Deregulation of miRNA in Helicobacter pylori-Induced Gastric MALT Lymphoma: From Mice to Human



Abstract
1. Introduction
2. Materials and Methods
2.1. Patient Samples
2.2. RNA Extraction
2.3. miRNA Reverse-Transcription
2.4. PCR Array
2.5. Quantitative Real-Time PCR
2.6. Statistical Analysis
3. Results
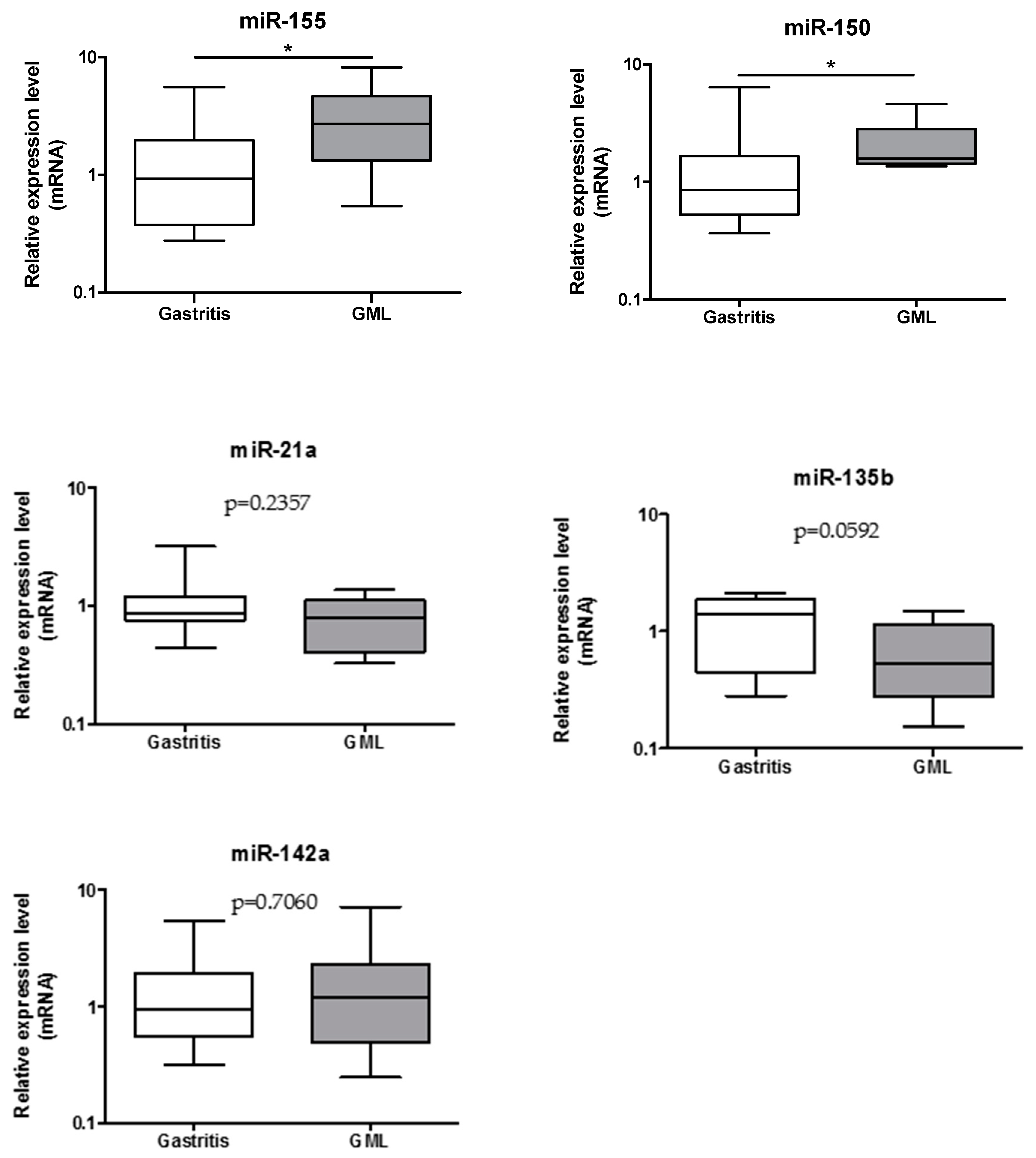
3.1. Relative Expression Levels of miR-155, miR-150, miR-21a, miR-135 and miR-142a in gastric MALT Lymphomas and Gastritis Patients
3.2. Investigation of miRNAs Expression by PCR Array
3.3. Validation of the Deregulation of the miRNAs in Gastric MALT Lymphoma
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zucca, E.; Copie-Bergman, C.; Ricardi, U.; Thieblemont, C.; Raderer, M.; Ladetto, M.; on behalf of the ESMO Guidelines Working Group. Gastric marginal zone lymphoma of MALT type: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2013, 24, vi144–vi148. [Google Scholar] [CrossRef] [PubMed]
- Zullo, A.; Hassan, C.; Ridola, L.; Repici, A.; Manta, R.; Andriani, A. Gastric MALT lymphoma: Old and new insights. Ann. Gastroenterol. 2014, 27, 27–33. [Google Scholar] [PubMed]
- Fischbach, W.; Goebeler-Kolve, M.-E.; Dragosics, B.; Greiner, A.; Stolte, M. Long term outcome of patients with gastric marginal zone B cell lymphoma of mucosa associated lymphoid tissue (MALT) following exclusive Helicobacter pylori eradication therapy: Experience from a large prospective series. Gut 2004, 53, 34–37. [Google Scholar] [CrossRef] [PubMed]
- Ullrich, A.; Fischbach, W.; Blettner, M. Incidence of gastric B-cell lymphomas: A population-based study in Germany. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2002, 13, 1120–1127. [Google Scholar] [CrossRef] [PubMed]
- Kusters, J.G.; van Vliet, A.H.M.; Kuipers, E.J. Pathogenesis of Helicobacter pylori Infection. Clin. Microbiol. Rev. 2006, 19, 449–490. [Google Scholar] [CrossRef] [PubMed]
- Vasilatou, D.; Sioulas, A.D.; Pappa, V.; Papanikolaou, I.S.; Triantafyllou, K.; Dimitriadis, G.D.; Papageorgiou, S.G. The role of miRNAs and epigenetic mechanisms in primary gastric mucosa-associated lymphoid tissue lymphoma. Future Oncol. 2016, 12, 1587–1593. [Google Scholar] [CrossRef] [PubMed]
- Bartels, C.L.; Tsongalis, G.J. MicroRNAs: Novel Biomarkers for Human Cancer. Clin. Chem. 2009, 55, 623–631. [Google Scholar] [CrossRef] [PubMed]
- Lawrie, C.H. microRNA expression in lymphoid malignancies: New hope for diagnosis and therapy? J. Cell. Mol. Med. 2008, 12, 1432–1444. [Google Scholar] [CrossRef]
- Thorns, C.; Kuba, J.; Bernard, V.; Senft, A.; Szymczak, S.; Feller, A.C.; Bernd, H.-W. Deregulation of a distinct set of microRNAs is associated with transformation of gastritis into MALT lymphoma. Virchows Arch. 2012, 460, 371–377. [Google Scholar] [CrossRef] [PubMed]
- Saito, Y.; Suzuki, H.; Tsugawa, H.; Imaeda, H.; Matsuzaki, J.; Hirata, K.; Hosoe, N.; Nakamura, M.; Mukai, M.; Saito, H.; et al. Overexpression of miR-142-5p and miR-155 in Gastric Mucosa-Associated Lymphoid Tissue (MALT) Lymphoma Resistant to Helicobacter pylori Eradication. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Fernández, C.; Bellosillo, B.; Ferraro, M.; Seoane, A.; Sánchez-González, B.; Pairet, S.; Pons, A.; Barranco, L.; Vela, M.C.; Gimeno, E.; et al. MicroRNAs 142-3p, miR-155 and miR-203 Are Deregulated in Gastric MALT Lymphomas Compared to Chronic Gastritis. Cancer Genomics Proteomics 2017, 14, 75–82. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Craig, V.J.; Cogliatti, S.B.; Rehrauer, H.; Wündisch, T.; Müller, A. Epigenetic Silencing of MicroRNA-203 Dysregulates ABL1 Expression and Drives Helicobacter-Associated Gastric Lymphomagenesis. Cancer Res. 2011, 71, 3616–3624. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, A.; E, M.; Quan, L.; Qu, Y.; Gu, A. Three novel microRNAs based on microRNA signatures for gastric mucosa-associated lymphoid tissue lymphoma. Neoplasma 2018, 65, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Floch, P.; Capdevielle, C.; Staedel, C.; Izotte, J.; Sifré, E.; Laur, A.M.; Giese, A.; Korolik, V.; Dubus, P.; Mégraud, F.; et al. Deregulation of MicroRNAs in Gastric Lymphomagenesis Induced in the d3Tx Mouse Model of Helicobacter pylori Infection. Front. Cell. Infect. Microbiol. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Chrisment, D.; Dubus, P.; Chambonnier, L.; Hocès de la Guardia, A.; Sifré, E.; Giese, A.; Capone, M.; Khairallah, C.; Costet, P.; Rousseau, B.; et al. Neonatal Thymectomy Favors Helicobacter pylori–Promoted Gastric Mucosa-Associated Lymphoid Tissue Lymphoma Lesions in BALB/c Mice. Am. J. Pathol. 2014, 184, 2174–2184. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Swerdlow, S.H.; Campo, E.; Harris, N.L.; Jaffe, E.S.; Pileri, S.A.; Stein, H.; Thiele, J. WHO Classification of Tumours of Haematopoietic and Lymphoid Tissues, Revised 4th ed.; International Agency for Research on Cancer (IARC): Lyon, France, 2017; Volume 2, ISBN 978-92-832-4494-3. [Google Scholar]
- Amiot, A.; Lévy, M.; Copie-Bergman, C.; Dupuis, J.; Szablewski, V.; Le Baleur, Y.; Baia, M.; Belhadj, K.; Sobhani, I.; Leroy, K.; et al. Rituximab, alkylating agents or combination therapy for gastric mucosa-associated lymphoid tissue lymphoma: A monocentric non-randomised observational study. Aliment. Pharmacol. Ther. 2014, 39, 619–628. [Google Scholar] [CrossRef]
- Tankovic, J.; Lamarque, D.; Lascols, C.; Soussy, C.J.; Delchier, J.C. Clarithromycin resistance of Helicobacter pylori has a major impact on the efficacy of the omeprazole-amoxicillin-clarithromycin therapy. Pathol. Biol. (Paris) 2001, 49, 528–533. [Google Scholar] [CrossRef]
- Ferreri, A.J.M.; Ernberg, I.; Copie-Bergman, C. Infectious agents and lymphoma development: Molecular and clinical aspects. J. Intern. Med. 2009, 265, 421–438. [Google Scholar] [CrossRef]
- Zhang, X.; Zhu, W.; Zhang, J.; Huo, S.; Zhou, L.; Gu, Z.; Zhang, M. MicroRNA-650 targets ING4 to promote gastric cancer tumorigenicity. Biochem. Biophys. Res. Commun. 2010, 395, 275–280. [Google Scholar] [CrossRef]
- Han, L.L.; Yin, X.R.; Zhang, S.Q. miR-650 Promotes the Metastasis and Epithelial–Mesenchymal Transition of Hepatocellular Carcinoma by Directly Inhibiting LATS2 Expression. Cell. Physiol. Biochem. 2018, 51, 1179–1192. [Google Scholar] [CrossRef]
- Zeng, Z.; Li, F.; Gao, F.; Sun, D.; Yao, L. Upregulation of miR-650 is correlated with down-regulation of ING4 and progression of hepatocellular carcinoma. J. Surg. Oncol. 2013, 107, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Liu, X.; Li, J.; Yang, J.; Zhang, E.; Yin, D.; Liu, Z.; Zhou, J.; Ding, Y.; Li, S.; et al. MiR-196a Is Upregulated in Gastric Cancer and Promotes Cell Proliferation by Downregulating p27kip1. Mol. Cancer Ther. 2012, 11, 842–852. [Google Scholar] [CrossRef] [PubMed]
- Xin, H.; Wang, C.; Liu, Z. miR-196a-5p promotes metastasis of colorectal cancer via targeting IκBα. BMC Cancer 2019, 19. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhang, W.; Ji, W. miR-196b is a prognostic factor of human laryngeal squamous cell carcinoma and promotes tumor progression by targeting SOCS2. Biochem. Biophys. Res. Commun. 2018, 501, 584–592. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Pan, J.; Lu, X.; Chi, P. Role of miR-196 and its target gene HoxB8 in the development and proliferation of human colorectal cancer and the impact of neoadjuvant chemotherapy with FOLFOX4 on their expression. Oncol. Lett. 2016, 12, 4041–4047. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Suo, A.-L.; Li, Z.-F.; Liu, L.-Y.; Tian, T.; Ni, L.; Zhang, W.-G.; Nan, K.-J.; Song, T.-S.; Huang, C. MicroRNA profiling of human gastric cancer. Mol. Med. Rep. 2009, 2, 963–970. [Google Scholar] [PubMed]
- Jiang, D.; Zhou, B.; Xiong, Y.; Cai, H. miR-135 regulated breast cancer proliferation and epithelial-mesenchymal transition acts by the Wnt/β-catenin signaling pathway. Int. J. Mol. Med. 2019. [Google Scholar] [CrossRef] [PubMed]
- Navarro, A.; Diaz, T.; Martinez, A.; Gaya, A.; Pons, A.; Gel, B.; Codony, C.; Ferrer, G.; Martinez, C.; Montserrat, E.; et al. Regulation of JAK2 by miR-135a: Prognostic impact in classic Hodgkin lymphoma. Blood 2009, 114, 2945–2951. [Google Scholar] [CrossRef]
- Wu, H.; Huang, M.; Cao, P.; Wang, T.; Shu, Y.; Liu, P. MiR-135a targets JAK2 and inhibits gastric cancer cell proliferation. Cancer Biol. Ther. 2012, 13, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Dou, W.; He, L.; Liang, S.; Tie, J.; Liu, C.; Li, T.; Lu, Y.; Mo, P.; Shi, Y.; et al. MicroRNA-7 functions as an anti-metastatic microRNA in gastric cancer by targeting insulin-like growth factor-1 receptor. Oncogene 2013, 32, 1363–1372. [Google Scholar] [CrossRef]
- Xie, J.; Chen, M.; Zhou, J.; Mo, M.-S.; Zhu, L.-H.; Liu, Y.-P.; Gui, Q.-J.; Zhang, L.; Li, G.-Q. miR-7 inhibits the invasion and metastasis of gastric cancer cells by suppressing epidermal growth factor receptor expression. Oncol. Rep. 2014, 31, 1715–1722. [Google Scholar] [CrossRef] [PubMed]
- Kong, D.; Piao, Y.-S.; Yamashita, S.; Oshima, H.; Oguma, K.; Fushida, S.; Fujimura, T.; Minamoto, T.; Seno, H.; Yamada, Y.; et al. Inflammation-induced repression of tumor suppressor miR-7 in gastric tumor cells. Oncogene 2012, 31, 3949–3960. [Google Scholar] [CrossRef] [PubMed]
- Tagscherer, K.E.; Fassl, A.; Sinkovic, T.; Richter, J.; Schecher, S.; Macher-Goeppinger, S.; Roth, W. MicroRNA-210 induces apoptosis in colorectal cancer via induction of reactive oxygen. Cancer Cell Int. 2016, 16. [Google Scholar] [CrossRef]
- Cheng, Z.; Geng, H.; Cheng, Y.; Dong, N.; Ning, F.; Yu, Z.; Jian, J.; Chen, S. Effects of MiR-210 on proliferation, apoptosis and invasion abilities of esophageal cancer cells. J. BUON 2018, 23, 814–819. [Google Scholar] [PubMed]
- Tsuchiya, S.; Fujiwara, T.; Sato, F.; Shimada, Y.; Tanaka, E.; Sakai, Y.; Shimizu, K.; Tsujimoto, G. MicroRNA-210 Regulates Cancer Cell Proliferation through Targeting Fibroblast Growth Factor Receptor-like 1 (FGFRL1). J. Biol. Chem. 2011, 286, 420–428. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, Y.; Yuan, W.; Qiu, S. MicroRNA-153 functions as a tumor suppressor in gastric cancer via targeting Kruppel-like factor 5. Exp. Ther. Med. 2018, 16, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Sun, J.; Bai, Z.; Li, H.; He, S.; Chen, R.; Che, X. MicroRNA-153 acts as a prognostic marker in gastric cancer and its role in cell migration and invasion. OncoTargets Ther. 2015, 8, 357–364. [Google Scholar]
- Li, G.; Xue, M.; Yang, F.; Jin, Y.; Fan, Y.; Li, W. CircRBMS3 promotes gastric cancer tumorigenesis by regulating miR-153–SNAI1 axis. J. Cell. Physiol. 2019, 234, 3020–3028. [Google Scholar] [CrossRef]
- He, Y.; Jiang, X.; Chen, J. The role of miR-150 in normal and malignant hematopoiesis. Oncogene 2014, 33, 3887–3893. [Google Scholar] [CrossRef]
- Wu, S.J.; Chen, J.; Wu, B.; Wang, Y.J.; Guo, K.Y. MicroRNA-150 enhances radiosensitivity by inhibiting the AKT pathway in NK/T cell lymphoma. J. Exp. Clin. Cancer Res. 2018, 37. [Google Scholar] [CrossRef]
- Xiao, C.; Calado, D.P.; Galler, G.; Thai, T.-H.; Patterson, H.C.; Wang, J.; Rajewsky, N.; Bender, T.P.; Rajewsky, K. MiR-150 Controls B Cell Differentiation by Targeting the Transcription Factor c-Myb. Cell 2007, 131, 146–159. [Google Scholar] [CrossRef] [PubMed]
- Gebauer, N.; Kuba, J.; Senft, A.; Schillert, A.; Bernard, V.; Thorns, C. MicroRNA-150 Is up-regulated in Extranodal Marginal Zone Lymphoma of MALT Type. Cancer Genomics - Proteomics 2014, 11, 51–56. [Google Scholar] [PubMed]
- Wu, Q.; Jin, H.; Yang, Z.; Luo, G.; Lu, Y.; Li, K.; Ren, G.; Su, T.; Pan, Y.; Feng, B.; et al. MiR-150 promotes gastric cancer proliferation by negatively regulating the pro-apoptotic gene EGR2. Biochem. Biophys. Res. Commun. 2010, 392, 340–345. [Google Scholar] [CrossRef] [PubMed]
- Lawrie, C.H. MicroRNAs and lymphomagenesis: A functional review. Br. J. Haematol. 2013, 160, 571–581. [Google Scholar] [CrossRef] [PubMed]
- Tagawa, H.; Ikeda, S.; Sawada, K. Role of microRNA in the pathogenesis of malignant lymphoma. Cancer Sci. 2013, 104, 801–809. [Google Scholar] [CrossRef] [PubMed]
- Tili, E.; Michaille, J.-J.; Croce, C.M. MicroRNAs play a central role in molecular dysfunctions linking inflammation with cancer. Immunol. Rev. 2013, 253, 167–184. [Google Scholar] [CrossRef] [PubMed]
- Babar, I.A.; Cheng, C.J.; Booth, C.J.; Liang, X.; Weidhaas, J.B.; Saltzman, W.M.; Slack, F.J. Nanoparticle-based therapy in an in vivo microRNA-155 (miR-155)-dependent mouse model of lymphoma. Proc. Natl. Acad. Sci. USA 2012, 109, E1695–E1704. [Google Scholar] [CrossRef] [PubMed]
- Oertli, M.; Engler, D.B.; Kohler, E.; Koch, M.; Meyer, T.F.; Muller, A. MicroRNA-155 Is Essential for the T Cell-Mediated Control of Helicobacter pylori Infection and for the Induction of Chronic Gastritis and Colitis. J. Immunol. 2011, 187, 3578–3586. [Google Scholar] [CrossRef]
- O’Connell, R.M.; Kahn, D.; Gibson, W.S.J.; Round, J.L.; Scholz, R.L.; Chaudhuri, A.A.; Kahn, M.E.; Rao, D.S.; Baltimore, D. MicroRNA-155 promotes autoimmune inflammation by enhancing inflammatory T cell development. Immunity 2010, 33, 607–619. [Google Scholar] [CrossRef]
- Xiao, B.; Liu, Z.; Li, B.-S.; Tang, B.; Li, W.; Guo, G.; Shi, Y.; Wang, F.; Wu, Y.; Tong, W.-D.; et al. Induction of microRNA-155 during Helicobacter pylori Infection and Its Negative Regulatory Role in the Inflammatory Response. J. Infect. Dis. 2009, 200, 916–925. [Google Scholar] [CrossRef]
- Hocès de la Guardia, A.; Staedel, C.; Kaafarany, I.; Clément, A.; Roubaud Baudron, C.; Mégraud, F.; Lehours, P. Inflammatory cytokine and microRNA responses of primary human dendritic cells cultured with Helicobacter pylori strains. Front. Microbiol. 2013, 4. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Deng, Z.; Wang, Z.; Wu, J.; Gu, T.; Jiang, Y.; Li, G. MicroRNA-155 in exosomes secreted from Helicobacter pylori infection macrophages immunomodulates inflammatory response. Am. J. Transl. Res. 2016, 8, 3700–3709. [Google Scholar] [PubMed]
- Koch, M.; Mollenkopf, H.-J.; Klemm, U.; Meyer, T.F. Induction of microRNA-155 is TLR- and type IV secretion system-dependent in macrophages and inhibits DNA-damage induced apoptosis. Proc. Natl. Acad. Sci. USA 2012, 109, E1153–E1162. [Google Scholar] [CrossRef] [PubMed]
- Du, M.-Q. MALT lymphoma: Many roads lead to nuclear factor-κb activation. Histopathology 2011. Available online: https://onlinelibrary.wiley.com/doi/full/10.1111/j.1365-2559.2010.03699.x (accessed on 24 April 2019). [CrossRef] [PubMed]
- O’Connell, R.M.; Chaudhuri, A.A.; Rao, D.S.; Baltimore, D. Inositol phosphatase SHIP1 is a primary target of miR-155. Proc. Natl. Acad. Sci. USA 2009, 106, 7113–7118. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Shen, Y.; Liu, M.; Bi, C.; Jiang, C.; Iqbal, J.; McKeithan, T.W.; Chan, W.C.; Ding, S.-J.; Fu, K. Quantitative Proteomics Reveals that miR-155 Regulates the PI3K-AKT Pathway in Diffuse Large B-Cell Lymphoma. Am. J. Pathol. 2012, 181, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Tomasini, R.; Seux, M.; Nowak, J.; Bontemps, C.; Carrier, A.; Dagorn, J.-C.; Pébusque, M.-J.; Iovanna, J.L.; Dusetti, N.J. TP53INP1 is a novel p73 target gene that induces cell cycle arrest and cell death by modulating p73 transcriptional activity. Oncogene 2005, 24, 8093–8104. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, Z.; Li, Y.; Zang, A. MicroRNA and signaling pathways in gastric cancer. Cancer Gene Ther. 2014, 21, 305–316. [Google Scholar] [CrossRef]
- Chen, C.; Zhang, Y.; Zhang, L.; Weakley, S.M.; Yao, Q. MicroRNA-196: Critical roles and clinical applications in development and cancer. J. Cell. Mol. Med. 2011, 15, 14–23. [Google Scholar] [CrossRef]
- Lu, Y.-C.; Chang, J.T.; Chan, E.-C.; Chao, Y.-K.; Yeh, T.-S.; Chen, J.-S.; Cheng, A.-J. miR-196, an Emerging Cancer Biomarker for Digestive Tract Cancers. J. Cancer 2016, 7, 650–655. [Google Scholar] [CrossRef]
- Lee, J.W.; Kim, N.; Park, J.H.; Kim, H.J.; Chang, H.; Kim, J.M.; Kim, J.-W.; Lee, D.H. Differential MicroRNA Expression Between Gastric Cancer Tissue and Non-cancerous Gastric Mucosa According to Helicobacter pylori Status. J. Cancer Prev. 2017, 22, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, W.; Liu, K.; Liu, S.; Ji, B.; Wang, Y. miR-138 suppresses cell proliferation and invasion by inhibiting SOX9 in hepatocellular carcinoma. Am. J. Transl. Res. 2016, 8, 2159–2168. [Google Scholar] [PubMed]
- Tang, X.; Jiang, J.; Zhu, J.; He, N.; Tan, J. HOXA4-regulated miR-138 suppresses proliferation and gefitinib resistance in non-small cell lung cancer. Mol. Genet. Genomics MGG 2019, 294, 85–93. [Google Scholar] [CrossRef] [PubMed]
- You, C.; Jin, L.; Xu, Q.; Shen, B.; Jiao, X.; Huang, X. Expression of miR-21 and miR-138 in colon cancer and its effect on cell proliferation and prognosis. Oncol. Lett. 2019, 17, 2271–2277. [Google Scholar] [CrossRef] [PubMed]
- Qu, M.; Zhu, Y.; Jin, M. MicroRNA-138 inhibits SOX12 expression and the proliferation, invasion and migration of ovarian cancer cells. Exp. Ther. Med. 2018, 16, 1629–1638. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Pan, Z.-G.; Shu, L.; Li, Q.-J. Podocalyxin-like, targeted by miR-138, promotes colorectal cancer cell proliferation, migration, invasion and EMT. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 8664–8674. [Google Scholar]
- Pang, L.; Li, B.; Zheng, B.; Niu, L.; Ge, L. miR-138 inhibits gastric cancer growth by suppressing SOX4. Oncol. Rep. 2017, 38, 1295–1302. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, C. MiR-153 regulates metastases of gastric cancer through Snail. Tumor Biol. 2016, 37, 15509–15515. [Google Scholar] [CrossRef]
- Yuan, Y.; Du, W.; Wang, Y.; Xu, C.; Wang, J.; Zhang, Y.; Wang, H.; Ju, J.; Zhao, L.; Wang, Z.; et al. Suppression of AKT expression by miR-153 produced anti-tumor activity in lung cancer. Int. J. Cancer 2015, 136, 1333–1340. [Google Scholar] [CrossRef]
- Masuda, M.; Miki, Y.; Hata, S.; Takagi, K.; Sakurai, M.; Ono, K.; Suzuki, K.; Yang, Y.; Abe, E.; Hirakawa, H.; et al. An induction of microRNA, miR-7 through estrogen treatment in breast carcinoma. J. Transl. Med. 2012, 10, S2. [Google Scholar] [CrossRef]
- Liu, Z.; Jiang, Z.; Huang, J.; Huang, S.; Li, Y.; Yu, S.; Yu, S.; Liu, X. miR-7 inhibits glioblastoma growth by simultaneously interfering with the PI3K/ATK and Raf/MEK/ERK pathways. Int. J. Oncol. 2014, 44, 1571–1580. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Shi, X.; Li, C.; Wang, X.; Hou, K.; Li, Z.; Zhang, X.; Fan, Y.; Qu, X.; Che, X.; et al. Long non-coding RNA UCA1 upregulation promotes the migration of hypoxia-resistant gastric cancer cells through the miR-7-5p/EGFR axis. Exp. Cell Res. 2018, 368, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Li, T.; Jiang, Y.; Pan, C.; Ding, Y.; Huang, Z.; Yu, H.; Kong, D. Overexpression of Circular RNA ciRS-7 Abrogates the Tumor Suppressive Effect of miR-7 on Gastric Cancer via PTEN/PI3K/AKT Signaling Pathway. J. Cell. Biochem. 2018, 119, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Kuo, S.-H.; Yeh, K.-H.; Chen, L.-T.; Lin, C.-W.; Hsu, P.-N.; Wu, M.-S.; Liou, J.-M.; Tsai, H.-J.; Tzeng, Y.-S.; Cheng, A.-L. Helicobacter pylori CagA Translocation Is Closely Associated with the Expression of CagA-signaling Molecules in Low-grade Gastric Mucosa-associated Lymphoid Tissue Lymphoma. Am. J. Surg. Pathol. 2015, 39, 761–766. [Google Scholar] [CrossRef] [PubMed]
- Floch, P.; Mégraud, F.; Lehours, P. Helicobacter pylori Strains and Gastric MALT Lymphoma. Toxins 2017, 9, 132. [Google Scholar] [CrossRef] [PubMed]
- Kuo, S.-H.; Wu, M.-S.; Yeh, K.-H.; Lin, C.-W.; Hsu, P.-N.; Chen, L.-T.; Cheng, A.-L. Novel Insights of Lymphomagenesis of Helicobacter pylori-Dependent Gastric Mucosa-Associated Lymphoid Tissue Lymphoma. Cancers 2019, 11, 547. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| miRNA | Fold-Regulation Value |
|---|---|
| miR-650 | 6.8305 |
| miR-196a-5p | 6.242 |
| miR-142-5p | 3.9504 |
| miR-138-5p | 3.9231 |
| miR-601 | 3.5357 |
| miR-196b-5p | 3.2761 |
| miR-141-3p | −3.0272 |
| miR-200a-5p | −3.0483 |
| miR-106b-5p | −3.1558 |
| miR-301a+3p | −3.1558 |
| miR-30a-5p | −3.1998 |
| miR-192-5p | −3.3357 |
| miR-192-3p | −4.193 |
| miR-20a-3p | −4.2222 |
| miR-429 | −4.3109 |
| miR-22-5p | −4.5567 |
| miR-335-5p | −4.8501 |
| miR-95-3p | −4.952 |
| miR-190a-5p | −6.1817 |
| miR-203a-3p | −6.6714 |
| miR-135b-5p | −6.859 |
| miR-205-5p | −6.859 |
| miR-153-3p | −7.6635 |
| miR-210-3p | −7.6635 |
| miR-7-5p | −7.7168 |
| miR-135a-5p | −9.305 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blosse, A.; Levy, M.; Robe, C.; Staedel, C.; Copie-Bergman, C.; Lehours, P. Deregulation of miRNA in Helicobacter pylori-Induced Gastric MALT Lymphoma: From Mice to Human. J. Clin. Med. 2019, 8, 845. https://doi.org/10.3390/jcm8060845
Blosse A, Levy M, Robe C, Staedel C, Copie-Bergman C, Lehours P. Deregulation of miRNA in Helicobacter pylori-Induced Gastric MALT Lymphoma: From Mice to Human. Journal of Clinical Medicine. 2019; 8(6):845. https://doi.org/10.3390/jcm8060845
Chicago/Turabian StyleBlosse, Alice, Michael Levy, Cyrielle Robe, Cathy Staedel, Christiane Copie-Bergman, and Philippe Lehours. 2019. "Deregulation of miRNA in Helicobacter pylori-Induced Gastric MALT Lymphoma: From Mice to Human" Journal of Clinical Medicine 8, no. 6: 845. https://doi.org/10.3390/jcm8060845
APA StyleBlosse, A., Levy, M., Robe, C., Staedel, C., Copie-Bergman, C., & Lehours, P. (2019). Deregulation of miRNA in Helicobacter pylori-Induced Gastric MALT Lymphoma: From Mice to Human. Journal of Clinical Medicine, 8(6), 845. https://doi.org/10.3390/jcm8060845