
Enhancing Growth and Glucosinolate Accumulation in Watercress (Nasturtium officinale L.) by Regulating Light Intensity and Photoperiod in Plant Factories


Abstract
:1. Introduction
2. Materials and Methods
2.1. Seedling Conditions
2.2. Treatments
2.3. Measurement of Photosynthetic Parameters and SPAD Value
2.4. Measurement of Plant Growth Parameters
2.5. Determination of the Individual Glucosinolate Concentration in Watercress
2.6. Statistical Analysis
3. Results
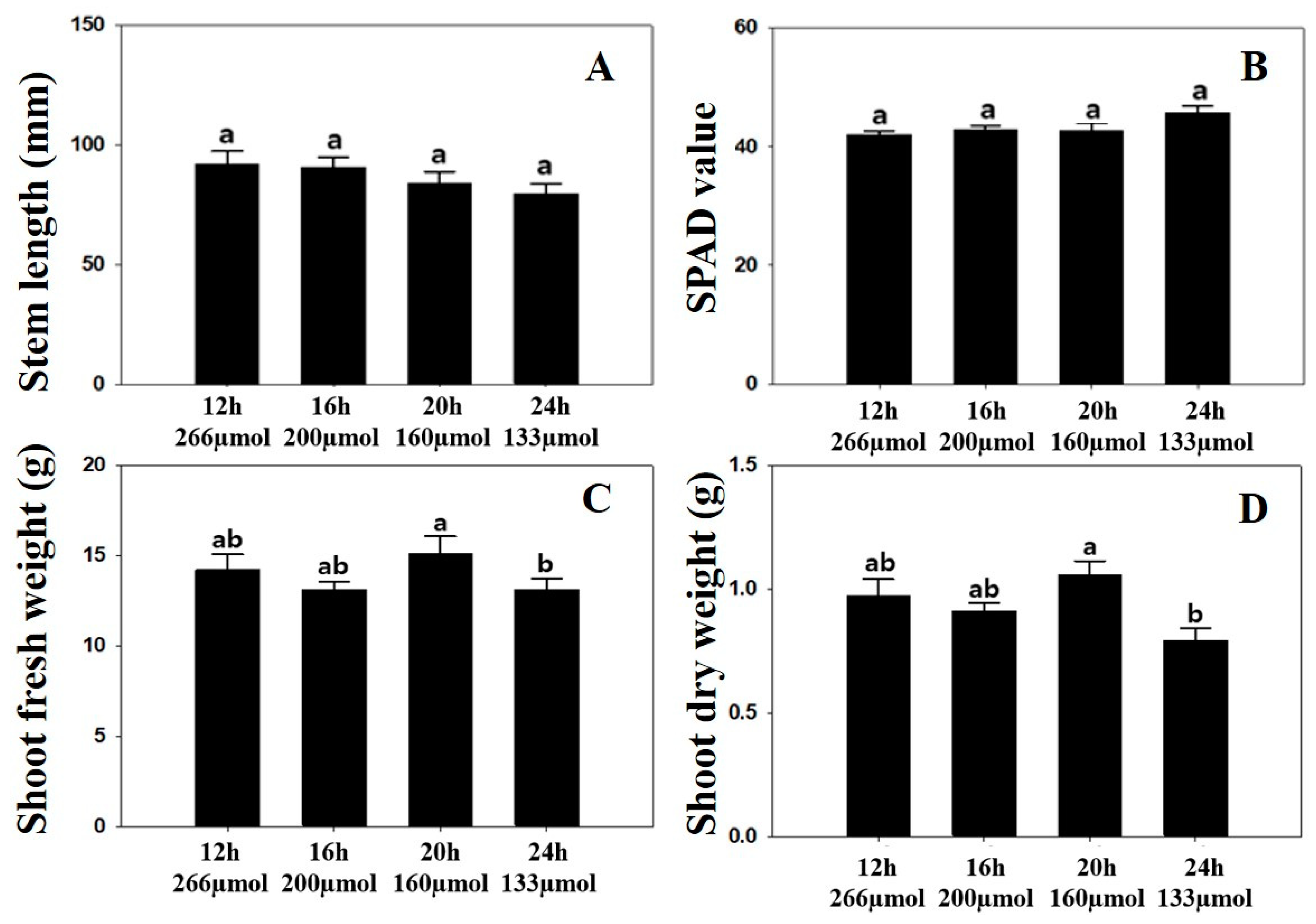
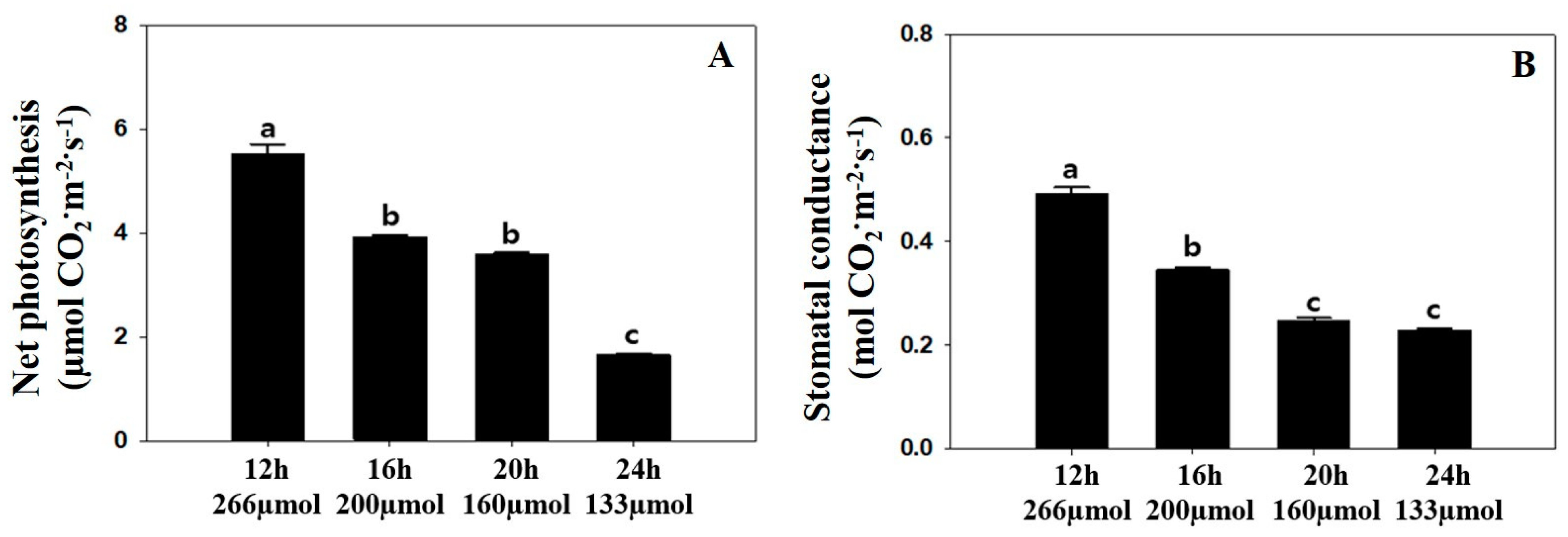
3.1. Plant Growth Parameters, Chlorophyll Content, and Photosynthetic Parameters
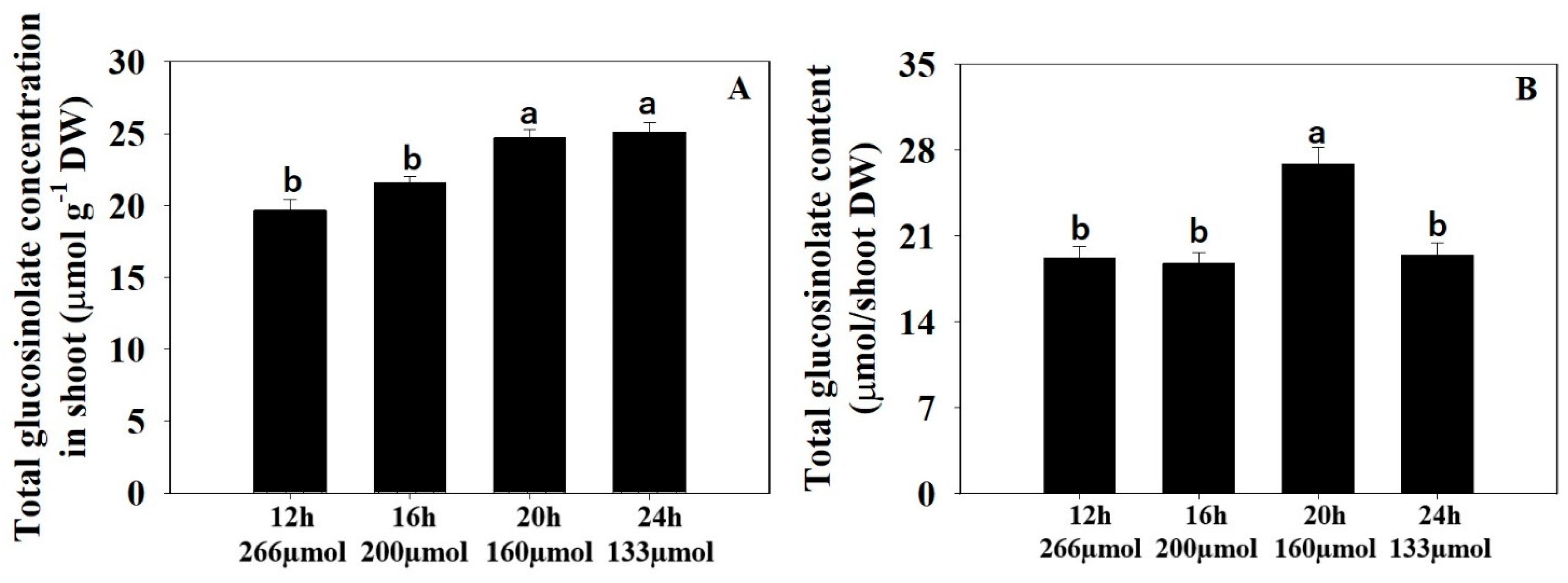
3.2. Total Glucosinolate Concentration and Content
4. Discussion
4.1. Plant Growth Parameters, Chlorophyll Content, and Photosynthetic Parameters
4.2. Total Glucosinolate Concentration and Content
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zeb, A. Phenolic profile and antioxidant potential of wild watercress (Nasturtium officinale L.). Springerplus 2015, 4, 714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klimek-Szczykutowicz, M.; Szopa, A.; Ekiert, H. Chemical composition, traditional and professional use in medicine, application in environmental protection, position in food and cosmetics industries, and biotechnological studies of Nasturtium officinale (watercress)—A review. Fitoterapia 2018, 129, 283–292. [Google Scholar] [CrossRef] [PubMed]
- EFSA European Food Safety Authority (EFSA). Available online: http://www.efsa.europa.eu/ (accessed on 25 January 2020).
- Noia, J.D. Defining powerhouse fruits and vegetables: A nutrient density approach. Prev. Chronic. Dis. 2014, 11, 130390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suroowan, S.; Mahomoodally, M.F. A comparative ethnopharmacological analysis of traditional medicine used against respiratory tract diseases in Mauritius. J. Ethnopharmacol. 2016, 177, 61–80. [Google Scholar] [CrossRef]
- Zafar, R.; Zahoor, M.; Shah, A.; Majid, F. Determination of antioxidants and antibacterial activities, total phenolic, polyphenol and pigment contents in Nasturtium officinale. Pharmacologyonline 2017, 1, 11–18. [Google Scholar]
- Klimek-Szczykutowicz, M.; Dziurka, M.; Blazevic, I.; Dulovic, A.; Granica, S.; Korona-Glowniak, I.; Ekiert, H.; Szopa, A. Phytochemical and biological activity studies on Nasturtium officinale (watercress) microshoot cultures grown in rita(r) temporary immersion systems. Molecules 2020, 25, 5257. [Google Scholar] [CrossRef]
- Al-Kodmany, K. The vertical farm: A review of developments and implications for the vertical city. Buildings 2018, 8, 24. [Google Scholar] [CrossRef] [Green Version]
- Ali, S.; Liu, Y.; Ishaq, M.; Shah, T.; Abdullah; Ilyas, A.; Din, I.U. Climate change and its impact on the yield of major food crops: Evidence from pakistan. Foods 2017, 6, 39. [Google Scholar] [CrossRef]
- Zhao, C.; Liu, B.; Piao, S.L.; Wang, X.H.; Lobell, D.B.; Huang, Y.; Huang, M.T.; Yao, Y.T.; Bassu, S.; Ciais, P.; et al. Temperature increase reduces global yields of major crops in four independent estimates. Proc. Natl. Acad. Sci. USA 2017, 114, 9326–9331. [Google Scholar] [CrossRef] [Green Version]
- Kozai, T. Resource use efficiency of closed plant production system with artificial light: Concept, estimation and application to plant factory. Proc. Jpn. Acad. Ser. 2013, 89, 447–461. [Google Scholar] [CrossRef] [Green Version]
- Kozai, T. Sustainable plant factory: Closed plant production systems with artificial light for high resource use efficiencies and quality produce. Acta Hortic. 2013, 1004, 27–40. [Google Scholar] [CrossRef]
- Zakurin, A.O.; Shchennikova, A.V.; Kamionskaya, A.M. Artificial-light culture in protected ground plant growing: Photosynthesis, photomorphogenesis, and prospects of LED application. Russ. J. Plant Physl. 2020, 67, 413–424. [Google Scholar] [CrossRef]
- Liu, Y.; Ren, X.X.; Jeong, B.R. Supplementary Light source affects growth, metabolism, and physiology of Adenophora triphylla (thunb.) A.DC. seedlings. Biomed. Res. Int. 2019, 2019. [Google Scholar] [CrossRef] [Green Version]
- Mao, H.P.; Hang, T.; Zhang, X.D.; Lu, N. Both multi-segment light intensity and extended photoperiod lighting strategies, with the same daily light integral, promoted lactuca sativa l. growth and photosynthesis. Agronomy 2019, 9, 857. [Google Scholar] [CrossRef] [Green Version]
- Kitaya, Y.; Niu, G.; Kozai, T.; Ohashi, M. Photosynthetic photon flux, photoperiod, and CO2 concentration affect growth and morphology of lettuce plug transplants. HortScience 1998, 33, 988–991. [Google Scholar] [CrossRef] [Green Version]
- Vlahos, J.; Heuvelink, E.; Martakis, G. A growth analysis study of three Achimenes cultivars grown under three light regimes. Sci. Hortic. 1991, 46, 275–282. [Google Scholar] [CrossRef]
- Zha, L.Y.; Liu, W.K. Effects of light quality, light intensity, and photoperiod on growth and yield of cherry radish grown under red plus blue LEDs. Hortic. Environ. Biote 2018, 59, 511–518. [Google Scholar] [CrossRef]
- Lu, N.; Bernardo, E.L.; Tippayadarapanich, C.; Takagaki, M.; Kagawa, N.; Yamori, W. Growth and accumulation of secondary metabolites in perilla as affected by photosynthetic photon flux density and electrical conductivity of the nutrient solution. Front. Plant Sci. 2017, 8, 708. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.J.; Kim, H.M.; Kim, H.M.; Jeong, B.R.; Lee, H.J.; Kim, H.J.; Hwang, S.J. Ice plant growth and phytochemical concentrations are affected by light quality and intensity of monochromatic light-emitting diodes. Hortic. Environ. Biote 2018, 59, 529–536. [Google Scholar] [CrossRef]
- Fan, X.X.; Xu, Z.G.; Liu, X.Y.; Tang, C.M.; Wang, L.W.; Han, X.L. Effects of light intensity on the growth and leaf development of young tomato plants grown under a combination of red and blue light. Sci. Hortic. 2013, 153, 50–55. [Google Scholar] [CrossRef]
- Kang, J.H.; KrishnaKumar, S.; Atulba, S.L.S.; Jeong, B.R.; Hwang, S.J. Light intensity and photoperiod influence the growth and development of hydroponically grown leaf lettuce in a closed-type plant factory system. Hortic. Environ. Biote 2013, 54, 501–509. [Google Scholar] [CrossRef]
- Yin, Y.H.; Yu, C.J.; Yu, L.; Zhao, J.S.; Sun, C.J.; Ma, Y.B.; Zhou, G.K. The influence of light intensity and photoperiod on duckweed biomass and starch accumulation for bioethanol production. Bioresour. Technol. 2015, 187, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Rehman, M.; Ullah, S.; Bao, Y.; Wang, B.; Peng, D.; Liu, L. Light-emitting diodes: Whether an efficient source of light for indoor plants? Environ. Sci. Pollut. Res. Int. 2017, 24, 24743–24752. [Google Scholar] [CrossRef] [PubMed]
- Taulavuori, E.; Taulavuori, K.; Holopainen, J.K.; Julkunen-Tiitto, R.; Acar, C.; Dincer, I. Targeted use of LEDs in improvement of production efficiency through phytochemical enrichment. J. Sci. Food Agric. 2017, 97, 5059–5064. [Google Scholar] [CrossRef] [PubMed]
- Engelen-Eigles, G.; Holden, G.; Cohen, J.D.; Gardner, G. The effect of temperature, photoperiod, and light quality on gluconasturtiin concentration in watercress (Nasturtium officinale R. Br.). J. Agric. Food Chem. 2006, 54, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.Y.; Kim, S.J.; Bok, K.J.; Lee, K.Y.; Park, J.S. Effect of different nutrient solution and light quality on growth and glucosinolate contents of watercress in hydroponics. J. Bio-Environ. Control 2018, 27, 371–380. [Google Scholar] [CrossRef]
- Lam, V.P.; Choi, J.; Kim, S.; Park, J.; Hernandez, R. Optimizing plant spacing and harvest time for yield and glucosinolate accumulation in watercress (Nasturtium officinale L.) grown in a hydroponic system. Hortic. Sci. Technol. 2019, 37, 733–743. [Google Scholar] [CrossRef]
- Cuong, D.M.; Park, C.H.; Bong, S.J.; Kim, N.S.; Kim, J.K.; Park, S.U. Enhancement of glucosinolate production in watercress (Nasturtium officinale) hairy roots by overexpressing cabbage transcription factors. J. Agric. Food Chem. 2019, 67, 4860–4867. [Google Scholar] [CrossRef]
- ISO Norm. Rapeseed-Determination of glucosinolate content. Part 1: Method using high-performance liquid chromatography. ISO 1992, 9167, 1–9. [Google Scholar]
- Li, Q.; Kubota, C. Effects of supplemental light quality on growth and phytochemicals of baby leaf lettuce. Environ. Exp. Bot. 2009, 67, 59–64. [Google Scholar] [CrossRef]
- Solovchenko, A.E.; Khozin-Goldberg, I.; Didi-Cohen, S.; Cohen, Z.; Merzlyak, M.N. Effects of light intensity and nitrogen starvation on growth, total fatty acids and arachidonic acid in the green microalga Parietochloris Incisa. J. Appl. Phycol. 2008, 20, 245–251. [Google Scholar] [CrossRef]
- Feng, L.Y.; Raza, M.A.; Li, Z.C.; Chen, Y.K.; Bin Khalid, M.H.; Du, J.B.; Liu, W.G.; Wu, X.L.; Song, C.; Yu, L.; et al. The influence of light intensity and leaf movement on photosynthesis characteristics and carbon balance of soybean. Front. Plant Sci. 2019, 9, 1925. [Google Scholar] [CrossRef] [PubMed]
- Gommers, C.M.M.; Visser, E.J.W.; Onge, K.R.S.; Voesenek, L.A.C.J.; Pierik, R. Shade tolerance: When growing tall is not an option. Trends Plant Sci. 2013, 18, 65–71. [Google Scholar] [CrossRef]
- Yang, F.; Huang, S.; Gao, R.C.; Liu, W.G.; Yong, T.W.; Wang, X.C.; Wu, X.L.; Yang, W.Y. Growth of soybean seedlings in relay strip intercropping systems in relation to light quantity and red:far-red ratio. Field. Crop Res. 2014, 155, 245–253. [Google Scholar] [CrossRef]
- Schumann, T.; Paul, S.; Melzer, M.; Dormann, P.; Jahns, P. Plant growth under natural light conditions provides highly flexible short-term acclimation properties toward high light stress. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Austin, J.; Jeon, Y.A.; Cha, M.-K.; Park, S.; Cho, Y.-Y. Effects of photoperiod, light intensity and electrical conductivity on the growth and yield of quinoa (Chenopodium quinoa Willd.) in a closedtype plant factory system. Hortic. Sci. Technol. 2016, 34, 405–413. [Google Scholar] [CrossRef]
- Gong, W.Z.; Qi, P.F.; Du, J.B.; Sun, X.; Wu, X.L.; Song, C.; Liu, W.G.; Wu, Y.S.; Yu, X.B.; Yong, T.W.; et al. Transcriptome analysis of shade-induced inhibition on leaf size in relay intercropped soybean. PLoS ONE 2014, 9, e98465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shafiq, I.; Hussain, S.; Raza, M.A.; Iqbal, N.; Ahsan Asghar, M.; Raza, A.; Fan, Y.F.; Mumtaz, M.; Shoaib, M.; Ansar, M.; et al. Crop photosynthetic response to light quality and light intensity. J. Integr. Agric. 2021, 20, 4–23. [Google Scholar] [CrossRef]
- Cen, Y.P.; Sage, R.F. The regulation of rubisco activity in response to variation in temperature and atmospheric CO2 partial pressure in sweet potato. Plant Physiol. 2005, 139, 979–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, B.Y.; Song, Y.X.; Song, C.; Cui, L.; Yong, T.W.; Yang, W.Y. Growth and photosynthetic responses of soybean seedlings to maize shading in relay intercropping system in Southwest China. Photosynthetica 2014, 52, 332–340. [Google Scholar] [CrossRef]
- Zhu, H.F.; Li, X.F.; Zhai, W.; Liu, Y.; Gao, Q.Q.; Liu, J.P.; Ren, L.; Chen, H.Y.; Zhu, Y.Y. Effects of low light on photosynthetic properties, antioxidant enzyme activity, and anthocyanin accumulation in purple pak-choi (Brassica campestris ssp Chinensis Makino). PLoS ONE 2017, 12, e0179305. [Google Scholar] [CrossRef] [PubMed]
- Huseby, S.; Koprivova, A.; Lee, B.R.; Saha, S.; Mithen, R.; Wold, A.B.; Bengtsson, G.B.; Kopriva, S. Diurnal and light regulation of sulphur assimilation and glucosinolate biosynthesis in Arabidopsis. J. Exp. Bot. 2013, 64, 1039–1048. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Zhu, Z.J.; Gerendas, J. Interactive effects of phosphorus supply and light intensity on glucosinolates in pakchoi (Brassica campestris L. ssp chinensis var. communis). Plant Soil. 2009, 323, 323–333. [Google Scholar] [CrossRef]
- Farhan, M.; Razak, S.A.; Pin, K.Y.; Chuah, A.L. Antioxidant activity and phenolic content of different parts of Orthosiphon stamineus grown under different light intensities. J. Trop. Sci. 2012, 24, 173–177. [Google Scholar]
- Carvalho, I.S.; Cavaco, T.; Carvalho, L.M.; Duque, P. Effect of photoperiod on flavonoid pathway activity in sweet potato (Ipomoea batatas (L.) Lam.) leaves. Food Chem. 2010, 118, 384–390. [Google Scholar] [CrossRef]
- Cho, J.Y.; Yoo, K.S.; Kim, J.; Choi, B.J.; Oh, W. Growth and bioactive compounds of lettuce as affected by light intensity and photoperiod in a plant factory using external electrode fluorescent lamps. Hortic. Sci. Technol. 2020, 38, 645–659. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Common Name | Side Chain Structure | Retention Time (min) | Response Factor |
|---|---|---|---|
| Glucobrassicin | indol-3-ylmethyl | 17.16 | 0.29 |
| 4- Methoxyglucobrassicin | 4-methoxyindol-3-ylmethyl | 16.05 | 0.25 |
| Glucohirsutin | 8-methylsulfinyloctyl | 13.78 | 1.1 |
| Glucosiberin | 7-methylsulfinylheptyl | 16.77 | 1 |
| Gluconasturtiin | 2-phenylethyl | 15.68 | 0.95 |
| Lighting Treatment | Individual Glucosinolate Concentration in Watercress Shoots (mg·g−1 DW) z | ||||
|---|---|---|---|---|---|
| Siber | Hirsu | Brassi | Metho | Nastur | |
| 12 h—266 µmol | 1.04b | 0.53 | 1.83ab | 0.58b | 15.69b |
| 16 h—200 µmol | 0.63b | 0.45 | 1.75ab | 0.68ab | 18.92a |
| 20 h—160 µmol | 3.36a | 0.59 | 1.40b | 0.94a | 18.81ab |
| 24 h—133 µmol | 0.46b | 0.53 | 2.21a | 0.79ab | 20.71a |
| Significance y | *** | NS | ** | *** | * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lam, V.P.; Choi, J.; Park, J. Enhancing Growth and Glucosinolate Accumulation in Watercress (Nasturtium officinale L.) by Regulating Light Intensity and Photoperiod in Plant Factories. Agriculture 2021, 11, 723. https://doi.org/10.3390/agriculture11080723
Lam VP, Choi J, Park J. Enhancing Growth and Glucosinolate Accumulation in Watercress (Nasturtium officinale L.) by Regulating Light Intensity and Photoperiod in Plant Factories. Agriculture. 2021; 11(8):723. https://doi.org/10.3390/agriculture11080723
Chicago/Turabian StyleLam, Vu Phong, Jaeyun Choi, and Jongseok Park. 2021. "Enhancing Growth and Glucosinolate Accumulation in Watercress (Nasturtium officinale L.) by Regulating Light Intensity and Photoperiod in Plant Factories" Agriculture 11, no. 8: 723. https://doi.org/10.3390/agriculture11080723
APA StyleLam, V. P., Choi, J., & Park, J. (2021). Enhancing Growth and Glucosinolate Accumulation in Watercress (Nasturtium officinale L.) by Regulating Light Intensity and Photoperiod in Plant Factories. Agriculture, 11(8), 723. https://doi.org/10.3390/agriculture11080723