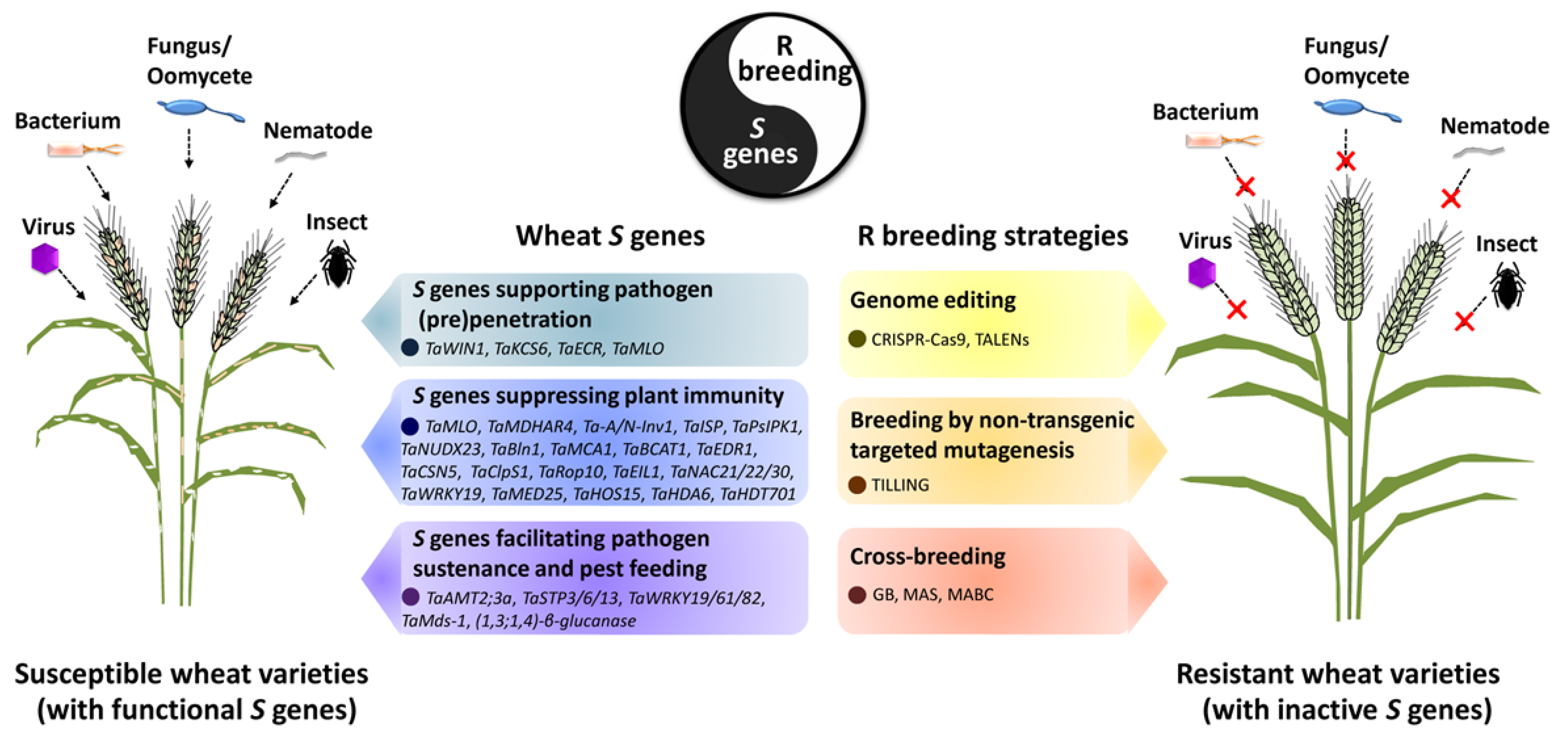{kind=link}
| Class | Wheat S Gene | S Gene Product Family | Pathogen/Pest Species | Contributions of Wheat S Genes to P&P Infections and Evidence | Application of S Genes in Resistance Breeding | Effector Targets | Reference |
|---|---|---|---|---|---|---|---|
| 1 | TaWIN1 | AP2-EREBP-type transcription factor | Blumeria graminis f. sp. tritici (Bgt) | Silencing of TaWIN1 by VIGS results in attenuated Bgt conidial germination. | None reported | None reported | [16] |
| 1 | TaKCS6 | 3-Ketoacyl-CoA synthase | Bgt | Silencing of TaKCS6 by VIGS leads to reduced Bgt conidial germination. | None reported | None reported | [17] |
| 1 | TaECR | Enoyl-CoA reductase | Bgt | Silencing of TaECR by VIGS results in decreased Bgt conidial germination. | None reported | None reported | [18] |
| 1, 2 | TaMLO | Integral membrane protein | Bgt | Knockout TaMLO by TILLING enhances wheat penetration and post-penetration resistance to Bgt. | Wheat Tamlo-R32 mutant generated by genome editing confers Bgt resistance without yield penalty. | None reported | [25,26,27,28] |
| 2 | TaMDHAR4 | Monodehydroascorbate reductase | Puccinia striiformis f. sp. Tritici (Pst) | Silencing of TaMDHAR4 by VIGS attenuates wheat susceptibility to Pst infection | None reported | None reported | [33] |
| 2 | Ta-A/N-Inv1 | Alkaline/neutral invertase | Pst | VIGS of Ta-A/N-Inv1 results in the wheat H2O2 over- accumulation, enhanced cell death, and reduced susceptibility to Pst infection | None reported | None reported | [34] |
| 2 | TaISP | Cytochrome b6-f component | Pst | Silencing of TaISP by VIGS reduces wheat photosynthesis and susceptibility to Pst. | None reported | Pst_12806 | [35] |
| 2 | TaNUDX23 | Nudix hydrolase | Pst | Knocking down of TaNUDX23 expression by VIGS attenuates Pst infection. | None reported | Pst18363 | [36] |
| 2 | TaBln1 | Cysteine-rich peptide | Pst | Silencing of TaBln1 results in the enhanced Ca2+ influx and attenuated accessibility to Pst. | None reported | None reported | [37] |
| 2 | TaMCA1 | Metacaspase | Pst | Knockdown of TaMCA1 expression by VIGS enhances wheat resistance to Pst. | None reported | None reported | [38] |
| 2 | TaBCAT1 | Branched-chain amino acid (BCAA) aminotransferase | Pst | TaBCAT1-silenced wheat plants exhibit enhanced levels of BCAAs and SA, as well as attenuated susceptibility to Pst. | None reported | None reported | [39] |
| 2 | TaEDR1 | Raf-like mitogen- activated protein kinase kinasekinase (MAPKKK) | Bgt | Knockout of TaEDR1 by TALENs results in attenuated wheat susceptibility to Bgt. | Wheat Taedr1 mutant generated by TALENs displays enhanced Bgt resistance without Bgt-induced cell death and obvious growth penalty. | None reported | [45] |
| 2 | TaCSN5 | COP9 subunit 5-like protein | Pst | TaCSN5-silenced wheat plants exhibit reduced susceptibility to Pst. | None reported | None reported | [46] |
| 2 | TaClpS1 | Caseinolytic peptidase (Clp) protease | Pst | Knockdown of TaClpS1 expression via VIGS leads to the enhanced wheat resistance to Pst. | None reported | None reported | [47] |
| 2 | TaROP10 | Small GTP-binding protein | Pst | The TaRop10-silenced wheat plant exhibits enhanced resistance to Pst. | None reported | None reported | [48] |
| 2 | TaEIL1 | ETHYLENE INSENSITIVE 3 (EIN3) family transcription factor | Pst | Silencing of TaEIL1 via VIGS enhances disease resistance against stripe rust. | None reported | None reported | [50] |
| 2 | TaNAC21/22/30 | NAC transcription factor | Pst | Silencing of TaNAC21, TaNAC22, and TaNAC30 attenuates wheat susceptibility to stripe rust. | None reported | None reported | [51,52] |
| 2 | TaMED25 | Mediator subunit | Bgt | Silencing of TaMED25 by VIGS enhances wheat resistance to Bgt. | None reported | None reported | [53] |
| 2 | TaPsIPK1 | Receptor-like cytoplasmic kinase | Pst | Overexpression of TaPsIPK1 enhances wheat susceptibility to Pst infection, but the silencing of TaPsIPK1 attenuates wheat susceptibility. | Inactivation of TaPsIPK1 by genome editing confers wheat broad-spectrum resistance against Pst without yield penalty. | PsSpg1 | [54] |
| 2 | TaWRKY19 | WRKY transcription factor | Pst | TaWRKY19-silenced or TaWRKY19-knockout wheat plants exhibit enhanced stripe rust resistance. | None reported | None reported | [55] |
| 2 | TaHOS15 | WD40-repeat protein | Bgt | Overexpression of TaHOS15 enhances wheat susceptibility to Bgt infection, but the silencing of TaHOS15 attenuates wheat susceptibility. | None reported | None reported | [59] |
| 2 | TaHDA6 | RPD3-type histone deacetylase | Bgt | Overexpression of TaHDA6 attenuates wheat powdery mildew resistance, but the silencing of TaHDA6 enhances wheat resistance. | None reported | None reported | [59] |
| 2 | TaHDT701 | HD2-type histone deacetylase | Bgt | Overexpression of TaHDT701 enhances wheat susceptibility to Bgt infection, but the silencing of TaHDT701 attenuates wheat susceptibility. | None reported | None reported | [60] |
| 3 | TaAMT2;3a | NH4+ transporter | Pst | Impeded Pst growth is observed in the TaAMT2;3a-silenced wheat leaves. | None reported | None reported | [61] |
| 3 | TaSTP3/6/13 | Sugartransporter | Pst | Silencing of TaSTP3, TaSTP6, and TaSTP13 by VIGS reduces wheat susceptibility to Pst. | None reported | None reported | [62,63,64,65] |
| 3 | TaWRKY19/61/82 | WRKY transcription factor | Pst | Pst growth is impeded in the TaWRKY19/61/82-silenced wheat leaves. | None reported | None reported | [65] |
| 4 | TaMds-1 | Small heatshock protein | Hessian fly | Silencing of TaMds-1 attenuates Hessian fly infestation. | None reported | None reported | [66] |
| 4 | (1,3;1,4)-β-glucanase | Glucanase | Russian wheat aphid (RWA) | Aphid reproduction is reduced in the (1,3;1,4)-β-glucanase-silenced wheat plants. | None reported | None reported | [67] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, M.; Yang, Z.; Chang, C. Susceptibility Is New Resistance: Wheat Susceptibility Genes and Exploitation in Resistance Breeding. Agriculture 2022, 12, 1419. https://doi.org/10.3390/agriculture12091419
Li M, Yang Z, Chang C. Susceptibility Is New Resistance: Wheat Susceptibility Genes and Exploitation in Resistance Breeding. Agriculture. 2022; 12(9):1419. https://doi.org/10.3390/agriculture12091419
Chicago/Turabian StyleLi, Mengmeng, Zige Yang, and Cheng Chang. 2022. "Susceptibility Is New Resistance: Wheat Susceptibility Genes and Exploitation in Resistance Breeding" Agriculture 12, no. 9: 1419. https://doi.org/10.3390/agriculture12091419
APA StyleLi, M., Yang, Z., & Chang, C. (2022). Susceptibility Is New Resistance: Wheat Susceptibility Genes and Exploitation in Resistance Breeding. Agriculture, 12(9), 1419. https://doi.org/10.3390/agriculture12091419