
Effect of Physical Activity, Smoking, and Sleep on Telomere Length: A Systematic Review of Observational and Intervention Studies

 ,
,  ,
,  , , ,
, , ,  ,
, 
Abstract
1. Introduction
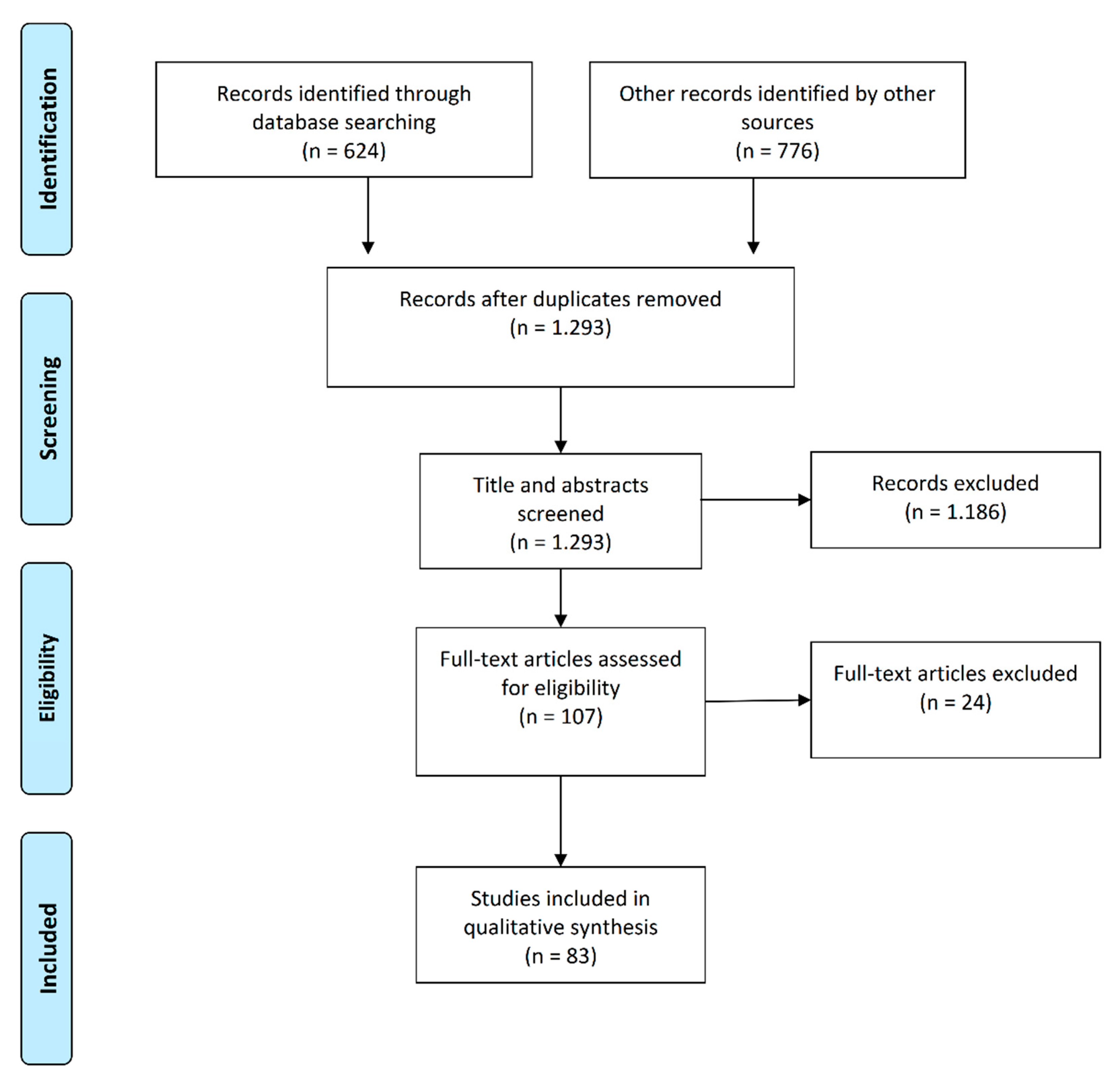
2. Materials and Methods
2.1. Search Strategy
2.2. Inclusion and Exclusion Criteria
2.3. Quality Control Analysis
3. Results
3.1. Association between PA and TL

{kind=link}
| Author, Reference | Year | Participants | TL Measurements | Tissue for TL Analysis | Exposure Assessment Methods | Results |
|---|---|---|---|---|---|---|
| Cassidy et al. [60] | 2010 | 2284 women from NHS | PCR | Blood | PA assessed by questionnaires | No association was observed between PA and TL |
| Dankel et al. [61] | 2017 | 4881 subjects from NHANES | PCR | Blood | PA assessed by questionnaires | PA was associated with increased TL, except among those who were overweight or obese |
| Ding et al. [62] | 2018 | 588 Chinese participants (18–64 y) | PCR | Blood | PA assessed by questionnaires | No associations were found between PA and TL |
| Du et al. [63] | 2012 | 7813 women (43–70 y) from NHS | PCR | Blood | PA assessed by questionnaires | Women with moderate or vigorous activity had a longer TL than less active |
| Fretts et al. [64] | 2018 | 2312 American Indian participants from SHFS | PCR | Blood | PA reported by pedometer | Participants in the upper PA quartiles had longer TL |
| Hastings et al. [65] | 2019 | 6731 participants from NHANES | PCR | Blood | PA determined with different tests (muscle strength, cardiorespiratory capacity (VO2) and walking speed | Participants with shorter telomeres performed worse on physical function tests |
| Kim et al. [66] | 2012 | 44 healthy postmenopausal women | PCR | Blood | PA reported by interview | Regular PA was associated with longer TL |
| Latifovic et al. [67] | 2016 | 477 subjects (20–50 y) | PCR | Blood | PA assessed by questionnaires | Vigorous PA related to longer TL |
| Loprinzi [68] | 2015 | 6405 participants from NHANES | PCR | Blood | Sedentary PA on screens (TV, computer …) reported by interview | Sedentary behavior based on screen free time was inversely associated with TL |
| Loprinzi et al. [69] | 2015 | 6503 participants from NHANES | PCR | Blood | PA reported by interview | Lower TL among those who performed less PA |
| Loprinzi et al. [70] | 2016 | 6474 participants from NHANES | PCR | Blood | PA reported by interview | The specific PA of running was the only activity associated with TL |
| Ogawa et al. [45] | 2017 | 6933 participants from NHANES | PCR | Blood | PA reported by interview and VO2max in subsample | Those who undertake vigorous PA present longer TL than those who practice moderate PA or do not perform PA |
| Sassenroth et al. [71] | 2015 | 815 participants from BASII | PCR | Blood | PA assessed by questionnaires | PA was associated with longer TL only when PA was regular and ≥10 y preceding the assessment |
| Savela et al. [72] | 2013 | 204 participants from HBS | SB | Blood | PA assessed by questionnaires | An U-shaped association was found. Moderate PA associated with longer TL |
| Shadyab et al. [73] | 2017 | 1476 elderly women from WHI | SB | Blood | PA assessed by questionnaires | Longer TL in the group of more active women compared to those who undertook less PA |
| Shadyab et al. [74] | 2017 | 1481 elderly women from WHI | SB | Blood | PA assessed by questionnaires and accelerometer | Longer sedentary time was associated with shorter TL in physically inactive older women |
| Shadyab et al. [75] | 2017 | 1405 elderly women from WHI | SB | Blood | PA assessed by questionnaires and accelerometer | Women at the highest level of total PA compared to the lowest level had longer TL, but significance was lost after adjustment for confounded terms |
| Silva et al. [76] | 2017 | Elderly subjects (65–85 y) with intense and moderate training, and never trained | Flow-FISH | Blood | PA assessed by questionnaires and VO2max | TL was longer in the intense training group compared to the untrained group |
| Song et al. [77] | 2010 | 170 subjects (18–80 y) | PCR ELISA | Blood | PA assessed by questionnaires | PA was inversely correlated with biomarkers of DNA damage, and these biomarkers were negatively associated with TL |
| Tucker LA [46] | 2017 | 5.823 adults from NHANES | PCR | Blood | PA assessed by questionnaires | PA was inversely associated with TL. High levels of PA were associated with longer telomeres, with a biological aging of 9 y difference compared with the sedentary ones |
| von Känel et al. [78] | 2017 | African (n = 96) and Caucasian (n = 107) schoolteachers from SAABPAS | PCR | Blood | PA measured by accelerometry | Regular PA of different intensity was not directly associated with TL |
| Williams et al. [79] | 2017 | 5674 adults from NFBC | PCR | Blood | PA determined by physical tests | TL was associated with greater aerobic fitness and muscular endurance only in young adults |
| Xue et al. [80] | 2017 | 518 adults from China | SB | Blood | PA assessed by questionnaires | Watching TV was associated with shorter TL However, doing moderate or vigorous PA was not associated with TL |
| Author, Reference | Year | Type | Participants | TL Measurements | Tissue for TL Analysis | Exposure Assessment Methods | Results |
|---|---|---|---|---|---|---|---|
| Colon et al. [81] | 2019 | CC | 7 male triathlon athletes and 7 active controls | PCR | Blood | PA assessed by questionnaires and VO2max | The triathlete subjects had longer telomeres than the active controls. Positive association between TL and VO2max |
| Denham et al. [27] | 2016 | CC | Polish endurance athletes (n = 61) and paired active controls (n = 61) | PCR GE | Blood | PA reported by questionnaires | Endurance athletes have longer TL, as well as a higher expression of TERT mRNA |
| Denham et al. [82] | 2013 | CC | 67 male ultramarathon runners and 63 controls | PCR | Blood | PA assessed by questionnaires | Ultramarathon runners had telomeres 11% longer than controls |
| LaRocca et al. [83] | 2010 | CC | 57 participants: differing by age and PA | SB | Blood | PA assessed by questionnaires and VO2max | TL of the older endurance-trained adults was greater than their sedentary peers. TL was positively related to VO2max |
| Magi et al. [84] | 2018 | CC | 26 men (20–50 y) | SB | Muscle | PA reported by interview | Regular physical training was positively with the maintenance of TL |
| Mathur et al. [85] | 2013 | CC | 17 marathon runners and 15 controls | FISH | Blood | Different tests were performed | No differences were observed in the TL of the different PA groups or tests |
| Muniesa et al. [86] | 2017 | CC | 125 young elite athletes | PCR | Blood | PA reported through a database | Elite athletes had a TL 12.4% longer than controls |
| Osthus et al. [87] | 2012 | CC | 10 young and 10 elderlies (50% endurance athletes, 50% medium level) | PCR | Muscle | PA assessed by questionnaires and VO2max | Older endurance athletes had longer TLs compared to their counterparts. These differences were not seen in the young participants |
| Puterman et al. [88] | 2010 | CC | 63 postmenopausal women | PCR | Blood | PA assessed by questionnaires | No association between PA and TL was observed. However, vigorous PA was linked to TL |
| Rae et al. [89] | 2010 | CC | 18 endurance runners vs. 19 controls | SB | Muscle | PA assessed by questionnaires | No significant differences were found between the mean and minimum TL between the endurance runners and controls |
| Simoes et al. [90] | 2017 | CC | Elite sprinters (n = 11) and untrained controls | PCR | Blood | PA reported by medical history | Elite sprinters have longer TL than their paired controls |
| Soares-Miranda et al. [91] | 2015 | L | 582 elderly subjects from CHS | SB | Blood | PA assessed by questionnaires and different tests | At baseline, higher PA was associated with longer TL. Prospective analyses show that changes in PA were associated with differences in changes in TL |
| Stenbäck et al. [92] | 2019 | L | 700 elderly subjects (Finland) | PCR | Blood | PA assessed by questionnaires and accelerometry | PA for 2-w was not associated with TL after adjustment. However, moderate PA was associated with longest TL |
| Weischer et al. [93] | 2014 | L | 4576 participants from CCHS | PCR | Blood | PA assessed by questionnaires | Increased PA was associated with short TL at baseline, but not with a change in TL during 10 years of follow-up |
| Author, Reference | Year | Participants | TL Measurements | Tissue for TL Analysis | Intervention | Results |
| Borghini et al. [94] | 2015 | 17 male endurance athletes and 32 controls | PCR | Saliva | Ultra-endurance race in athletes (330 km in a maximum time of 150 h) | Endurance athletes present longer TL than controls. Acute extreme intervention increased TL attrition compared to baseline |
| Brandao et al. [95] | 2020 | 20 obese sedentary women (20–40 y) | PCR | Blood | Combined aerobic and strength training (55 min/d, 3 times/w, 8 w) | Combination training for 8 w promoted an increase in TL, fat-free mass, and physical performance |
| Chilton et al. [96] | 2014 | 22 healthy young men, nonsmokers | GE | Blood | 30 min of continuous running on a treadmill at 80% O2 | Increased expression of the key telomeric gene TERT mRNA, associated with longer TL |
| Diman et al. [97] | 2016 | 10 healthy and moderately active young men | FISH GE | Muscle | Two cycling exercise groups: low intensity (50% VO2) and a high intensity (75% VO2). | Endurance cycling exercise increased the TERRA gene expression, a key player in telomere integrity, associated with longer TL |
| Dimauro et al. [98] | 2017 | 24 men, 12 T2D and 12 controls. | FISH | Blood | Two groups: untrained and trained (moderate PA over at least 1 y) | Individuals who participated in the regular exercise program showed longer TL |
| Friedenreich et al. [99] | 2018 | 212 Healthy women aged (50–74 y). | PCR | Blood | Two groups: aerobic exercise (45 min, 5 d/w for y, n = 99) vs. inactivity | No association between aerobic exercise and TL |
| Laye et al. [100] | 2012 | 8 participants in the “2010 Bornholm Ultramarathon 7” | PCR GE | Muscle and whole blood | Carrying out 7 marathons on 7 consecutive d | A positive regulation of gene expression of the components of the shelterin complex was found without observing changes in TL |
| Mason C et [101] | 2013 | 439 women with overweight or obesity (50–75 y) | PCR | Blood | Four groups: weight loss with diet (n = 118), PA (n = 117), diet + exercise (n = 117) and control (n = 87); for 12 m | After 12 m, no changes were seen in the TL of the intervention groups compared to the control group |
| Puterman et al. [102] | 2018 | 68 careers of elderly people | PCR | Blood | Two groups: aerobic training and control for 24 w | Significant changes in TL were observed in aerobic exercise intervention group |
| Sjögren et al. [103] | 2014 | 49 participants (68 y) with low PA | PCR | Blood | Two groups: PA and minimal PA. Intervention for 6 m | The decrease in seated time in the intervention group was associated with TL |
| Werner et al. [44] | 2019 | 125 inactive participants | PCR FlowFISH FACS | Blood | Three groups: aerobic endurance training; high intensity exercises; resistance training | Endurance training and high intensity, but not strength training, was associated with increased telomerase activity and TL |
| Zietzer et al. [104] | 2017 | 26 youth and 14 elderly participants | PCR | Blood | Randomization to 2 trials: -ISRT-1: crossover trial (2 separate sessions). -ISRT-2: 1 session of ISRT, and subsequently 5 d/w of 90 min. | Telomerase activity in young individuals increased with exercise. In contrast, short-term ISRT exercises were not associated with telomerase activity. However, after 5 w of daily ISRT, telomerase increased |
3.2. Association between Telomere Length and Tobacco Smoking
| Author, Reference | Year | Participants | TL Measurements | Tissue for TL Analysis | Exposure Assessment Methods | Results |
|---|---|---|---|---|---|---|
| Cassidy et al. [60] | 2010 | 2284 women from NHS | PCR | Blood | Tobacco smoking reported by questionnaire | Tobacco use was not associated with TL |
| Flannagan et al. [105] | 2017 | 20 smokers and 20 non-smokers | PCR | Blood | Tobacco smoking reported by questionnaire | No association was detected between smoking and TL |
| Gao et al. [106] | 2016 | 548 participants from ESTHER | PCR and methylation | Blood | Tobacco smoking reported by questionnaire | Lower methylation was observed in current smokers associated with TL |
| Huzen et al. [107] | 2014 | 8592 subjects from PREVEND | PCR | Blood | Tobacco smoking reported by questionnaire | Dose-dependent association between number of cigarettes smoked and baseline TL |
| Khan et al. [108] | 2019 | 5864 participants from NHANES | PCR | Blood | Tobacco smoking reported by questionnaire | No association was detected between TL and smoker status, but there was an inverse association between the number of cigarettes smoked and shorter TL |
| Latifovic et al. [67] | 2016 | 477 subjects (20–50 y) | PCR | Blood | Tobacco smoking reported by questionnaire | Smokers had shorter TL than those who had never smoked |
| Lei et al. [109] | 2020 | 500 African Americans from FACHS | Methylation arrays | Blood | Tobacco smoking reported by questionnaire | Associations between smoking and aging by differences in methylation |
| Lu et al. [110] | 2019 | 2256 participants from WHI and JHS; 1078 participants from FHS | SB and methylation arrays | Blood | Tobacco smoking reported by questionnaire | Being a smoker was associated with lower values of mtlDNA (level of DNA methylation as an estimator of TL). In addition, smoking was associated with a shorter TL |
| Lu et al. [111] | 2017 | 1303 non-smoking adult participants from GS:GFHS | PCR | Blood | Tobacco smoking reported by questionnaire | TL decreased more rapidly with increasing age among passive smokers compared to those who were not exposed |
| Nawrot et al. [112] | 2010 | 216 non-smokers and 89 smokers from FLEMENGHO | SB | Blood | Tobacco smoking reported by questionnaire | The TL of smokers was shorter than in non-smokers |
| Needham et al. [113] | 2013 | 5360 subjects from NHANES | PCR | Blood | Tobacco smoking reported by questionnaire (number of cigarettes/d and years of smoking) | Smokers of 60 packs of cigarettes/y or more showed significantly shorter TL than those who had never smoked |
| Patel et al. [114] | 2017 | 461 subjects NHANES | PCR and GE | Blood | Tobacco smoking reported by questionnaire | The number of cigarettes smoked/d was associated with a shorter TL, but no differences were detected in the expression levels of candidate genes |
| Rode et al. [115] | 2014 | 55,568 participants from CGPS | PCR | Blood | Tobacco smoking reported by questionnaire | An association was observed between high cumulative tobacco use and short TL |
| Song et al. [77] | 2010 | 170 adults (18–80 y) | PCR | Blood | Tobacco smoking reported by questionnaire | Inverse association between smoking and TL |
| Sulastri et al. [116] | 2017 | 130 Minangkabau men (40–50 y) | PCR | Blood | Tobacco smoking reported by questionnaire | Smoking was a risk factor for telomere shortening |
| Sun et al. [117] | 2012 | 5862 women from NHS | PCR | Blood | Tobacco smoking reported by questionnaire | Compared to current smokers, women who had never smoked had longer TLs |
| Verde et al. [118] | 2015 | 147 healthy smokers from Spain | PCR | Blood | Smoking habit was determined by survey and Fagerström test | Association between cumulative tobacco use and years of smoking and reduction in TL. No significant differences between the values of metabolized nicotine and TL |
| Weischer et al. [93] | 2014 | 4576 participants from CCHS | PCR | Blood | Tobacco smoking reported by questionnaire | Short TL was associated with increased tobacco use |
| Wulaningsih et al. [119] | 2016 | 6456 participants from NHANES | PCR | Blood | Questionnaires for smoking and serum cotinine levels were evaluated | Being a smoker was associated with a 50 bp decrease in TL compared to those who had never smoked |
| Yun et al. [120] | 2019 | 1037 adults (729 white and 308 African American) | SB | Blood | Tobacco smoking reported by questionnaire | An association was observed between smoking and TL. Body weight had a suppressing effect on this association |
| Author, Reference | Year | Type of Study | Participants | TL Measurements | Tissue for TL Analysis | Exposure Assessment Methods | Results |
|---|---|---|---|---|---|---|---|
| Zhang et al. [121] | 2016 | Longitudinal | 5624 participants from HRS, 16 y follow up | PCR | Saliva | Tobacco smoking reported by questionnaire | Shorter TL observed in smokers. The number of cigarettes was also inversely associated with TL in women, but not in men. Sex-specific prospective changes on smokers and TL were detected |
| Bendix et al. [122] | 2014 | Longitudinal | 1356 individuals (30–70 y). 10-y follow-up | PCR | Blood | Tobacco smoking reported by questionnaire | Tobacco influences the change in TL after follow-up, with smokers having a greater risk of telomere shortening |
| Chen et al. [123] | 2011 | Longitudinal | Follow-up of 271 participants at 3 times | SB | Blood | Tobacco smoking reported by survey | Smoking at baseline was associated with shorter TL and accelerated shortening |
| Marcon et al. [124] | 2017 | Twin study (case-control) | 22 homozygous twins (one smoker and the other non-smoker) | SB and methylation arrays | Blood | Tobacco smoking reported by questionnaire, and urine cotinine | Statistically significant higher values of TL in smokers compared to non-smoking twins |
| Müezzinler et al. [125] | 2015 | Longitudinal | 3600 elderly adults. Follow-up on 1000 participants | PCR | Blood | Tobacco smoking reported by questionnaire | Smoking was inversely associated with TL at baseline. On average, current smokers had 73 bp shorter telomeres, but this relationship could not be shown longitudinally |
| Strandberg et al. [126] | 2011 | Longitudinal | 622 participants from Finland (30–45 y). Follow-up at 30–40 years | SB | Blood | Tobacco smoking reported by questionnaire | Age-adjusted TL was significantly longer among lifelong non-smokers compared to past or present smokers |
| Walters et al. [127] | 2014 | Case-Control | Healthy smokers (n = 29) vs. non-smokers (n = 29) | SB | Epithelial cells or whole blood | Tobacco smoking reported by questionnaire, and urine cotinine | No differences were observed between TL and being a smoker or not |
3.3. Association between Telomere Length and Sleep
| Author, Reference | Year | Type of Study | Participants | TL Measurements | Tissue for TL Analysis | Exposure Assessment Methods | Results |
|---|---|---|---|---|---|---|---|
| Cribbet et al. [128] | 2014 | Cross-sectional | 154 subjects (45–77 y) | PCR | Blood | Sleep quality and duration by questionnaire (PSQI) | Better quality of sleep was significantly associated with longer TL. Sleep durations of >7 h/night or ≤5 h/night associated with shorter TL in older adults. Sleep duration in middle-aged adults was not associated with TL |
| Grieshober et al. [129] | 2019 | Cross-sectional | 3145 post-menopausal women (EA, AA) from the WHIS | SB | Blood | Sleep duration and sleep disturbance by questionnaires (WHIIRS) | Longer TL was associated with longer sleep duration in the whole sample (associations being strongest among AA). No associations were observed with sleep disturbances |
| Jackowska et al. [130] | 2012 | Cross-sectional | 494 men and women (mean age 63.3 y) from the WIIS | PCR | Blood | Sleep duration assessed by interview | Sleep duration was associated with TL in men, but not in women. TL was 6% shorter in men who sleep ≤5 h/night compared to >7 h/night |
| Liang et al. [43] | 2011 | Cross-sectional | 4117 women from the NHS | PCR | Blood | Sleep duration and rotating night shifts assessed by questionnaires | Sleep duration was associated with TL (compared to women in the 9 h/night, those in the <6 h/night category, had decreased TL). However, heterogeneity by age was observed (sleep duration and TL were only associated in women <50 y, and not older). No significant associations were found between rotating shift history and TL |
| Nguyen et al. [131] | 2020 | Cross-sectional | 1070 parents/careers with a (mean age 44 y), from the LSAC | PCR | Blood | Sleep behavior determined by interview and actigraphy over 8 d | Sleep duration and most other sleep metrics were not associated with TL |
| Prather et al. [132] | 2015 | Cross-sectional | 87 obese adults from a trial (San Francisco, CA) at baseline | PCR (DTC) | Blood | Sleep quality and duration by questionnaire (PSQI) | Poorer global sleep quality was associated with shorter TL in lymphocytes, but not in granulocytes. Sleep duration was not related to TL |
| Prather et al. [133] | 2011 | Cross-sectional | 245 women (49 to 66 y) | PCR | Blood | Sleep quality and duration by questionnaire (adapted from the PSQI) | Lower sleep quality was associated to shorter TL |
| Tempaku et al. [134] | 2018 | Cross-sectional | 925 participants from EPISONO cohort | PCR | Blood | Sleep quality and duration assessed by questionnaires (PSQI, UNIFESP, Epworth, ISI, PSPDN), and polysomnography | Insomnia disorders and long sleepers were associated with shorter TL |
| Wynchank et al. [135] | 2019 | Longitudinal | 2936 European subjects from the NSDA analyzed at two waves 6 y apart | PCR | Blood | Sleep duration and insomnia symptoms by questionnaire (ISR). Chronotype determined by the MCTQ | Indicators of delayed circadian rhythm, Late MSFsc, late sleep onset time, and moderately late chronotype, were associated with shorter TL at both waves. No predictors showed accelerated TL attrition over 6 y. Sleep duration and insomnia were not associated with TL |
| Zgheib et al. [136] | 2018 | Cross-sectional | 497 Lebanese (men and women >18 y) | PCR | Blood | Sleep habits determined by questionnaire (3 questions) | Difficulties to sleep (but not short sleep duration) were associated with shorter TL |
| Zhao H [137] | 2017 | Cross-sectional | 12,178 Mexican Americans (20–85 y, 80% women) from the MM:MACS | PCR | Blood | Sleep duration determined by questionnaire | TL was associated with sleeping time per d (longer in those participants who slept at least 9 h/d, followed by those who slept between 7–8 h/d, and shortest in those who slept ≤6 h/d) |
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Abdellatif, M.; Sedej, S.; Carmona-Gutierrez, D.; Madeo, F.; Kroemer, G. Autophagy in Cardiovascular Aging. Circ. Res. 2018, 123, 803–824. [Google Scholar] [CrossRef]
- Johnson, N.B.; Hayes, L.D.; Brown, K.; Hoo, E.C.; Ethier, K.A.; Centers for Disease Control and Prevention (CDC). CDC National Health Report: Leading causes of morbidity and mortality and associated behavioral risk and protective factors—United States, 2005–2013. MMWR Suppl. 2014, 63, 3–27. [Google Scholar] [PubMed]
- North, B.J.; Sinclair, D.A. The intersection between aging and cardiovascular disease. Circ. Res. 2012, 110, 1097–1108. [Google Scholar] [CrossRef] [PubMed]
- Paneni, F.; Diaz Canestro, C.; Libby, P.; Luscher, T.F.; Camici, G.G. The Aging Cardiovascular System: Understanding It at the Cellular and Clinical Levels. J. Am. Coll. Cardiol. 2017, 69, 1952–1967. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, S.; Pesce, M. Stiffness and Aging in Cardiovascular Diseases: The Dangerous Relationship between Force and Senescence. Int. J. Mol. Sci. 2021, 22, 3404. [Google Scholar] [CrossRef]
- Strait, J.B.; Lakatta, E.G. Aging-associated cardiovascular changes and their relationship to heart failure. Heart Fail. Clin. 2012, 8, 143–164. [Google Scholar] [CrossRef]
- Wagner, K.H.; Cameron-Smith, D.; Wessner, B.; Franzke, B. Biomarkers of Aging: From Function to Molecular Biology. Nutrients 2016, 8, 338. [Google Scholar] [CrossRef] [PubMed]
- Ismail, L.; Materwala, H.; Al Kaabi, J. Association of risk factors with type 2 diabetes: A systematic review. Comput. Struct. Biotechnol. J. 2021, 19, 1759–1785. [Google Scholar] [CrossRef]
- Zhu, B.; Bronas, U.G.; Fritschi, C. Sleep assessment in aging adults with type 2 diabetes: Agreement between actigraphy and sleep diaries. Sleep Med. 2018, 46, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Buford, T.W. Hypertension and aging. Ageing Res. Rev. 2016, 26, 96–111. [Google Scholar] [CrossRef]
- Fan, Z.Y.; Yang, Y.; Zhang, C.H.; Yin, R.Y.; Tang, L.; Zhang, F. Prevalence and Patterns of Comorbidity among Middle-Aged and Elderly People in China: A Cross-Sectional Study Based on CHARLS Data. Int. J. Gen. Med. 2021, 14, 1449–1455. [Google Scholar] [CrossRef] [PubMed]
- Higashi, Y.; Kihara, Y.; Noma, K. Endothelial dysfunction and hypertension in aging. Hypertens Res. 2012, 35, 1039–1047. [Google Scholar] [CrossRef] [PubMed]
- Berben, L.; Floris, G.; Wildiers, H.; Hatse, S. Cancer and Aging: Two Tightly Interconnected Biological Processes. Cancers 2021, 13, 1400. [Google Scholar] [CrossRef]
- Cano, A.; Turowski, P.; Ettcheto, M.; Duskey, J.T.; Tosi, G.; Sanchez-Lopez, E.; Garcia, M.L.; Camins, A.; Souto, E.B.; Ruiz, A.; et al. Nanomedicine-based technologies and novel biomarkers for the diagnosis and treatment of Alzheimer’s disease: From current to future challenges. J. Nanobiotechnol. 2021, 19, 122. [Google Scholar] [CrossRef]
- Fahy, G.M.; Brooke, R.T.; Watson, J.P.; Good, Z.; Vasanawala, S.S.; Maecker, H.; Leipold, M.D.; Lin, D.T.S.; Kobor, M.S.; Horvath, S. Reversal of epigenetic aging and immunosenescent trends in humans. Aging Cell 2019, 18, e13028. [Google Scholar] [CrossRef] [PubMed]
- Freitas-Simoes, T.M.; Cofan, M.; Blasco, M.A.; Soberon, N.; Foronda, M.; Serra-Mir, M.; Roth, I.; Valls-Pedret, C.; Domenech, M.; Ponferrada-Ariza, E.; et al. Walnut Consumption for Two Years and Leukocyte Telomere Attrition in Mediterranean Elders: Results of a Randomized Controlled Trial. Nutrients 2018, 10, 1907. [Google Scholar] [CrossRef]
- Pataky, M.W.; Young, W.F.; Nair, K.S. Hormonal and Metabolic Changes of Aging and the Influence of Lifestyle Modifications. Mayo Clin. Proc. 2021, 96, 788–814. [Google Scholar] [CrossRef]
- Zhang, W.; Song, M.; Qu, J.; Liu, G.H. Epigenetic Modifications in Cardiovascular Aging and Diseases. Circ. Res. 2018, 123, 773–786. [Google Scholar] [CrossRef]
- Marian, A.J.; Bhatnagar, A.; Bolli, R.; Izpisua Belmonte, J.C. Introduction to Cardiovascular Aging Compendium. Circ. Res. 2018, 123, 737–739. [Google Scholar] [CrossRef]
- Falus, A.; Marton, I.; Borbenyi, E.; Tahy, A.; Karadi, P.; Aradi, J.; Stauder, A.; Kopp, M. A challenging epigenetic message: Telomerase activity is associated with complex changes in lifestyle. Cell Biol. Int. 2011, 35, 1079–1083. [Google Scholar] [CrossRef]
- Fouquerel, E.; Opresko, P.L. Convergence of The Nobel Fields of Telomere Biology and DNA Repair. Photochem. Photobiol. 2017, 93, 229–237. [Google Scholar] [CrossRef]
- Martinez, P.; Blasco, M.A. Heart-Breaking Telomeres. Circ. Res. 2018, 123, 787–802. [Google Scholar] [CrossRef] [PubMed]
- Munoz-Lorente, M.A.; Cano-Martin, A.C.; Blasco, M.A. Mice with hyper-long telomeres show less metabolic aging and longer lifespans. Nat. Commun. 2019, 10, 4723. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Pan, X.; Liu, L.; Liu, N. Telomere length maintenance, shortening, and lengthening. J. Cell Physiol. 2014, 229, 1323–1329. [Google Scholar] [CrossRef] [PubMed]
- Bekaert, S.; De Meyer, T.; Rietzschel, E.R.; De Buyzere, M.L.; De Bacquer, D.; Langlois, M.; Segers, P.; Cooman, L.; Van Damme, P.; Cassiman, P.; et al. Telomere length and cardiovascular risk factors in a middle-aged population free of overt cardiovascular disease. Aging Cell 2007, 6, 639–647. [Google Scholar] [CrossRef] [PubMed]
- Aviv, A. Genetics of leukocyte telomere length and its role in atherosclerosis. Mutat. Res. 2012, 730, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Denham, J.; O’Brien, B.J.; Prestes, P.R.; Brown, N.J.; Charchar, F.J. Increased expression of telomere-regulating genes in endurance athletes with long leukocyte telomeres. J. Appl. Physiol. 2016, 120, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, M.; Pusceddu, I.; Marz, W.; Herrmann, W. Telomere biology and age-related diseases. Clin. Chem. Lab. Med. 2018, 56, 1210–1222. [Google Scholar] [CrossRef]
- Mons, U.; Muezzinler, A.; Schottker, B.; Dieffenbach, A.K.; Butterbach, K.; Schick, M.; Peasey, A.; De Vivo, I.; Trichopoulou, A.; Boffetta, P.; et al. Leukocyte Telomere Length and All-Cause, Cardiovascular Disease, and Cancer Mortality: Results From Individual-Participant-Data Meta-Analysis of 2 Large Prospective Cohort Studies. Am. J. Epidemiol. 2017, 185, 1317–1326. [Google Scholar] [CrossRef]
- Rizvi, S.; Raza, S.T.; Mahdi, F. Telomere length variations in aging and age-related diseases. Curr. Aging Sci. 2014, 7, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Goglin, S.E.; Farzaneh-Far, R.; Epel, E.S.; Lin, J.; Blackburn, E.H.; Whooley, M.A. Change in Leukocyte Telomere Length Predicts Mortality in Patients with Stable Coronary Heart Disease from the Heart and Soul Study. PLoS ONE 2016, 11, e0160748. [Google Scholar] [CrossRef]
- Turner, K.J.; Vasu, V.; Griffin, D.K. Telomere Biology and Human Phenotype. Cells 2019, 8, 73. [Google Scholar] [CrossRef] [PubMed]
- Brouilette, S.; Singh, R.K.; Thompson, J.R.; Goodall, A.H.; Samani, N.J. White cell telomere length and risk of premature myocardial infarction. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 842–846. [Google Scholar] [CrossRef] [PubMed]
- Brouilette, S.W.; Moore, J.S.; McMahon, A.D.; Thompson, J.R.; Ford, I.; Shepherd, J.; Packard, C.J.; Samani, N.J.; West of Scotland Coronary Prevention Study, G. Telomere length, risk of coronary heart disease, and statin treatment in the West of Scotland Primary Prevention Study: A nested case-control study. Lancet 2007, 369, 107–114. [Google Scholar] [CrossRef]
- Carty, C.L.; Kooperberg, C.; Liu, J.; Herndon, M.; Assimes, T.; Hou, L.; Kroenke, C.H.; LaCroix, A.Z.; Kimura, M.; Aviv, A.; et al. Leukocyte Telomere Length and Risks of Incident Coronary Heart Disease and Mortality in a Racially Diverse Population of Postmenopausal Women. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 2225–2231. [Google Scholar] [CrossRef]
- De Meyer, T.; Nawrot, T.; Bekaert, S.; De Buyzere, M.L.; Rietzschel, E.R.; Andres, V. Telomere Length as Cardiovascular Aging Biomarker: JACC Review Topic of the Week. J. Am. Coll. Cardiol. 2018, 72, 805–813. [Google Scholar] [CrossRef]
- Haycock, P.C.; Heydon, E.E.; Kaptoge, S.; Butterworth, A.S.; Thompson, A.; Willeit, P. Leucocyte telomere length and risk of cardiovascular disease: Systematic review and meta-analysis. BMJ 2014, 349, g4227. [Google Scholar] [CrossRef]
- Okuda, K.; Khan, M.Y.; Skurnick, J.; Kimura, M.; Aviv, H.; Aviv, A. Telomere attrition of the human abdominal aorta: Relationships with age and atherosclerosis. Atherosclerosis 2000, 152, 391–398. [Google Scholar] [CrossRef]
- Sharifi-Sanjani, M.; Oyster, N.M.; Tichy, E.D.; Bedi, K.C., Jr.; Harel, O.; Margulies, K.B.; Mourkioti, F. Cardiomyocyte-Specific Telomere Shortening is a Distinct Signature of Heart Failure in Humans. J. Am. Heart Assoc. 2017, 6, 25211. [Google Scholar] [CrossRef]
- Willeit, P.; Willeit, J.; Brandstatter, A.; Ehrlenbach, S.; Mayr, A.; Gasperi, A.; Weger, S.; Oberhollenzer, F.; Reindl, M.; Kronenberg, F.; et al. Cellular aging reflected by leukocyte telomere length predicts advanced atherosclerosis and cardiovascular disease risk. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 1649–1656. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, E.H.; Epel, E.S.; Lin, J. Human telomere biology: A contributory and interactive factor in aging, disease risks, and protection. Science 2015, 350, 1193–1198. [Google Scholar] [CrossRef] [PubMed]
- Opresko, P.L.; Shay, J.W. Telomere-associated aging disorders. Ageing Res. Rev. 2017, 33, 52–66. [Google Scholar] [CrossRef] [PubMed]
- Liang, G.; Schernhammer, E.; Qi, L.; Gao, X.; De Vivo, I.; Han, J. Associations between rotating night shifts, sleep duration, and telomere length in women. PLoS ONE 2011, 6, e23462. [Google Scholar] [CrossRef] [PubMed]
- Werner, C.M.; Hecksteden, A.; Morsch, A.; Zundler, J.; Wegmann, M.; Kratzsch, J.; Thiery, J.; Hohl, M.; Bittenbring, J.T.; Neumann, F.; et al. Differential effects of endurance, interval, and resistance training on telomerase activity and telomere length in a randomized, controlled study. Eur. Heart J. 2019, 40, 34–46. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, E.F.; Leveille, S.G.; Wright, J.A.; Shi, L.; Camhi, S.M.; You, T. Physical Activity Domains/Recommendations and Leukocyte Telomere Length in U.S. Adults. Med. Sci. Sports Exerc. 2017, 49, 1375–1382. [Google Scholar] [CrossRef] [PubMed]
- Tucker, L.A. Physical activity and telomere length in U.S. men and women: An NHANES investigation. Prev. Med. 2017, 100, 145–151. [Google Scholar] [CrossRef]
- Crous-Bou, M.; Molinuevo, J.L.; Sala-Vila, A. Plant-Rich Dietary Patterns, Plant Foods and Nutrients, and Telomere Length. Adv. Nutr. 2019, 10, S296–S303. [Google Scholar] [CrossRef]
- Freitas-Simoes, T.M.; Ros, E.; Sala-Vila, A. Nutrients, foods, dietary patterns and telomere length: Update of epidemiological studies and randomized trials. Metabolism 2016, 65, 406–415. [Google Scholar] [CrossRef]
- Galie, S.; Canudas, S.; Muralidharan, J.; Garcia-Gavilan, J.; Bullo, M.; Salas-Salvado, J. Impact of Nutrition on Telomere Health: Systematic Review of Observational Cohort Studies and Randomized Clinical Trials. Adv. Nutr. 2020, 11, 576–601. [Google Scholar] [CrossRef]
- Marti, A.; Echeverria, R.; Morell-Azanza, L.; Ojeda-Rodriguez, A. Telomeres and diet quality. Nutr. Hosp. 2017, 34, 1226–1245. [Google Scholar] [CrossRef]
- Perez, L.M.; Amaral, M.A.; Mundstock, E.; Barbe-Tuana, F.M.; Guma, F.; Jones, M.H.; Machado, D.C.; Sarria, E.E.; Marques, E.M.M.; Preto, L.T.; et al. Effects of Diet on Telomere Length: Systematic Review and Meta-Analysis. Public Health Genom. 2017, 20, 286–292. [Google Scholar] [CrossRef] [PubMed]
- Rafie, N.; Golpour Hamedani, S.; Barak, F.; Safavi, S.M.; Miraghajani, M. Dietary patterns, food groups and telomere length: A systematic review of current studies. Eur. J. Clin. Nutr. 2017, 71, 151–158. [Google Scholar] [CrossRef]
- Bray, N.W.; Pieruccini-Faria, F.; Bartha, R.; Doherty, T.J.; Nagamatsu, L.S.; Montero-Odasso, M. The effect of physical exercise on functional brain network connectivity in older adults with and without cognitive impairment. A systematic review. Mech. Ageing Dev. 2021, 196, 111493. [Google Scholar] [CrossRef] [PubMed]
- Duggal, N.A.; Niemiro, G.; Harridge, S.D.R.; Simpson, R.J.; Lord, J.M. Can physical activity ameliorate immunosenescence and thereby reduce age-related multi-morbidity? Nat. Rev. Immunol. 2019, 19, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Harridge, S.D.; Lazarus, N.R. Physical Activity, Aging, and Physiological Function. Physiology 2017, 32, 152–161. [Google Scholar] [CrossRef]
- Jakovljevic, D.G. Physical activity and cardiovascular aging: Physiological and molecular insights. Exp. Gerontol. 2018, 109, 67–74. [Google Scholar] [CrossRef]
- Orkaby, A.R.; Forman, D.E. Physical activity and CVD in older adults: An expert’s perspective. Expert Rev. Cardiovasc. Ther. 2018, 16, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Rosique-Esteban, N.; Diaz-Lopez, A.; Martinez-Gonzalez, M.A.; Corella, D.; Goday, A.; Martinez, J.A.; Romaguera, D.; Vioque, J.; Aros, F.; Garcia-Rios, A.; et al. Leisure-time physical activity, sedentary behaviors, sleep, and cardiometabolic risk factors at baseline in the PREDIMED-PLUS intervention trial: A cross-sectional analysis. PLoS ONE 2017, 12, e0172253. [Google Scholar] [CrossRef]
- Talar, K.; Hernandez-Belmonte, A.; Vetrovsky, T.; Steffl, M.; Kalamacka, E.; Courel-Ibanez, J. Benefits of Resistance Training in Early and Late Stages of Frailty and Sarcopenia: A Systematic Review and Meta-Analysis of Randomized Controlled Studies. J. Clin. Med. 2021, 10, 1630. [Google Scholar] [CrossRef]
- Cassidy, A.; De Vivo, I.; Liu, Y.; Han, J.; Prescott, J.; Hunter, D.J.; Rimm, E.B. Associations between diet, lifestyle factors, and telomere length in women. Am. J. Clin. Nutr. 2010, 91, 1273–1280. [Google Scholar] [CrossRef] [PubMed]
- Dankel, S.J.; Loenneke, J.P.; Loprinzi, P.D. The impact of overweight/obesity duration and physical activity on telomere length: An application of the WATCH paradigm. Obes. Res. Clin. Pract. 2017, 11, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.X.; Zhang, Y.H.; Xu, X.Z.; Zhang, J.; Ming, S.U.N.; Liu, D.; Zhao, Z.Y.; Zhou, Y.; Zhang, Q.; Wang, Y.X. Association between Physical Activity and Telomere Length in a North Chinese Population: A China Suboptimal Health Cohort Study. Biomed. Environ. Sci. 2018, 31, 394–398. [Google Scholar] [CrossRef] [PubMed]
- Du, M.; Prescott, J.; Kraft, P.; Han, J.; Giovannucci, E.; Hankinson, S.E.; De Vivo, I. Physical activity, sedentary behavior, and leukocyte telomere length in women. Am. J. Epidemiol. 2012, 175, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Fretts, A.M.; Mete, M.; Howard, B.V.; Best, L.G.; Siscovick, D.S.; Eilat-Adar, S.; Zhao, J. Physical activity and telomere length in American Indians: The Strong Heart Study. Eur. J. Epidemiol. 2018, 33, 497–500. [Google Scholar] [CrossRef] [PubMed]
- Hastings, W.J.; Shalev, I.; Belsky, D.W. Comparability of biological aging measures in the National Health and Nutrition Examination Study, 1999–2002. Psychoneuroendocrinology 2019, 106, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Ko, J.H.; Lee, D.C.; Lim, I.; Bang, H. Habitual physical exercise has beneficial effects on telomere length in postmenopausal women. Menopause 2012, 19, 1109–1115. [Google Scholar] [CrossRef]
- Latifovic, L.; Peacock, S.D.; Massey, T.E.; King, W.D. The Influence of Alcohol Consumption, Cigarette Smoking, and Physical Activity on Leukocyte Telomere Length. Cancer Epidemiol. Biomark. Prev. 2016, 25, 374–380. [Google Scholar] [CrossRef]
- Loprinzi, P.D. Leisure-Time Screen-Based Sedentary Behavior and Leukocyte Telomere Length: Implications for a New Leisure-Time Screen-Based Sedentary Behavior Mechanism. Mayo Clin. Proc. 2015, 90, 786–790. [Google Scholar] [CrossRef] [PubMed]
- Loprinzi, P.D.; Loenneke, J.P.; Blackburn, E.H. Movement-Based Behaviors and Leukocyte Telomere Length among US Adults. Med. Sci. Sports Exerc. 2015, 47, 2347–2352. [Google Scholar] [CrossRef] [PubMed]
- Loprinzi, P.D.; Sng, E. Mode-specific physical activity and leukocyte telomere length among U.S. adults: Implications of running on cellular aging. Prev. Med. 2016, 85, 17–19. [Google Scholar] [CrossRef]
- Sassenroth, D.; Meyer, A.; Salewsky, B.; Kroh, M.; Norman, K.; Steinhagen-Thiessen, E.; Demuth, I. Sports and Exercise at Different Ages and Leukocyte Telomere Length in Later Life—Data from the Berlin Aging Study II (BASE-II). PLoS ONE 2015, 10, e0142131. [Google Scholar] [CrossRef]
- Savela, S.; Saijonmaa, O.; Strandberg, T.E.; Koistinen, P.; Strandberg, A.Y.; Tilvis, R.S.; Pitkala, K.H.; Miettinen, T.A.; Fyhrquist, F. Physical activity in midlife and telomere length measured in old age. Exp. Gerontol. 2013, 48, 81–84. [Google Scholar] [CrossRef] [PubMed]
- Shadyab, A.H.; LaMonte, M.J.; Kooperberg, C.; Reiner, A.P.; Carty, C.L.; Manini, T.M.; Hou, L.; Di, C.; Macera, C.A.; Gallo, L.C.; et al. Leisure-time physical activity and leukocyte telomere length among older women. Exp. Gerontol. 2017, 95, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Shadyab, A.H.; Macera, C.A.; Shaffer, R.A.; Jain, S.; Gallo, L.C.; LaMonte, M.J.; Reiner, A.P.; Kooperberg, C.; Carty, C.L.; Di, C.; et al. Associations of Accelerometer-Measured and Self-Reported Sedentary Time With Leukocyte Telomere Length in Older Women. Am. J. Epidemiol. 2017, 185, 172–184. [Google Scholar] [CrossRef] [PubMed]
- Shadyab, A.H.; LaMonte, M.J.; Kooperberg, C.; Reiner, A.P.; Carty, C.L.; Manini, T.M.; Hou, L.; Di, C.; LaCroix, A.Z. Association of Accelerometer-Measured Physical Activity With Leukocyte Telomere Length Among Older Women. J. Gerontol. A Biol. Sci. Med. Sci. 2017, 72, 1532–1537. [Google Scholar] [CrossRef]
- Silva, L.C.; de Araujo, A.L.; Fernandes, J.R.; Matias Mde, S.; Silva, P.R.; Duarte, A.J.; Garcez Leme, L.E.; Benard, G. Moderate and intense exercise lifestyles attenuate the effects of aging on telomere length and the survival and composition of T cell subpopulations. Age 2016, 38, 24. [Google Scholar] [CrossRef]
- Song, Z.; von Figura, G.; Liu, Y.; Kraus, J.M.; Torrice, C.; Dillon, P.; Rudolph-Watabe, M.; Ju, Z.; Kestler, H.A.; Sanoff, H.; et al. Lifestyle impacts on the aging-associated expression of biomarkers of DNA damage and telomere dysfunction in human blood. Aging Cell 2010, 9, 607–615. [Google Scholar] [CrossRef] [PubMed]
- von Kanel, R.; Bruwer, E.J.; Hamer, M.; de Ridder, J.H.; Malan, L. Association between objectively measured physical activity, chronic stress and leukocyte telomere length. J. Sports Med. Phys. Fit. 2017, 57, 1349–1358. [Google Scholar] [CrossRef]
- Williams, D.M.; Buxton, J.L.; Kantomaa, M.T.; Tammelin, T.H.; Blakemore, A.I.F.; Jarvelin, M.R. Associations of Leukocyte Telomere Length With Aerobic and Muscular Fitness in Young Adults. Am. J. Epidemiol. 2017, 185, 529–537. [Google Scholar] [CrossRef] [PubMed]
- Xue, H.M.; Liu, Q.Q.; Tian, G.; Quan, L.M.; Zhao, Y.; Cheng, G. Television Watching and Telomere Length Among Adults in Southwest China. Am. J. Public Health 2017, 107, 1425–1432. [Google Scholar] [CrossRef] [PubMed]
- Colon, M.; Hodgson, A.; Donlon, E.; Murphy, J.E.J. Effects of Competitive Triathlon Training on Telomere Length. J Aging Phys. Act. 2019, 27, 510–514. [Google Scholar] [CrossRef]
- Denham, J.; Nelson, C.P.; O’Brien, B.J.; Nankervis, S.A.; Denniff, M.; Harvey, J.T.; Marques, F.Z.; Codd, V.; Zukowska-Szczechowska, E.; Samani, N.J.; et al. Longer leukocyte telomeres are associated with ultra-endurance exercise independent of cardiovascular risk factors. PLoS ONE 2013, 8, e69377. [Google Scholar] [CrossRef]
- LaRocca, T.J.; Seals, D.R.; Pierce, G.L. Leukocyte telomere length is preserved with aging in endurance exercise-trained adults and related to maximal aerobic capacity. Mech. Ageing Dev. 2010, 131, 165–167. [Google Scholar] [CrossRef]
- Magi, F.; Dimauro, I.; Margheritini, F.; Duranti, G.; Mercatelli, N.; Fantini, C.; Ripani, F.R.; Sabatini, S.; Caporossi, D. Telomere length is independently associated with age, oxidative biomarkers, and sport training in skeletal muscle of healthy adult males. Free Radic. Res. 2018, 52, 639–647. [Google Scholar] [CrossRef] [PubMed]
- Mathur, S.; Ardestani, A.; Parker, B.; Cappizzi, J.; Polk, D.; Thompson, P.D. Telomere length and cardiorespiratory fitness in marathon runners. J. Investig. Med. 2013, 61, 613–615. [Google Scholar] [CrossRef] [PubMed]
- Muniesa, C.A.; Verde, Z.; Diaz-Urena, G.; Santiago, C.; Gutierrez, F.; Diaz, E.; Gomez-Gallego, F.; Pareja-Galeano, H.; Soares-Miranda, L.; Lucia, A. Telomere Length in Elite Athletes. Int. J. Sports Physiol. Perform. 2017, 12, 994–996. [Google Scholar] [CrossRef]
- Osthus, I.B.; Sgura, A.; Berardinelli, F.; Alsnes, I.V.; Bronstad, E.; Rehn, T.; Stobakk, P.K.; Hatle, H.; Wisloff, U.; Nauman, J. Telomere length and long-term endurance exercise: Does exercise training affect biological age? A pilot study. PLoS ONE 2012, 7, e52769. [Google Scholar] [CrossRef]
- Puterman, E.; Lin, J.; Blackburn, E.; O’Donovan, A.; Adler, N.; Epel, E. The power of exercise: Buffering the effect of chronic stress on telomere length. PLoS ONE 2010, 5, e10837. [Google Scholar] [CrossRef]
- Rae, D.E.; Vignaud, A.; Butler-Browne, G.S.; Thornell, L.E.; Sinclair-Smith, C.; Derman, E.W.; Lambert, M.I.; Collins, M. Skeletal muscle telomere length in healthy, experienced, endurance runners. Eur. J. Appl. Physiol. 2010, 109, 323–330. [Google Scholar] [CrossRef]
- Simoes, H.G.; Sousa, C.V.; Dos Santos Rosa, T.; da Silva Aguiar, S.; Deus, L.A.; Rosa, E.; Amato, A.A.; Andrade, R.V. Longer Telomere Length in Elite Master Sprinters: Relationship to Performance and Body Composition. Int. J. Sports Med. 2017, 38, 1111–1116. [Google Scholar] [CrossRef]
- Soares-Miranda, L.; Imamura, F.; Siscovick, D.; Jenny, N.S.; Fitzpatrick, A.L.; Mozaffarian, D. Physical Activity, Physical Fitness, and Leukocyte Telomere Length: The Cardiovascular Health Study. Med. Sci. Sports Exerc. 2015, 47, 2525–2534. [Google Scholar] [CrossRef] [PubMed]
- Stenback, V.; Mutt, S.J.; Leppaluoto, J.; Gagnon, D.D.; Makela, K.A.; Jokelainen, J.; Keinanen-Kiukaanniemi, S.; Herzig, K.H. Association of Physical Activity With Telomere Length Among Elderly Adults-The Oulu Cohort 1945. Front. Physiol. 2019, 10, 444. [Google Scholar] [CrossRef]
- Weischer, M.; Bojesen, S.E.; Nordestgaard, B.G. Telomere shortening unrelated to smoking, body weight, physical activity, and alcohol intake: 4,576 general population individuals with repeat measurements 10 years apart. PLoS Genet. 2014, 10, e1004191. [Google Scholar] [CrossRef] [PubMed]
- Borghini, A.; Giardini, G.; Tonacci, A.; Mastorci, F.; Mercuri, A.; Mrakic-Sposta, S.; Moretti, S.; Andreassi, M.G.; Pratali, L. Chronic and acute effects of endurance training on telomere length. Mutagenesis 2015, 30, 711–716. [Google Scholar] [CrossRef] [PubMed]
- Brandao, C.F.C.; Nonino, C.B.; de Carvalho, F.G.; Nicoletti, C.F.; Noronha, N.Y.; San Martin, R.; de Freitas, E.C.; Junqueira-Franco, M.V.M.; Marchini, J.S. The effects of short-term combined exercise training on telomere length in obese women: A prospective, interventional study. Sports Med. Open 2020, 6, 5. [Google Scholar] [CrossRef]
- Chilton, W.L.; Marques, F.Z.; West, J.; Kannourakis, G.; Berzins, S.P.; O’Brien, B.J.; Charchar, F.J. Acute exercise leads to regulation of telomere-associated genes and microRNA expression in immune cells. PLoS ONE 2014, 9, e92088. [Google Scholar] [CrossRef]
- Diman, A.; Boros, J.; Poulain, F.; Rodriguez, J.; Purnelle, M.; Episkopou, H.; Bertrand, L.; Francaux, M.; Deldicque, L.; Decottignies, A. Nuclear respiratory factor 1 and endurance exercise promote human telomere transcription. Sci. Adv. 2016, 2, e1600031. [Google Scholar] [CrossRef]
- Dimauro, I.; Sgura, A.; Pittaluga, M.; Magi, F.; Fantini, C.; Mancinelli, R.; Sgadari, A.; Fulle, S.; Caporossi, D. Regular exercise participation improves genomic stability in diabetic patients: An exploratory study to analyse telomere length and DNA damage. Sci. Rep. 2017, 7, 4137. [Google Scholar] [CrossRef]
- Friedenreich, C.M.; Wang, Q.; Ting, N.S.; Brenner, D.R.; Conroy, S.M.; McIntyre, J.B.; Mickle, A.; Courneya, K.S.; Beattie, T. Effect of a 12-month exercise intervention on leukocyte telomere length: Results from the ALPHA Trial. Cancer Epidemiol. 2018, 56, 67–74. [Google Scholar] [CrossRef]
- Laye, M.J.; Solomon, T.P.; Karstoft, K.; Pedersen, K.K.; Nielsen, S.D.; Pedersen, B.K. Increased shelterin mRNA expression in peripheral blood mononuclear cells and skeletal muscle following an ultra-long-distance running event. J. Appl. Physiol. 2012, 112, 773–781. [Google Scholar] [CrossRef] [PubMed]
- Mason, C.; Risques, R.A.; Xiao, L.; Duggan, C.R.; Imayama, I.; Campbell, K.L.; Kong, A.; Foster-Schubert, K.E.; Wang, C.Y.; Alfano, C.M.; et al. Independent and combined effects of dietary weight loss and exercise on leukocyte telomere length in postmenopausal women. Obesity 2013, 21, E549–E554. [Google Scholar] [CrossRef] [PubMed]
- Puterman, E.; Weiss, J.; Lin, J.; Schilf, S.; Slusher, A.L.; Johansen, K.L.; Epel, E.S. Aerobic exercise lengthens telomeres and reduces stress in family caregivers: A randomized controlled trial-Curt Richter Award Paper 2018. Psychoneuroendocrinology 2018, 98, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Sjögren, P.; Fisher, R.; Kallings, L.; Svenson, U.; Roos, G.; Hellenius, M.L. Stand up for health--avoiding sedentary behaviour might lengthen your telomeres: Secondary outcomes from a physical activity RCT in older people. Br. J. Sports Med. 2014, 48, 1407–1409. [Google Scholar] [CrossRef] [PubMed]
- Zietzer, A.; Buschmann, E.E.; Janke, D.; Li, L.; Brix, M.; Meyborg, H.; Stawowy, P.; Jungk, C.; Buschmann, I.; Hillmeister, P. Acute physical exercise and long-term individual shear rate therapy increase telomerase activity in human peripheral blood mononuclear cells. Acta Physiol. 2017, 220, 251–262. [Google Scholar] [CrossRef]
- Flannagan, K.S.; Jansen, E.C.; Rozek, L.S.; Rentschler, K.M.; Roman, A.V.; Ramirez-Zea, M.; Villamor, E.; Nine Mesoamerican Countries Metabolic Syndrome Study (NiMeCoMeS) Group. Sociodemographic correlates and family aggregation of leukocyte telomere length in adults and children from Mesoamerica. Am. J. Hum. Biol. 2017, 29, e22942. [Google Scholar] [CrossRef]
- Gao, X.; Mons, U.; Zhang, Y.; Breitling, L.P.; Brenner, H. DNA methylation changes in response to active smoking exposure are associated with leukocyte telomere length among older adults. Eur. J. Epidemiol. 2016, 31, 1231–1241. [Google Scholar] [CrossRef] [PubMed]
- Huzen, J.; Wong, L.S.; van Veldhuisen, D.J.; Samani, N.J.; Zwinderman, A.H.; Codd, V.; Cawthon, R.M.; Benus, G.F.; van der Horst, I.C.; Navis, G.; et al. Telomere length loss due to smoking and metabolic traits. J. Intern. Med. 2014, 275, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.J.; Gebreab, S.Y.; Gaye, A.; Crespo, P.R.; Xu, R.; Davis, S.K. Associations of smoking indicators and cotinine levels with telomere length: National Health and Nutrition Examination Survey. Prev. Med. Rep. 2019, 15, 100895. [Google Scholar] [CrossRef] [PubMed]
- Lei, M.K.; Gibbons, F.X.; Simons, R.L.; Philibert, R.A.; Beach, S.R.H. The Effect of Tobacco Smoking Differs across Indices of DNA Methylation-Based Aging in an African American Sample: DNA Methylation-Based Indices of Smoking Capture These Effects. Genes 2020, 11, 311. [Google Scholar] [CrossRef] [PubMed]
- Lu, A.T.; Seeboth, A.; Tsai, P.C.; Sun, D.; Quach, A.; Reiner, A.P.; Kooperberg, C.; Ferrucci, L.; Hou, L.; Baccarelli, A.A.; et al. DNA methylation-based estimator of telomere length. Aging 2019, 11, 5895–5923. [Google Scholar] [CrossRef]
- Lu, L.; Johnman, C.; McGlynn, L.; Mackay, D.F.; Shiels, P.G.; Pell, J.P. Association between exposure to second-hand smoke and telomere length: Cross-sectional study of 1303 non-smokers. Int. J. Epidemiol. 2017, 46, 1978–1984. [Google Scholar] [CrossRef] [PubMed]
- Nawrot, T.S.; Staessen, J.A.; Holvoet, P.; Struijker-Boudier, H.A.; Schiffers, P.; Van Bortel, L.M.; Fagard, R.H.; Gardner, J.P.; Kimura, M.; Aviv, A. Telomere length and its associations with oxidized-LDL, carotid artery distensibility and smoking. Front Biosci 2010, 2, 1164–1168. [Google Scholar] [CrossRef][Green Version]
- Needham, B.L.; Adler, N.; Gregorich, S.; Rehkopf, D.; Lin, J.; Blackburn, E.H.; Epel, E.S. Socioeconomic status, health behavior, and leukocyte telomere length in the National Health and Nutrition Examination Survey, 1999–2002. Soc. Sci. Med. 2013, 85, 1–8. [Google Scholar] [CrossRef]
- Patel, C.J.; Manrai, A.K.; Corona, E.; Kohane, I.S. Systematic correlation of environmental exposure and physiological and self-reported behaviour factors with leukocyte telomere length. Int. J. Epidemiol. 2017, 46, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Rode, L.; Bojesen, S.E.; Weischer, M.; Nordestgaard, B.G. High tobacco consumption is causally associated with increased all-cause mortality in a general population sample of 55,568 individuals, but not with short telomeres: A Mendelian randomization study. Int. J. Epidemiol. 2014, 43, 1473–1483. [Google Scholar] [CrossRef] [PubMed]
- Sulastri, D.; Lestari, Y.; Afriwardi; Desmawati. Relationship Between Body Composition and Smoking Habit with Telomere Length of Minangkabau Ethnicity Men, in West Sumatera, Indonesia. Pak. J. Biol. Sci. 2017, 20, 516–522. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Shi, L.; Prescott, J.; Chiuve, S.E.; Hu, F.B.; De Vivo, I.; Stampfer, M.J.; Franks, P.W.; Manson, J.E.; Rexrode, K.M. Healthy lifestyle and leukocyte telomere length in U.S. women. PLoS ONE 2012, 7, e38374. [Google Scholar] [CrossRef] [PubMed]
- Verde, Z.; Reinoso-Barbero, L.; Chicharro, L.; Garatachea, N.; Resano, P.; Sanchez-Hernandez, I.; Rodriguez Gonzalez-Moro, J.M.; Bandres, F.; Santiago, C.; Gomez-Gallego, F. Effects of cigarette smoking and nicotine metabolite ratio on leukocyte telomere length. Environ. Res. 2015, 140, 488–494. [Google Scholar] [CrossRef] [PubMed]
- Wulaningsih, W.; Serrano, F.E.; Utarini, A.; Matsuguchi, T.; Watkins, J.; Network, P.R. Smoking, second-hand smoke exposure and smoking cessation in relation to leukocyte telomere length and mortality. Oncotarget 2016, 7, 60419–60431. [Google Scholar] [CrossRef] [PubMed]
- Yun, M.; Li, S.; Yan, Y.; Zhang, T.; Bazzano, L.; He, J.; Chen, W. Suppression effect of body weight on the association between cigarette smoking and telomere length: The Bogalusa Heart Study. Aging 2019, 11, 9893–9900. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Lauderdale, D.S.; Pierce, B.L. Sex-Specific and Time-Varying Associations between Cigarette Smoking and Telomere Length among Older Adults. Am. J. Epidemiol. 2016, 184, 922–932. [Google Scholar] [CrossRef] [PubMed]
- Bendix, L.; Thinggaard, M.; Fenger, M.; Kolvraa, S.; Avlund, K.; Linneberg, A.; Osler, M. Longitudinal changes in leukocyte telomere length and mortality in humans. J. Gerontol. A Biol. Sci. Med. Sci. 2014, 69, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Kimura, M.; Kim, S.; Cao, X.; Srinivasan, S.R.; Berenson, G.S.; Kark, J.D.; Aviv, A. Longitudinal versus cross-sectional evaluations of leukocyte telomere length dynamics: Age-dependent telomere shortening is the rule. J. Gerontol. A Biol. Sci. Med. Sci. 2011, 66, 312–319. [Google Scholar] [CrossRef] [PubMed]
- Marcon, F.; Siniscalchi, E.; Andreoli, C.; Allione, A.; Fiorito, G.; Medda, E.; Guarrera, S.; Matullo, G.; Crebelli, R. Telomerase activity, telomere length and hTERT DNA methylation in peripheral blood mononuclear cells from monozygotic twins with discordant smoking habits. Environ. Mol. Mutagen. 2017, 58, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Muezzinler, A.; Mons, U.; Dieffenbach, A.K.; Butterbach, K.; Saum, K.U.; Schick, M.; Stammer, H.; Boukamp, P.; Holleczek, B.; Stegmaier, C.; et al. Smoking habits and leukocyte telomere length dynamics among older adults: Results from the ESTHER cohort. Exp. Gerontol. 2015, 70, 18–25. [Google Scholar] [CrossRef]
- Strandberg, T.E.; Saijonmaa, O.; Tilvis, R.S.; Pitkala, K.H.; Strandberg, A.Y.; Miettinen, T.A.; Fyhrquist, F. Association of telomere length in older men with mortality and midlife body mass index and smoking. J. Gerontol. A Biol. Sci. Med. Sci. 2011, 66, 815–820. [Google Scholar] [CrossRef] [PubMed]
- Walters, M.S.; De, B.P.; Salit, J.; Buro-Auriemma, L.J.; Wilson, T.; Rogalski, A.M.; Lief, L.; Hackett, N.R.; Staudt, M.R.; Tilley, A.E.; et al. Smoking accelerates aging of the small airway epithelium. Respir Res. 2014, 15, 94. [Google Scholar] [CrossRef]
- Cribbet, M.R.; Carlisle, M.; Cawthon, R.M.; Uchino, B.N.; Williams, P.G.; Smith, T.W.; Gunn, H.E.; Light, K.C. Cellular aging and restorative processes: Subjective sleep quality and duration moderate the association between age and telomere length in a sample of middle-aged and older adults. Sleep 2014, 37, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Grieshober, L.; Wactawski-Wende, J.; Hageman Blair, R.; Mu, L.; Liu, J.; Nie, J.; Carty, C.L.; Hale, L.; Kroenke, C.H.; LaCroix, A.Z.; et al. A Cross-Sectional Analysis of Telomere Length and Sleep in the Women’s Health Initiative. Am. J. Epidemiol. 2019, 188, 1616–1626. [Google Scholar] [CrossRef]
- Jackowska, M.; Hamer, M.; Carvalho, L.A.; Erusalimsky, J.D.; Butcher, L.; Steptoe, A. Short sleep duration is associated with shorter telomere length in healthy men: Findings from the Whitehall II cohort study. PLoS ONE 2012, 7, e47292. [Google Scholar] [CrossRef]
- Nguyen, M.T.; Lycett, K.; Olds, T.; Matricciani, L.; Vryer, R.; Ranganathan, S.; Burgner, D.; Saffery, R.; Wake, M. Objectively measured sleep and telomere length in a population-based cohort of children and midlife adults. Sleep 2020, 43, zsz200. [Google Scholar] [CrossRef]
- Prather, A.A.; Gurfein, B.; Moran, P.; Daubenmier, J.; Acree, M.; Bacchetti, P.; Sinclair, E.; Lin, J.; Blackburn, E.; Hecht, F.M.; et al. Tired telomeres: Poor global sleep quality, perceived stress, and telomere length in immune cell subsets in obese men and women. Brain Behav. Immun. 2015, 47, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Prather, A.A.; Puterman, E.; Lin, J.; O’Donovan, A.; Krauss, J.; Tomiyama, A.J.; Epel, E.S.; Blackburn, E.H. Shorter leukocyte telomere length in midlife women with poor sleep quality. J. Aging Res. 2011, 2011, 721390. [Google Scholar] [CrossRef]
- Tempaku, P.; Hirotsu, C.; Mazzotti, D.; Xavier, G.; Maurya, P.; Brietzke, E.; Belangero, S.; Poyares, D.; Bittencourt, L.; Tufik, S. Long Sleep Duration, Insomnia, and Insomnia With Short Objective Sleep Duration Are Independently Associated with Short Telomere Length. J. Clin. Sleep Med. 2018, 14, 2037–2045. [Google Scholar] [CrossRef]
- Wynchank, D.; Bijlenga, D.; Penninx, B.W.; Lamers, F.; Beekman, A.T.; Kooij, J.J.S.; Verhoeven, J.E. Delayed sleep-onset and biological age: Late sleep-onset is associated with shorter telomere length. Sleep 2019, 42, zsz139. [Google Scholar] [CrossRef]
- Zgheib, N.K.; Sleiman, F.; Nasreddine, L.; Nasrallah, M.; Nakhoul, N.; Isma’eel, H.; Tamim, H. Short Telomere Length is Associated with Aging, Central Obesity, Poor Sleep and Hypertension in Lebanese Individuals. Aging Dis. 2018, 9, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Han, L.; Chang, D.; Ye, Y.; Shen, J.; Daniel, C.R.; Gu, J.; Chow, W.-H.; Wu, X. Social-Demographics, Health Behaviors, and Telomere Length in the Mexican American Mano a Mano Cohort. Oncotarget 2017, 8, 96553–96567. [Google Scholar] [CrossRef]
- Sella, E.; Miola, L.; Toffalini, E.; Borella, E. The Relationship between Sleep Quality and Quality of Life in Aging: A Systematic Review and Meta-Analysis. Health Psychol. Rev. 2021, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Taillard, J.; Gronfier, C.; Bioulac, S.; Philip, P.; Sagaspe, P. Sleep in Normal Aging, Homeostatic and Circadian Regulation and Vulnerability to Sleep Deprivation. Brain Sci. 2021, 11, 1003. [Google Scholar] [CrossRef]
- Carrier, J.; Semba, K.; Deurveilher, S.; Drogos, L.; Cyr-Cronier, J.; Lord, C.; Sekerovick, Z. Sex Differences in Age-Related Changes in the Sleep-Wake Cycle. Front. Neuroendocrinol. 2017, 47, 66–85. [Google Scholar] [CrossRef]
- Evans, M.A.; Buysse, D.J.; Marsland, A.L.; Wright, A.G.C.; Foust, J.; Carroll, L.W.; Kohli, N.; Mehra, R.; Jasper, A.; Srinivasan, S.; et al. Meta-Analysis of Age and Actigraphy-Assessed Sleep Characteristics across the Lifespan. Sleep 2021, 44, zsab088. [Google Scholar] [CrossRef]
- Kocevska, D.; Lysen, T.S.; Dotinga, A.; Koopman-Verhoeff, M.E.; Luijk, M.P.C.M.; Antypa, N.; Biermasz, N.R.; Blokstra, A.; Brug, J.; Burk, W.J.; et al. Sleep Characteristics across the Lifespan in 1.1 Million People from the Netherlands, United Kingdom and United States: A Systematic Review and Meta-Analysis. Nat. Hum. Behav. 2021, 5, 113–122. [Google Scholar] [CrossRef]
- Buysse, D.J.; Reynolds, C.F.; Monk, T.H.; Berman, S.R.; Kupfer, D.J. The Pittsburgh Sleep Quality Index: A New Instrument for Psychiatric Practice and Research. Psychiatry Res. 1989, 28, 193–213. [Google Scholar] [CrossRef]
- Ye, L.; Elstgeest, L.E.M.; Zhang, X.; Alhambra-Borrás, T.; Tan, S.S.; Raat, H. Factors Associated with Physical, Psychological and Social Frailty among Community-Dwelling Older Persons in Europe: A Cross-Sectional Study of Urban Health Centres Europe (UHCE). BMC Geriatr. 2021, 21, 422. [Google Scholar] [CrossRef]
- Tsai, M.-C.; Yeh, T.-L.; Hsu, H.-Y.; Hsu, L.-Y.; Lee, C.-C.; Tseng, P.-J.; Chien, K.-L. Comparison of Four Healthy Lifestyle Scores for Predicting Cardiovascular Events in a National Cohort Study. Sci. Rep. 2021, 11, 22146. [Google Scholar] [CrossRef] [PubMed]
- Sotos-Prieto, M.; Mattei, J.; Cook, N.R.; Hu, F.B.; Willett, W.C.; Chiuve, S.E.; Rimm, E.B.; Sesso, H.D. Association Between a 20-Year Cardiovascular Disease Risk Score Based on Modifiable Lifestyles and Total and Cause-Specific Mortality Among US Men and Women. J. Am. Heart Assoc. 2018, 7, e010052. [Google Scholar] [CrossRef] [PubMed]
- Parameswaran, G.; Ray, D.W. Sleep, Circadian Rhythms, and Type 2 Diabetes Mellitus. Clin. Endocrinol. 2021, 96, 12–20. [Google Scholar] [CrossRef]
- Lohman, T.; Bains, G.; Berk, L.; Lohman, E. Predictors of Biological Age: The Implications for Wellness and Aging Research. Gerontol. Geriatr. Med. 2021, 7. [Google Scholar] [CrossRef]
- Gruber, H.-J.; Semeraro, M.D.; Renner, W.; Herrmann, M. Telomeres and Age-Related Diseases. Biomedicines 2021, 9, 1335. [Google Scholar] [CrossRef]
- Rostron, B.L.; Chang, C.M.; Pechacek, T.F. Estimation of cigarette smoking-attributable morbidity in the United States. JAMA Intern. Med. 2014, 174, 1922–1928. [Google Scholar] [CrossRef] [PubMed]
- Chilton, W.; O’Brien, B.; Charchar, F. Telomeres, Aging and Exercise: Guilty by Association? Int. J. Mol. Sci. 2017, 18, 2573. [Google Scholar] [CrossRef]
- Von Zglinicki, T.; Pilger, R.; Sitte, N. Accumulation of single-strand breaks is the major cause of telomere shortening in human fibroblasts. Free Radic. Biol. Med. 2000, 28, 64–74. [Google Scholar] [CrossRef]
- Grandner, M.A. Sleep, Health, and Society. Sleep Med. Clin. 2017, 12, 1–22. [Google Scholar] [CrossRef] [PubMed]
- St-Onge, M.P. Sleep-obesity relation: Underlying mechanisms and consequences for treatment. Obes. Rev. 2017, 18 (Suppl. 1), 34–39. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, E.; Yorifuji, T.; Ueshima, K.; Takao, S.; Sugiyama, M.; Ohta, T.; Ishikawa-Takata, K.; Doi, H. Sleep duration, sleep quality and cardiovascular disease mortality among the elderly: A population-based cohort study. Prev. Med. 2009, 49, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Tempaku, P.F.; Mazzotti, D.R.; Tufik, S. Telomere length as a marker of sleep loss and sleep disturbances: A potential link between sleep and cellular senescence. Sleep Med. 2015, 16, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Lotfi, R.A.; El Zawahry, K.M.; Kamar, Z.A.; Hashem, Z. Effects of smoking on human telomerase reverse transcriptase expression in the skin. Int. J. Dermatol. 2014, 53, 1205–1212. [Google Scholar] [CrossRef]
- Gardner, M.; Bann, D.; Wiley, L.; Cooper, R.; Hardy, R.; Nitsch, D.; Martin-Ruiz, C.; Shiels, P.; Sayer, A.A.; Barbieri, M.; et al. Gender and Telomere Length: Systematic Review and Meta-Analysis. Exp. Gerontol. 2014, 51, 15–27. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barragán, R.; Ortega-Azorín, C.; Sorlí, J.V.; Asensio, E.M.; Coltell, O.; St-Onge, M.-P.; Portolés, O.; Corella, D. Effect of Physical Activity, Smoking, and Sleep on Telomere Length: A Systematic Review of Observational and Intervention Studies. J. Clin. Med. 2022, 11, 76. https://doi.org/10.3390/jcm11010076
Barragán R, Ortega-Azorín C, Sorlí JV, Asensio EM, Coltell O, St-Onge M-P, Portolés O, Corella D. Effect of Physical Activity, Smoking, and Sleep on Telomere Length: A Systematic Review of Observational and Intervention Studies. Journal of Clinical Medicine. 2022; 11(1):76. https://doi.org/10.3390/jcm11010076
Chicago/Turabian StyleBarragán, Rocío, Carolina Ortega-Azorín, Jose V. Sorlí, Eva M. Asensio, Oscar Coltell, Marie-Pierre St-Onge, Olga Portolés, and Dolores Corella. 2022. "Effect of Physical Activity, Smoking, and Sleep on Telomere Length: A Systematic Review of Observational and Intervention Studies" Journal of Clinical Medicine 11, no. 1: 76. https://doi.org/10.3390/jcm11010076
APA StyleBarragán, R., Ortega-Azorín, C., Sorlí, J. V., Asensio, E. M., Coltell, O., St-Onge, M.-P., Portolés, O., & Corella, D. (2022). Effect of Physical Activity, Smoking, and Sleep on Telomere Length: A Systematic Review of Observational and Intervention Studies. Journal of Clinical Medicine, 11(1), 76. https://doi.org/10.3390/jcm11010076