
Traumatic Brain Injury Induces a Differential Immune Response in Polytrauma Patients; Prospective Analysis of CD69 Expression on T Cells and Platelet Expansion


Abstract
:1. Introduction
2. Material and Methods
2.1. Patients
2.2. Reagents
2.3. Blood Samples and Data Retrieval
2.4. Flow Cytometry
2.5. Thromboelastometry
2.6. Subgroup Analysis for Patients with Traumatic Brain Injury
2.7. Statistics
3. Results
3.1. Study Population
3.2. Late Increase in CD69 Expression on CD4+ Tregs
3.3. Increase in CD69 Expression in Patients without Severe TBI on CD4+ Tregs and Th17 Cells
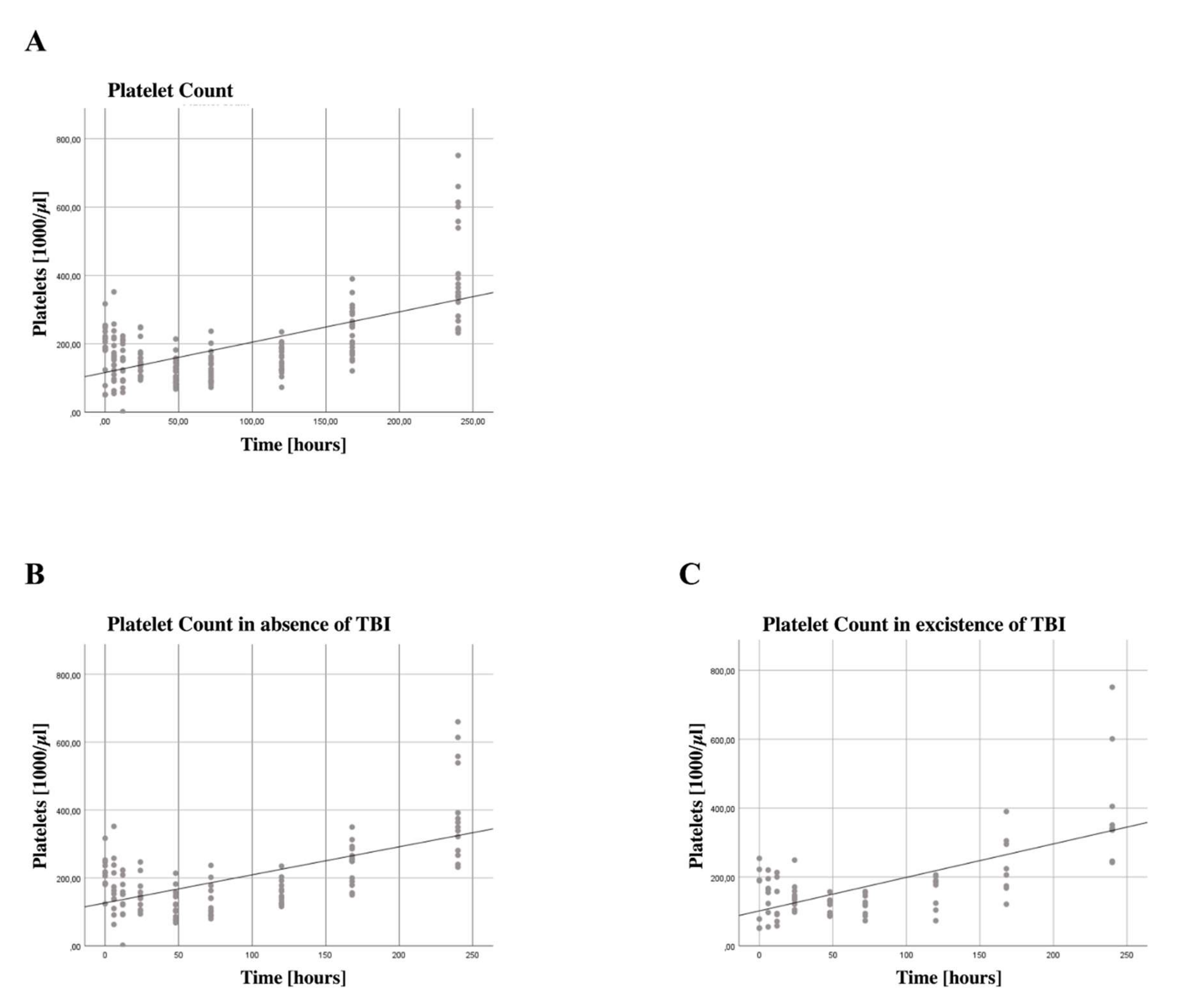
3.4. Increased Platelet Counts
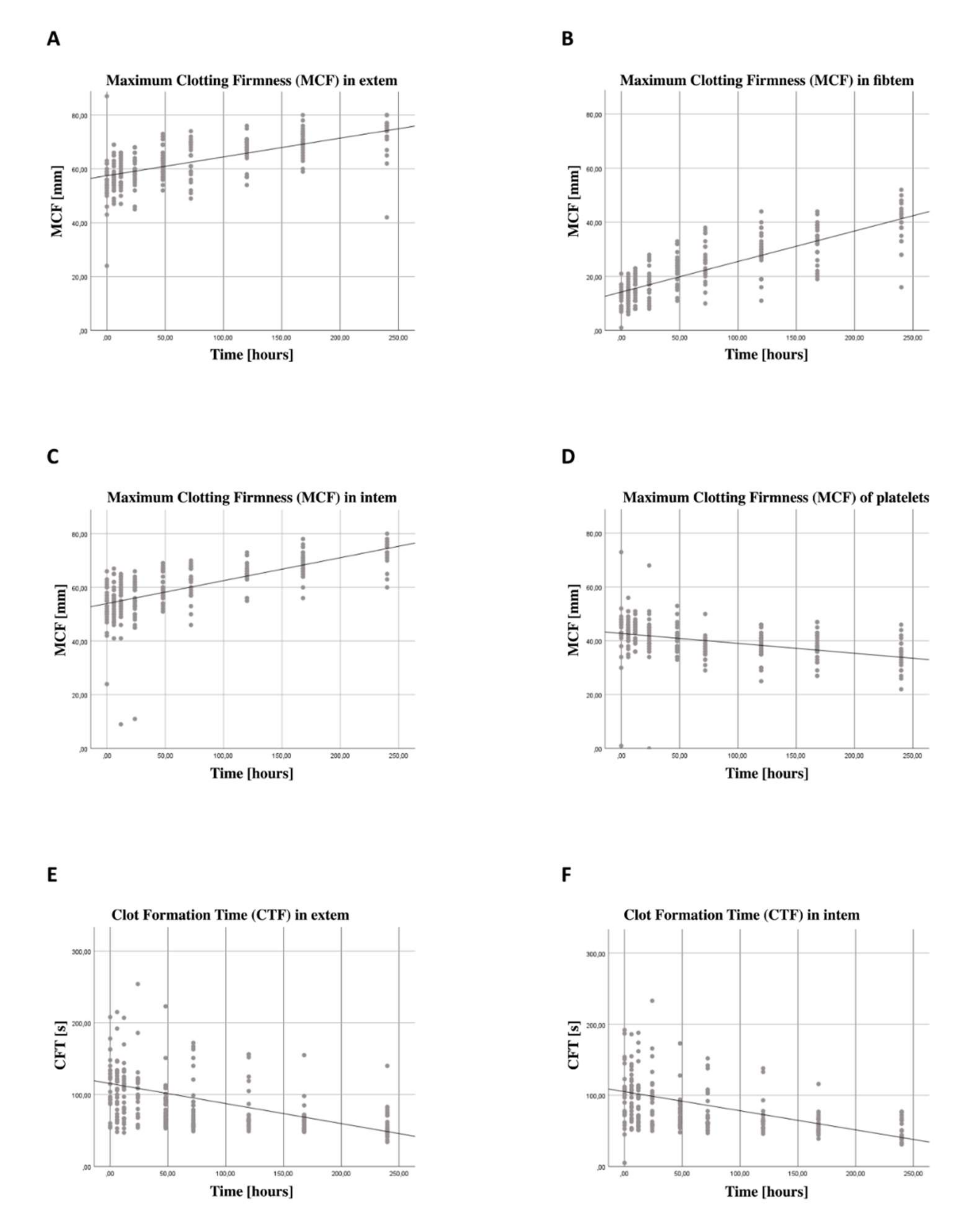
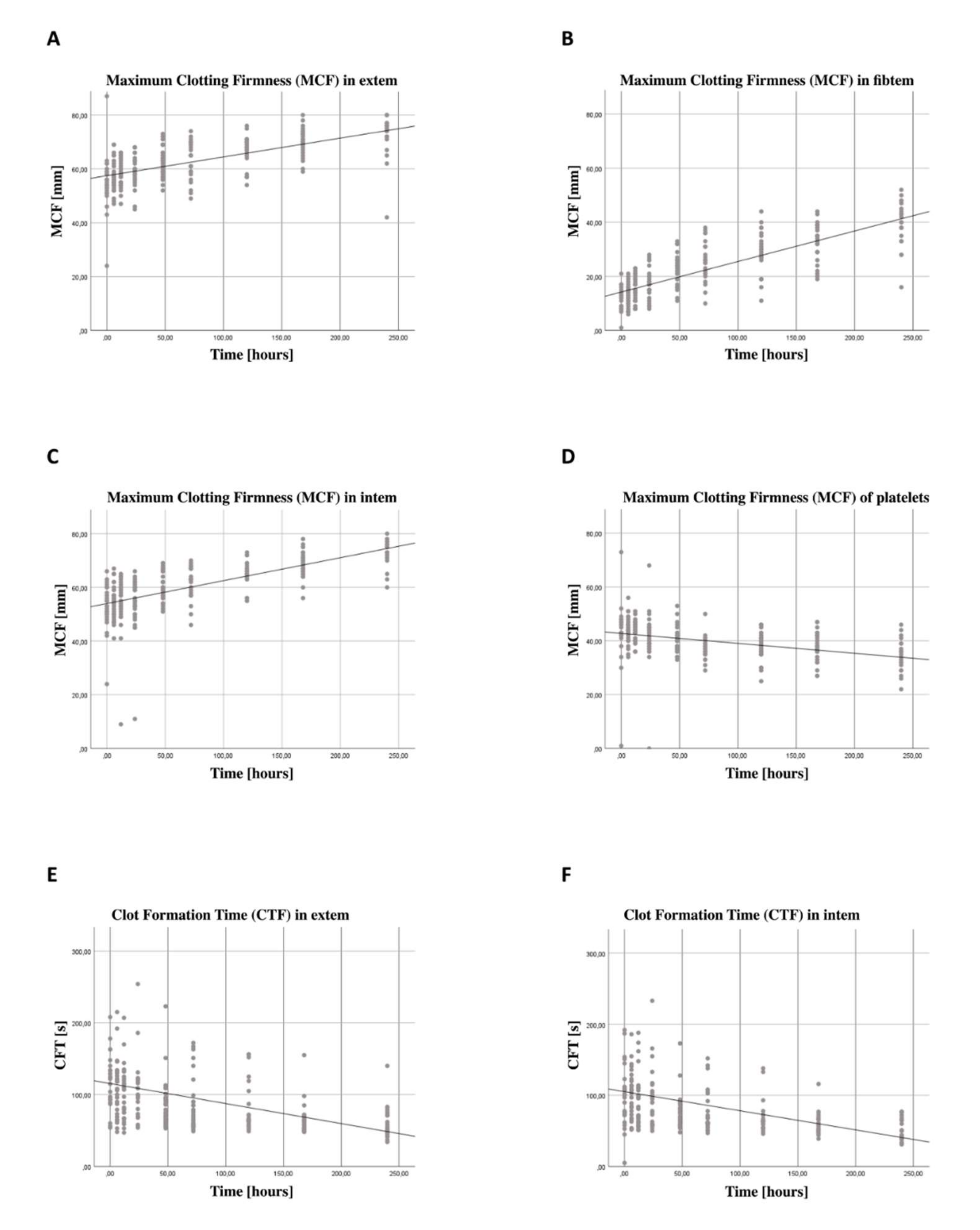
3.5. Increase in Coagulation Function Independent from Thrombocyte Function
3.6. Subgroup Analysis for Patients with Traumatic Brain Injury
4. Discussion
4.1. CD69 Expression on CD4+ Tregs and Th17 Cells
4.2. Platelet Count and Platelet Function
4.3. Thromboelastometry
4.4. Influence of TBI on the Expression of CD69 on CD4+ Tregs and Th17 Cells
4.5. Increase in Platelet Count for Severe TBI and Non-Severe TBI Patients
4.6. Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| APC | Allophycocyanin |
| AIS | Abbreviated Injury Severity Score |
| B | Regression coefficient |
| CARS | Compensatory anti-inflammatory response syndrome |
| CD | Cluster of differentiation |
| CFT | Clot formation time |
| CI | Confidence interval |
| CT | Clotting time |
| EDTA | Ethylenediaminetetraacetic acid |
| FoxP3 | Forkhead box P3 |
| GEE | Generalized estimation equations |
| GITR | Glucocorticoid-induced TNFR-related protein |
| IL | Interleukin |
| ISS | Injury severity score |
| MCF | Maximum clot firmness |
| MFI | Median fluorescence intensity |
| MODS | Multiple organ dysfunction syndrome |
| p | Probability value |
| PBA | PBS, Bovine Serum Albumin, NaN3 |
| PBS | Phosphate-buffered saline |
| PE | Phycoerythrin |
| R | Correlation coefficient |
| SE | Standard error |
| SIRS | Systemic inflammatory response syndrome |
| TBI | Traumatic brain injury |
| TCR | T cell receptor |
| Th17 | T helper 17 |
| TEG | Thromboelastography |
| TLR | Toll-like receptor |
| TNF | Tumor necrosis factor |
| TNFR2 | Tumor necrosis factor receptor 2 |
| vWF | Von Willebrand factor |
References
- Heron, M. Deaths: Leading Causes for 2019. Natl. Vital. Stat. Rep. 2021, 70, 1–114. [Google Scholar] [PubMed]
- Lenz, A.; Franklin, G.A.; Cheadle, W.G. Systemic inflammation after trauma. Injury 2007, 38, 1336–1345. [Google Scholar] [CrossRef] [PubMed]
- Hoover, L.; Bochicchio, G.V.; Napolitano, L.M.; Joshi, M.; Bochicchio, K.; Meyer, W.; Scalea, T.M. Systemic Inflammatory Response Syndrome and Nosocomial Infection in Trauma. J. Trauma 2006, 61, 310–316, discussion 316–317. [Google Scholar] [CrossRef] [PubMed]
- Keel, M.; Trentz, O. Pathophysiology of polytrauma. Injury 2005, 36, 691–709. [Google Scholar] [CrossRef]
- Baue, A.E.; Durham, R.; Faist, E. Systemic inflammatory response syndrome (SIRS), multiple organ dysfunction syndrome (MODS), multiple organ failure (MOF): Are we winning the battle? Shock 1998, 10, 79–89. [Google Scholar] [CrossRef]
- Bergmann, C.B.; Beckmann, N.; Salyer, C.E.; Hanschen, M.; Crisologo, P.A.; Caldwell, C.C. Potential Targets to Mitigate Trauma- or Sepsis-Induced Immune Suppression. Front. Immunol. 2021, 12, 622601. [Google Scholar] [CrossRef]
- Stoecklein, V.M.; Osuka, A.; Lederer, J.A. Trauma equals danger--damage control by the immune system. J. Leukoc. Biol. 2012, 92, 539–551. [Google Scholar] [CrossRef]
- Föhse, L.; Suffner, J.; Suhre, K.; Wahl, B.; Lindner, C.; Lee, C.-W.; Schmitz, S.; Haas, J.D.; Lamprecht, S.; Koenecke, C.; et al. High TCR diversity ensures optimal function andhomeostasis of Foxp3+ regulatory Tcells. Eur. J. Immunol. 2011, 41, 3101–3113. [Google Scholar] [CrossRef]
- Valencia, X.; Stephens, G.; Goldbach-Mansky, R.; Wilson, M.; Shevach, E.M.; Lipsky, P.E. TNF downmodulates the function of human CD4+CD25hi T-regulatory cells. Blood 2006, 108, 253–261. [Google Scholar] [CrossRef]
- Wan, Y.Y.; Flavell, R.A. Regulatory T-cell functions are subverted and converted owing to attenuated Foxp3 expression. Nature 2007, 445, 766–770. [Google Scholar] [CrossRef] [Green Version]
- Dasgupta, A.; Mahapatra, M.; Saxena, R. Flow cytometric immunophenotyping of regulatory T cells in chronic lymphocytic leukemia: Comparative assessment of various markers and use of novel antibody panel with CD127 as alternative to transcription factor FoxP3. Leuk. Lymphoma 2013, 54, 778–789. [Google Scholar] [CrossRef] [PubMed]
- Heuer, J.G.; Zhang, T.; Zhao, J.; Ding, C.; Cramer, M.; Justen, K.L.; Vonderfecht, S.L.; Na, S. Adoptive Transfer of In Vitro-Stimulated CD4+CD25+ Regulatory T Cells Increases Bacterial Clearance and Improves Survival in Polymicrobial Sepsis. J. Immunol. 2005, 174, 7141–7146. [Google Scholar] [CrossRef] [PubMed]
- Wisnoski, N.; Chung, C.-S.; Chen, Y.; Huang, X.; Ayala, A. The contribution of CD4+ CD25+ T-regulatory-cells to immune suppression in sepsis. Shock 2007, 27, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Hanschen, M.; Tajima, G.; O’Leary, F.; Ikeda, K.; Lederer, J.A. Injury Induces Early Activation of T-Cell Receptor Signaling Pathways in CD4+ Regulatory T Cells. Shock 2011, 35, 252–257. [Google Scholar] [CrossRef]
- Hefele, F.; Ditsch, A.; Krysiak, N.; Caldwell, C.C.; Biberthaler, P.; van Griensven, M.; Huber-Wagner, S.; Hanschen, M. Trauma Induces Interleukin-17A Expression on Th17 Cells and CD4+ Regulatory T Cells as Well as Platelet Dysfunction. Front. Immunol. 2019, 10, 2389. [Google Scholar] [CrossRef]
- Brucklacher-Waldert, V.; Steinbach, K.; Lioznov, M.; Kolster, M.; Hölscher, C.; Tolosa, E. Phenotypical Characterization of Human Th17 Cells Unambiguously Identified by Surface IL-17A Expression. J. Immunol. 2009, 183, 5494–5501. [Google Scholar] [CrossRef]
- Martin-Orozco, N.; Muranski, P.; Chung, Y.; Yang, X.O.; Yamazaki, T.; Lu, S.; Hwu, P.; Restifo, N.P.; Overwijk, W.W.; Dong, C. T Helper 17 Cells Promote Cytotoxic T Cell Activation in Tumor Immunity. Immunity 2009, 31, 787–798. [Google Scholar] [CrossRef]
- Noack, M.; Miossec, P. Th17 and regulatory T cell balance in autoimmune and inflammatory diseases. Autoimmun. Rev. 2014, 13, 668–677. [Google Scholar] [CrossRef]
- Hampton, T. Platelets’ Role in Adaptive Immunity May Contribute to Sepsis and Shock. JAMA 2018, 319, 1311–1312. [Google Scholar] [CrossRef]
- Semple, J.W.; Italiano, J.E., Jr.; Freedman, J. Platelets and the immune continuum. Nat. Rev. Immunol. 2011, 11, 264–274. [Google Scholar] [CrossRef]
- Klinger, M.H.; Jelkmann, W. Review: Role of Blood Platelets in Infection and Inflammation. J. Interf. Cytokine Res. 2002, 22, 913–922. [Google Scholar] [CrossRef] [PubMed]
- Elzey, B.D.; Sprague, D.L.; Ratliff, T.L. The emerging role of platelets in adaptive immunity. Cell. Immunol. 2005, 238, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Huang, Z.; Stålesen, R.; Hansson, G.K.; Li, N. Platelets provoke distinct dynamics of immune responses by differentially regulating CD4+ T-cell proliferation. J. Thromb. Haemost. 2014, 12, 1156–1165. [Google Scholar] [CrossRef]
- Davenport, R.A.; Brohi, K. Coagulopathy in trauma patients: Importance of thrombocyte function? Curr. Opin. Anaesthesiol. 2009, 22, 261–266. [Google Scholar] [CrossRef] [PubMed]
- MacLeod, J.B.A.; Lynn, M.; McKenney, M.G.; Cohn, S.M.; Murtha, M. Early Coagulopathy Predicts Mortality in Trauma. J. Trauma 2003, 55, 39–44. [Google Scholar] [CrossRef]
- Solomon, C.; Traintinger, S.; Ziegler, B.; Hanke, A.; Rahe-Meyer, N.; Voelckel, W.; Schöchl, H. Platelet function following trauma. A multiple electrode aggregometry study. Thromb. Haemost. 2011, 106, 322–330. [Google Scholar] [CrossRef]
- Bergmann, C.B.; Hefele, F.; Unger, M.; Huber-Wagner, S.; Biberthaler, P.; Van Griensven, M.; Hanschen, M. Platelets modulate the immune response following trauma by interaction with CD4+ T regulatory cells in a mouse model. Immunol. Res. 2016, 64, 508–517. [Google Scholar] [CrossRef]
- Bock, M.; Bergmann, C.B.; Jung, S.; Kalbitz, M.; Relja, B.; Huber-Wagner, S.; Biberthaler, P.; van Griensven, M.; Hanschen, M. The posttraumatic activation of CD4+ T regulatory cells is modulated by TNFR2- and TLR4-dependent pathways, but not by IL-10. Cell. Immunol. 2018, 331, 137–145. [Google Scholar] [CrossRef]
- Bock, M.; Bergmann, C.B.; Jung, S.; Biberthaler, P.; Heimann, L.; Hanschen, M. Platelets differentially modulate CD4+ Treg activation via GPIIa/IIIb-, fibrinogen-, and PAR4-dependent pathways. Immunol. Res. 2022, 70, 185–196. [Google Scholar] [CrossRef]
- Loane, D.J.; Kumar, A. Microglia in the TBI brain: The good, the bad, and the dysregulated. Exp. Neurol. 2016, 275 Pt 3, 316–327. [Google Scholar] [CrossRef] [Green Version]
- Maegele, M.; Schöchl, H.; Menovsky, T.; Maréchal, H.; Marklund, N.; Buki, A.; Stanworth, S. Coagulopathy and haemorrhagic progression in traumatic brain injury: Advances in mechanisms, diagnosis, and management. Lancet Neurol. 2017, 16, 630–647. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, F.; Dong, J.-F. Coagulopathy induced by traumatic brain injury: Systemic manifestation of a localized injury. Blood 2018, 131, 2001–2006. [Google Scholar] [CrossRef] [PubMed]
- Cossarizza, A.; Chang, H.; Radbruch, A.; Akdis, M.; Andrä, I.; Annunziato, F.; Bacher, P.; Barnaba, V.; Battistini, L.; Bauer, W.M.; et al. Guidelines for the use of flow cytometry and cell sorting in immunological studies. Eur. J. Immunol. 2017, 47, 1584–1797. [Google Scholar] [CrossRef] [PubMed]
- Lang, T.; Von Depka, M. Possibilities and limitations of thrombelastometry/-graphy. Hamostaseologie 2006, 26, S20–S29. [Google Scholar]
- Sancho, D.; Gómez, M.; Sánchez-Madrid, F. CD69 is an immunoregulatory molecule induced following activation. Trends Immunol. 2005, 26, 136–140. [Google Scholar] [CrossRef] [PubMed]
- González-Amaro, R.; Cortés, J.R.; Sánchez-Madrid, F.; Martín, P. Is CD69 an effective brake to control inflammatory diseases? Trends Mol. Med. 2013, 19, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Deknuydt, F.; Bioley, G.; Valmori, D.; Ayyoub, M. IL-1β and IL-2 convert human Treg into T(H)17 cells. Clin. Immunol. 2009, 131, 298–307. [Google Scholar] [CrossRef]
- Ng, M.S.Y.; Tung, J.P.; Fraser, J.F. Platelet Storage Lesions: What More Do We Know Now? Transfus. Med. Rev. 2018, 32, 144–154. [Google Scholar] [CrossRef]
- Jenne, C.N.; Urrutia, R.; Kubes, P. Platelets: Bridging hemostasis, inflammation, and immunity. Int. J. Lab. Hematol. 2013, 35, 254–261. [Google Scholar] [CrossRef]
- Gerdes, N.; Zhu, L.; Ersoy, M.; Hermansson, A.; Hjemdahl, P.; Hu, H.; Hansson, G.K.; Li, N. Platelets regulate CD4+ T-cell differentiation via multiple chemokines in humans. Thromb. Haemost. 2011, 106, 353–362. [Google Scholar] [CrossRef]
- Rugeri, L.; Levrat, A.; David, J.S.; Delecroix, E.; Floccard, B.; Gros, A.; Allaouchiche, B.; Negrier, C. Diagnosis of early coagulation abnormalities in trauma patients by rotation thrombelastography. J. Thromb. Haemost. 2007, 5, 289–295. [Google Scholar] [CrossRef]
- Engelman, D.T.; Gabram, S.G.; Allen, L.; Ens, G.E.; Jacobs, L.M. Hypercoagulability following Multiple Trauma. World J. Surg. 1996, 20, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Cap, A.; Hunt, B.J. The pathogenesis of traumatic coagulopathy. Anaesthesia 2015, 70 (Suppl. S1), 96-e34. [Google Scholar] [CrossRef] [PubMed]
- Levi, M.; van der Poll, T.; Büller, H.R. Bidirectional Relation Between Inflammation and Coagulation. Circulation 2004, 109, 2698–2704. [Google Scholar] [CrossRef]
- Schouten, M.; Wiersinga, W.J.; Levi, M.; Van Der Poll, T. Inflammation, endothelium, and coagulation in sepsis. J. Leukoc. Biol. 2008, 83, 536–545. [Google Scholar] [CrossRef]
- Bateman, R.M.; Sharpe, M.D.; Ellis, C.G. Bench-to-bedside review: Microvascular dysfunction in sepsis –hemodynamics, oxygen transport, and nitric oxide. Crit. Care 2003, 7, 359–373. [Google Scholar] [CrossRef] [PubMed]
- Alam, S.I.; Rehman, S.U.; Kim, M.O. Nicotinamide Improves Functional Recovery via Regulation of the RAGE/JNK/NF-κB Signaling Pathway after Brain Injury. J. Clin. Med. 2019, 8, 271. [Google Scholar] [CrossRef]
- Russo, M.V.; McGavern, D.B. Inflammatory neuroprotection following traumatic brain injury. Science 2016, 353, 783–785. [Google Scholar] [CrossRef]
- Raposo, C.; Graubardt, N.; Cohen, M.; Eitan, C.; London, A.; Berkutzki, T.; Schwartz, M. CNS Repair Requires Both Effector and Regulatory T Cells with Distinct Temporal and Spatial Profiles. J. Neurosci. 2014, 34, 10141–10155. [Google Scholar] [CrossRef]
- Gardner, R.C.; Yaffe, K. Epidemiology of mild traumatic brain injury and neurodegenerative disease. Mol. Cell. Neurosci. 2015, 66, 75–80. [Google Scholar] [CrossRef]
- Cui, J.; Li, H.; Chen, Z.; Dong, T.; He, X.; Wei, Y.; Li, Z.; Duan, J.; Cao, T.; Chen, Q.; et al. Thrombo-Inflammation and Immunological Response in Ischemic Stroke: Focusing on Platelet-Tregs Interaction. Front. Cell. Neurosci. 2022, 16, 955385. [Google Scholar] [CrossRef] [PubMed]
- Mortimer, J.A.; Van Duijn, C.M.; Chandra, V.; Fratiglioni, L.; Graves, A.B.; Heyman, A.; Jorm, A.F.; Kokmen, E.; Kondo, K.; Rocca, W.A.; et al. Head Trauma as a Risk Factor for Alzheimer’s Disease: A Collaborative Re-Analysis of Case-Control Studies. Int. J. Epidemiol. 1991, 20 (Suppl. S2), S28–S35. [Google Scholar] [CrossRef] [PubMed]
- Jassam, Y.N.; Izzy, S.; Whalen, M.; McGavern, D.B.; El Khoury, J. Neuroimmunology of Traumatic Brain Injury: Time for a Paradigm Shift. Neuron 2017, 95, 1246–1265. [Google Scholar] [CrossRef] [PubMed]
- Needham, E.; Helmy, A.; Zanier, E.; Jones, J.; Coles, A.; Menon, D. The immunological response to traumatic brain injury. J. Neuroimmunol. 2019, 332, 112–125. [Google Scholar] [CrossRef]
- Prodan, C.I.; Vincent, A.S.; Dale, G.L. Coated-Platelet Levels Increase with Number of Injuries in Patients with Mild Traumatic Brain Injury. J. Neurotrauma 2016, 33, 818–824. [Google Scholar] [CrossRef] [PubMed]
- Ploplis, V.A.; Donahue, D.L.; Sandoval-Cooper, M.J.; MorenoCaffaro, M.; Sheets, P.; Thomas, S.G.; Walsh, M.; Castellino, F.J. Systemic Platelet Dysfunction Is the Result of Local Dysregulated Coagulation and Platelet Activation in the Brain in a Rat Model of Isolated Traumatic Brain Injury. J. Neurotrauma 2014, 31, 1672–1675. [Google Scholar] [CrossRef]
- Castellino, F.J.; Chapman, M.P.; Donahue, D.L.; Thomas, S.; Moore, E.E.; Wohlauer, M.V.; Fritz, B.; Yount, R.; Ploplis, V.; Davis, P.; et al. Traumatic brain injury causes platelet adenosine diphosphate and arachidonic acid receptor inhibition independent of hemorrhagic shock in humans and rats. J. Trauma Acute Care Surg. 2014, 76, 1169–1176. [Google Scholar] [CrossRef] [PubMed]
- Roozenbeek, B.; Maas, A.I.R.; Menon, D.K. Changing patterns in the epidemiology of traumatic brain injury. Nat. Rev. Neurol. 2013, 9, 231–236. [Google Scholar] [CrossRef]
- Sollazzo, D.; Trabanelli, S.; Curti, A.; Vianelli, N.; Lemoli, R.M.; Catani, L. Circulating CD4+CD161+CD196+ Th17 cells are not increased in immune thrombocytopenia. Haematologica 2011, 96, 632–634. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
| Characteristics | Frequency (Percentage) | Mean ± SD |
|---|---|---|
| Patient number | 30 (100%) | |
| Age [years] | 50.5 ± 18.5 | |
| Sex | ||
| male | 18 (60%) | |
| female | 12 (40%) | |
| ISS | 33.2 ± 11.5 | |
| Survivors | 26 (87%) | |
| ICU stay [days] | 13.3 ± 15.3 | |
| Number of surgeries | 3.9 ± 2.4 | |
| AIS score | ||
| Head/neck | 20 (67%) | 2.6 ± 1.3 |
| Face | 15 (50%) | 1.7 ± 1.0 |
| Chest | 24 (80%) | 3.2 ± 1.2 |
| Abdominal/pelvic contents | 11 (37%) | 2.4 ± 1.6 |
| Extremities/pelvic girdle | 27 (90%) | 3.0 ± 0.8 |
| External | 19 (63%) | 1.3 ± 0.7 |
| Injury mechanism | ||
| Road traffic accident | 20 (66.7%) | |
| Suspected suicide attempt | 5 (16.7%) | |
| Others | 5 (16.6%) |
| Value | All Patients | Non-Severe TBI Population (AIS Head Region 1–2) | Severe TBI Population (AIS Head Region ≥ 3) | |||
|---|---|---|---|---|---|---|
| B | p-Value | B | p-Value | B | p-Value | |
| CD69 on CD4+ Tregs | 0.033 | <0.001 | 0.040 | <0.001 | 0.020 | 0.202 |
| CD69 on CD4+ Th17 cells | 0.058 | 0.093 | 0.028 | 0.003 | 0.101 | 0.202 |
| Platelet count | 23.190 | <0.001 | 20.456 | <0.001 | 28.443 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ditsch, A.; Hunold, L.; Hefele, F.; Greve, F.; Mair, O.; Biberthaler, P.; Heimann, L.; Hanschen, M. Traumatic Brain Injury Induces a Differential Immune Response in Polytrauma Patients; Prospective Analysis of CD69 Expression on T Cells and Platelet Expansion. J. Clin. Med. 2022, 11, 5315. https://doi.org/10.3390/jcm11185315
Ditsch A, Hunold L, Hefele F, Greve F, Mair O, Biberthaler P, Heimann L, Hanschen M. Traumatic Brain Injury Induces a Differential Immune Response in Polytrauma Patients; Prospective Analysis of CD69 Expression on T Cells and Platelet Expansion. Journal of Clinical Medicine. 2022; 11(18):5315. https://doi.org/10.3390/jcm11185315
Chicago/Turabian StyleDitsch, Alexander, Lea Hunold, Friederike Hefele, Frederik Greve, Olivia Mair, Peter Biberthaler, Laura Heimann, and Marc Hanschen. 2022. "Traumatic Brain Injury Induces a Differential Immune Response in Polytrauma Patients; Prospective Analysis of CD69 Expression on T Cells and Platelet Expansion" Journal of Clinical Medicine 11, no. 18: 5315. https://doi.org/10.3390/jcm11185315
APA StyleDitsch, A., Hunold, L., Hefele, F., Greve, F., Mair, O., Biberthaler, P., Heimann, L., & Hanschen, M. (2022). Traumatic Brain Injury Induces a Differential Immune Response in Polytrauma Patients; Prospective Analysis of CD69 Expression on T Cells and Platelet Expansion. Journal of Clinical Medicine, 11(18), 5315. https://doi.org/10.3390/jcm11185315