
Sorghum as a Potential Valuable Aquafeed Ingredient: Nutritional Quality and Digestibility

 ,
, 
Abstract
1. Introduction
2. Sorghum Biology and Agriculture
- -
- Grain sorghums (Caffrotum group) are cultivated for their grain, similar to corn/maize, and used to make human foods and beverages and in animal feeding.
- -
- Sweet sorghums (Saccharatum group) are cultivated for their stalks and foliage, which are used to produce syrups, silage, and, increasingly, biofuels.
- -
- Grass sorghums (Sudan grass) are cultivated as animal fodder consumed directly as a pasture or hay crop or as silage.
- -
- Broom corn (Technicum group) are woody varieties cultivated for their fibrous structures used to make brooms and brushes [24].
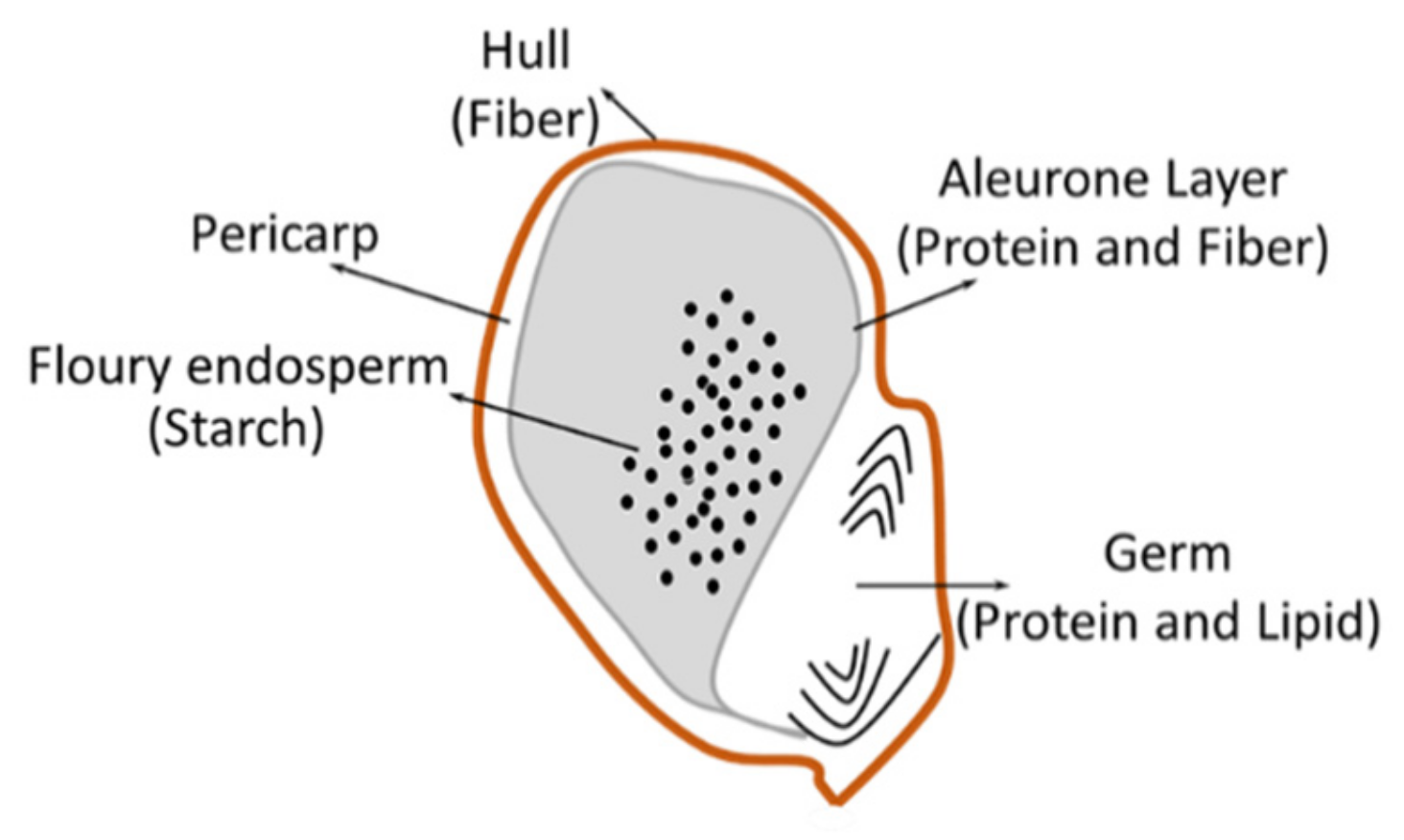
3. Nutrient Composition of Sorghum
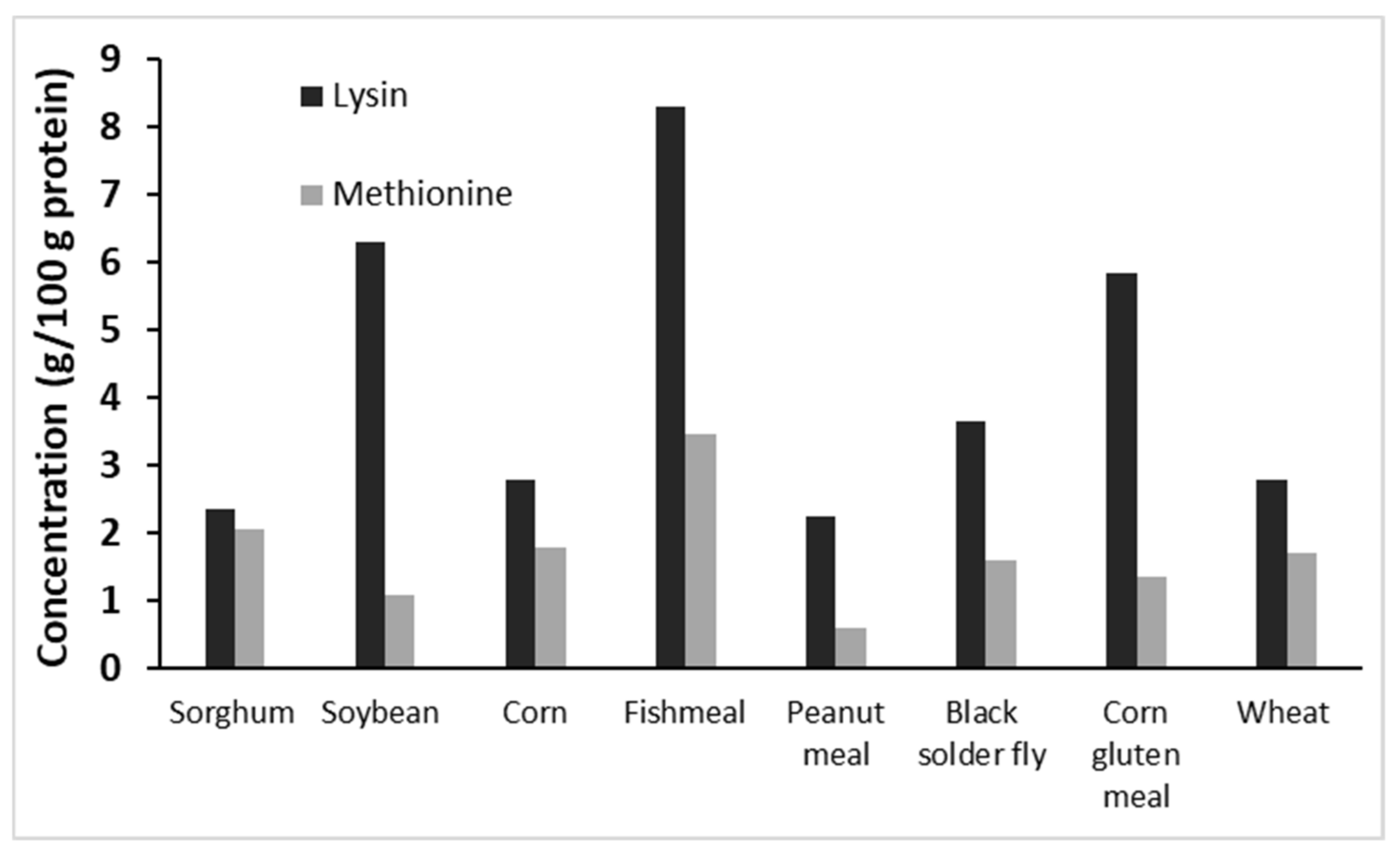
3.1. Protein Content and Amino Acids Compositions
3.2. Fat and Fatty Acids
3.3. Micronutrients
3.4. Fiber
3.5. Starch
3.6. Anti-Nutritional Factors (ANFs)
4. Sorghum in Animal Nutrition
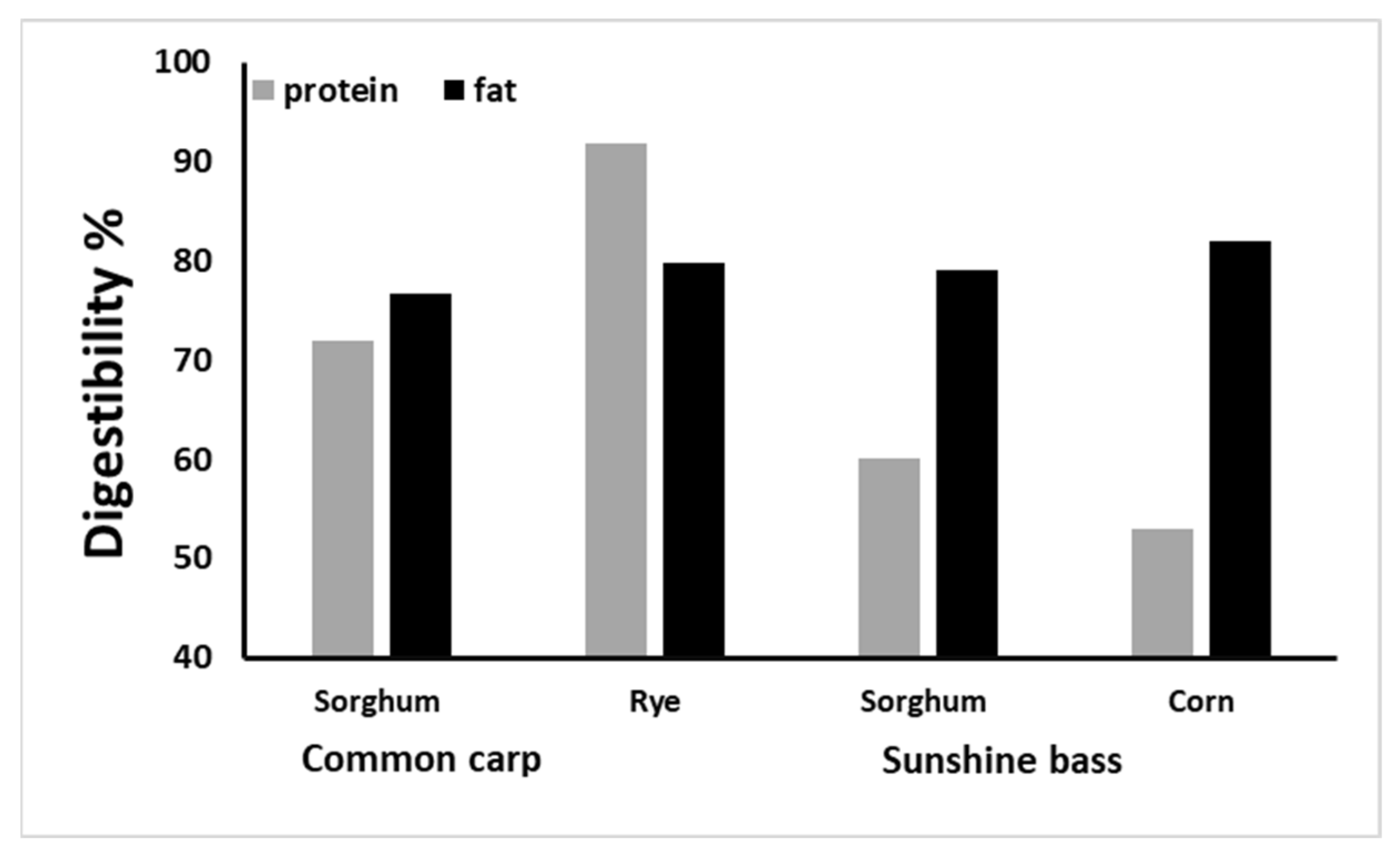
4.1. Aquatic Livestock
4.2. Supplementation Strategies for Improving Sorghum Performance in Aquafeed
4.3. Reducing the Impact of Anti-Nutritional Factors
5. Improving Sorghum Digestibility
5.1. Fermentation
5.2. Chemical Modification
5.3. Enzymatic Hydrolysis
5.4. Thermo-Mechanical Treatment
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. FAOSTAT Statistical Database; Food and Agriculture Organisation of the United Nations: Rome, Italy, 2020. [Google Scholar]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food security: The challenge of feeding 9 billion people. Science 2010, 327, 812–818. [Google Scholar] [CrossRef] [PubMed]
- Valin, H.; Sands, R.D.; Van der Mensbrugghe, D.; Nelson, G.C.; Ahammad, H.; Blanc, E.; Bodirsky, B.; Fujimori, S.; Hasegawa, T.; Havlik, P. The future of food demand: Understanding differences in global economic models. Agric. Econ. 2014, 45, 51–67. [Google Scholar] [CrossRef]
- Nyachoti, C.; Atkinson, J.; Leeson, S. Sorghum tannins: A review. World’s Poult. Sci. J. 1997, 53, 5–21. [Google Scholar] [CrossRef]
- Aderolu, A.Z.; Kuton, M.P.; Odu-Onikosi, S.G. Substitution effect of sorghum meal for maize meal in the diet of catfish (Clarias gariepinus, Burchell, 1822) juvenile. Res. J. Fis. Hyd 2009, 4, 41–45. [Google Scholar]
- Stamenković, O.S.; Siliveru, K.; Veljković, V.B.; Banković-Ilić, I.B.; Tasić, M.B.; Ciampitti, I.A.; Đalović, I.G.; Mitrović, P.M.; Sikora, V.Š.; Prasad, P.V. Production of biofuels from sorghum. Renew. Sustain. Energy Rev. 2020, 124, 109769. [Google Scholar] [CrossRef]
- Bean, S.; Wilson, J.; Moreau, R.; Galant, A.; Awika, J.; Kaufman, R.; Adrianos, S.; Ioerger, B. Structure and composition of the sorghum grain. In Sorghum: State of the Art and Future Perspectives; American Society of Agronomy: Madison, WI, USA, 2016. [Google Scholar]
- USDA-NASS. Crop Production, 2019 Summary. Available online: https://downloads.usda.library.cornell.edu/usda-esmis/files/k3569432s/sj139j59z/1257b842j/cropan20.pdf (accessed on 13 May 2021).
- Jiddere, G.; Filli, K.B. The effect of feed moisture and barrel temperature on the essential amino acids profile of sorghum malt and bambara groundnut based extrudates. J. Food Processing Technol. 2015, 6, 1000448. [Google Scholar]
- Obizoba, I.C.; Atii, J. Effect of soaking, sprouting, fermentation and cooking on nutrient composition and some anti-nutritional factors of sorghum (Guinesia) seeds. Plant Foods Hum. Nutr. 1991, 41, 203–212. [Google Scholar] [CrossRef]
- Mohammed, N.A.; Ahmed, I.A.M.; Babiker, E.E. Nutritional evaluation of sorghum flour (Sorghum bicolor L. Moench) during processing of injera. World Acad. Sci. Eng. Technol. 2011, 51, 99–103. [Google Scholar]
- Aruna, C.; Visarada, K.; Bhat, B.V.; Tonapi, V.A. Breeding Sorghum for Diverse end Uses; Woodhead Publishing: Cambridge, MA, USA, 2018. [Google Scholar]
- Wang, L.; Weller, C.L.; Schlegel, V.L.; Carr, T.P.; Cuppett, S.L. Supercritical CO2 extraction of lipids from grain sorghum dried distillers grains with solubles. Bioresour. Technol. 2008, 99, 1373–1382. [Google Scholar] [CrossRef]
- Van Amerongen, A. Sorghum Surges. Available online: http://ethanolproducer.com/articles/6338/sorghum-surges (accessed on 13 May 2021).
- Cui, X.; Kavvada, O.; Huntington, T.; Scown, C.D. Strategies for near-term scale-up of cellulosic biofuel production using sorghum and crop residues in the US. Environ. Res. Lett. 2018, 13, 124002. [Google Scholar] [CrossRef]
- Baral, N.R.; Dahlberg, J.; Putnam, D.; Mortimer, J.C.; Scown, C.D. Supply cost and life-cycle greenhouse gas footprint of dry and ensiled biomass sorghum for biofuel production. ACS Sustain. Chem. Eng. 2020, 8, 15855–15864. [Google Scholar] [CrossRef]
- Hepher, B. Nutrition of pond fishes; Cambridge University Press: Cambridge, UK, 1988. [Google Scholar]
- Al-Ogaily, S.; Al-Asgah, N.; Ali, A. Effect of feeding different grain sources on the growth performance and body composition of tilapia, Oreochromis niloticus (L.). Aquac. Res. 1996, 27, 523–529. [Google Scholar] [CrossRef]
- Alavi, S.; Mazumdar, S.D.; Taylor, J.R. Modern convenient sorghum and millet food, beverage and animal feed products, and their technologies. In Sorghum and Millets; Elsevier: Amsterdam, The Netherlands, 2019; pp. 293–329. [Google Scholar]
- Froehlich, H.E.; Runge, C.A.; Gentry, R.R.; Gaines, S.D.; Halpern, B.S. Comparative terrestrial feed and land use of an aquaculture-dominant world. Proc. Natl. Acad. Sci. USA 2018, 115, 5295–5300. [Google Scholar] [CrossRef]
- Kraugerud, O.F.; Jørgensen, H.Y.; Svihus, B. Physical properties of extruded fish feed with inclusion of different plant (legumes, oilseeds, or cereals) meals. Anim. Feed Sci. Technol. 2011, 163, 244–254. [Google Scholar] [CrossRef]
- Lemlioglu-Austin, D. Sorghum: Obliging alternative and ancient grain. Cereal Foods World 2014, 59, 12–20. [Google Scholar] [CrossRef]
- Damodaran, S. Amino acids, peptides and proteins. Fennema’s Food Chem. 2008, 4, 425–439. [Google Scholar]
- Thain, M.; Hickman, M. Penguin Dictionary of Biology; Penguin Books: New York, NY, USA, 2004. [Google Scholar]
- Hahn, D.; Rooney, L. Effect of genotype on tannins and phenols of sorghum. Cereal Chem 1986, 63, 4–8. [Google Scholar]
- Price, M.L.; Van Scoyoc, S.; Butler, L.G. A critical evaluation of the vanillin reaction as an assay for tannin in sorghum grain. J. Agric. Food Chem. 1978, 26, 1214–1218. [Google Scholar] [CrossRef]
- Khoddami, A.; Messina, V.; Vadabalija Venkata, K.; Farahnaky, A.; Blanchard, C.L.; Roberts, T.H. Sorghum in foods: Functionality and potential in innovative products. Crit. Rev. Food Sci. Nutr. 2021, 1–17. [Google Scholar] [CrossRef]
- Tuinstra, M.R. Food-grade sorghum varieties and production considerations: A review. J. Plant Interact. 2008, 3, 69–72. [Google Scholar] [CrossRef]
- Stefoska-Needham, A.; Beck, E.J.; Johnson, S.K.; Tapsell, L.C. Sorghum: An underutilized cereal whole grain with the potential to assist in the prevention of chronic disease. Food Rev. Int. 2015, 31, 401–437. [Google Scholar] [CrossRef]
- Xiong, Y.; Zhang, P.; Warner, R.D.; Fang, Z. Sorghum grain: From genotype, nutrition, and phenolic profile to its health benefits and food applications. Compr. Rev. Food Sci. Food Saf. 2019, 18, 2025–2046. [Google Scholar] [CrossRef]
- Wall, J.; Blessin, C. Composition and structure of sorghum grains. Cereal Sci. Today 1969, 14, 264–268. [Google Scholar]
- Taylor, J.; Duodu, K.G. Sorghum and Millets: Chemistry, Technology, and Nutritional Attributes; Elsevier: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Gassem, M.A.; Osman, M.A. Proximate composition and the content of sugars, amino acids and anti-nutritional factors of three sorghum varieties. Agric. Res. Cent. King Saud Univ. Res. Bull. 2003, 125, 5–19. [Google Scholar]
- Pontieri, P.; Troisi, J.; Calcagnile, M.; Bean, S.R.; Tilley, M.; Aramouni, F.; Boffa, A.; Pepe, G.; Campiglia, P.; Del Giudice, F. Chemical composition, fatty acid and mineral content of food-grade white, red and black sorghum varieties grown in the mediterranean environment. Foods 2022, 11, 436. [Google Scholar] [CrossRef]
- Udachan, I.S.; Sahoo, A.; Hend, G. Extraction and characterization of sorghum (Sorghum bicolor L. Moench) starch. Int. Food Res. J. 2012, 19, 315–319. [Google Scholar]
- Douglas, J.; Sullivan, T.; Bond, P.; Struwe, F. Nutrient composition and metabolizable energy values of selected grain sorghum varieties and yellow corn. Poult. Sci. 1990, 69, 1147–1155. [Google Scholar] [CrossRef]
- Miller, D. Minerals. In Food Chemistry; Fennema Marcel Dekker: New York, NY, USA, 1996; pp. 617–649. [Google Scholar]
- Wong, J.H.; Marx, D.B.; Wilson, J.D.; Buchanan, B.B.; Lemaux, P.G.; Pedersen, J.F. Principal component analysis and biochemical characterization of protein and starch reveal primary targets for improving sorghum grain. Plant Sci. 2010, 179, 598–611. [Google Scholar] [CrossRef]
- Murty, D.; Renard, C. Sorghum. Crops in Tropical Africa; Raemaekers, R.H., Ed.; Directorate General for International Cooperation. Ministry of Foreign Affairs, External Trade and International Cooperation: Brussels, Belgium, 2001; pp. 68–96. [Google Scholar]
- Virupaksha, T.; Sastry, L. Protein content and amino acid composition of some varieties of grain sorghum. J. Agric. Food Chem. 1968, 16, 199–203. [Google Scholar] [CrossRef]
- Waggle, D.; Deyoe, C. Relationship between protein level and amino acid composition of sorghum grain. Feedstuffs 1966, 38, 18. [Google Scholar]
- Mossé, J.; Huet, J.; Baudet, J. The amino acid composition of wheat grain as a function of nitrogen content. J. Cereal Sci. 1985, 3, 115–130. [Google Scholar] [CrossRef]
- Portz, L.; Cyrino, J.E.P. Digestibility of nutrients and amino acids of different protein sources in practical diets by largemouth bass Micropterus salmoides (Lacepéde, 1802). Aquac. Res. 2004, 35, 312–320. [Google Scholar] [CrossRef]
- Breuer, L.H., Jr.; Dohm, C.K. Comparative nutritive value of several sorghum grain varieties and hybrids. J. Agric. Food Chem. 1972, 20, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Sikka, K.C.; Johari, R.P. Comparative nutritive value and amino acid content of different varieties of sorghum and effect of lysine fortification. J. Agric. Food Chem. 1979, 27, 962–965. [Google Scholar] [CrossRef]
- Duodu, K.; Nunes, A.; Delgadillo, I.; Parker, M.; Mills, E.; Belton, P.; Taylor, J. Effect of grain structure and cooking on sorghum and maize in vitro protein digestibility. J. Cereal Sci. 2002, 35, 161–174. [Google Scholar] [CrossRef]
- Hamaker, B.R.; Kirleis, A.W.; Mertz, E.T.; Axtell, J.D. Effect of cooking on the protein profiles and in vitro digestibility of sorghum and maize. J. Agric. Food Chem. 1986, 34, 647–649. [Google Scholar] [CrossRef]
- De Cindio, B.; Gabriele, D.; Pollini, C.M.; Peressini, D.; Sensidoni, A. Filled snack production by co-extrusion-cooking: 2. Effect of processing on cereal mixtures. J. Food Eng. 2002, 54, 63–73. [Google Scholar] [CrossRef]
- Glennie, C. Endosperm cell wall modification in sorghum grain during germination. Cereal Chem. 1984, 61, 285–289. [Google Scholar]
- Knudsen, K.B.; Munck, L. Dietary fibre contents and compositions of sorghum and sorghum-based foods. J. Cereal Sci. 1985, 3, 153–164. [Google Scholar] [CrossRef]
- Gram, N.H. The ultrastructure of germinating barley seeds. II. Breakdown of starch granules and cell walls of the endosperm in three barley varieties. Carlsberg Res. Commun. 1982, 47, 173. [Google Scholar] [CrossRef]
- Shull, J.M.; Watterson, J.J.; Kirleis, A. Purification and immunocytochemical localization of kafirins in Sorghum bicolor (L. Moench) endosperm. Protoplasma 1992, 171, 64–74. [Google Scholar] [CrossRef]
- Shewry, P.R.; Tatham, A.S. The prolamin storage proteins of cereal seeds: Structure and evolution. Biochem. J. 1990, 267, 1–12. [Google Scholar] [CrossRef]
- Belton, P.; Delgadillo, I.; Halford, N.; Shewry, P. Kafirin structure and functionality. J. Cereal Sci. 2006, 44, 272–286. [Google Scholar] [CrossRef]
- Watterson, J.; Shull, J.; Kirleis, A. and Opaque Endosperm of Sorghum bicolor. Cereal Chem 1970, 70, 452–457. [Google Scholar]
- Dowling, L.; Arndt, C.; Hamaker, B. Economic viability of high digestibility sorghum as feed for market broilers. Agron. J. 2002, 94, 1050–1058. [Google Scholar] [CrossRef]
- Laidlaw, H.; Mace, E.; Williams, S.; Sakrewski, K.; Mudge, A.; Prentis, P.; Jordan, D.; Godwin, I. Allelic variation of the β-, γ-and δ-kafirin genes in diverse Sorghum genotypes. Theor. Appl. Genet. 2010, 121, 1227–1237. [Google Scholar] [CrossRef]
- Hamaker, B.; Kirleis, A.; Butler, L.; Axtell, J.; Mertz, E. Improving the in vitro protein digestibility of sorghum with reducing agents. Proc. Natl. Acad. Sci. 1987, 84, 626–628. [Google Scholar] [CrossRef]
- Da Silva, L.S.; Taylor, J.; Taylor, J.R. Transgenic sorghum with altered kafirin synthesis: Kafirin solubility, polymerization, and protein digestion. J. Agric. Food Chem. 2011, 59, 9265–9270. [Google Scholar] [CrossRef]
- Rooney, L. Processing methods to improve nutritional value of sorghum for livestock. In Proceedings of the Sorghum Nutritional Quality Conference, Purdue University, West Lafayette, ID, USA, 26 February–1 March 1990; pp. 206–210. [Google Scholar]
- Baldwin, A.; Sniegowski, M. Fatty acid compositions of lipids from corn and grain sorghum kernels. J. Am. Oil Chem. Soc. 1951, 28, 24–27. [Google Scholar] [CrossRef]
- Osman, R.; Abd El-Gelil, F.; El-Noamany, H.; Dawood, M.G. Oil content and fatty acid composition of some varieties of barley and sorghum grains. Grasas Y Aceites 2000, 51, 157–162. [Google Scholar] [CrossRef]
- Kamath, V.; Niketh, S.; Chandrashekar, A.; Rajini, P. Chymotryptic hydrolysates of α-kafirin, the storage protein of sorghum (Sorghum bicolor) exhibited angiotensin converting enzyme inhibitory activity. Food Chem. 2007, 100, 306–311. [Google Scholar] [CrossRef]
- Kulamarva, A.G.; Sosle, V.R.; Raghavan, G.V. Nutritional and rheological properties of sorghum. Int. J. Food Prop. 2009, 12, 55–69. [Google Scholar] [CrossRef]
- Jambunathan, R.; Subramanian, V. Grain quality and utilization of sorghum and pearl millet. In Proceedings of the International Biotechnology Workshop, ICRISAT Center, Hyderabad, India, 12–15 January 1987; pp. 133–139. [Google Scholar]
- Lodge, S.; Stock, R.; Klopfenstein, T.; Shain, D.; Herold, D. Evaluation of corn and sorghum distillers byproducts. J. Anim. Sci. 1997, 75, 37–43. [Google Scholar] [CrossRef]
- McNeill, J.; Potter, G.; Riggs, J.; Rooney, L. Chemical and physical properties of processed sorghum grain carbohydrates. J. Anim. Sci. 1975, 40, 335–341. [Google Scholar] [CrossRef]
- Hibberd, C.; Wagner, D.; Schemm, R.; Mitchell, E., Jr.; Weibel, D.; Hintz, R. Digestibility characteristics of isolated starch from sorghum and corn grain. J. Anim. Sci. 1982, 55, 1490–1497. [Google Scholar] [CrossRef]
- Dreher, M.L.; Dreher, C.J.; Berry, J.W.; Fleming, S.E. Starch digestibility of foods: A nutritional perspective. Crit. Rev. Food Sci. Nutr. 1984, 20, 47–71. [Google Scholar] [CrossRef]
- Davis, A.; Hoseney, R. Grain sorghum condensed tannins. I. Isolation, estimation, and selective adsorption by starch. Cereal Chem 1979, 56, 310–314. [Google Scholar]
- Atteh, J. Principles and Practice of Livestock Feed Manufacturing; ADLEK Printers: Ilorin, Nigeria, 2002; pp. 52–58. [Google Scholar]
- Doggett, S.W.; Green, J.P.; Cantril, S.T. Efficacy of radiation therapy alone for limited squamous cell carcinoma of the anal canal. Int. J. Radiat. Oncol. Biol. Phys. 1988, 15, 1069–1072. [Google Scholar] [CrossRef]
- Purseglove, J.W. Tropical Crops. Dicotyledons 1 and 2; Cambridge University Press: Cambridge, UK, 1968. [Google Scholar]
- Kumar, R. Anti-nutritional factors, the potential risks of toxicity and methods to alleviate them. In Legume Trees and Other Fodder Trees as Protein Sources for Livestock; FAO Corporate Document Repository: Kuala Lumpur, Malaysia, 1992; Volume 102, pp. 145–160. [Google Scholar]
- Myer, R.; Gorbet, D. Waxy and normal grain sorghums with varying tannin contents in diets for young pigs. Anim. Feed. Sci. Technol. 1985, 12, 179–186. [Google Scholar] [CrossRef]
- Nelson, T.S.; Stephenson, E.L.; Burgos, A.; Floyd, J.; York, J.O. Effect of tannin content and dry matter digestion on energy utilization and average amino acid availability of hybrid sorghum grains. Poult. Sci. 1975, 54, 1620–1623. [Google Scholar] [CrossRef]
- Dykes, L.; Rooney, L.W. Sorghum and millet phenols and antioxidants. J. Cereal Sci. 2006, 44, 236–251. [Google Scholar] [CrossRef]
- Dykes, L. Sorghum phytochemicals and their potential impact on human health. Sorghum 2019, 1931, 121–140. [Google Scholar]
- Treviño, J.; Ortiz, L.; Centeno, C. Effect of tannins from faba beans (Vicia faba) on the digestion of starch by growing chicks. Anim. Feed Sci. Technol. 1992, 37, 345–349. [Google Scholar] [CrossRef]
- Elkin, R.G.; Freed, M.B.; Hamaker, B.R.; Zhang, Y.; Parsons, C.M. Condensed tannins are only partially responsible for variations in nutrient digestibilities of sorghum grain cultivars. J. Agric. Food Chem. 1996, 44, 848–853. [Google Scholar] [CrossRef]
- Etuk, E.; Ifeduba, A.; Okata, U.; Chiaka, I.; Okoli, I.C.; Okeudo, N.; Esonu, B.; Udedibie, A.; Moreki, J. Nutrient composition and feeding value of sorghum for livestock and poultry: A review. J. Anim. Sci. Adv. 2012, 2, 510–524. [Google Scholar]
- Hancock, J.; Bramel-Cox, P. Use of sorghum grain for feeding livestock and poultry. In Expected Impacts of Sorghum Farm Program Target Price Policies and Other Factors Affecting Demand: Completion Report for the Grain Sorghum Federation; Texas Agricultural Experiment Station: College Station, TX, USA, 1992. [Google Scholar]
- Butler, L.G. Sorghum polyphenols. Toxic. Plant Orig. 1989, 4, 95–121. [Google Scholar]
- Awika, J.M.; Rooney, L.W. Sorghum phytochemicals and their potential impact on human health. Phytochemistry 2004, 65, 1199–1221. [Google Scholar] [CrossRef]
- Iren, L. Sorghum and millets, in cultivated plants, primarily as food sources. Encycl. Life Support Syst. 2004, 1–7. [Google Scholar]
- Mabelebele, M.; Gous, R.; O’Neil, H.M.; Iji, P. Whole sorghum inclusion and feed form on performance and nutrient digestibility of broiler chickens. J. Appl. Anim. Nutr. 2018, 6. [Google Scholar] [CrossRef]
- Carter, P. Spectrophotometric determination of serum iron at the submicrogram level with a new reagent (ferrozine). Anal. Biochem. 1971, 40, 450–458. [Google Scholar] [CrossRef]
- D’mello, J. Antinutritional factors and mycotoxins. In Farm Animal Metabolism and Nutrition; CABI Publishing: Wallingford, UK, 2000; pp. 383–403. [Google Scholar]
- Degani, G. Digestible energy in dietary sorghum, wheat bran, and rye in the common carp (Cyprinus carpio L.). Isr. J. Aquac. 2006, 58, 71–77. [Google Scholar] [CrossRef]
- Rawles, S.D.; Gatlin III, D.M. Nutrient digestibility of common feedstuffs in extruded diets for sunshine bass Morone chrysops♀×M. saxatilis♂. J. World Aquac. Soc. 2000, 31, 570–579. [Google Scholar] [CrossRef]
- González-Félix, M.L.; Davis, D.A.; Rossi, W., Jr.; Perez-Velazquez, M. Evaluation of apparent digestibility coefficient of energy of various vegetable feed ingredients in Florida pompano, Trachinotus carolinus. Aquaculture 2010, 310, 240–243. [Google Scholar] [CrossRef]
- Yones, A.; Metwalli, A.A. Dietary sorghum starch influences growth performance, apparent digestibility coefficient and some hepatic enzyme activities of carbohydrate metabolism in Hypridered tilapia (Oreochromis mossambicus × O. niloticus) fingerlings. Int. J. Fish. Aquac. Res. 2016, 1, 1–16. [Google Scholar]
- Bruce, T.; Harris, H.W. Expanding Utilization and Increasing Value of Sorghum Fractions via Microbial Conversion; United Sorghum Checkoff Program (USCP): Lubbock, TX, USA, 2017; pp. 1–36. [Google Scholar]
- Balogun, A.; Fagbenro, O. Use of macadamia presscake as a protein feedstuff in practical diets for tilapia, Oreochromis niloticus (L.). Aquac. Res. 1995, 26, 371–377. [Google Scholar] [CrossRef]
- US Grains Council. Sorghum as Source of Ingredient to Replace Cassava for Feeding Pangasius in Vietnam; US Grains Council: Washington, DC, USA, 2017. [Google Scholar]
- Signor, A.; Lewandowski, V.; Silva, R.A.d.; Fries, E.M.; Schuller, J.M. Effect of phytase on digestibility of corn, sorghum and wheat bran by silver catfish (Rhamdia voulezi). Acta Scientiarum. Anim. Sci. 2016, 38, 355–359. [Google Scholar] [CrossRef]
- Lins Rodrigues, M.; Souza dos Santos Sanchez, M.; Ernzen Pessini, J.; Weiler, K.A.; Deparis, A.; Boscolo, W.R.; Bittencourt, F.; Signor, A. Replacement of corn by sorghum and phytase supplementation in silver catfish (Rhamdia quelen) diets: Growth performance, physiological variables and bone mineralization. J. Appl. Anim. Res. 2020, 48, 142–150. [Google Scholar] [CrossRef]
- Zahler, J.D. Improving the Nutritional Characteristics of Plant Feedstuff By-Products Using Fungal Metabolism; South Dakota State University: Brookings, SD, USA, 2018. [Google Scholar]
- Hussein, E.E.S.; Dabrowski, K.; El-Saidy, D.M.; Lee, B.J. Enhancing the growth of Nile tilapia larvae/juveniles by replacing plant (gluten) protein with algae protein. Aquac. Res. 2013, 44, 937–949. [Google Scholar] [CrossRef]
- Eissa, A.E.; Attia, M.M.; Elgendy, M.Y.; Ismail, G.A.; Sabry, N.M.; Prince, A.; Mahmoud, M.A.; El-Demerdash, G.O.; Abdelsalam, M.; Derwa, H.I. Streptococcus, Centrocestus formosanus and Myxobolus tilapiae concurrent infections in farmed Nile tilapia (Oreochromis niloticus). Microb. Pathog. 2021, 158, 105084. [Google Scholar] [CrossRef]
- Cavalcante, R.B.; Telli, G.S.; Tachibana, L.; de Carla Dias, D.; Oshiro, E.; Natori, M.M.; da Silva, W.F.; Ranzani-Paiva, M.J. Probiotics, Prebiotics and Synbiotics for Nile tilapia: Growth performance and protection against Aeromonas hydrophila infection. Aquac. Rep. 2020, 17, 100343. [Google Scholar] [CrossRef]
- Urriola, P.; Hoehler, D.; Pedersen, C.; Stein, H.; Shurson, G. Amino acid digestibility of distillers dried grains with solubles, produced from sorghum, a sorghum-corn blend, and corn fed to growing pigs. J. Anim. Sci. 2009, 87, 2574–2580. [Google Scholar] [CrossRef]
- Barekatain, M.; Antipatis, C.; Choct, M.; Iji, P. Interaction between protease and xylanase in broiler chicken diets containing sorghum distillers’ dried grains with solubles. Anim. Feed Sci. Technol. 2013, 182, 71–81. [Google Scholar] [CrossRef]
- Adedeji, O.E.; Jegede, D.E.; Abdulsalam, K.O.; Umeohia, U.E.; Ajayi, O.A.; Iboyi, J.E. Effect of processing treatments on the proximate, functional and sensory properties of soy-sorghum-roselle complementary food. Br. J. Appl. Sci. Technol. 2015, 6, 635. [Google Scholar] [CrossRef]
- Li, M.; Robinson, E.; Oberle, D.; Lucas, P. Effects of various corn distillers by-products on growth, feed efficiency, and body composition of channel catfish, Ictalurus punctatus. Aquac. Nutr. 2010, 16, 188–193. [Google Scholar] [CrossRef]
- Cheng, Z.J.; Hardy, R.W. Effects of microbial phytase supplementation in corn distiller’s dried grain with solubles on nutrient digestibility and growth performance of rainbow trout, Oncorhynchus mykiss. J. Appl. Aquac. 2004, 15, 83–100. [Google Scholar] [CrossRef]
- Webster, C. Evaluation of distillers’ grains with solubles in prepared channel catfish diets. Trans. Ky. Acad. Sci. 1991, 51, 135–138. [Google Scholar]
- Lee, S.M.; Pan, B.S. Effect of dietary sorghum distillery residue on hematological characteristics of cultured grey mullet (Mugil cephalus)—An animal model for prescreening antioxidant and blood thinning activities. J. Food Biochem. 2003, 27, 1–18. [Google Scholar] [CrossRef]
- Furuya, W.M.; Furlan, A.C.; Rossetto, V.; Furuya, B. Sorghum a potential energy source in tilapia feed. Glob. Aquac. Advocate 2003, 1–4. [Google Scholar]
- Rodrigues, S.R.; Dalmoro, V.; dos Santos, J.H. An evaluation of Acacia mearnsii tannin as an aluminum corrosion inhibitor in acid, alkaline, and neutral media. Mater. Corros. 2020, 71, 1160–1174. [Google Scholar] [CrossRef]
- Osman, M.A. Changes in sorghum enzyme inhibitors, phytic acid, tannins and in vitro protein digestibility occurring during Khamir (local bread) fermentation. Food Chem. 2004, 88, 129–134. [Google Scholar] [CrossRef]
- Dicko, M.H.; Gruppen, H.; Traoré, A.S.; van Berkel, W.J.; Voragen, A.G. Evaluation of the effect of germination on phenolic compounds and antioxidant activities in sorghum varieties. J. Agric. Food Chem. 2005, 53, 2581–2588. [Google Scholar] [CrossRef] [PubMed]
- Dirar, H. The Indigenous Fermented Foods and Beverages of Sudan: A Study in African Food and Nutrition; Cambridge University Press: Cambridge, UK, 1991; pp. 23–40. [Google Scholar]
- Elmaki, H.B.; Babiker, E.; El Tinay, A.H. Changes in chemical composition, grain malting, starch and tannin contents and protein digestibility during germination of sorghum cultivars. Food Chem. 1999, 64, 331–336. [Google Scholar] [CrossRef]
- Awika, J.M.; Dykes, L.; Gu, L.; Rooney, L.W.; Prior, R.L. Processing of sorghum (Sorghum bicolor) and sorghum products alters procyanidin oligomer and polymer distribution and content. J. Agric. Food Chem. 2003, 51, 5516–5521. [Google Scholar] [CrossRef] [PubMed]
- Rom, D.; Shull, J.; Chandrashekar, A.; Kirleis, A. on In Vitro Protein Digestibility and Microstructure of Sorghum Flour. Cereal Chem 1992, 69, 178–181. [Google Scholar]
- Oria, M.; Hamaker, B.; Schull, J. In vitro protein digestibility of developing and mature sorghum grain in relation to α-, β-, and γ-kafirin disulfide crosslinking. J. Cereal Sci. 1995, 22, 85–93. [Google Scholar] [CrossRef]
- Maclean, W.C., Jr.; RomaÑa, G.L.d.; Placko, R.P.; Graham, G.G. Protein quality and digestibility of sorghum in preschool children: Balance studies and plasma free amino acids. J. Nutr. 1981, 111, 1928–1936. [Google Scholar] [CrossRef]
- Mudge, S.R.; Campbell, B.C.; Mustapha, N.B.; Godwin, I.D. Genomic approaches for improving grain quality of sorghum. In The Sorghum Genome; Springer: Berlin/Heidelberg, Germany, 2016; pp. 189–205. [Google Scholar]
- Mehlo, L.; Mbambo, Z.; Bado, S.; Lin, J.; Moagi, S.M.; Buthelezi, S.; Stoychev, S.; Chikwamba, R. Induced protein polymorphisms and nutritional quality of gamma irradiation mutants of sorghum. Mutat. Res. 2013, 749, 66–72. [Google Scholar] [CrossRef]
- Yousif, N.E.; El Tinay, A.H. Effect of fermentation on sorghum protein fractions and in vitro protein digestibility. Plant Foods Hum. Nutr. 2001, 56, 175–182. [Google Scholar] [CrossRef]
- Taylor, J.; Belton, P.S. Sorghum. In Pseudocereals and Less Common Cereals; Springer: Berlin/Heidelberg, Germany, 2002; pp. 25–91. [Google Scholar]
- Elkhalifa, A.E.O.; Bernhardt, R.; Bonomi, F.; Iametti, S.; Pagani, M.A.; Zardi, M. Fermentation modifies protein/protein and protein/starch interactions in sorghum dough. Eur. Food Res. Technol. 2006, 222, 559–564. [Google Scholar] [CrossRef]
- Babiker, E.; Kato, A. Improvement of the functional properties of sorghum protein by protein-polysaccharide and protein-protein complexes. Food Nahr. 1998, 42, 286–289. [Google Scholar] [CrossRef]
- Zhang, G.; Hamaker, B.R. Low α-amylase starch digestibility of cooked sorghum flours and the effect of protein. Cereal Chem. 1998, 75, 710–713. [Google Scholar] [CrossRef]
- Yang, P.; Seib, P.A. Low-input wet-milling of grain sorghum for readily accessible starch and animal feed. Cereal Chem. 1995, 72, 498–503. [Google Scholar]
- Xu, X. In vitro digestibility of starch in sorghum differing in endosperm hardness and flour particle size. Ph.D. Thesis, Kansas State University, Manhattan, KS, USA, 2008. [Google Scholar]
- Rooney, L.; Pflugfelder, R. Factors affecting starch digestibility with special emphasis on sorghum and corn. J. Anim. Sci. 1986, 63, 1607–1623. [Google Scholar] [CrossRef]
- Fapojuwo, O.; Maga, J.; Jansen, G. Effect of extrusion cooking on in vitro protein digestibility of sorghum. J. Food Sci. 1987, 52, 218–219. [Google Scholar] [CrossRef]
- Hamaker, B.; Mertz, E.; Axtell, J. Effect of extrusion on sorghum kafirin solubility. Cereal Chem. 1994, 71, 515. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Pros | Cons |
|---|---|
| Large global production | Low protein content |
| Sustainable | Lower protein and starch digestibility |
| Non-GMO | Negative impact on organ health |
| Emerging nutrient source | Deficiency in lysine and threonine |
| Good results in omnivorous and herbivores | Anti-nutritional factors |
| Great source for value-added products | Limited studies in aquaculture |
| High antioxidant level |
| Commodity | Price (US Dollar/Pound) |
|---|---|
| Sorghum | 0.06 |
| Wheat | 0.08 |
| Corn | 0.06 |
| Soybean | 0.14 |
| Country | Sorghum Production (1000 MT) | Changes (%) | |
|---|---|---|---|
| 2019–2020 | 2020–2021 | ||
| USA | 8.67 | 9.47 | 9.2 |
| Nigeria | 6.67 | 6.90 | 3.5 |
| Ethiopia | 5.20 | 5.20 | 0.0 |
| Sudan | 4.00 | 5.00 | 25.0 |
| Mexico | 4.30 | 4.50 | 4.7 |
| India | 4.73 | 3.85 | −18.7 |
| China | 3.60 | 3.55 | −1.4 |
| Argentina | 2.50 | 3.20 | 28.0 |
| Brazil | 2.25 | 2.10 | −6.8 |
| Niger | 1.97 | 1.90 | −3.6 |
| Parameters | White | Red | Black | Hamra | Shahla | Baidha | Dadar | Parbhani | CSH-5 | CSH-9 | Texas | Red | White | Super Sack |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| [34] | [33] | [35] | Authors | |||||||||||
| Moisture (%) | 11.86 | 11.92 | 11.44 | 8.33 | 8.58 | 8.43 | 9.99 | 8.51 | 8.10 | 9.80 | 13.5 | 11.6 | 11.1 | 12.2 |
| Crude protein (%) | 6.14 | 6.85 | 7.28 | 14.80 | 14.51 | 14.75 | 11.02 | 10.65 | 8.90 | 9.60 | 14.9 | 15 | 10.9 | 13.4 |
| Crude fiber (%) | 5.37 | 6.46 | 7.78 | 2.26 | 1.71 | 1.64 | 1.90 | 1.58 | 1.40 | 2.70 | 1.6 | 1.0 | 1.6 | 1.6 |
| Total carbohydrate (%) | 73.17 | 71.32 | 70.07 | 68.34 | 69.65 | 68.81 | 71.89 | 73.35 | 76.20 | 70.65 | 74.8 | 64.2 | 70.3 | 63.8 |
| Crude fat (%) | 2.23 | 2.00 | 1.55 | 4.33 | 3.58 | 4.47 | 2.30 | 2.80 | 2.50 | 2.70 | 3.16 | 3.93 | 3.94 | 4.01 |
| Ash (%) | 1.22 | 1.44 | 1.88 | 1.94 | 1.97 | 1.90 | 1.09 | 0.92 | 1.55 | 1.75 | 2.85 | 2.55 | 2.58 | 2.53 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zarei, M.; Amirkolaei, A.K.; Trushenski, J.T.; Sealey, W.M.; Schwarz, M.H.; Ovissipour, R. Sorghum as a Potential Valuable Aquafeed Ingredient: Nutritional Quality and Digestibility. Agriculture 2022, 12, 669. https://doi.org/10.3390/agriculture12050669
Zarei M, Amirkolaei AK, Trushenski JT, Sealey WM, Schwarz MH, Ovissipour R. Sorghum as a Potential Valuable Aquafeed Ingredient: Nutritional Quality and Digestibility. Agriculture. 2022; 12(5):669. https://doi.org/10.3390/agriculture12050669
Chicago/Turabian StyleZarei, Mohammad, Abdolsamad K. Amirkolaei, Jesse T. Trushenski, Wendy M. Sealey, Michael H. Schwarz, and Reza Ovissipour. 2022. "Sorghum as a Potential Valuable Aquafeed Ingredient: Nutritional Quality and Digestibility" Agriculture 12, no. 5: 669. https://doi.org/10.3390/agriculture12050669
APA StyleZarei, M., Amirkolaei, A. K., Trushenski, J. T., Sealey, W. M., Schwarz, M. H., & Ovissipour, R. (2022). Sorghum as a Potential Valuable Aquafeed Ingredient: Nutritional Quality and Digestibility. Agriculture, 12(5), 669. https://doi.org/10.3390/agriculture12050669