Mechanisms of Testicular Disruption from Exposure to Bisphenol A and Phtalates

 , and
, and 
Abstract
1. Introduction—The Endocrine Disruptor “Dilemma”
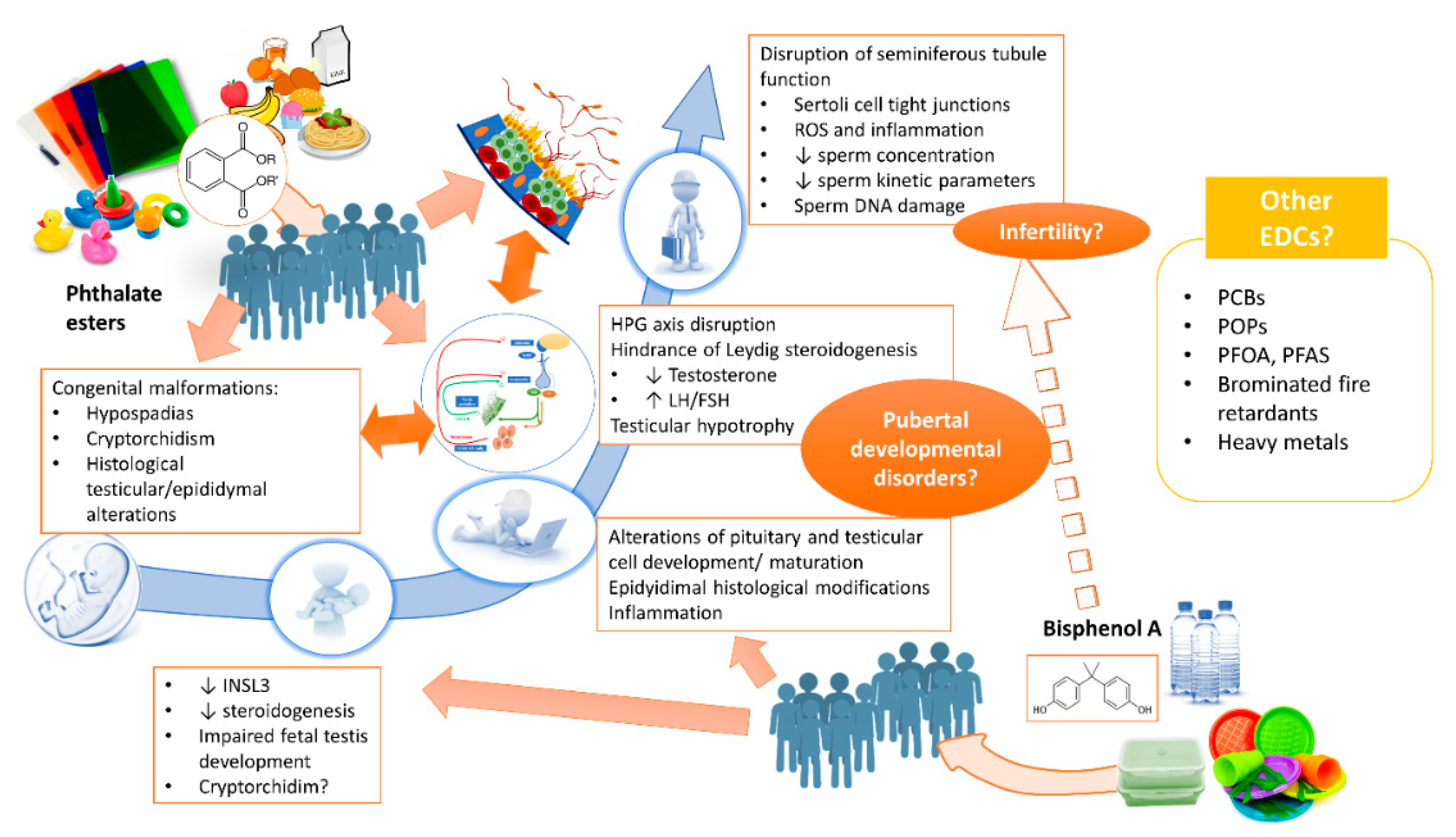
2. Bisphenol A and Phthalate Esters
2.1. Pre-Natal Exposure: Can Testicular Function Already be Compromised in Utero?
2.2. Postnatal Exposure: Are There Repercussions for Adult Testicular Function?
3. Clinical Data: The Real-Life Impact of BPA and Phthalates on Testicular Function
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gore, A.C.; Chappell, V.A.; Fenton, S.E.; Flaws, J.A.; Nadal, A.; Prins, G.S.; Toppari, J.; Zoeller, R.T. Executive Summary to EDC-2: The Endocrine Society’s second Scientific Statement on endocrine-disrupting chemicals. Endocr. Rev. 2015, 36, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Barouki, R. Endocrine disruptors: Revisiting concepts and dogma in toxicology. Comptes Rendus-Biol. 2017, 340, 410–413. [Google Scholar] [CrossRef] [PubMed]
- Kiyama, R.; Wada-Kiyama, Y. Estrogenic endocrine disruptors: Molecular mechanisms of action. Environ. Int. 2015, 83, 11–40. [Google Scholar] [CrossRef]
- Müllerová, D.; Kopecký, J. White adipose tissue: Storage and effector site for environmental pollutants. Physiol. Res. 2007, 56, 375–381. [Google Scholar]
- Pestana, D.; Faria, G.; Sá, C.; Fernandes, V.C.; Teixeira, D.; Norberto, S.; Faria, A.; Meireles, M.; Marques, C.; Correia-Sá, L.; et al. Persistent organic pollutant levels in human visceral and subcutaneous adipose tissue in obese individuals-Depot differences and dysmetabolism implications. Environ. Res. 2014, 133, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Jacobs, D.R.; Park, H.Y.; Carpenter, D.O. A role of low dose chemical mixtures in adipose tissue in carcinogenesis. Environ. Int. 2017, 108, 170–175. [Google Scholar] [CrossRef]
- De Kretser, D.M.; Loveland, K.L.; Meinhardt, A.; Simorangkir, D.; Wreford, N. Spermatogenesis. Hum. Reprod. 1998, 13, 1–8. [Google Scholar] [CrossRef]
- Neto, F.T.L.; Bach, P.V.; Najari, B.B.; Li, P.S.; Goldstein, M. Spermatogenesis in humans and its affecting factors. Semin. Cell Dev. Biol. 2016, 59, 10–26. [Google Scholar] [CrossRef]
- Tavares, R.S.; Escada-Rebelo, S.; Correia, M.; Mota, P.C.; Ramalho-Santos, J. The non-genomic effects of endocrine-disrupting chemicals on mammalian sperm. Reproduction 2016, 151, R1–R13. [Google Scholar] [CrossRef]
- Yeung, B.H.; Wan, H.T.; Law, A.Y.; Wong, C.K. Endocrine disrupting chemicals: Multiple effects on testicular signaling and spermatogenesis. Spermatogenesis 2011, 1, 231–239. [Google Scholar] [CrossRef]
- Kabir, E.R.; Rahman, M.S.; Rahman, I. A review on endocrine disruptors and their possible impacts on human health. Environ. Toxicol. Pharmacol. 2015, 40, 241–258. [Google Scholar] [CrossRef] [PubMed]
- Eskenazi, B.; Warner, M.; Brambilla, P.; Signorini, S.; Ames, J.; Mocarelli, P. The Seveso accident: A look at 40 years of health research and beyond. Environ. Int. 2018, 121, 71–84. [Google Scholar] [CrossRef]
- Vandenberg, L.N.; Colborn, T.; Hayes, T.B.; Heindel, J.J.; Jacobs, D.R.; Lee, D.H.; Shioda, T.; Soto, A.M.; vom Saal, F.S.; Welshons, W.V.; et al. Hormones and endocrine-disrupting chemicals: Low-dose effects and nonmonotonic dose responses. Endocr. Rev. 2012, 33, 378–455. [Google Scholar] [CrossRef]
- Welshons, W.V.; Thayer, K.A.; Judy, B.M.; Taylor, J.A.; Curran, E.M.; vom Saal, F.S. Large effects from small exposures. I. Mechanisms for endocrine-disrupting chemicals with estrogenic activity. Environ. Health Perspect. 2003, 111, 994–1006. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Jacobs, D.R. New approaches to cope with possible harms of low-dose environmental chemicals. J. Epidemiol. Community Health 2019, 73, 193–197. [Google Scholar] [CrossRef]
- Lee, D.H. Evidence of the possible harm of endocrine-disrupting chemicals in humans: Ongoing debates and key issues. Endocrinol. Metab. 2018, 33, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Kortenkamp, A. Which chemicals should be grouped together for mixture risk assessments of male reproductive disorders? Mol. Cell. Endocrinol. 2020, 499, 110581. [Google Scholar] [CrossRef] [PubMed]
- Philips, E.M.; Kahn, L.G.; Jaddoe, V.W.V.; Shao, Y.; Asimakopoulos, A.G.; Kannan, K.; Steegers, E.A.P.; Trasande, L. First Trimester Urinary Bisphenol and Phthalate Concentrations and Time to Pregnancy: A Population-Based Cohort Analysis. J. Clin. Endocrinol. Metab. 2018, 103, 3540–3547. [Google Scholar] [CrossRef] [PubMed]
- Murata, M.; Kang, J.H. Bisphenol A (BPA) and cell signaling pathways. Biotechnol. Adv. 2018, 36, 311–327. [Google Scholar] [CrossRef]
- Wang, H.; Ding, Z.; Shi, Q.M.; Ge, X.; Wang, H.X.; Li, M.X.; Chen, G.; Wang, Q.; Ju, Q.; Zhang, J.P.; et al. Anti-androgenic mechanisms of Bisphenol A involve androgen receptor signaling pathway. Toxicology 2017, 387, 10–16. [Google Scholar] [CrossRef]
- Alonso-Magdalena, P.; Ropero, A.B.; Soriano, S.; García-Arévalo, M.; Ripoll, C.; Fuentes, E.; Quesada, I.; Nadal, Á. Bisphenol-A acts as a potent estrogen via non-classical estrogen triggered pathways. Mol. Cell. Endocrinol. 2012, 355, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Xi, W.; Lee, C.K.F.; Yeung, W.S.B.; Giesy, J.P.; Wong, M.H.; Zhang, X.; Hecker, M.; Wong, C.K.C. Effect of perinatal and postnatal bisphenol A exposure to the regulatory circuits at the hypothalamus-pituitary-gonadal axis of CD-1 mice. Reprod. Toxicol. 2011, 31, 409–417. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, D.; Yanagiba, Y.; Duan, Z.; Ito, Y.; Okamura, A.; Asaeda, N.; Tagawa, Y.; Li, C.M.; Taya, K.; Zhang, S.Y.; et al. Bisphenol A may cause testosterone reduction by adversely affecting both testis and pituitary systems similar to estradiol. Toxicol. Lett. 2010, 194, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Díaz, S.; Mitchell, A.A.; Kelley, K.E.; Calafat, A.M.; Hauser, R. Medications as a potential source of exposure to phthalates in the U.S. population. Environ. Health Perspect. 2009, 117, 185–189. [Google Scholar] [CrossRef]
- Christen, V.; Crettaz, P.; Oberli-Schrämmli, A.; Fent, K. Antiandrogenic activity of phthalate mixtures: Validity of concentration addition. Toxicol. Appl. Pharmacol. 2012, 259, 169–176. [Google Scholar] [CrossRef]
- Takeuchi, S.; Iida, M.; Kobayashi, S.; Jin, K.; Matsuda, T.; Kojima, H. Differential effects of phthalate esters on transcriptional activities via human estrogen receptors α and β, and androgen receptor. Toxicology 2005, 210, 223–233. [Google Scholar] [CrossRef]
- Engel, A.; Buhrke, T.; Imber, F.; Jessel, S.; Seidel, A.; Völkel, W.; Lampen, A. Agonistic and antagonistic effects of phthalates and their urinary metabolites on the steroid hormone receptors ERα, ERβ, and AR. Toxicol. Lett. 2017, 277, 54–63. [Google Scholar] [CrossRef]
- Wisniewski, P.; Romano, R.M.; Kizys, M.M.L.; Oliveira, K.C.; Kasamatsu, T.; Giannocco, G.; Chiamolera, M.I.; Dias-da-Silva, M.R.; Romano, M.A. Adult exposure to bisphenol A (BPA) in Wistar rats reduces sperm quality with disruption of the hypothalamic-pituitary-testicular axis. Toxicology 2015, 329, 1–9. [Google Scholar] [CrossRef]
- Rahman, M.S.; Kwon, W.S.; Lee, J.S.; Yoon, S.J.; Ryu, B.Y.; Pang, M.G. Bisphenol-A Affects Male Fertility via Fertility-related Proteins in Spermatozoa. Sci. Rep. 2015, 5, 9169. [Google Scholar] [CrossRef]
- Mitro, S.D.; Johnson, T.; Zota, A.R. Cumulative Chemical Exposures during Pregnancy and Early Development. Curr. Environ. Health Rep. 2015, 2, 367–378. [Google Scholar] [CrossRef]
- Gaudriault, P.; Mazaud-Guittot, S.; Lavoué, V.; Coiffec, I.; Lesné, L.; Dejucq-Rainsford, N.; Scholze, M.; Kortenkamp, A.; Jégou, B. Endocrine disruption in human fetal testis explants by individual and combined exposures to selected pharmaceuticals, pesticides, and environmental pollutants. Environ. Health Perspect. 2017, 125, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Conley, J.M.; Lambright, C.S.; Evans, N.; Cardon, M.; Furr, J.; Wilson, V.S.; Gray, L.E.J. Mixed “Antiandrogenic” Chemicals at Low Individual Doses Produce Reproductive Tract Malformations in the Male Rat. Toxicol. Sci. 2018, 164, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Howdeshell, K.L.; Furr, J.; Lambright, C.R.; Rider, C.V.; Wilson, V.S.; Gray, L.E. Cumulative effects of dibutyl phthalate and diethylhexyl phthalate on male rat reproductive tract development: Altered fetal steroid hormones and genes. Toxicol. Sci. 2007, 99, 190–202. [Google Scholar] [CrossRef] [PubMed]
- Howdeshell, K.L.; Rider, C.V.; Wilson, V.S.; Gray, L.E.J. Mechanisms of action of phthalate esters, individually and in combination, to induce abnormal reproductive development in male laboratory rats. Environ. Res. 2008, 108, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.T.; Ma, L.; Yuan, L.; Wang, X.R.; Zhang, W. Study on developmental abnormalities in hypospadiac male rats induced by maternal exposure to di-n-butyl phthalate (DBP). Toxicology 2007, 232, 286–293. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.J.; Jiang, J.T.; Ma, L.; Zhang, J.; Hong, Y.; Liao, K.; Liu, Q.; Liu, G.H. Molecular and toxicologic research in newborn hypospadiac male rats following in utero exposure to di-n-butyl phthalate (DBP). Toxicology 2009, 260, 120–125. [Google Scholar] [CrossRef]
- Barakat, R.; Lin, P.C.P.; Rattan, S.; Brehm, E.; Canisso, I.F.; Abosalum, M.E.; Flaws, J.A.; Hess, R.; Ko, C.M. Prenatal exposure to DEHP induces premature reproductive senescence in male mice. Toxicol. Sci. 2017, 156, 96–108. [Google Scholar] [CrossRef]
- Barakat, R.; Seymore, T.; Lin, P.C.P.; Park, C.J.; Ko, C.M.J. Prenatal exposure to an environmentally relevant phthalate mixture disrupts testicular steroidogenesis in adult male mice. Environ. Res. 2019, 172, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Chevalier, N.; Brucker-Davis, F.; Lahlou, N.; Coquillard, P.; Pugeat, M.; Pacini, P.; Panaïa-Ferrari, P.; Wagner-Mahler, K.; Fénichel, P. A negative correlation between insulin-like peptide 3 and bisphenol A in human cord blood suggests an effect of endocrine disruptors on testicular descent during fetal development. Hum. Reprod. 2015, 30, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Wilson, V.S.; Lambright, C.; Furr, J.; Ostby, J.; Wood, C.; Held, G.; Gray, L.E. Phthalate ester-induced gubernacular lesions are associated with reduced insl3 gene expression in the fetal rat testis. Toxicol. Lett. 2004, 146, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Ge, R.S.; Chen, G.R.; Hu, G.X.; Dong, L.; Lian, Q.Q.; Hardy, D.O.; Sottas, C.M.; Li, X.K.; Hardy, M.P. Involvement of testicular growth factors in fetal Leydig cell aggregation after exposure to phthalate in utero. Proc. Natl. Acad. Sci. USA 2008, 105, 7218–7222. [Google Scholar] [CrossRef] [PubMed]
- Pathirana, I.N.; Kawate, N.; Tsuji, M.; Takahashi, M.; Hatoya, S.; Inaba, T.; Tamada, H. In vitro effects of estradiol-17β, monobutyl phthalate and mono-(2-ethylhexyl) phthalate on the secretion of testosterone and insulin-like peptide 3 by interstitial cells of scrotal and retained testes in dogs. Theriogenology 2011, 76, 1227–1233. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Bu, T.; Su, H.; Chen, Z.; Liang, Y.; Zhang, G.; Zhu, D.; Shan, Y.; Xu, R.; Hu, Y.; et al. In utero exposure to diisononyl phthalate caused testicular dysgenesis of rat fetal testis. Toxicol. Lett. 2015, 232, 466–474. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Chen, X.; Hu, G.; Li, L.; Su, H.; Wang, Y.; Chen, D.; Zhu, Q.; Li, C.; Li, J.; et al. Effects of in utero exposure to dicyclohexyl phthalate on rat fetal leydig cells. Int. J. Environ. Res. Public Health 2016, 13, 246. [Google Scholar] [CrossRef] [PubMed]
- Aydoğan Ahbab, M.; Barlas, N. Influence of in utero di-n-hexyl phthalate and dicyclohexyl phthalate on fetal testicular development in rats. Toxicol. Lett. 2015, 233, 125–137. [Google Scholar] [CrossRef]
- Lara, N.L.M.; Van Den Driesche, S.; MacPherson, S.; França, L.R.; Sharpe, R.M. Dibutyl phthalate induced testicular dysgenesis originates after seminiferous cord formation in rats. Sci. Rep. 2017, 7, 2521. [Google Scholar] [CrossRef]
- Hu, G.; Li, J.; Shan, Y.; Li, X.; Zhu, Q.; Li, H.; Wang, Y.; Chen, X.; Lian, Q.; Ge, R.S. In utero combined di-(2-ethylhexyl) phthalate and diethyl phthalate exposure cumulatively impairs rat fetal Leydig cell development. Toxicology 2018, 395, 23–33. [Google Scholar] [CrossRef]
- Fisher, J.S.; Macpherson, S.; Marchetti, N.; Sharpe, R.M. Human “testicular dysgenesis syndrome”: A possible model using in-utero exposure of the rat to dibutyl phthalate. Hum. Reprod. 2003, 18, 1383–1394. [Google Scholar] [CrossRef]
- Kilcoyne, K.R.; Smith, L.B.; Atanassova, N.; Macpherson, S.; McKinnell, C.; Van Den Driesche, S.; Jobling, M.S.; Chambers, T.J.G.; De Gendt, K.; Verhoeven, G.; et al. Fetal programming of adult Leydig cell function by androgenic effects on stem/progenitor cells. Proc. Natl. Acad. Sci. USA 2014, 111, E1924–E1932. [Google Scholar] [CrossRef]
- N’Tumba-Byn, T.; Moison, D.; Lacroix, M.; Lecureuil, C.; Lesage, L.; Prud’homme, S.M.; Pozzi-Gaudin, S.; Frydman, R.; Benachi, A.; Livera, G.; et al. Differential Effects of Bisphenol A and Diethylstilbestrol on Human, Rat and Mouse Fetal Leydig Cell Function. PLoS ONE 2012, 7, e51579. [Google Scholar] [CrossRef]
- Ben Maamar, M.; Lesne, L.; Desdoits-Lethimonier, C.; Coiffec, I.; Lassurguère, J.; Lavoué, V.; Deceuninck, Y.; Antignac, J.P.; Le Bizec, B.; Perdu, E.; et al. An investigation of the endocrine-disruptive effects of bisphenol A in human and rat fetal testes. PLoS ONE 2015, 10, e0128051. [Google Scholar]
- Lv, Y.; Li, L.; Fang, Y.; Chen, P.; Wu, S.; Chen, X.; Ni, C.; Zhu, Q.; Huang, T.; Lian, Q.; et al. In utero exposure to bisphenol A disrupts fetal testis development in rats. Environ. Pollut. 2019, 246, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Shi, W.; Wang, X.; Song, P.; Zhong, X. Bisphenol A Exposure during Pregnancy Alters the Mortality and Levels of Reproductive Hormones and Genes in Offspring Mice. BioMed Res. Int. 2017, 2017, 3585809. [Google Scholar] [CrossRef] [PubMed]
- Ullah, A.; Pirzada, M.; Jahan, S.; Ullah, H.; Turi, N.; Ullah, W.; Siddiqui, M.F.; Zakria, M.; Lodhi, K.Z.; Khan, M.M. Impact of low-dose chronic exposure to bisphenol A and its analogue bisphenol B, bisphenol F and bisphenol S on hypothalamo-pituitary-testicular activities in adult rats: A focus on the possible hormonal mode of action. Food Chem. Toxicol. 2018, 121, 24–36. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Han, C.; Li, S.; Cui, Y.; Bao, Y.; Shi, W. Maternal exposure to bisphenol A during pregnancy interferes ovaries development of F1 female mice. Environ. Sci. Pollut. Res. 2019, 26, 23491–23504. [Google Scholar] [CrossRef]
- Shi, M.; Sekulovski, N.; MacLean, J.A.; Hayashi, K. Prenatal exposure to bisphenol A analogues on male reproductive functions in mice. Toxicol. Sci. 2018, 163, 620–631. [Google Scholar] [CrossRef]
- Gules, O.; Yildiz, M.; Naseer, Z.; Tatar, M. Effects of folic acid on testicular toxicity induced by bisphenol-A in male Wistar rats. Biotech. Histochem. 2019, 94, 26–35. [Google Scholar] [CrossRef]
- Srivastava, S.; Gupta, P. Alteration in apoptotic rate of testicular cells and sperms following administration of Bisphenol A (BPA) in Wistar albino rats. Environ. Sci. Pollut. Res. 2018, 25, 21635–21643. [Google Scholar] [CrossRef]
- Dere, E.; Anderson, L.M.; Huse, S.M.; Spade, D.J.; McDonnell-Clark, E.; Madnick, S.J.; Hall, S.J.; Camacho, L.; Lewis, S.M.; Vanlandingham, M.M.; et al. Effects of continuous bisphenol A exposure from early gestation on 90day old rat testes function and sperm molecular profiles: A CLARITY-BPA consortium study. Toxicol. Appl. Pharmacol. 2018, 347, 1–9. [Google Scholar] [CrossRef]
- Atanassova, N.; McKinnell, C.; Turner, K.J.; Walker, M.; Fisher, J.S.; Morley, M.; Millar, M.R.; Groome, N.P.; Sharpe, R.M. Comparative effects of neonatal exposure of male rats to potent and weak (environmental) estrogens on spermatogenesis at puberty and the relationship, to adult testis size and fertility: Evidence for stimulatory effects of low estrogen levels. Endocrinology 2000, 141, 3898–3907. [Google Scholar] [CrossRef]
- Meng, Y.; Lin, R.; Wu, F.; Sun, Q.; Jia, L. Decreased capacity for sperm production induced by perinatal bisphenol a exposure is associated with an increased inflammatory response in the offspring of C57BL/6 male mice. Int. J. Environ. Res. Public Health 2018, 15, 2158. [Google Scholar] [CrossRef] [PubMed]
- Valentino, R.; D’Esposito, V.; Ariemma, F.; Cimmino, I.; Beguinot, F.; Formisano, P. Bisphenol A environmental exposure and the detrimental effects on human metabolic health: Is it necessary to revise the risk assessment in vulnerable population? J. Endocrinol. Investig. 2016, 39, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Izumi, Y.; Yamaguchi, K.; Ishikawa, T.; Ando, M.; Chiba, K.; Hashimoto, H.; Shiotani, M.; Fujisawa, M. Molecular changes induced by bisphenol-A in rat Sertoli cell culture. Syst. Biol. Reprod. Med. 2011, 57, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Ogo, F.M.; de Lion Siervo, G.E.M.; Staurengo-Ferrari, L.; de Oliveira Mendes, L.; Luchetta, N.R.; Vieira, H.R.; Fattori, V.; Verri, W.A.; Scarano, W.R.; Fernandes, G.S.A. Bisphenol A Exposure Impairs Epididymal Development during the Peripubertal Period of Rats: Inflammatory Profile and Tissue Changes. Basic Clin. Pharmacol. Toxicol. 2018, 122, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Van’T Erve, T.J.; Rosen, E.M.; Barrett, E.S.; Nguyen, R.H.N.; Sathyanarayana, S.; Milne, G.L.; Calafat, A.M.; Swan, S.H.; Ferguson, K.K. Phthalates and Phthalate Alternatives Have Diverse Associations with Oxidative Stress and Inflammation in Pregnant Women. Environ. Sci. Technol. 2019, 53, 3258–3267. [Google Scholar] [CrossRef]
- Weinberger, B.; Vetrano, A.M.; Archer, F.E.; Marcella, S.W.; Wartenberg, D.; Robson, M.G.; Klim, J.; Azhar, S.; Wang, L.; Rich, D.Q. Effects of Maternal Exposure To Phthalates and Bisphenol A during Pregnancy on Gestational Age. J. Matern.-Fetal Neonatal Med. 2014, 27, 323–327. [Google Scholar] [CrossRef]
- Ferguson, K.K.; McElrath, T.F.; Mukherjee, B.; Loch-Caruso, R.; Meeker, J.D. Associations between maternal biomarkers of phthalate exposure and inflammation using repeated measurements across pregnancy. PLoS ONE 2015, 10, e0212958. [Google Scholar] [CrossRef]
- Eckstrum, K.S.; Weis, K.E.; Baur, N.G.; Yoshihara, Y.; Raetzman, L.T. Icam5 Expression Exhibits Sex Differences in the Neonatal Pituitary and Is Regulated by Estradiol and Bisphenol A. Endocrinology 2016, 157, 1408–1420. [Google Scholar] [CrossRef]
- Eckstrum, K.S.; Edwards, W.; Banerjee, A.; Wang, W.; Flaws, J.A.; Katzenellenbogen, J.A.; Kim, S.H.; Raetzman, L.T. Effects of Exposure to the Endocrine-Disrupting Chemical Bisphenol A During Critical Windows of Murine Pituitary Development. Endocrinology 2018, 159, 119–131. [Google Scholar] [CrossRef]
- Ullah, A.; Pirzada, M.; Jahan, S.; Ullah, H.; Razak, S.; Rauf, N.; Khan, M.J.; Mahboob, S.Z. Prenatal BPA and its analogs BPB, BPF, and BPS exposure and reproductive axis function in the male offspring of Sprague Dawley rats. Hum. Exp. Toxicol. 2019, 38, 1344–1365. [Google Scholar] [CrossRef]
- Gao, H.T.; Xu, R.; Cao, W.X.; Qian, L.L.; Wang, M.; Lu, L.; Xu, Q.; Yu, S.Q. Effects of six priority controlled phthalate esters with long-term low-dose integrated exposure on male reproductive toxicity in rats. Food Chem. Toxicol. 2017, 101, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Ha, M.; Guan, X.; Wei, L.; Li, P.; Yang, M.; Liu, C. Di-(2-ethylhexyl) phthalate inhibits testosterone level through disturbed hypothalamic-pituitary-testis axis and ERK-mediated 5α-Reductase 2. Sci. Total Environ. 2016, 563–564, 566–575. [Google Scholar] [CrossRef] [PubMed]
- Sobarzo, C.M.; Rosana, N.D.M.; Livia, L.; Berta, D.; Schteingart, H.F. Mono-(2-ethylhexyl) phthalate (MEHP) affects intercellular junctions of Sertoli cell: A potential role of oxidative stress. Reprod. Toxicol. 2015, 58, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Aly, H.A.A.; Hassan, M.H.; El-Beshbishy, H.A.; Alahdal, A.M.; Osman, A.M.M. Dibutyl phthalate induces oxidative stress and impairs spermatogenesis in adult rats. Toxicol. Ind. Health 2016, 32, 1467–1477. [Google Scholar] [CrossRef] [PubMed]
- Nelli, G.; Pamanji, S.R. Di-n-butyl phthalate prompts interruption of spermatogenesis, steroidogenesis, and fertility associated with increased testicular oxidative stress in adult male rats. Environ. Sci. Pollut. Res. 2017, 24, 18563–18574. [Google Scholar] [CrossRef] [PubMed]
- Sathyanarayana, S.; Grady, R.; Barrett, E.S.; Redmon, B.; Nguyen, R.H.N.; Barthold, J.S.; Bush, N.R.; Swan, S.H. First trimester phthalate exposure and male newborn genital anomalies. Environ. Res. 2016, 151, 777–782. [Google Scholar] [CrossRef]
- Hart, R.J.; Frederiksen, H.; Doherty, D.A.; Keelan, J.A.; Skakkebaek, N.E.; Minaee, N.S.; McLachlan, R.; Newnham, J.P.; Dickinson, J.E.; Pennell, C.E.; et al. The possible impact of antenatal exposure to ubiquitous phthalates upon male reproductive function at 20 years of age. Front. Endocrinol. (Lausanne) 2018, 9, 288. [Google Scholar] [CrossRef]
- Joensen, U.N.; Frederiksen, H.; Jensen, M.B.; Lauritsen, M.P.; Olesen, I.A.; Lassen, T.H.; Andersson, A.M.; Jørgensen, N. Phthalate excretion pattern and testicular function: A study of 881 healthy Danish men. Environ. Health Perspect. 2012, 120, 1397–1403. [Google Scholar] [CrossRef]
- Han, X.; Cui, Z.; Zhou, N.; Ma, M.; Li, L.; Li, Y.; Lin, H.; Ao, L.; Shu, W.; Liu, J.; et al. Urinary phthalate metabolites and male reproductive function parameters in Chongqing general population, China. Int. J. Hyg. Environ. Health 2014, 217, 271–278. [Google Scholar] [CrossRef]
- Lenters, V.; Portengen, L.; Smit, L.A.M.; Jönsson, B.A.G.; Giwercman, A.; Rylander, L.; Lindh, C.H.; Spanò, M.; Pedersen, H.S.; Ludwicki, J.K.; et al. Phthalates, perfluoroalkyl acids, metals and organochlorines and reproductive function: A multipollutant assessment in Greenlandic, Polish and Ukrainian men. Occup. Environ. Med. 2015, 72, 385–393. [Google Scholar] [CrossRef]
- Wang, Y.X.; Zeng, Q.; Sun, Y.; Yang, P.; Wang, P.; Li, J.; Huang, Z.; You, L.; Huang, Y.H.; Wang, C.; et al. Semen phthalate metabolites, semen quality parameters and serum reproductive hormones: A cross-sectional study in China. Environ. Pollut. 2016, 211, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Axelsson, J.; Rylander, L.; Rignell-Hydbom, A.; Jönsson, B.A.G.; Lindh, C.H.; Giwercman, A. Phthalate exposure and reproductive parameters in young men from the general Swedish population. Environ. Int. 2015, 85, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.X.; Zeng, Q.; Sun, Y.; You, L.; Wang, P.; Li, M.; Yang, P.; Li, J.; Huang, Z.; Wang, C.; et al. Phthalate exposure in association with serum hormone levels, sperm DNA damage and spermatozoa apoptosis: A cross-sectional study in China. Environ. Res. 2016, 150, 557–565. [Google Scholar] [CrossRef] [PubMed]
- Thurston, S.W.; Mendiola, J.; Bellamy, A.R.; Levine, H.; Wang, C.; Sparks, A.; Redmon, J.B.; Drobnis, E.Z.; Swan, S.H. Phthalate exposure and semen quality in fertile US men. Andrology 2016, 4, 632–638. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wang, H.; Tian, M.; Zhang, J.; Panuwet, P.; D’Souza, P.E.; Barr, D.B.; Huang, Q.; Xia, Y.; Shen, H. Phthalate metabolites related to infertile biomarkers and infertility in Chinese men. Environ. Pollut. 2017, 231, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Yang, H.; Zhou, N.; Sun, L.; Bao, H.; Tan, L.; Chen, H.; Ling, X.; Zhang, G.; Huang, L.; et al. Phthalate exposure, even below US EPA reference doses, was associated with semen quality and reproductive hormones: Prospective MARHCS study in general population. Environ. Int. 2017, 104, 58–68. [Google Scholar] [CrossRef]
- Al-Saleh, I.; Coskun, S.; Al-Doush, I.; Al-Rajudi, T.; Abduljabbar, M.; Al-Rouqi, R.; Palawan, H.; Al-Hassan, S. The relationships between urinary phthalate metabolites, reproductive hormones and semen parameters in men attending in vitro fertilization clinic. Sci. Total Environ. 2019, 658, 982–995. [Google Scholar] [CrossRef]
- Lassen, T.H.; Frederiksen, H.; Jensen, T.K.; Petersen, J.H.; Joensen, U.N.; Main, K.M.; Skakkebaek, N.E.; Juul, A.; Jørgensen, N.; Andersson, A.M. Urinary bisphenol a levels in young men: Association with reproductive hormones and semen quality. Environ. Health Perspect. 2014, 122, 478–484. [Google Scholar] [CrossRef]
- Knez, J.; Kranvogl, R.; Breznik, B.P.; Vončina, E.; Vlaisavljević, V. Are urinary bisphenol A levels in men related to semen quality and embryo development after medically assisted reproduction? Fertil. Steril. 2014, 101, 215–221. [Google Scholar] [CrossRef]
- Adoamnei, E.; Mendiola, J.; Vela-Soria, F.; Fernández, M.F.; Olea, N.; Jørgensen, N.; Swan, S.H.; Torres-Cantero, A.M. Urinary bisphenol A concentrations are associated with reproductive parameters in young men. Environ. Res. 2018, 161, 122–128. [Google Scholar] [CrossRef]
- Ji, H.; Miao, M.; Liang, H.; Shi, H.; Ruan, D.; Li, Y.; Wang, J.; Yuan, W. Exposure of environmental Bisphenol A in relation to routine sperm parameters and sperm movement characteristics among fertile men. Sci. Rep. 2018, 8, 17548. [Google Scholar] [CrossRef] [PubMed]
- Pollard, S.H.; Cox, K.J.; Blackburn, B.E.; Wilkins, D.G.; Carrell, D.T.; Stanford, J.B.; Porucznik, C.A. Male exposure to bisphenol A (BPA) and semen quality in the Home Observation of Periconceptional Exposures (HOPE) cohort. Reprod. Toxicol. 2019, 90, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.; Ko, D.H.; Lee, W.; Kim, K.R.; Chun, S.; Song, J.; Min, W.K. Body fluid concentrations of bisphenol A and their association with in vitro fertilization outcomes. Hum. Fertil. 2019, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bonde, J.P.; Flachs, E.M.; Rimborg, S.; Glazer, C.H.; Giwercman, A.; Ramlau-Hansen, C.H.; Hougaard, K.S.; Høyer, B.B.; Hærvig, K.K.; Petersen, S.B.; et al. The epidemiologic evidence linking prenatal and postnatal exposure to endocrine disrupting chemicals with male reproductive disorders: A systematic review and meta-analysis. Hum. Reprod. Update 2016, 23, 104–125. [Google Scholar] [CrossRef] [PubMed]
- Meeker, J.D.; Calafat, A.M.; Hauser, R. Urinary metabolites of di(2-ethylhexyl) phthalate are associated with decreased steroid hormone levels in adult men. J. Androl. 2010, 30, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Lambrot, R.; Muczynski, V.; Lecureuil, C.; Angenard, G.; Coffigny, H.; Pairault, C.; Moison, D.; Frydman, R.; Habert, R.; Rouiller-Fabre, V. Phthalates impair germ cell development in the human fetal testis in vitro without change in testosterone production. Environ. Health Perspect. 2009, 117, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Desdoits-Lethimonier, C.; Lesné, L.; Gaudriault, P.; Zalko, D.; Antignac, J.P.; Deceuninck, Y.; Platel, C.; Dejucq-Rainsford, N.; Mazaud-Guittot, S.; Jégou, B. Parallel assessment of the effects of bisphenol A and several of its analogs on the adult human testis. Hum. Reprod. 2017, 32, 1465–1473. [Google Scholar] [CrossRef]


{kind=link}
| Paper | Year | N subjects | EDCs investigated | Subjects with EDCs >LOD | Population | Sperm Parameters | Hormones |
|---|---|---|---|---|---|---|---|
| Joensen et al. [78] | 2012 | 881 | urinary MEP, MnBP, MiBP, MBzP, MEHP, MEHHP, MEOHP, MECPP, MOP, MCPP, MiNP, MHiNP, MOiNP, MCiOP | MOP 2.5%; others 79.6–100% | general population | semen parameters were not significantly associated with the urine concentrations of various phthalate esters | urinary MEHP was associated with total and free testosterone, but not with gonadotropins |
| Han et al. [79] | 2014 | 232 | MBP, MEP, MEHP, MBzP, phthalic acid | MBzP 8.9%; MEHP 58.9%; MEP 77.6%; others 100% | men with unknown fertility attending a fertility center | weak association between MBP and sperm concentration (OR 1.97; 95%CI 0.97–4.04); no associations with sperm DNA integrity | no correlations between phthalates and serum hormones after adjustments for confounders |
| Lenters et al. [80] | 2014 | 602 | DEHP, MEHHP, 5OH-MEHP, MEOHP, 5oxo-MEHP, MECPP, 5cx-MEPP, DiNP, MHiNP, 7OH-MMeOP, MOiNP, 7oxo-MMeOP, MOiCP, PFAS, PFOS, PFOA, PFHxS, PFNA, PFDA, PFUnDA, PFDoDA, cadmium, lead, mercury, PCB153, p,p’-DDE, HCB | phtalates 39–100%; metals 100%; perfluoroalkylic acids 29–100%; organochlorines 93–100%; | male partners of pregnant women | no associations between phthalates and sperm parameters; PCB-153 negatively associated with sperm motility | DiNP metabolites negatively associated with testosterone |
| Wang et al. [81] | 2015 | 687 | semen plasma MMP, MEP, MBP, MBzP, MEHP, MEHHP, MEOHP and MOP | MOP 13%; MBzP 29%; MMP 35%; others 67–100% | men with unknown fertility, attending a fertility clinic | negative associations with semen volume semen volume (MBP, MEHP, MEHHP, MEOHP), sperm kinetics parameters (MBzP, MEHP), normal forms (MBzP) | no associations with reproductive hormones |
| Axelsson et al. [82] | 2015 | 314 | urinary and serum MEHP, MECPP, MEHHP, MEOHP, MCiOP, MHiNP, MOiNP, MBP, MBzP, MEP | >97% | general population | DHEP metabolites and MECPP in particular were negatively associated with sperm motility; MEHP was positively associated with HDS. | N/A |
| Wang et al. [83] | 2016 | 1040 (483 reproductive hormones; 509 DNA integrity; 467 sperm apoptosis) | MMP, MEP, MBP, MBzP, MEHP, MEHHP, MEOHP, MEHP urinari | >90% | men with unknown fertility, attending a fertility clinic | MEHP, MEHHP, MEOHP: association with increased DNA fragmentation and apoptosis | MEHP: negative association with Estradiol, total and free Testosterone. |
| Thurston et al. [84] | 2016 | 420 | urinary MEHP, MEHHP, MEOHP, MECPP, MBP, MiBP, MCPP, MBzP, MEP | 77–100% | partners of pregnant women | MiBP: positive association with motility; MBzP: positive association with total sperm count | N/A |
| Liu et al. [85] | 2017 | 139 infertile vs. 150 fertile | urinary MMP, MEP, MBP, MBzP, MEHP, MEHHP, MEOHP | MMP 3.5%; MBzP 50.9%; others >99% | infertile vs. fertile subjects | MEHHP was significantly riduced in cases vs. fertile controls | N/A |
| Chen et al. [86] | 2017 | 796 | urinary MMP, MEP, MiBP, MnBP, MCHP, MCPP, MnOP, MEHHP, MECPP, MEOHP, MEHP, MBzP, MiNP | MCHP 2.5%; MiNP 17.3%; MnOP 32.5%; MBzP 35.6%; others >88% | general population | negative associations with sperm volume (MiBP, MEHP), concentration (MEP), motility (MEP, MnBP, MCPP, MnOP), normal forms (MEHP, MnOP, MBzP) | negative associations with estradiol (MnOP, MEHHP, MECPP, MEOHP), testosterone (MMP, MiBP, MnBP, MEOHP); positive associations with gonadotropins (MBzP) |
| Al-Saleh et al. [87] | 2019 | 599 | MEP, MiBP, MnBP, MBzP, MECPP, MEHHP, MEOHP, MEHP | MBzP: 26.1%; others: >96% | infertile | MECPP, MEHHP, MEOHP and calculated ΣDEHP: positive association with sperm concentration; calculated excreted %MEHP: negative association with sperm concentration | MiBP and MEHHP were inversely associated with testosterone and FSH, respectively. Estradiol was positively associated with MEP. %MEHP was positively associated with gonadotropins (FSH, LH) |
| Paper | Year | N subjects | EDCs investigated | Subjects with EDCs >LOD | Population | Sperm Parameters | Hormones |
|---|---|---|---|---|---|---|---|
| Lassen et al. [88] | 2014 | 308 | urinary BPA | 98% | general population | highest BPA concentration quartile had reduced % progressive motility vs. lower quartile. | highest BPA quartiles had higher LH, testosterone and estradiol compared to lower quartile. |
| Knez et al. [89] | 2014 | 149 | urinary BPA | 98% | infertile | BPA: negative association with sperm concentration and vitality | N/A |
| Adoamnei et al. [90] | 2018 | 215 | urinary BPA | 95% | healthy volounteers | negative association between BPA and sperm concentration, total sperm count | positive association between BPA and LH |
| Ji et al. [91] | 2018 | 500 | urinary BPA | 73.6% | fertile | BPA was associated with several CASA kinetics parameters (positive associations: LIN, STR, WOB; negative associations: ALH)) | N/A |
| Pollard et al. [92] | 2019 | 161 | urinary BPA | 87% | men with unknown fertility | abnormal morphology was associated with higher BPA mean urinary concentrations | N/A |
| Kim et al. [93] | 2019 | 146 | Urinary, semen plasma, blood BPA | 43.1% (urine); 71.3% (semen plasma); 77.6% (blood) | infertile | no associations with sperm concentration and motility; no significant effect on embryo embryo quality, oocytes retrieved, pregnancy | N/A |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pallotti, F.; Pelloni, M.; Gianfrilli, D.; Lenzi, A.; Lombardo, F.; Paoli, D. Mechanisms of Testicular Disruption from Exposure to Bisphenol A and Phtalates. J. Clin. Med. 2020, 9, 471. https://doi.org/10.3390/jcm9020471
Pallotti F, Pelloni M, Gianfrilli D, Lenzi A, Lombardo F, Paoli D. Mechanisms of Testicular Disruption from Exposure to Bisphenol A and Phtalates. Journal of Clinical Medicine. 2020; 9(2):471. https://doi.org/10.3390/jcm9020471
Chicago/Turabian StylePallotti, Francesco, Marianna Pelloni, Daniele Gianfrilli, Andrea Lenzi, Francesco Lombardo, and Donatella Paoli. 2020. "Mechanisms of Testicular Disruption from Exposure to Bisphenol A and Phtalates" Journal of Clinical Medicine 9, no. 2: 471. https://doi.org/10.3390/jcm9020471
APA StylePallotti, F., Pelloni, M., Gianfrilli, D., Lenzi, A., Lombardo, F., & Paoli, D. (2020). Mechanisms of Testicular Disruption from Exposure to Bisphenol A and Phtalates. Journal of Clinical Medicine, 9(2), 471. https://doi.org/10.3390/jcm9020471