Stem Cells on Biomaterials for Synthetic Grafts to Promote Vascular Healing

 ,
,  ,
,
Abstract
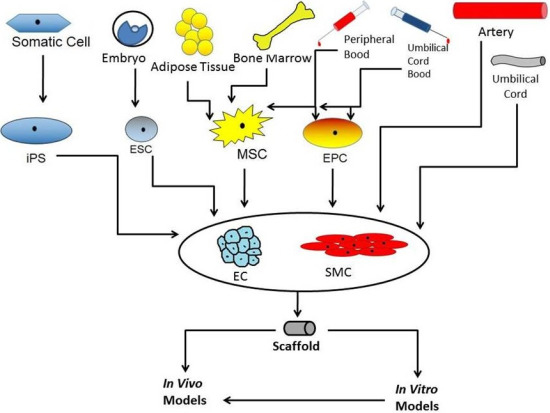
:
1. Introduction
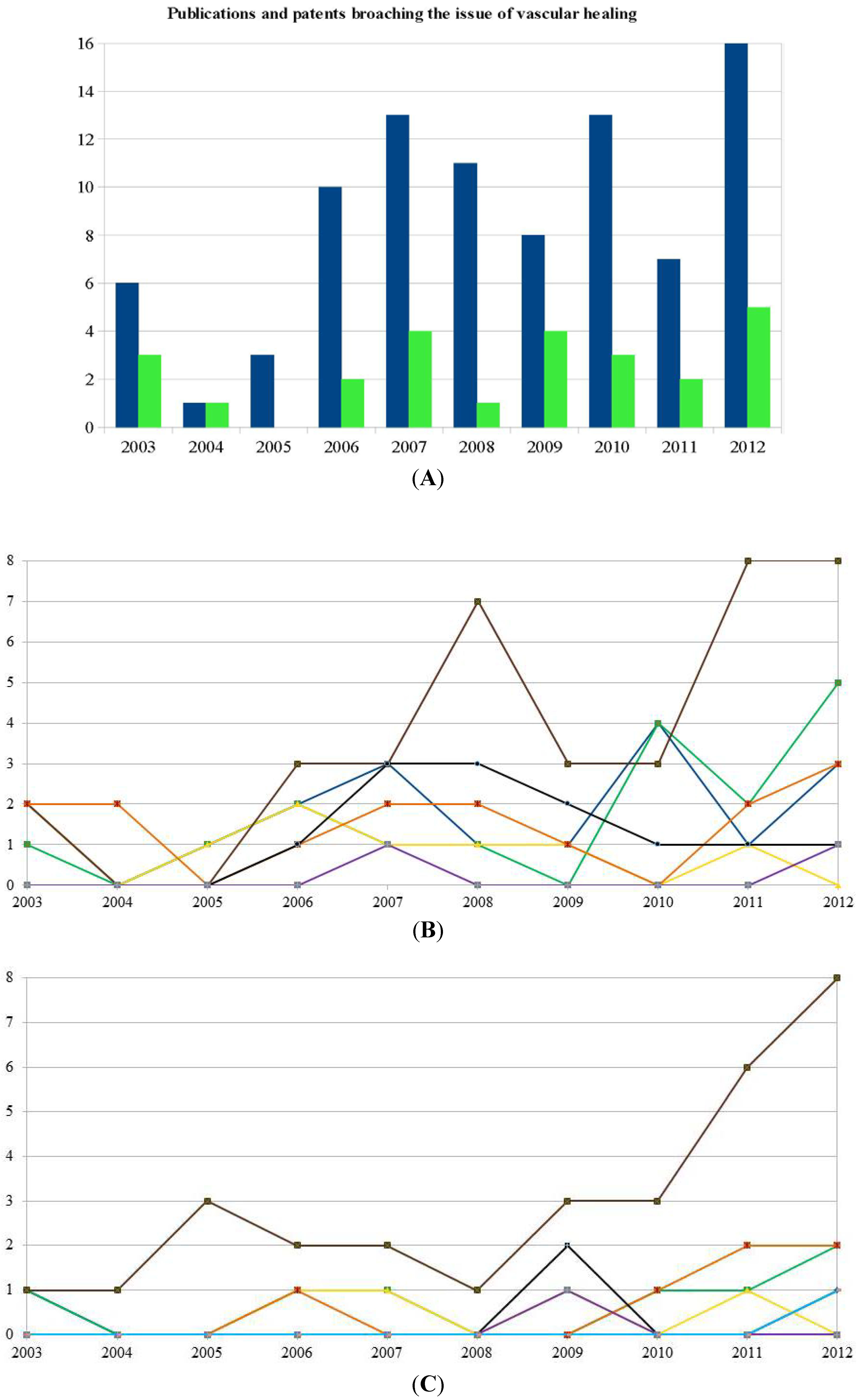
2. Stem Cells for Vascular Grafts
2.1. Embryonic Stem Cells
2.1.1. Endothelial Cell and Smooth Muscle Cell Markers Expressed in Differentiated ESCs
2.1.2. Embryonic Stem Cells for in Vitro 3D Culture Models
2.1.3. Embryonic Stem Cells for Vascular Grafts in Animal Models
2.1.4. Patents on Embryonic Stem Cells for Vascular Grafts
2.2. Induced Pluripotent Stem Cells
Induced Pluripotent Stem Cells for Vascular Grafts
2.3. Mesenchymal Stem Cells
2.3.1. Endothelial Cell and Smooth Muscle Cell Markers Expressed in Differentiated MSCs
2.3.2. MSCs for in Vitro Differentiation Assays
2.3.3. MSCs for Vascular Grafts in Animal Models
2.4. Endothelial and Smooth Muscle Progenitor Cells
2.4.1. EC and SMC Expressed in Differentiation Assays of Vascular Precursor Cells
2.4.2. Endothelial and Smooth Muscle Progenitor Cells for Vascular Grafts in Animal Models
2.5. Patents on Adult Stem and Precursor Cell Isolation and Maintenance
3. Primary Tissue Cells for Vascular Grafts
3.1. Endothelial Cells
3.2. Smooth Muscle Cells
3.3. Endothelial Cells and Smooth Muscle Cells for Vascular Grafts
Patents on Endothelial Cells and Smooth Muscle Cells for Vascular Grafts
3.4. Other Primary Tissue Cells for Vascular Grafts
4. Summary of Cell Types for Vascular Grafts
5. Scaffolds for Stem Cell Engineering

5.1. Scaffolds Materials
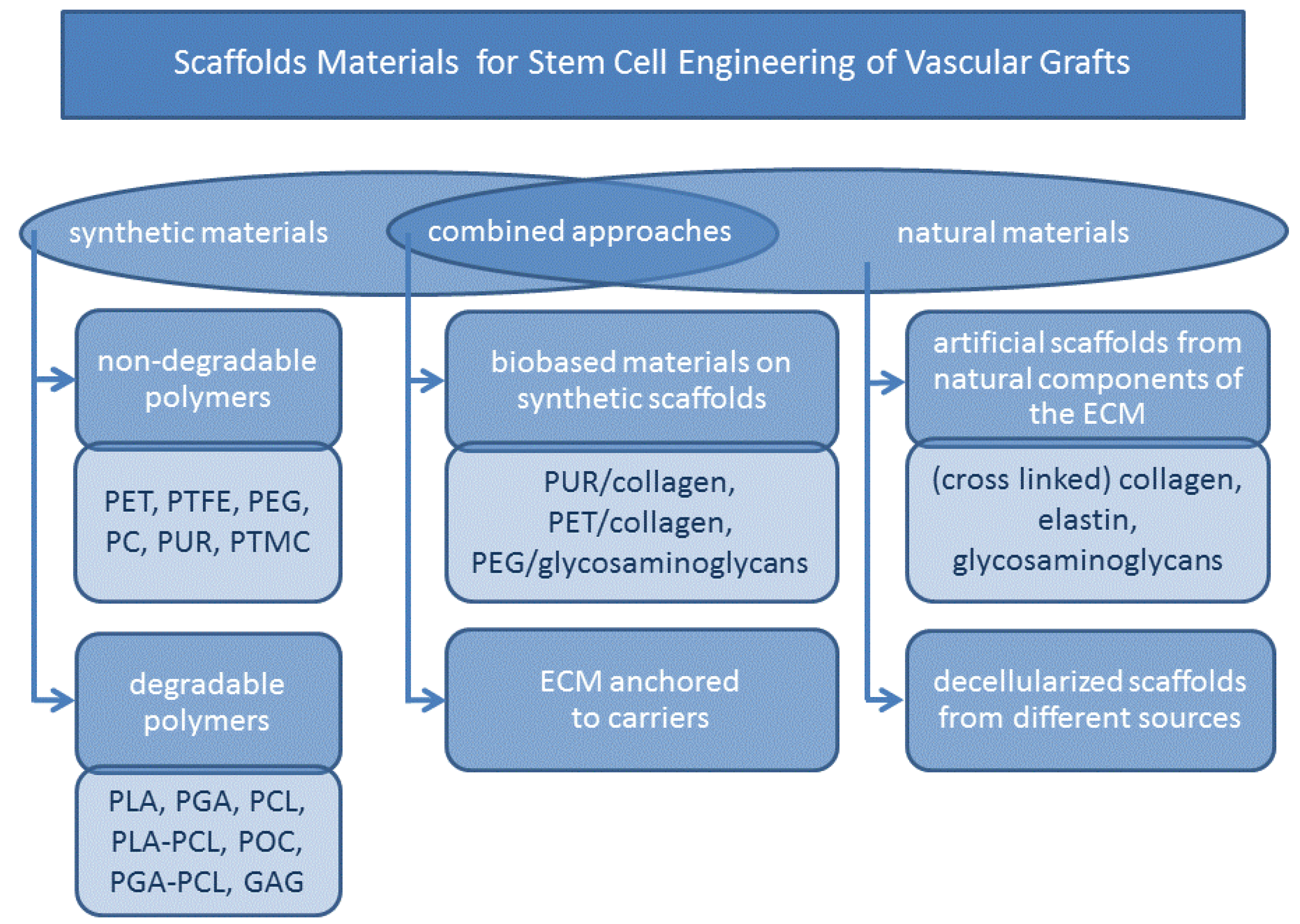
5.1.1. Synthetic Polymers
5.1.2. Natural Polymers
5.2. Scaffold Fabrication Methods
5.2.1. Conventional and Nano-based Fabrication Methods
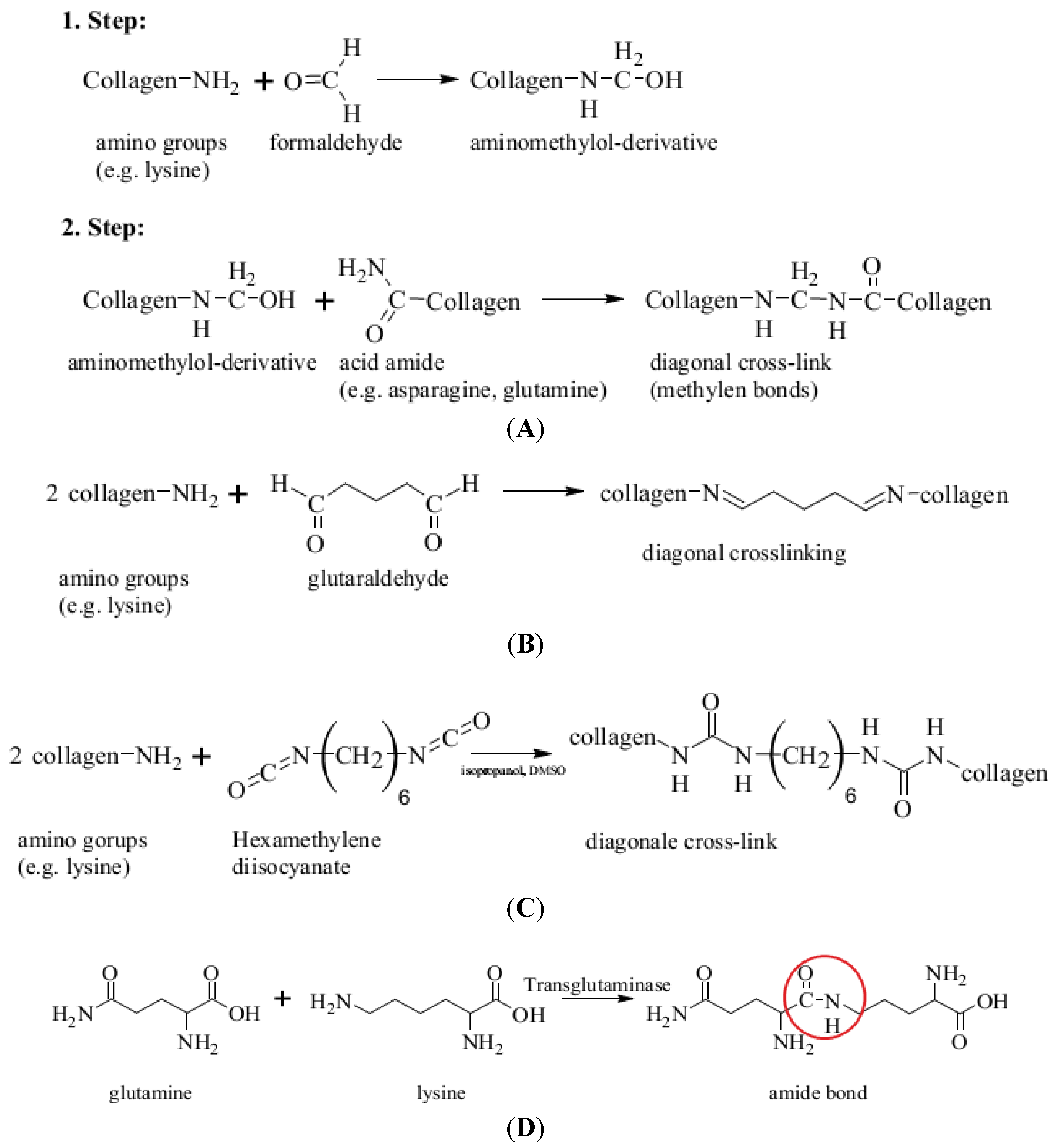
5.2.2. Network Formation via Polymer Cross-Linking
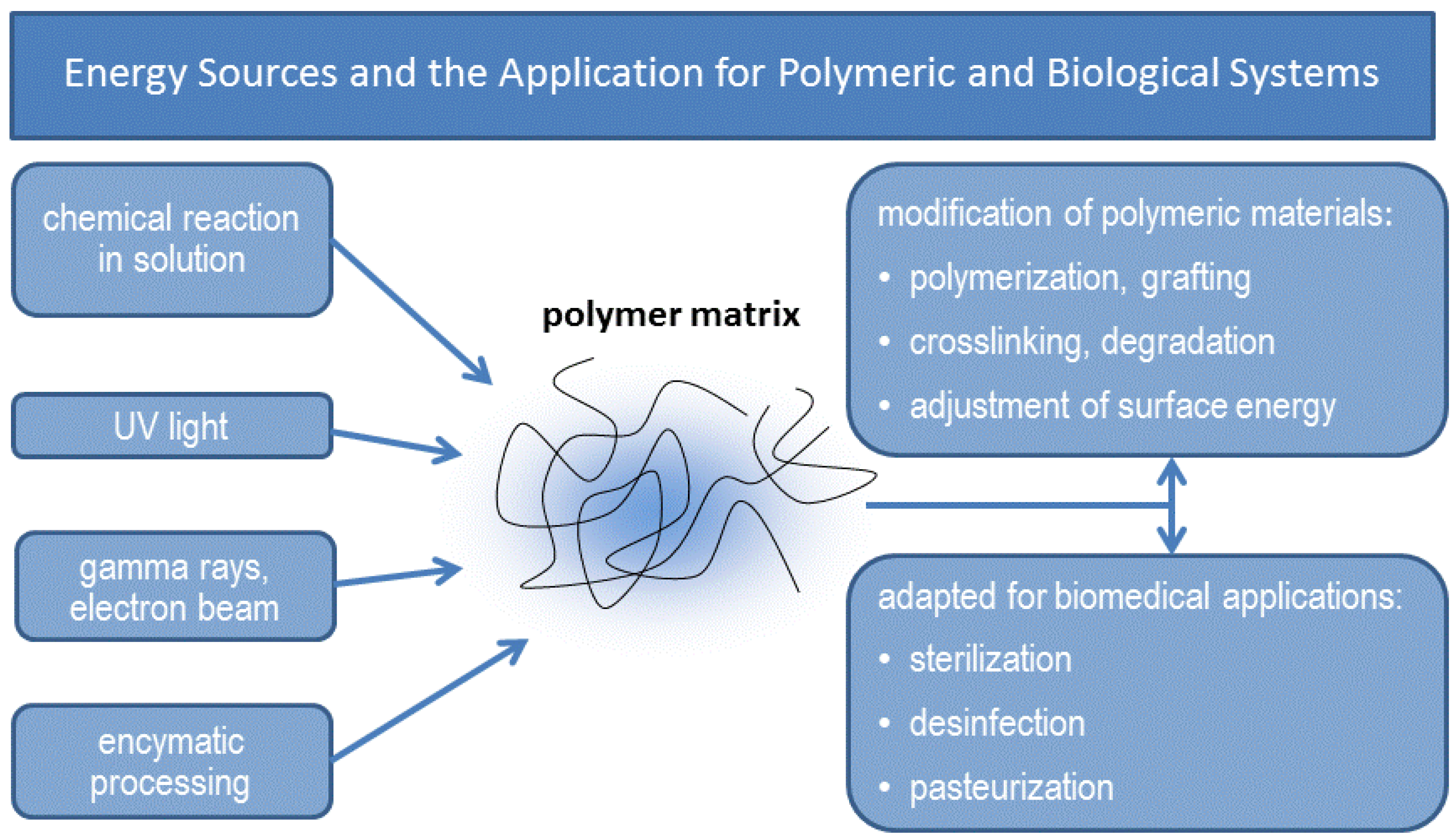

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Irradiation target | Net effect | Dose range (kGy) |
|---|---|---|
| Food | Cold pasteurization | 0.3–30 |
| Medical disposable items | Sterilization | 10–60 |
| Cellulose and derivates | Degradation | 5–50 |
| Polymer coatings | Curing | 30–160 |
| Polyolefin foams | Crosslinking | 40–80 |
| Heat-shrinkable materials | Memory imparted | 75–250 |
| Soft materials (especially rubber) | Cross-linking/Vulcanization | 80–400 |
| Fluoropolymers | Degradation | 500–1500 |
| Gemstones | Coloration | >10,000 |
6. Summary of Scaffolds for Vascular Grafts
7. Conclusions
Abbreviations
| AP | alkaline phosphatase |
| ATSC | adipose-tissue derived stem cell |
| 3D | three-dimensional |
| Ac-LDL | acetylated-low density protein |
| Ang-1 | angiopoietin-1 |
| ASC | adult stem cell |
| α-SMA | alpha-smooth muscle actin |
| bFGF | basic fibroblast growth factor |
| BM-EPC | bone marrow-derived progenitor cell |
| BM-MSC | bone marrow-derived mesenchymal stem cell |
| BrdU | bromedesoxyuridine |
| CD | cluster of determinants |
| DMSO | dimethylsulfoxid |
| EC | endothelial cell |
| EGM-2 | endothelial growth medium-2 |
| eNOS | endothelial nitric monoxide synthase |
| EPC | endothelial progenitor cell |
| ePET | expanded poly(ethylene terephtalate) |
| ESC | embryonic stem cell |
| FACS | fluorescent activated cell sorting |
| FDM | fused deposition modeling |
| GAG | glycosaminoglycan |
| HA | hyaluronic acid |
| hATSC | human adipose tissue-derived cell |
| HC | heavy chain |
| HCAEC | human coronary artery endothelial cell |
| hCASMC | human coronary artery smooth muscle cell |
| hEC | human endothelial cell |
| hEPC | human endothelial progenitor cell |
| hESC | human embryonic stem cell |
| HGF | hepatocyte growth factor |
| HGF | hematopoietic growth factor |
| HLA-DR | human leukocyte antigen |
| hMSC | human mesenchymal stem cell |
| hSMC | human smooth muscle cell |
| hsp | heat shock protein |
| HSVSMC | human saphenous vein smooth muscle cell |
| HUASMC | human umbilical artery smooth muscle cell |
| HUVEC | human umbilical vein endothelial cell |
| HUVSMC | human umbilical vein smooth muscle cell |
| IGF-1 | insulin-like growth factor-1 |
| iPS | induced pluripotent stem cell |
| KDR/Flk-1 | kinase insert domian receptor/fetal liver kinase-1 |
| MI | moycardium infarction |
| MSC | mesenchymal stem cell |
| NHS | N-hydroxysuccinimide |
| PB-EPC | peripheral blood-derived endothelial progenitor cell |
| PB-MSC | peripheral blood derived mesenchymal stem cell |
| PCL | poly(caprolactone) |
| PDGF | platelet-derived growth factor |
| PECAM-1 | platelet endothelial cell adhesion molecule-1 |
| PEG | poly(ethylene glycol) |
| PET | poly(ethylene terephtalate) |
| PGA | polyglycolic acid |
| PHB | poly(hydroxybutyrate) |
| PLA | polylactic acid |
| PLA-PGA | poly(lactic glycolic acid) |
| PLA-PCL | poly(lactic acid)caprolactone |
| POC | poly(1,8-octanediol-co-citric acid) |
| PTHrP | parathyroid hormone-related protein |
| PTFE | poly(tetrafluoroethylene) |
| PTMC | poly(1,3-trimethylene carbonate) |
| PU | polyurethanes |
| NG2 | nerval/glial antigen 2 |
| SDF | stromal cell-derived factor |
| SL | stereolithography |
| SLA | selective laser ablation |
| SLS | selective laser sintering |
| SM | smooth muscle |
| SMC | smooth muscle cell |
| SMPC | smooth muscle progenitor cell |
| TEBV | tissue-engineered blood vessel |
| TEGV | tissue-engineered vascular graft |
| TGF-β | transforming growth factor-beta |
| TGM | transglutaminase |
| UB-EPC | umbilical cord blood endothelial progenitor cell |
| UC-EPC | umbilical cord endothelial progenitor cell |
| UV | ultraviolet |
| VCAM | vascular cell adhesion molecule |
| VE-cadherin | vascular endothelial cadherin |
| VEGF | vascular endothelial growth factor |
Acknowledgments
Conflicts of Interest
References
- Limbach, C.A.; Lange, M.; Schulze, M.; Tobiasch, E. Recent patents on biomedical applications for the treatment of atherosclerosis. Recent Pat. Regen. Med. 2012, 2, 75–102. [Google Scholar]
- Donovan, P.J.; Gearhart, J. The end of the beginning for pluripotent stem cells. Nature 2001, 414, 92–97. [Google Scholar] [CrossRef]
- Liu, S. iPS cells: A more critical review. Stem Cells Dev. 2008, 17, 391–397. [Google Scholar] [CrossRef]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef]
- Takahashi, K.; Okita, K.; Nakagawa, M.; Yamanaka, S. Induction of pluripotent stem cells from fibroblast cultures. Nat. Protoc. 2007, 2, 3081–3089. [Google Scholar] [CrossRef]
- Schulze, M.; Tobiasch, E. Artificial Scaffolds and Mesenchymal Stem Cells for Hard Tissues. In Tissue Engineering III: Cell—Surface Interactions for Tissue Culture; Scheper, T., Kaspar, C., Pörtner, R., Witte, F., Eds.; Springerverlag: Berlin, Germany, 2011; Volume 126, pp. 153–194. [Google Scholar]
- Jin, H.J.; Bae, Y.K.; Kim, M.; Kwon, S.J.; Jeon, H.B.; Choi, S.J.; Kim, S.W.; Yang, Y.S.; Oh, W.; Chang, J.W. Comparative analysis of human mesenchymal stem cells from bone marrow adipose tissue, and umbilical cord blood as source of cell therapy. Int. J. Mol. Sci. 2013, 14, 17986–18001. [Google Scholar] [CrossRef]
- Calloni, R.; Cordero, E.A.; Henriques, J.A.; Bonatto, D. Reviewing and updating the major molecular markers for stem cells. Stem Cells Dev. 2013, 22, 1455–1476. [Google Scholar] [CrossRef]
- Shamblott, M.J.; Axelman, J.; Littlefield, J.W.; Blumenthal, P.D.; Huggins, G.R.; Cui, Y. Human embryonic germ cell derivatives express a broad range of developmentally distinct markers and proliferate extensively in vitro. Proc. Natl. Acad. Sci. USA 2001, 98, 113–118. [Google Scholar]
- Huang, H.; Nakayama, Y.; Qin, K.; Yamamoto, K.; Ando, J.; Yamashita, J.; Itoh, H.; Kanda, K.; Yaku, H.; Okamoto, Y.; et al. Differentiation from embryonic stem cells to vascular wall cells under in vitro pulsatile flow loading. J. Artif. Organs 2005, 8, 110–118. [Google Scholar] [CrossRef]
- Nakagami, H.; Nakagawa, N.; Takeya, Y.; Kashiwagi, K.; Ishida, C.; Hayashi, S.; Aoki, M.; Matsumoto, K.; Nakamura, T.; Ogihara, T.; et al. Model of vasculogenesis from embryonic stem cells for vascular research and regenerative medicine. Hypertension 2006, 48, 112–119. [Google Scholar] [CrossRef]
- Ferreira, L.S.; Gerecht, S.; Shieh, H.F.; Watson, N.; Rupnick, M.A.; Dallabrida, S.M.; Vunjak-Novakovic, G.; Langer, R. Vascular progenitor cells isolated from human embryonic stem cells give rise to endothelial and smooth muscle like cells and form vascular networks in vivo. Circ. Res. 2007, 101, 286–294. [Google Scholar] [CrossRef]
- Abilez, O.; Benharash, P.; Mehrotra, M.; Miyamoto, E.; Gale, A.; Picquet, J.; Xu, C.; Zarins, C. A novel culture system shows that stem cells can be grown in 3D and under physiologic pulsatile conditions for tissue engineering of vascular grafts. J. Surg. Res. 2006, 132, 170–178. [Google Scholar] [CrossRef]
- Xiong, Q.; Hill, K.L.; Li, Q.; Suntharalingam, P.; Mansoor, A.; Wang, X.; Jameel, M.N.; Zhang, P.; Swingen, C.; Kaufman, D.S.; et al. A fibrin patch-based enhanced delivery of human embryonic stem cell-derived vascular cell transplantation in a porcine model of postinfarction left ventricular remodeling. Stem Cells 2011, 29, 367–375. [Google Scholar] [CrossRef]
- Shen, G.; Tsung, H.C.; Wu, C.F.; Liu, X.Y.; Wang, X.Y.; Liu, W.; Cui, L.; Cao, Y.L. Tissue engineering of blood vessels with endothelial cells differentiated from mouse embryonic stem cells. Cell Res. 2003, 13, 335–341. [Google Scholar] [CrossRef]
- McDevitt, T.C.; Ramachandran, N.R. A Cellularized Biomaterial from Embryonic Stem Cells. EU Patent 1965812, 8 November 2006. [Google Scholar]
- Yeryemyeyev, A.; Artyem, V.; Svyetlakov, A.V.; Bol’shakov, I.N.; Shyeina, J.I.; Polstjanoj, A.M. Method for Preparation of a Cardiomyocyte Matrix. EU Patent 2559755, 1 April 2011. [Google Scholar]
- Hayashi, S. Blood Vessel-Specific Organogenesis from Embryonic Stem Cells on Three-Dimensional Matrigel Layer. U.S. Patent 20070003526, 24 March 2004. [Google Scholar]
- Wernig, M.; Meissner, A.; Foreman, R.; Brambrink, T.; Ku, M.; Hochedlinger, K. In vitro reprogramming of fibroblasts into a pluripotent ES-cell-like state. Nature 2007, 448, 318–324. [Google Scholar] [CrossRef]
- Yu, J.; Vodyanik, M.; Smuga-Otto, K.; Antosiewicz-Bourget, J.; Frane, J.L.; Tian, S. Induced pluripotent stem cell lines derived from human somatic cells. Science 2007, 318, 1917–1920. [Google Scholar] [CrossRef]
- Stadtfeld, M.; Nagaya, M.; Utikal, J.; Weir, G.; Hochedlinger, K. Induced pluripotent stem cells generated without viral integration. Science 2008, 322, 945–949. [Google Scholar] [CrossRef]
- Kim, J.B.; Zaehres, H.; Wu, G.; Gentile, L.; Ko, K.; Sebastiano, V. Pluripotent stem cells induced from adult neural stem cells by reprogramming with two factors. Nature 2008, 454, 646–650. [Google Scholar] [CrossRef]
- Kim, J.B.; Zaehres, H.; Schöler, R. Generation of Induced Pluripotent Stem (IPs) Cells. WO Patent 2009144008, 26 May 2009. [Google Scholar]
- Yamanaka, S.; Tanabe, K. Method for Producing Induced Pluripotent Stem Cells. WO Patent 2010050626, 30 October 2009. [Google Scholar]
- Choi, K.D.; Yu, J.; Smuga-Otto, K.; Salvagiotto, G.; Rehrauer, W.; Vodyanik, M.; Thomson, J. Hematopoietic and endothelial differentiation of human induced pluripotent stem cells. Stem Cells 2009, 27, 559–567. [Google Scholar]
- Lian, Q.; Zhang, Y.; Zhang, J.; Zhang, H.K.; Wu, X.; Zhang, Y. Functional mesenchymal stem cells derived from human induced pluripotent stem cells attenuate limb ischemia in mice. Circulation 2010, 121, 1113–1123. [Google Scholar] [CrossRef]
- Hibino, N.; Duncan, D.R.; Nalbandian, A.; Yi, T.; Qyang, Y.; Shinoka, T.; Breuer, C.K. Evaluation of the use of an induced pluripotent stem cell sheet for the construction of tissue-engineered vascular grafts. J. Thorac. Cardiovasc. Surg. 2012, 143, 696–739. [Google Scholar] [CrossRef]
- Tobiasch, E. Adult Human Mesenchymal Stem Cells as Source for Future Tissue Engineering. In Forschungsspitzen und Spitzenforschung; Zacharias, C., Ter Horst, K.W., Witt, K.U., Eds.; Springverlag: Berlin, Germany, 2008; pp. 329–338. [Google Scholar]
- Haddouti, E.M.; Skroch, M.; Zippel, N.; Müller, C.; Birova, B.; Pansky, A.; Kleinfeld, C.; Winter, M.; Tobiasch, E. Human dental follicle precursor cells of wisdom teeth: Isolation and differentiation towards osteoblasts for implants with and without scaffolds. Mater. Sci. Eng. Technol. 2009, 40, 732–737. [Google Scholar]
- Tobiasch, E. Differentiation Potential of Adult Human Mesenchymal Stem Cells. In Stem Cell Engineering; Artmann, G.M., Hescheler, J., Minger, S., Eds.; Springerverlag: Berlin, Germany, 2010; pp. 61–77. [Google Scholar]
- Zippel, N.; Schulze, M.; Tobiasch, E. Biomaterials and mesenchymal stem cells for regenerative medicine. Recent Pat. Biotechnol. 2010, 4, 1–22. [Google Scholar] [CrossRef]
- Khan, D.; Kleinfeld, C.; Winter, M.; Tobiasch, E. Oral Tissues as Source for Bone Regeneration in Dental Implantology. In Tissue Regeneration—From Basic Biology to Clinical Application; Davies, J., Ed.; InTech: Rijeka, Croatia, 2012; pp. 325–335. [Google Scholar]
- Zhang, Y.; Khan, D.; Delling, J.; Tobiasch, E. Mechanisms underlying the osteo- and adipo-differentiation of human mesenchymal stem cells. Sci. World J. 2012, 2012. [Google Scholar] [CrossRef]
- Hass, R.; Kasper, C.; Böhm, S.; Jacobs, R. Different populations and sources of human mesenchymal stem cells (MSC): A comparison of adult and neonatal tissue-derived MSC. Cell Commun. Signal. 2011, 9. [Google Scholar] [CrossRef]
- Dominici, M.; Blanc, K.L.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Zhang, Y.; Tobiasch, E. The Role of Purinergic Receptors in Stem Cells in Their Derived Consecutive Tissues. In Adult Stem Cell Standardization; Di Nardo, P., Ed.; River Publishers: Roma, Italy, 2011; pp. 73–98. [Google Scholar]
- Zippel, N.; Limbach, C.A.; Ratajski, N.; Urban, C.; Luparello, C.; Pansky, A. Purinergic receptors influence the differentiation of human mesenchymal stem cells. Stem Cells Dev. 2012, 21, 884–900. [Google Scholar] [CrossRef]
- Longo, A.; Librizzi, M.; Naselli, F.; Caradonna, F.; Tobiasch, E.; Luparello, C. PTHrP in differentiating human mesenchymal stem cells: Transcript isoform expression, promoter methylation, and protein accumulation. Biochemie 2013, 95, 1888–1896. [Google Scholar] [CrossRef]
- Zuk, P.A.; Zhu, M.; Ashjian, P.; de Ugarte, D.A.; Huang, J.I.; Mizuno, H. Human adipose tissue is a source of multipotent stem cells. Mol. Biol. Cell 2002, 13, 4279–4295. [Google Scholar] [CrossRef]
- Pansky, A.; Roitzheim, B.; Tobiasch, E. Differentiation potential of adult of human mesenchymal stem cells. Clin. Lab. 2007, 53, 81–84. [Google Scholar]
- Birnbaum, T.; Hildebrandt, J.; Nuebling, G.; Sostak, P.; Straube, A. Glioblastoma-dependent differentiation and angiogenic potential of human mesenchymal stem cells in vitro. J. Neurooncol. 2011, 105, 57–65. [Google Scholar] [CrossRef]
- Kim, J.H.; Park, I.S.; Park, Y.; Jung, Y.; Kim, S.H.; Kim, S.H. Therapeutic angiogenesis of three-dimensionally cultured adipose-derived stem cells in rat infarcted hearts. Cytotherapy 2013, 15, 542–556. [Google Scholar] [CrossRef]
- Duffy, G.P.; McFadden, T.M.; Byrne, E.M.; Gill, S.L.; Farrell, E.; O’Brien, F.J. Towards in vitro vascularization of collagen-GAG scaffolds. Eur. Cells Mater. 2011, 21, 15–30. [Google Scholar]
- Song, Y.; Kamphuis, M.M.; Zhang, Z.; Sterk, L.M.; Vermes, I.; Poot, A.A.; Feijen, J.; Grijpma, D.W. Flexible and elastic porous poly(trimethylene carbonate) structures for use in vascular tissue engineering. Acta Biomater. 2010, 6, 1269–1277. [Google Scholar] [CrossRef]
- Tsigkou, O.; Pomerantseva, I.; Spencer, J.A.; Redondo, P.A.; Hart, A.R.; O’Doherty, E.; Lin, Y.; Friedrich, C.C.; Daheron, L.; Lin, C.P.; et al. Engineered vascularized bone grafts. Proc. Natl. Acad. Sci. USA 2010, 107, 3311–3316. [Google Scholar] [CrossRef]
- Gong, Z.; Niklason, L.E. Small-diameter human vessel wall engineered from bone marrow-derived mesenchymal stem cells (hMSCs). FASEB J. 2008, 22, 1635–1648. [Google Scholar] [CrossRef]
- Kim, B.S.; Choi, J.S.; Kim, J.D.; Choi, Y.C.; Cho, Y.W. Recellularization of decellularized human adipose tissue-derived extracellular matrix sheets with other human cell types. Cell Tissue Res. 2012, 348, 559–567. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, S.; Zhou, J.; Wang, J.; Zhen, M.; Liu, Y.; Chen, J.; Qi, Z. The development of a tissue-engineered artery using decellularized scaffold and autologous ovine mesenchymal stem cells. Biomaterials 2010, 31, 296–307. [Google Scholar] [CrossRef]
- Dufourcq, P.; Descamps, B.; Tojais, N.F.; Leroux, L.; Oses, P.; Daret, D.; Moreau, C.; Lamazière, J.M.; Couffinhal, T.; Duplàa, C. Secreted frizzled-related protein-1 enhances mesenchymal stem cell function in angiogenesis and contributes to neovessel maturation. Stem Cells 2008, 26, 2991–3001. [Google Scholar] [CrossRef]
- Altman, A.M.; Yan, Y.; Matthias, N.; Bai, X.; Rios, C.; Mathur, A.B.; Song, Y.H.; Alt, E.U. IFATS collection: Human adipose-derived stem cells seeded on a silk fibroin-chitosan scaffold enhance wound repair in a murine soft tissue injury model. Stem Cells 2009, 27, 250–258. [Google Scholar] [CrossRef]
- Chung, E.; Nam, S.Y.; Ricles, L.M.; Emelianov, S.Y.; Suggs, L.J. Evaluation of gold nanotracers to track adipose-derived stem cells in a PEGylated fibrin gel for dermal tissue engineering applications. Int. J. Nanomed. 2013, 8, 325–336. [Google Scholar]
- Matsumura, G.; Miyagawa-Tomita, S.; Shin’oka, T.; Ikada, Y.; Kurosawa, H. First evidence that bone marrow cells contribute to the construction of tissue-engineered vascular autografts in vivo. Circulation 2003, 108, 1729–1734. [Google Scholar] [CrossRef]
- Cho, S.W.; Jeon, O.; Lim, J.E.; Gwak, S.J.; Kim, S.S.; Choi, C.Y.; Kim, D.I.; Kim, B.S. Preliminary experience with tissue engineering of a venous vascular patch by using bone marrow-derived cells and a hybrid biodegradable polymer scaffold. J. Vasc. Surg. 2006, 44, 1329–1340. [Google Scholar] [CrossRef]
- Lim, S.H.; Cho, S.W.; Park, J.C.; Jeon, O.; Lim, J.M.; Kim, S.S.; Kim, B.S. Tissue-engineered blood vessels with endothelial nitric oxide synthase activity. J. Biomed. Mater. Res. 2008, 85, 537–546. [Google Scholar]
- Mendelson, K.; Aikawa, E.; Mettler, B.A.; Sales, V.; Martin, D.; Mayer, J.E.; Schoen, F.J. Healing and remodeling of bioengineered pulmonary artery patches implanted in sheep. Cardiovasc. Pathol. 2007, 16, 277–282. [Google Scholar] [CrossRef]
- Derval, N.; Barandon, L.; Dufourcq, P.; Leroux, L.; Lamazière, J.M.; Daret, D.; Couffinhal, T.; Duplàa, C. Epicardial deposition of endothelial progenitor and mesenchymal stem cells in a coated muscle patch after myocardial infarction in a murine model. Eur. J. Cardiothorac. Surg. 2008, 34, 248–254. [Google Scholar] [CrossRef]
- Ashara, T.; Murohara, T.; Sullivan, A.; Silver, M.; van der Zee, R.; Li, T. Isolation of putative progenitor endo cells for angiogenesis. Science 1997, 275, 964–967. [Google Scholar] [CrossRef]
- Shi, Q.; Rafii, S.; Wu, M.H.; Wijelath, E.S.; Yu, C.; Ishida, A. Evidence for circulating bone marrow-derived endothelial cells. Blood 1998, 92, 362–367. [Google Scholar]
- Reyes, M.; Dudek, A.; Jahagirdar, B.; Koodie, L.; Marker, P.H.; Verfaillie, C.M. Origin of endothelial progenitors in human postnatal bone marrow. J. Clin. Investig. 2002, 109, 337–346. [Google Scholar]
- Phuc, P.V.; Nqoc, V.B.; Lam, D.H.; Viet, P.Q.; Nqoc, P.K. Isolation of three important types of stem cells from the same samples of banked umbilical cord blood. Cell Tissue Bank. 2012, 13, 341–351. [Google Scholar] [CrossRef]
- Shirota, T.; He, H.; Yasui, H.; Matsuda, T. Human endothelial progenitor cell-seeded hybrid graft: Proliferative and antithrombogenic potentials in vitro and fabrication processing. Tissue Eng. 2003, 9, 127–136. [Google Scholar] [CrossRef]
- Westman, J.; Nilsson, M.; Ornitz, D.M.; Svahn, C. Synthesis and fibroblast growth factor binding of oligosaccharides related to heparin and heparan sulphate. Carbohydr. Res. 1995, 14, 95–113. [Google Scholar]
- Amini, A.R.; Laurencin, C.T.; Nukavarapu, S.P. Differential analysis of peripheral blood- and bone marrow-derived endothelial progenitor cells for enhanced vascularization in bone tissue engineering. J. Orthop. Res. 2012, 30, 1507–1515. [Google Scholar] [CrossRef]
- Davis, M.E.; Motion, J.P.; Narmoneva, D.A.; Takahashi, T.; Hakuno, D.; Kamm, R.D.; Zhang, S.; Lee, R.T. Injectable self-assembling peptide nanofibers create intramyocardial microenvironments for endothelial cells. Circulation 2005, 111, 442–450. [Google Scholar] [CrossRef]
- Sainz, J.; Al Haj Zen, A.; Caligiuri, G.; Demerens, C.; Urbain, D.; Lemitre, M.; Lafont, A. Isolation of “side population” progenitor cells from healthy arteries of adult mice. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 281–286. [Google Scholar]
- Sreerekha, P.R.; Divya, P.; Krishnan, L.K. Adult stem cell homing and differentiation in vitro on composite fibrin matrix. Cell Prolif. 2006, 39, 301–312. [Google Scholar] [CrossRef]
- Wu, X.; Rabkin-Aikawa, E.; Guleserian, K.J.; Perry, T.E.; Masuda, Y.; Sutherland, F.W.; Schoen, F.J.; Mayer, J.E.; Bischoff, J. Tissue-engineered microvessels on three-dimensional biodegradable scaffolds using human endothelial progenitor cells. Am. J. Physiol. Heart Circ. Physiol. 2004, 287, 480–487. [Google Scholar] [CrossRef]
- Allen, J.; Khan, S.; Serrano, M.C.; Ameer, G. Characterization of porcine circulating progenitor cells: Toward a functional endothelium. Tissue Eng. Part A 2008, 14, 183–194. [Google Scholar] [CrossRef]
- Cherqui, S.; Kingdon, K.M.; Thorpe, C.; Kurian, S.M.; Salomon, D.R. Lentiviral gene delivery of vMIP-II to transplanted endothelial cells and endothelial progenitors is proangiogenic in vivo. Mol. Ther. 2007, 15, 1264–1272. [Google Scholar] [CrossRef]
- Schmeckpeper, J.; Ikeda, Y.; Kumar, A.H.; Metharom, P.; Russell, S.J.; Caplice, N.M. Lentiviral tracking of vascular differentiation in bone marrow progenitor cells. Differentiation 2009, 78, 169–176. [Google Scholar] [CrossRef]
- Fioretta, E.S.; Fledderus, J.O.; Baaijens, F.P.; Bouten, C.V. Influence of substrate stiffness on circulating progenitor cell fate. J. Biomech. 2012, 45, 736–744. [Google Scholar] [CrossRef]
- Clever, Y.P.; Cremers, B.; Krauss, B.; Böhm, M.; Speck, U.; Laufs, U.; Scheller, B. Paclitaxel and sirolimus differentially affect growth and motility of endothelial progenitor cells and coronary artery smooth muscle cells. EuroIntervention 2011, 7, 32–42. [Google Scholar] [CrossRef]
- Cottone, R.J.; Yoklavich, M.; Parker, S. Progenitor Endothelial Cell Capturing with a Drug Eluting Implantable Medical Device. U.S. Patent 0120172970, 29 December 2011. [Google Scholar]
- Melero-Martin, J.M.; Khan, Z.A.; Picard, A.; Wu, X.; Paruchuri, S.; Bischoff, J. In vivo vasculogenic potential of human blood-derived endothelial progenitor cells. Blood 2007, 109, 4761–4768. [Google Scholar] [CrossRef]
- Griese, D.P.; Ehsan, A.; Melo, L.G.; Kong, D.; Zhang, L.; Mann, M.J.; Pratt, R.E.; Mulligan, R.C.; Dzau, V.J. Isolation and transplantation of autologous circulating endothelial cells into denuded vessels and prosthetic grafts: Implications for cell-based vascular therapy. Circulation 2003, 108, 2710–2715. [Google Scholar] [CrossRef]
- He, H.; Shirota, T.; Yasui, H.; Matsuda, T. Canine endothelial progenitor cell-lined hybrid vascular graft with nonthrombogenic potential. J. Thorac. Cardiovasc. Surg. 2003, 126, 455–464. [Google Scholar] [CrossRef]
- Yu, J.; Wang, A.; Tang, Z.; Henry, J.; Li-Ping, L.B.; Zhu, Y.; Yuan, F.; Huang, F.; Li, S. The effect of stromal cell-derived factor-1α/heparin coating of biodegradable vascular grafts on the recruitment of both endothelial and smooth muscle progenitor cells for accelerated regeneration. Biomaterials 2012, 33, 8062–8074. [Google Scholar] [CrossRef]
- Noishiki, Y.; Seki, F.; Yasuhiro, A.; Akihiro, S.Y. Method for Harvesting Bone Marrow and Its Medical Apparatus. U.S. Patent 20090187116, 5 April 2005. [Google Scholar]
- Muschler, G.F. Apparatus and Method for Harvesting Bone Marrow. U.S. Patent 20070055282, 9 November 2006. [Google Scholar]
- Sciorra, L. Multipotent Stem Cell-Based Culture Systems and Models. WO Patent 2013049628, 28 September 2012. [Google Scholar]
- Harman, R.J.; Sand, T.T. Methods of Preparing and Using Stem Cell Compositions and Kits Comprising the Same. EU Patent 1678295, 7 October 2004. [Google Scholar]
- Kieda, C.; Grillon, C.; Lamerant-Fayel, N.; Paprocka, M.; Krawczenko, A.; Goszyk-Dus, D. Human and Murine Stem-Cell Lines: Models of Endothelial Cell Precursors. EU Patent 2524031, 11 January 2011. [Google Scholar]
- Ma, T.; Junho, K. Mesenchymal Stem Cells (MSC) Expansion Methods and Materials. WO Patent 2012079086, 11 December 2011. [Google Scholar]
- Xianqun, F.; Yefei, W.; Huifang, Z. Method for in-Vitro Fusion of Stem Cells and Porous Biomaterial. CN Patent 102477419, 26 November 2010. [Google Scholar]
- Gostjeva, E.V.; Thilly, W.G. Wound Healing Metakaryotic Stem Cells and Methods of Use Thereof. WO Patent 2012061073, 24 October 2011. [Google Scholar]
- Amoh, Y.; Li, L.; Yang, M.; Jiang, P. Angiogensis Models Using Nestin-Expressing Stem Cells to Image Nascent Blood Vessels. U.S. Patent 20050170330, 28 October 2004. [Google Scholar]
- Lee, J.W.; Park, M.S.K.; Yun, H. Method for Inducing in Vivo Migration of Stem Cells. WO Patent 2012121451, 21 February 2011. [Google Scholar]
- Hantash, B. Differentiation of Mesenchymal Stem Cells into Fibroblasts, Compositions Comprising Mesenchymal Stem Cell-Derived Fibroblasts, and Methods of Using the Same. WO Patent 2009155334, 17 June 2009. [Google Scholar]
- Guoqi, T.; Yigang, W.; Baisong, Z. Technique for Heart Disease External Differentiation Therapy by Utilizing Stem Cells Ofmasticatory Muscles and Orbicularis Oculi Muscles. CN Patent 102703382, 25 June 2012. [Google Scholar]
- Nanxue, S.L.; Lidong, G.; Xuetao, P.; Cixian, B.; Fan, Y.; Yunfang, W.; Huimin, Y. Structural Method and Application of Tissue Engineering Adipose Tissue. CN Patent 1912109, 9 August 2005. [Google Scholar]
- Shortkroff, S.; Khoury, J.; Tarrant, L.J.B.; Claesson, H.P.I.; Smith, R.L. A Method for Improvement of Differentiation of Mesenchymal Stem Cells Using a Double-Structured Tissue Implant. EU Patent 182887, 20 August 2008. [Google Scholar]
- Park, K.; Deok, K.; Hee, J.H.; Dong, K.; Hong, Y.; Jin, C.; Heung, J.; Jang, J.W. Method for Surface Modification of Polymeric Scaffold for Stem Cell Transplantation Using Decellularized Extracellular Matrix. U.S. Patent 20100267143, 28 October 2009. [Google Scholar]
- Aicher, W.; Angres, B. Isolierung von Mesenchymalen Stammzellen. DE Patent 102010023837, 7 June 2010. [Google Scholar]
- Hamada, Y.; Matsumura, N.; Egusa, H.; Kaneda, Y.; Okazaki, M. Mesenchymal Cell Proliferation Stimulator and Skeletal System Biomaterial. WO Patent 2008026634, 29 August 2007. [Google Scholar]
- Cho, M. Scaffold for Articular Cartilage Regeneration and Method for Manufacturing Same. U.S. Patent 20130084636, 22 February 2011. [Google Scholar]
- Komeno, K.; Ono, S.; Tanne, K. Material for Graft and Method for Culturing Anaplastic Mesenchymal Stem Cell. JP Patent 2006122147, 26 October 2004. [Google Scholar]
- Huang, L.H. Cell Tissue Gel Containing Collagen and Hyaluronan. EU Patent 2515867, 21 December 2010. [Google Scholar]
- Abatangelo, G.; Callegaro, L. A Biologic Material Comprising an Efficient Culture of Bone Marrow Stem Cells Partially or Completely Differentiated into Connective Tissue Cells and a Three-Dimensional Biocompatible and Biodegradable Matrix Consisting of a Hyaluronic Acid Derivative. EU Patent 0863776, 19 November 1996. [Google Scholar]
- Choi, Y.S.; Noh, S.; Eun, L.; Youngjun, J.; Sang, Y.; Kim, S.M.; Han, K.; Chung, H.M. Cartilage Cell Treating Agent Comprising Collagen, Hyaluronic Acid Derivative, and Stem Cell Derived from Mammal Umbilical Cord. WO Patent 2013009102, 11 July 2012. [Google Scholar]
- Min, B.; Hyoun, P.; So, R.; Park, S.H. Method for Differenciating Mesenchymal Stem Cell and Culturing Chondrocytes Using Alginate Coated Fibrin/Ha Composite Scaffold. KR Patent 20090013425, 1 August 2007. [Google Scholar]
- Shimizu, T.; Haraguchi, Y.; Aoki, S.; Okano, M.; Ono, T.; Rin, K.; Horii, A. Differentiation Method to Smooth Muscle Cell of Mesenchymal Stem Cell, Production Method of Smooth Muscle Implant, and Smooth Muscle Implant. JP Patent 2012235730, 11 May 2011. [Google Scholar]
- Westenfelder, C. Therapeutic Method for Kidney Disease and Multi Organ Failure due to Mesenchymal Stem Cell, and Mesenchymal Stem Cell-Conditioned Medium. JP Patent 2012246298, 26 July 2012. [Google Scholar]
- Arai, K. Method for Producing Collagen, Cell and DNA. JP Patent 2007089575, 28 August 2006. [Google Scholar]
- Brouard, S.; Otterbein, L.E.; Anrather, J.; Tobiasch, E.; Bach, F.H.; Choi, A.M.; Soares, M.P. Carbon monoxide generated by hemeoxygenase 1 suppresses endothelial cell apoptosis. J. Exp. Med. 2000, 192, 1015–1026. [Google Scholar] [CrossRef]
- Brouard, S.; Berberat, P.O.; Tobiasch, E.; Seldon, M.P.; Bach, F.H.; Soares, M.P. Hemeoxygenase-1-derived carbon monoxide requires the activation of transcription factor NF-kappa B to protect endothelial cells from tumor necrosis factor-alpha-mediated apoptosis. J. Biol. Chem. 2002, 277, 17950–17961. [Google Scholar]
- Soares, M.P.; Usheva, A.; Brouard, S.; Berberat, P.O.; Gunther, L.; Tobiasch, E.; Bach, F.H. Modulation of endothelial cell apoptosis by heme oxygenase-1-derived carbon monoxide. Antioxid. Redox Signal. 2002, 4, 321–329. [Google Scholar]
- Jaffe, E.A.; Nachman, R.L.; Becker, C.G.; Minich, C.R. Culture of human endothelial cells derived from umbilical veins. Identification by morphologic and immunologic criteria. J. Clin. Investig. 1973, 52, 2745–2756. [Google Scholar] [CrossRef]
- Voyta, J.C.; Via, D.P.; Butterfield, C.E.; Zetter, B.R. Identification and isolation of endothelial cells based on their increased uptake of acetylated-low density protein. J. Cell Biol. 1984, 99, 2034–2040. [Google Scholar] [CrossRef]
- Knighton, D.R. Method for Isolating Wound Capillary Endothelial Cells. WO Patent 1990012085, 11 April 1990. [Google Scholar]
- Aldons, J.L. Atherosclerosis. Nature 2000, 407, 233–241. [Google Scholar] [CrossRef]
- Ray, J.L.; Leach, R.; Herbert, J.M.; Bentson, M. Isolation of vascular smooth muscle cells from a single murine aorta. Methods Cell Sci. 2001, 23, 185–188. [Google Scholar] [CrossRef]
- Ribeiro, M.P.; Relvas, R.; Chiquita, I.J. Isoalation of human umbilical artery smooth muscle cells (HUASMC). J. Vis. Exp. 2010, 41. [Google Scholar] [CrossRef]
- Fillinger, M.F.; O’Connor, S.E.; Wagner, R.J.; Cronenwett, J.L. The effect of endothelial cell coculture on smooth muscle cell proliferation. J. Vasc. Surg. 1993, 17, 1058–1067. [Google Scholar]
- Wallace, C.S.; Truskey, G.A. Direct-contact co-culture between smooth muscle cells and endothelial cells inhibits TNF-alpha-mediated endothelial cell activation. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, 338–346. [Google Scholar] [CrossRef]
- Chen, L.; Xi, T.; Yang, Z.; Wang, W.; Liu, Q.; Wu, Y.; Gu, Y.; Wang, J.; Feng, Z. Effect of surface property of different polyether-ester copolymers on growth of smooth muscle cells and endothelial cells. Zhongguo Xiu Fu Chong Jian Wai Ke Za Zhi 2005, 19, 481–485. [Google Scholar]
- Baguneid, M.; de Mel, A.; Yildirimer, L.; Fuller, B.J.; Hamilton, G.; Seifalian, A.M. In vivo study of a model tissue-engineered small-diameter vascular bypass graft. Biotechnol. Appl. Biochem. 2011, 58, 14–24. [Google Scholar] [CrossRef]
- Boni, L.; Chalajour, F.; Sasaki, T.; Snyder, R.L.; Boyd, W.D.; Riemer, R.K.; Reddy, V.M. Reconstruction of pulmonary artery with porcine small intestinal submucosa in a lamb surgical model: Viability and growth potential. J. Thorac. Cardiovasc. Surg. 2012, 144, 963–969. [Google Scholar]
- Callegari, A.; Bollini, S.; Iop, L.; Chiavegato, A.; Torregrossa, G.; Pozzobon, M.; Gerosa, G.; de Coppi, P.; Elvassore, N.; Sartore, S. Neovascularization induced by porous collagen scaffold implanted on intact and cryoinjured rat hearts. Biomaterials 2007, 28, 5449–5461. [Google Scholar] [CrossRef]
- Saif, J.; Schwarz, T.M.; Chau, D.Y.; Henstock, J.; Sami, P.; Leicht, S.F.; Hermann, P.C.; Alcala, S.; Mulero, F.; Shakesheff, K.M.; et al. Combination of injectable multiple growth factor-releasing scaffolds and cell therapy as an advanced modality to enhance tissue neovascularization. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 1897–1904. [Google Scholar] [CrossRef]
- Stoeckius, M.; Erat, A.; Fujikawa, T.; Hiromura, M.; Koulova, A.; Otterbein, L.; Bianchi, C.; Tobiasch, E.; Dagon, Y.; Sellke, F.W.; et al. Essential roles of Raf/ERK/MAPK pathway, YY1, and Ca+2 influx in the growth arrest of human vascular smooth muscle cells by bilirubin. J. Biol. Chem. 2012, 287, 15418–15426. [Google Scholar] [CrossRef]
- Nabel, E.G.; Nabel, G.J. Inhibition of Arterial Smooth Muscle Cell Proliferation. EU Patent 0752002, 1 March 1995. [Google Scholar]
- Suda, T.; Katagiri, T.; Kodaira, K.; Goto, M.; Higashio, K. Therapeutic Agent for Arteriosclerosis. WO Patent 2007007665, 7 July 2006. [Google Scholar]
- Wolinsky, H. Method for the Prevention of Restenosis. U.S. Patent 4824436, 7 April 1986. [Google Scholar]
- Altman, P. Method of Treating Coronary Arteries with Perivascular Delivery of Therapeutic Agents. U.S. Patent 20110293592, 3 August 2011. [Google Scholar]
- Edelman, E.; Nathan, A.; Nugent, M.A. Inhibition of Vascular Occlusion Following Vascular Intervention. EU Patent 0850073, 9 May 1996. [Google Scholar]
- Uchida, S. Vascular Prothesis. WO Patent 2012115275, 23 February 2012. [Google Scholar]
- Takano, T. Artificial Blood Vessel. JP Patent 04303451, 30 March 1991. [Google Scholar]
- Cao, Y.L. Method and Apparatus for Constructing Blood Vessel in Vitro in the Tissue Project. CN Patent 1236056, 9 September 2002. [Google Scholar]
- Burleigh, D. Therapeutic Treatment for Inhibiting Blood Vessel Blockage Using a Polypeptide. EP Patent 722334, 14 September 1994. [Google Scholar]
- Jianhong, M.; Chunli, H.; Chuhong, Z.; Dajun, Y.; Li, L. Biological Artificial Blood Vessel Capable of in Vivo Capturing Endothelial Ancestral Cell. CN Patent 101066477, 17 May 2007. [Google Scholar]
- Iwazawa, R.; Nakamura, K. Scaffold for Vascular Endothelial Cell Migration. EU Patent 2578246, 29 August 2012. [Google Scholar]
- Hoganson, D.M.; Vacanti, J.P. System and Method for in Vitro Blood Vessel Modeling. WO Patent 2009102571, 11 February 2009. [Google Scholar]
- Kellar, R.S.; Landeen, L.K.; Shepherd, B.R.; Naughton, G.K.; Ratcliffe, A.; Williams, S.K. Scaffold-based three-dimensional human fibroblast culture provides a structural matrix that supports angiogenesis in infarcted heart tissue. Circulation 2001, 104, 2063–2068. [Google Scholar] [CrossRef]
- Tosun, Z.; McFetridge, P.S. Improved recellularization of ex vivo vascular scaffolds using direct transport gradients to modulate ECM remodeling. FASEB J. 2005, 19, 1546–1548. [Google Scholar]
- Formigli, L.; Perna, A.M.; Meacci, E.; Cinci, L.; Margheri, M.; Nistri, S.; Tani, A.; Silvertown, J.; Orlandini, G.; Porciani, C.; et al. Paracrine effects of transplanted myoblasts and relaxin on post-infarction heart remodelling. J. Cell. Mol. Med. 2007, 11, 1087–1100. [Google Scholar] [CrossRef]
- Pařízek, M.; Novotná, K.; Bačáková, L. The role of smooth muscle cells in vessel wall pathophysiology and re-construction using bioactive synthetic polymers. Physiol. Res. 2011, 60, 419–437. [Google Scholar]
- Chlupač, J.; Filova, E.; Bacakova, L. Blood vessel replacement: 50 Years of development and tissue engineering paradigms in vascular surgery. Physiol. Res. 2009, 58, 119–139. [Google Scholar]
- Engler, A.; Sen, S.; Sweeney, H.; Discher, D. Matrix elasticity directs stem cell lineage specification. Cell 2006, 126, 677–698. [Google Scholar] [CrossRef]
- Gilbert, P.; Havenstritte, K.; Magnusson, K.; Sacco, A.; Leonardi, N.; Kraft, P.; Nguyen, N.; Thrun, S.; Lutolf, M.; Blau, H. Substrate elasticity regulates skeletal muscle stem cell self-renewal in culture. Science 2010, 329, 1078–1081. [Google Scholar] [CrossRef]
- Holst, J.; Watson, S.; Lord, M.S.; Eamegdool, S.S.; Bax, D.V.; Nivison-Smith, L.B.; Kondyurin, A.; Ma, L.; Oberhauser, A.F.; Weiss, A.S.; et al. Substrate elasticity provides mechanical signals for the expansion of hemopoietic stem and progenitor cells. Nat. Biotechnol. 2010, 28, 1123–1128. [Google Scholar] [CrossRef]
- Benoit, D.; Schwartz, M.; Durney, A.; Anseth, K. Small functional groups for controlled differentiation of hydrogel-encapsulated human mesenchymal stem cells. Nat. Mater. 2008, 7, 816–823. [Google Scholar] [CrossRef]
- Schmedlen, R.; Elbjeirami, W.; Gobin, A.; West, J. Tissue engineered small-diameter vascular grafts. Clin. Plast. Surg. 2003, 30, 507–517. [Google Scholar] [CrossRef]
- Sawhney, A.; Pathak, C.; Hubbel, J. Bioerodible hydrogels based on photopolymerized poly(ethylene glycol)-co-poly(α-hydroxy acid)diacrylate macromers. Macromelecules 1993, 26, 581–587. [Google Scholar] [CrossRef]
- Szycher, M. Szycher’s Handbook of Polyurethane, 2nd ed.; CRC Press LLC: Boca Raton, FL, USA, 2013. [Google Scholar]
- Wright, J. Using Polyurethanes in Medical Applications. Available online: http://www.mddionline.com/article/using-polyurethanes-medical-applications (accessed on 24 June 2013).
- Baer, G.; Wilson, T.; Matthews, D.; Maitland, D. Shape-memory behavior of thermally stimulated polyurethane for medical applications. J. Appl. Polym. Sci. 2007, 103, 3882–3892. [Google Scholar] [CrossRef]
- McBane, J.E.M.; Sharifpoor, S.; Labow, R.S.; Ruel, M.; Suuronen, E.J.; Santerre, J.P. Tissue engineering a small diameter vessel substitute: Engineering constructs with select biomaterials and cells. Curr. Vasc. Pharmacol. 2012, 10, 347–360. [Google Scholar] [CrossRef]
- Romaškevič, T.; Budrienė, S.; Pielichowski, K.; Pielichowski, J. Application of polyurethane-based materials for immobilization of enzymes and cells: A review. Chemija 2006, 17, 74–89. [Google Scholar]
- Prewitz, M.; Seib, F.; Pompe, T.; Werner, C. Polymeric biomaterials for stem cell bioengineering. Macromol. Rapid Commun. 2012, 33, 1420–1431. [Google Scholar] [CrossRef]
- Szilagyi, D.; France, L.; Smith, R. Clinical use of an elastic Dacron prosthesis. Arch. Surg. 1958, 77, 538–551. [Google Scholar]
- Van Damme, H.; Deprez, M.; Creemers, E.; Limet, R. Intrinsic structural failure of polyester (dacron) vascular grafts. A general review. Acta Chir. Belg. 2005, 105, 249–255. [Google Scholar]
- Takamoto, T.; Ichinohe, N.; Tabata, Y. Proliferation of rat mesenchymal stem cells in collagen sponges reinforced with poly(ethylene terephthalate) fibers by stirring culture method. J. Biomater. Sci. Polym. 2012, 23, 1741–1753. [Google Scholar]
- Prokoph, S.; Chavakis, E.; Levental, K.; Zieris, A.; Freudenberg, U.; Dimmeler, S.; Werner, C. Sustained delivery of SDF-1α from heparin-based hydrogels to attract circulating pro-angiogenic cells. Biomaterials 2012, 33, 4792–4800. [Google Scholar] [CrossRef]
- Kurane, A.; Vyavahare, N. In vivo vascular tissue engineering: Influence of cytokine and implant location on tissue specific cellular recruitment. J. Tissue Eng. Regen. Med. 2009, 3, 280–289. [Google Scholar] [CrossRef]
- Huang, N.F.; Li, S. Regulation of the matrix microenvironment for stem cell engineering and regenerative medicine. Ann. Biomed. Eng. 2011, 39, 1201–1214. [Google Scholar] [CrossRef]
- Prewitz, M.K.; Seib, P.F.; von Bonin, M.; Friedrichs, J.; Stibel, A.; Niehage, C.; Müller, K.; Anastassiadis, K.; Waskow, C.; Hoflack, B.; et al. Tightly anchored tissue-mimetic matrices as instructive stem cell microenvironments. Nat. Methods 2013, 10. [Google Scholar] [CrossRef]
- Embuscado, M.E.; Huber, K.C. Edible Films and Coatings for Food Applications; Springer: Dordrecht, The Netherlands, 2009; pp. 245–250. [Google Scholar]
- Le Ricousse-Roussanne, S.; Barateau, V.; Contreres, J.O.; Boval, B.; Kraus-Berthier, L.; Tobelem, G. Ex vivo differentiated endothelial and smooth muscle cells from human cord blood progenitors home to the angiogenic tumor vasculature. Cardiovasc. Res. 2004, 62, 176–184. [Google Scholar] [CrossRef]
- Bohrer, C.; Ruth, P.; Schatton, W. Verfahren zur Herstellung von Porösen Schwämmen aus Gereinigtem Marinen Kollagen. DE Patent 102005063438 A1, 24 February 2005. [Google Scholar]
- Donzelli, E.; Salvade, A.; Mimo, P.; Vigano, M.; Morrone, M.; Papagna, R.; Carini, F.; Zaopo, A.; Miloso, M.; Baldoni, M.; et al. Mesenchymal stem cells cultured on a collagen scaffold: In vitro osteogenic differentiation. Arch. Oral Biol. 2007, 52, 64–73. [Google Scholar] [CrossRef]
- Ruszczak, Z. Effect of collagen matrices on dermal wound healing. Adv. Drug Deliv. Rev. 2003, 55, 1595–1611. [Google Scholar] [CrossRef]
- Chvapil, M. Collagen sponge: Theory and practice of medical applications. J. Biomed. Mater. Res. 1977, 11, 721–741. [Google Scholar] [CrossRef]
- Casu, B.; Lindahl, U. Structure and biological interactions of heparin and heparan sulfate. Adv. Carbohydr. Chem. Biochem. 2001, 57, 159–206. [Google Scholar]
- Avci, F.Y.; de Angelis, P.L.; Liu, J.; Linhardt, R.J. Enzymatic Synthesis of Glycosaminoglycanes: Improving on Nature, Frontiers in Modern Carbohydrate Chemistry. In ACS Symposium Series 960; Demchenko, A.V., Ed.; ACS: Washington, DC, USA, 2007; pp. 253–284. [Google Scholar]
- Linhardt, R.J.; Claude, S. Hudson award address in carbohydrate chemistry. Heparin: Structure and activity. J. Med. Chem. 2003, 46, 2551–2554. [Google Scholar] [CrossRef]
- Jaques, L.B. Heparin: An old drug with a new paradigm. Science 1979, 206, 528–533. [Google Scholar]
- Linhardt, R.J. Heparin: An important drug enters its seventh decade. Chem. Ind. 1991, 2, 45–50. [Google Scholar]
- Dougher, A.M.; Wasserstrom, H.; Torley, L.; Shridaran, L.; Westdock, P.; Hileman, R.E.; Fromm, J.R.; Anderberg, R.; Lyman, S.; Linhardt, R.J.; et al. Identification of a heparin binding peptide on the extracellular domain of the KDR VEGF receptor. Growth Factors 1997, 14, 257–268. [Google Scholar] [CrossRef]
- Lucas, H.; Basten, J.E.; van Dinther, T.G.; Meuleman, D.G.; van Aelst, S.F.; van Boeckel, C.A. Syntheses of heparin-like pentamers containing opened uronic acid moieties. Tetrahedron 1990, 46, 8207–8228. [Google Scholar] [CrossRef]
- Van Boeckel, C.A.; Petitou, M. The unique antithrombin III binding domain of heparin: A lead to new synthetic antithrombotics. Angew. Chem. Int. Ed. 1993, 32, 1671–1690. [Google Scholar] [CrossRef]
- Karst, N.A.; Linhardt, R.J. Recent chemical and enzymatic approaches to the synthesis of glycosaminoglycan oligosaccharides. Curr. Med. Chem. 2003, 10, 1993–2031. [Google Scholar] [CrossRef]
- Bacakova, L.; Svorcik, V. Cell Colonization Control by Physical and Chemical Modification of Materials. In Cell Growth Processes: New Research, 1st ed.; Kimura, D., Ed.; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2008; pp. 5–56. [Google Scholar]
- Sachlos, E.; Czernuszka, J. Making tissue engineering scaffolds work. Eur. Cells Mater. 2003, 30, 29–39. [Google Scholar]
- Moroni, L.; Wijn, J.; van Blitterswijkc, A. Integrating novel technologies to fabricate smart scaffolds. J. Biomater. Sci. Polym. Ed. 2008, 19, 543–572. [Google Scholar] [CrossRef]
- Anderson, S.B.; Lin, C.C.; Kuntzler, D.V.; Anseth, K.S. The performance of human mesenchymal stem cells encapsulated in cell-degradable polymer-peptide hydrogels. Biomaterials 2011, 32, 3564–3574. [Google Scholar] [CrossRef]
- Wan, A.C.A.; Ying, J.Y. Nanomaterials for in situ cell delivery and tissue regeneration. Adv. Drug Deliv. Rev. 2010, 62, 731–740. [Google Scholar] [CrossRef]
- Ma, P.X. Biomimetic materials for tissue engineering. Adv. Drug Deliv. Rev. 2008, 60, 184–198. [Google Scholar] [CrossRef]
- Goldstein, A.; Zhu, G.; Morris, G. Effect of osteoblastic culture conditions on the structure of poly(d,l-lactic-co-glycolic acid) foam scaffolds. Tissue Eng. 1999, 5, 421–434. [Google Scholar] [CrossRef]
- Nasibulin, A.G.; Anisimov, A.S.; Pikhitsa, P.V.; Jiang, H.; Brown, D.P.; Choi, M.; Kauppinen, E.I. Investigations of nanobud formation. Chem. Phys. Lett. 2007, 446, 109–114. [Google Scholar] [CrossRef]
- Bauer, S.; Park, J.; von der Mark, K.; Schmuki, P. Improved attachment of mesenchymal stem cells on super hydrophobic TiO2 nanotubes. Acta Biomater. 2008, 4, 1576–1582. [Google Scholar] [CrossRef]
- Park, J.; Bauer, S.; Schmuki, P. Narrow window in nanoscale dependent activation of endothelial cell growth and differentiation on TiO2 nanotube surfaces. Nano Lett. 2009, 9, 3157–3164. [Google Scholar] [CrossRef]
- Zhang, L.; Webster, T. Nanotechnology and nanomaterials: Promises for improved tissue regeneration. Nano Today 2009, 4, 66–80. [Google Scholar] [CrossRef]
- Kim, K.; Dean, D.; Lu, A.Q. Early osteogenic signal expression of rat bone marrow stromal cells is influenced by both hydroxyapatite nanoparticles content and initial cell seeding density in biodegradable nanocomposites scaffolds. Acta Biomater. 2011, 7, 1249–1264. [Google Scholar] [CrossRef]
- Oliveira, J.M.; Sousa, R.A.; Malafaya, P.B. In vivo study of dendron-like nanoparticles for stem cells “tune-up”: From nano to tissues. Nanomedicine 2011, 7, 914–924. [Google Scholar]
- Wang, J.; Yu, X. Preparation, characterization and in vitro analysis of novel structured nanofibrous scaffolds for bone tissue engineering. Acta Biomater. 2010, 6, 3004–3012. [Google Scholar] [CrossRef]
- Kretlow, J.; Mikos, A. From material to tissue: Biomaterial development, scaffold fabrication, and tissue engineering. AIChE J. 2008, 54, 3048–3067. [Google Scholar] [CrossRef]
- Wei, G.; Ma, P.X. Nanostructured biomaterials for regeneration, nano-scaled drug release systems incorporated into nanostructured biomaterials represents a novel and promising strategy to tissue regeneration. Adv. Funct. Mater. 2008, 18, 3568–3582. [Google Scholar] [CrossRef]
- Burdick, J.A.; Vunjak-Novakovic, G. Engineered microenvironments for controlled stem cell differentiation. Tissue Eng. 2009, 15, 205–219. [Google Scholar]
- Peltola, S.; Sanna, M.; Melchels, F.; Grijpma, D.; Kellomaki, M. A review of rapid prototyping techniques for tissue engineering purposes. Ann. Med. 2008, 40, 268–280. [Google Scholar] [CrossRef]
- Bens, A.; Seitz, H.; Bermes, H.; Emons, M.; Pansky, A.; Roitzheim, B. Non-toxic flexible photopolymers for medical stereolithography technology. Rapid Prototyp. J. 2007, 13, 38–47. [Google Scholar] [CrossRef]
- Sitharaman, B.; Avti, P.K.; Schaefer, K. A novel nanoparticle-enhanced photoacustic stimulus for bone tissue engineering. Tissue Eng. Part A 2011, 17, 1851–1858. [Google Scholar] [CrossRef]
- Lim, Y.C.; Johnson, J.; Fei, Z.Z. Micropatterning and characterization of electrospun poly(epsilon-caprolactone)/gelatin nanofiber tissue scaffolds by femtosecond laser ablation for tissue engineering applications. Biotechnol. Bioeng. 2011, 108, 116–126. [Google Scholar] [CrossRef]
- Fan, D.M.; Akkaraju, G.R.; Couch, E.F. The role of nanostructured mesoporous silicon in discriminating in vitro calcification for electrospun composite tissue engineering scaffolds. Nanoscale 2011, 3, 354–361. [Google Scholar] [CrossRef]
- Qiang, L.; Qing, L.F. Preparation of 3-D regenerated fibroin scaffolds with freeze drying method and freeze drying/foaming technique. J. Mater. Sci. Mater. Med. 2006, 17, 1349–1356. [Google Scholar] [CrossRef]
- Gagnieu, C. Vernetzbare Kollagenderivate, Verfahren zu Ihrer Herstellung und Ihre Verwendung zur Herstellung von Biomaterialien. DE Patent 69315483T2, 2 July 1998. [Google Scholar]
- Zeugolis, D.I.; Paul, G.R.; Attenburrow, G. Cross-linking of extruded collagen fibers: A biomimetic three-dimensional scaffold for tissue engineering applications. J. Biomed. Mater. Res. Part A 2009, 89, 895–908. [Google Scholar] [CrossRef]
- Fratzl, P. Collagen—Structure and Mechanics, 1st ed.; Springer: New York, NY, USA, 2008; pp. 1–13. [Google Scholar]
- Kling, S. Biomechanical Response of Normal and Cross-Linked Corneas; Diplomarbeit, Fachhochschule und Berufskollegs NTA: Isny (Allgäu), Germany, 2009. [Google Scholar]
- Nimni, M.E.; Wise, L.; Trantolo, D.J.; Altobelli, D.E.; Yaszemski, M.J. Collagen: Molecular Structure and Biomaterial Properties. In Encyclopedic Handbook of Biomaterials and Bioengineering: Part A: Materials, 1st ed.; Taylor & Francis: New York, NY, USA, 1995; Volume 1, pp. 1229–1243. [Google Scholar]
- Khor, E. Methods for the treatment of collagenous tissues for bioprostheses. Biomaterials 1997, 18, 95–105. [Google Scholar] [CrossRef]
- Pischinger, A. Das System der Grundregulation—Grundlagen Einer Ganzheitsbiologischen Medizin, 11th ed.; Karl, F., Ed.; Haug Verlag: Stuttgart, Germany, 2010; pp. 67–73. [Google Scholar]
- Reich, G. Kollagen: Eine Einführung in Methoden, Ergebnisse und Probleme der Kollagenforschung, 1st ed.; Theodor Steinkopff: Dresden, Germany, 1966; pp. 103–106. [Google Scholar]
- Hamaguchi, P.Y.; Shiku, Y.; Tanaka, M. Property improvement of fish water soluble protein films by dialdehyde starch (DAS) and/or sodium dodecyl sulfate (SDS) treatments. J. Package Technol. Sci. 2003, 12, 271–283. [Google Scholar]
- Audic, J.-L.; Chaufer, B. Caseinate based biodegradable films with improved water resistance. J. Appl. Polym. Sci. 2010, 117, 1828–1836. [Google Scholar]
- Angele, P.; Abke, J.; Kujat, R.; Faltermeier, H.; Schumann, D.; Nerlich, M.; Kinner, B.; Englert, C.; Ruszczak, Z.; Mehrl, R.; et al. Influence of different collagen species on physico-chemical properties of cross-linked collagen matrices. Biomaterials 2004, 25, 2831–2841. [Google Scholar] [CrossRef]
- Sommer, I.; Kunz, P.M. Improving the water resistance of biodegradable collagen films. J. Appl. Polym. Sci. 2012, 125, 27–41. [Google Scholar] [CrossRef]
- Renner, M.U. Gelatinefilme als Arzneistoffträger, 1st ed.; Cuvillier Verlag: Göttingen, Germany, 2004; pp. 26–28. [Google Scholar]
- Gratzer, P.F.; Pereira, C.A.; Lee, J.M. Solvent environment modulates effects of glutaraldehyde crosslinking on tissue-derived biomaterials. J. Biomed. Mater. Res. 1996, 31, 533–543. [Google Scholar] [CrossRef]
- Jayakrishnan, A.; Jameela, S.R. Glutaraldehyde as a fixative in bioprostheses and drug delivery matrices. Biomaterials 1996, 17, 471–484. [Google Scholar] [CrossRef]
- Burness, D.M.; Pouradier, J. The Hardening of Gelatin and Emulsions. In The Theory of Photographic Process, 5th ed.; James, T.H., Ed.; Macmillian & Co.: New York, NY, USA, 1977; pp. 51–67. [Google Scholar]
- Petite, H.; Rault, I.; Huc, A.; Menasche, P.; Herbage, D. Use of the acyl azide method for cross-linking collagen-rich tissues such as pericardium. J. Biomed. Mater. Res. 1990, 24, 179–187. [Google Scholar] [CrossRef]
- Marquié, C. Chemical reactions in cottonseed protein cross-linking by formaldehyde, glutaraldehyde, and glyoxal for the formation of protein films with enhanced mechanical properties. J. Agric. Food Chem. 2001, 49, 4676–4681. [Google Scholar] [CrossRef]
- Cheung, D.T.; Nimni, M.E. Mechanism of crosslinking of proteins by glutaraldehyde II. Reaction with monomeric and polymeric collagen. Connect. Tissue Res. 1982, 10, 201–216. [Google Scholar] [CrossRef]
- Damink, L.H.; Dijkstra, P.J.; van Luyn, M.J.A.; van Wachem, P.B.; Nieuwenhuis, P.; Feijen, J. Crosslinking of dermal sheep collagen using a water-soluble carbodiimide. Biomaterials 1996, 17, 765–773. [Google Scholar] [CrossRef]
- Rault, I.; Frei, V.; Herbage, D. Evaluation of different chemical methods for cross-linking collagen gel, films and sponges. J. Mater. Sci. Mater. Med. 1996, 7, 215–221. [Google Scholar] [CrossRef]
- Bedino, J.H. Champion—An Expanding Encyclopedia of Mortuary Practices, 1st ed.; Champion Company: Springfield, IL, USA, 2003; p. 649. [Google Scholar]
- Maser, F. Wursthüllen auf Basis von Kollagen. 1. Freiberger Kollagensymposium; Bundesforschungsanstalt für Ernährung und Lebensmittel: Freiberg, Germany, 1996. [Google Scholar]
- Marzec, E.; Pietrucha, K. The effect of different methods of cross-linking of collagen on its dielectric properties. Biophys. Chem. 2008, 132, 89–96. [Google Scholar] [CrossRef]
- Stachel, I.; Schwarzenbolz, U.; Henle, T.; Meyer, M. Cross-linking of type I collagen with microbial transglutaminase: Identification of cross-linking sites. Biomacromol 2010, 11, 698–705. [Google Scholar] [CrossRef]
- Chambi, H.; Grosso, C. Edible films produced with gelatin and casein crosslinked with transglutaminase. Food Res. Int. 2006, 39, 458–466. [Google Scholar] [CrossRef]
- Taylor, M.M.; Marmer, W.N.; Brown, E.M. Characterization of biopolymers prepared from gelatin and sodium caseinate for potential use in leather processing. J. Am. Leather Chem. Assoc. 2005, 100, 149–159. [Google Scholar]
- Marx, C. Optimierung Einer Mikrobiellen Transglutaminase Mittels Random Mutagenese. Dissertation; Martin-Luther-Universität Halle-Wittenberg: Halle, Germany, 2008. [Google Scholar]
- Kaminski, A.; Grazka, E.; Jastrzebska, A.; Marowska, J.; Gut, G.; Wojciechowski, A. Effect of accelerated electron beam on mechanical properties of human cortical bone: Influence of different processing methods. Cell Tissue Bank. 2012, 13, 375–386. [Google Scholar] [CrossRef]
- Weadock, K.; Olson, R.M.; Silver, F.H. Evaluation of collagen crosslinking techniques. Biomater. Med. Devices Artif. Organs 1983, 11, 293–318. [Google Scholar]
- Kamińska, A.; Sionkowska, A. Effect of UV radiation on the infrared spectra of collagen. Polym. Degrad. Stab. 1996, 51, 19–26. [Google Scholar] [CrossRef]
- Raiskup, F.; Spoerl, E. Corneal crosslinking with riboflavin and ultraviolet AI principles. Ocul. Surf. 2013, 11, 65–74. [Google Scholar] [CrossRef]
- Hayes, S.; Kamma-Lorger, C.S.; Boote, C.; Young, R.D.; Quantock, A.J. The effect of riboflavin/UVA collagen cross-linking therapy on the structure and hydrodynamic behaviour of the ungulate and rabbit corneal stroma. PLoS One 2013, 8, e52860. [Google Scholar]
- Cheung, D.T.; Perelman, N.; Tong, D.; Nimni, M.E. The effect of γ-irradiation on collagen molecules, isolated α-chains, and crosslinked native fibers. J. Biomed. Mater. Res. 2004, 24, 581–589. [Google Scholar]
- Drobny, J.G. Radiation Technology for Polymers, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2010; pp. 5–12. [Google Scholar]
- Cheng, S. Radiation Processing of Polymer Materials and Its Industrial Applications, 1st ed.; John Wiley & Sons: New York, NY, USA, 2011. [Google Scholar]
- Jiang, B.; Wu, Z.; Zhao, H.; Tang, F.; Lu, J.; Wie, Q.; Zhang, X. Electron beam irradiation modification of collagen membranes. Biomaterials 2006, 27, 15–23. [Google Scholar] [CrossRef]
- Seto, A.; Gatt, C.J.; Dunn, M.G. Radioprotection of tendon tissue via crosslinking and free radical scavenging. Clin. Orthop. Relat. Res. 2008, 466, 1788–1795. [Google Scholar] [CrossRef]
- Zuwei, M.; Zhengwei, M.; Gao, C. Surface modification and property analysis of biomedical polymers used for tissue engineering. Colloid Surf. B 2007, 60, 137–157. [Google Scholar] [CrossRef]
- Charlesby, A. Use of high energy radiation for crosslinking and degradation. Radiat. Phys. Chem. 1977, 9, 17–29. [Google Scholar]
- Menges, G. Werkstoffkunde Kunststoffe, 4th ed.; Carl Hanser: München, Germany, 1998; pp. 287–290. [Google Scholar]
- Flory, P.J.; Rehner, J. Statistical mechanics of cross-linked polymer networks II. Swelling. J. Chem. Phys. 1943, 11, 512–520. [Google Scholar] [CrossRef]
- Gottlieb, R.; Kaiser, C.; Gohs, U.; Arndt, K.F. Temperature sensitive hydrogels based on hydroxypropyl cellulose by high energy irradiation. Macromol. Symp. 2007, 254, 361–369. [Google Scholar]
- Lappan, U.; Geibler, U.; Gohs, U.; Uhlmann, S. Influence of irradiation temperature on grafting of styrene into poly(tetrafluoroethylene-co-hexafluoropropylene) films. Macromol. Mater. Eng. 2009, 294, 510–515. [Google Scholar] [CrossRef]
- Abke, J. Verbesserung der Biokompatibilität Metallischer Implantate Durch Kovalente Anbindung Einer Quervernetzten Kollagenschicht. Dissertation, University Regensburg, Regensburg, Germany, 28 May 2003. [Google Scholar]
- Dahl, B.J.; Spotts, E.; Truong, J.Q. Corneal collagen crosslinking: An introduction and literature review. Optometry 2012, 83, 33–42. [Google Scholar] [CrossRef]
- Suljovrujic, E.; Ignjatovic, N.; Uskokovic, D. Gamma irradiation processing of hydroxylapatite/poly-l-lactide composite biomaterial. Radiat. Phys. Chem. 2003, 67, 357–379. [Google Scholar]
- Heger, A.; Dorschner, H.; Dunsch, L.; Ihme, B.; Lunkwitz, K. Technologie der Strahlenchemie von Polymeren, 1st ed.; Carl Hanser: München, Germany, 1990. [Google Scholar]
- Hirose, M.; Yamato, M.; Kwon, O.H.; Harimoto, M.; Kushida, A.; Shimizu, T.; Kikushi, A.; Okano, T. Temperature-responsive surface for novel co-culture systems of hepatocytes with endothelial celle: 2-D pattern and double layered co-cultures. Yonsei Med. J. 2000, 41, 803–813. [Google Scholar]
- Röthemeyer, F.; Sommer, F. Kautschuktechnologie, 2nd ed.; Carl Hanser: München, Germany, 2006; p. 327. [Google Scholar]
- Becker, G.W.; Braun, D. Kunststoffhandbuch, 2nd ed.; Carl Hanser: München, Germany, 1990; Volume 1, p. 956. [Google Scholar]
- Khorasani, M.T.; Mirzadeh, H.; Irani, S. Plasma surface modification of poly(l-lactic acid) and poly(lactic-co-glycolic acid) films for improvement of nerve cells adhesion. Radiat. Phys. Chem. 2007, 77, 280–287. [Google Scholar] [CrossRef]
- Leonard, D.; Pick, L.T.; Farrar, D.F.; Dickson, G.R.; Orr, J.F.; Buchanan, F.J. The modification of PLA and PLGA using electron-beam radiation. J. Biomed. Mater. Res. 2009, 89, 567–574. [Google Scholar]
- Loo, S.C.J.; Ooi, C.P.; Boey, Y.C.F. Influence of electron-beam radiation on the hydrolytic degradation behavior of poly(lactide-co-glycolide) (PGLA). Biomaterials 2006, 26, 3809–3817. [Google Scholar]
- Miao, P.; Wu, D.; Zhao, C.; Xu, G.; Zeng, K.; Wang, Y.; Fu, Q.; Yang, G. Modification of poly(d,l-lactic acid)-co-poly(ethylene glycol) copolymer by low energy electron beam (EB) radiation. e-Polymers 2010, 6, 1–12. [Google Scholar]
- Quynh, T.M.; Mitomo, H.; Nagasawa, N.; Wada, Y.; Yoshii, F.; Tamada, M. Properties of crosslinked polylactides (PLLA & PDLA) by radiation and its biodegradability. Eur. Polym. J. 2007, 43, 1779–1785. [Google Scholar]
- Rasal, R.M.; Janorkar, A.J.; Hirt, D.E. Poly(lactic acid) modifications. Prog. Polym. Sci. 2010, 35, 338–356. [Google Scholar] [CrossRef]
- Wang, S.; Cui, W.; Bei, J. Bulk and surface modifications of polylactide. Anal. Bioanal. Chem. 2005, 381, 547–556. [Google Scholar] [CrossRef]
- Heinrich, G.; Straube, E.; Helmis, G. Rubber elasticity of polymer networks: Theories. Adv. Polym. Sci. 1988, 85, 33–87. [Google Scholar] [CrossRef]
- Neuhaus-Steinmetz, H. Penetration Depth of the Radiation Dose and Dose Yield for Low Energy Electron Beam Accelerators. In Proceedings of the Radtech Europe, Mediterraneo, Italy, 2–6 May 1993.
- Sterilisation von Produkten für die Gesundheitsfürsorge—Strahlen Teil 2: Festlegung der Sterilisationsdosis. Available online: http://www.rki.de/DE/Content/Infekt/Krankenhaushygiene/Kommission/Downloads/Medprod_Rili_2012.pdf?__blob=publicationFile (accessed on 12 December 2013).
- Heilmann, A.; Schwegmann, C.H. Beschleunigte elektronen sterilisieren materialschonend. Dev. Med. 2010, 7–8, pp. 30–32. Available online: http://www.bgs.eu/uploads/media/DeviceMed_juli_august_2010_BGS_Fresenius_EBeam.pdf (accessed on 29 May 2013).
- Cleland, M.R. Radiation processing: Basic concepts and practical aspects. J. Ind. Radiat. Technol. 1983, 1, 191–218. [Google Scholar]
- Hiraoka, Y.; Kimura, Y.; Ueda, H.; Tabata, Y. Fabrication and biocompatibility of collagen sponge reinforced with poly(glycolic acid) fiber. Tissue Eng. 2003, 9, 1101–1112. [Google Scholar] [CrossRef]
- Melchiorri, A.J.; Hibino, N.; Fisher, J.P. Strategies and techniques to enhance the in situ endothelialization of small-diameter biodegradable polymeric vascular grafts. Tissue Eng. Part B 2013, 19. [Google Scholar] [CrossRef]
- Perán, M.; García, M.A.; Lopez-Ruiz, E.; Jiménez, G.; Marchal, J.A. How can nanotechnology help to repair the body? Advances in cardiac, skin, bone, cartilage, nerve and cardiac tissue regeneration. Materials 2013, 6, 1333–1359. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Babczyk, P.; Conzendorf, C.; Klose, J.; Schulze, M.; Harre, K.; Tobiasch, E. Stem Cells on Biomaterials for Synthetic Grafts to Promote Vascular Healing. J. Clin. Med. 2014, 3, 39-87. https://doi.org/10.3390/jcm3010039
Babczyk P, Conzendorf C, Klose J, Schulze M, Harre K, Tobiasch E. Stem Cells on Biomaterials for Synthetic Grafts to Promote Vascular Healing. Journal of Clinical Medicine. 2014; 3(1):39-87. https://doi.org/10.3390/jcm3010039
Chicago/Turabian StyleBabczyk, Patrick, Clelia Conzendorf, Jens Klose, Margit Schulze, Kathrin Harre, and Edda Tobiasch. 2014. "Stem Cells on Biomaterials for Synthetic Grafts to Promote Vascular Healing" Journal of Clinical Medicine 3, no. 1: 39-87. https://doi.org/10.3390/jcm3010039
APA StyleBabczyk, P., Conzendorf, C., Klose, J., Schulze, M., Harre, K., & Tobiasch, E. (2014). Stem Cells on Biomaterials for Synthetic Grafts to Promote Vascular Healing. Journal of Clinical Medicine, 3(1), 39-87. https://doi.org/10.3390/jcm3010039