
Metabolomic Alteration of Oral Keratinocytes and Fibroblasts in Hypoxia

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Primary Cell Culture under Hypoxic Conditions
2.2. Metabolite Extraction
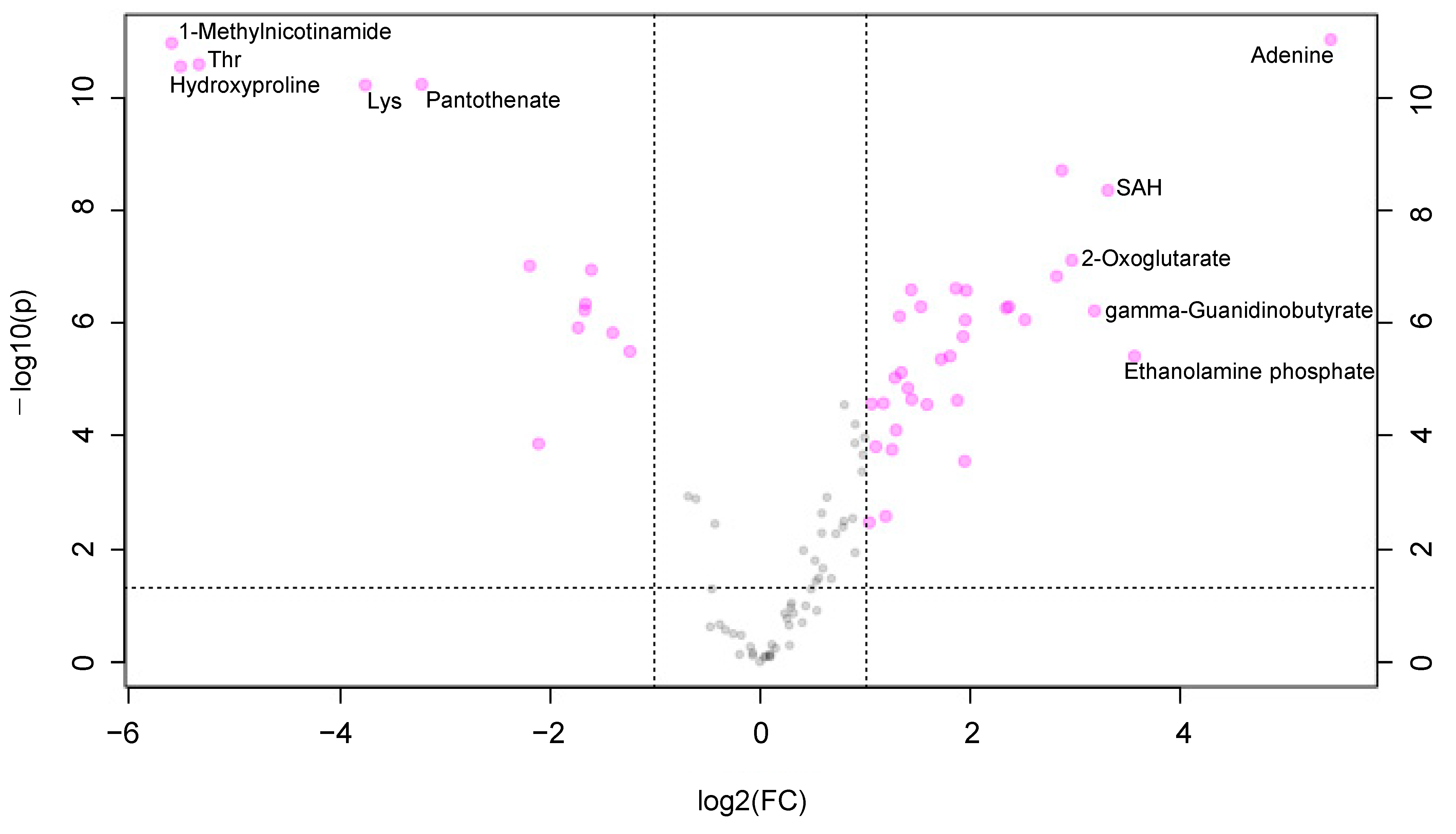
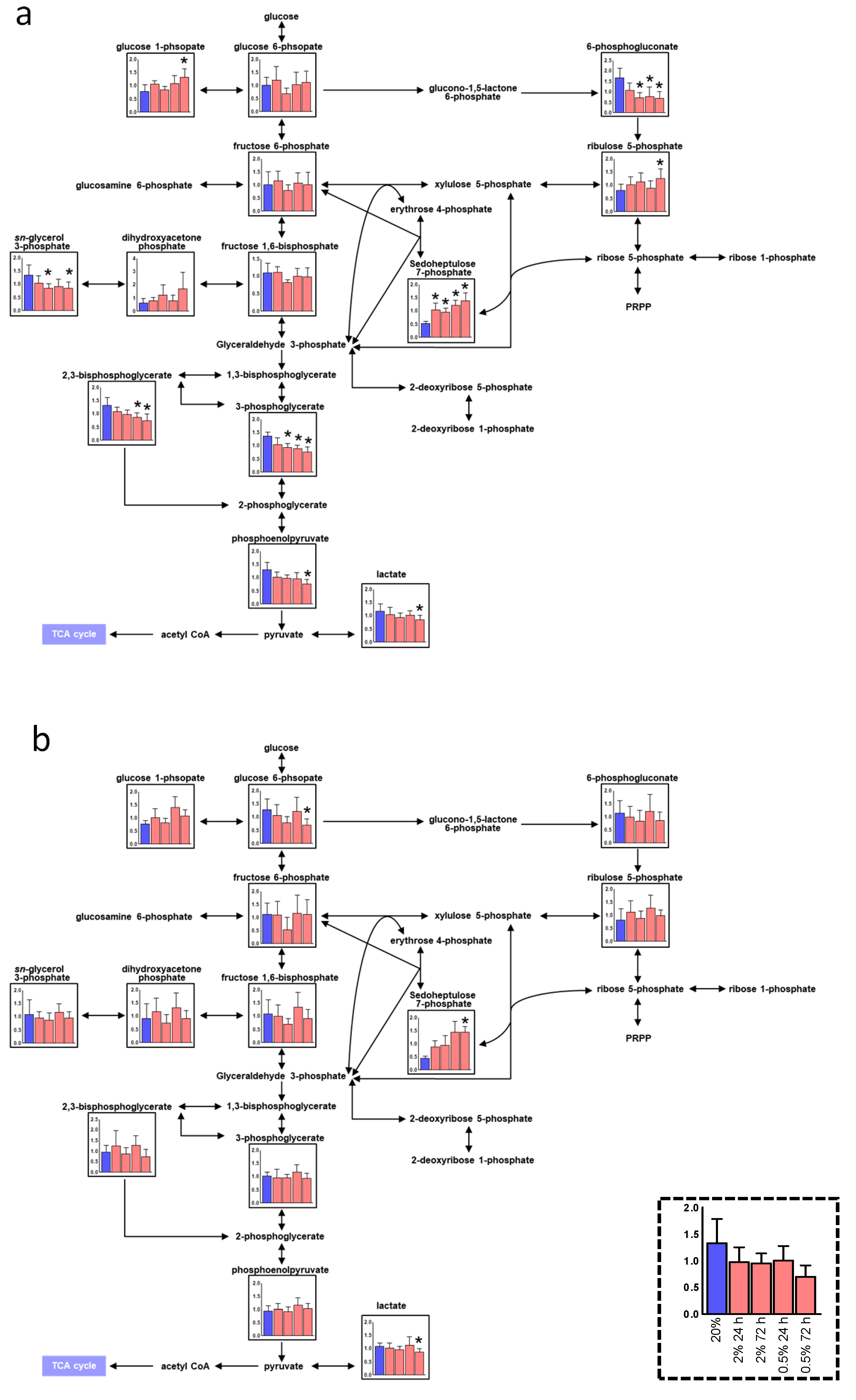
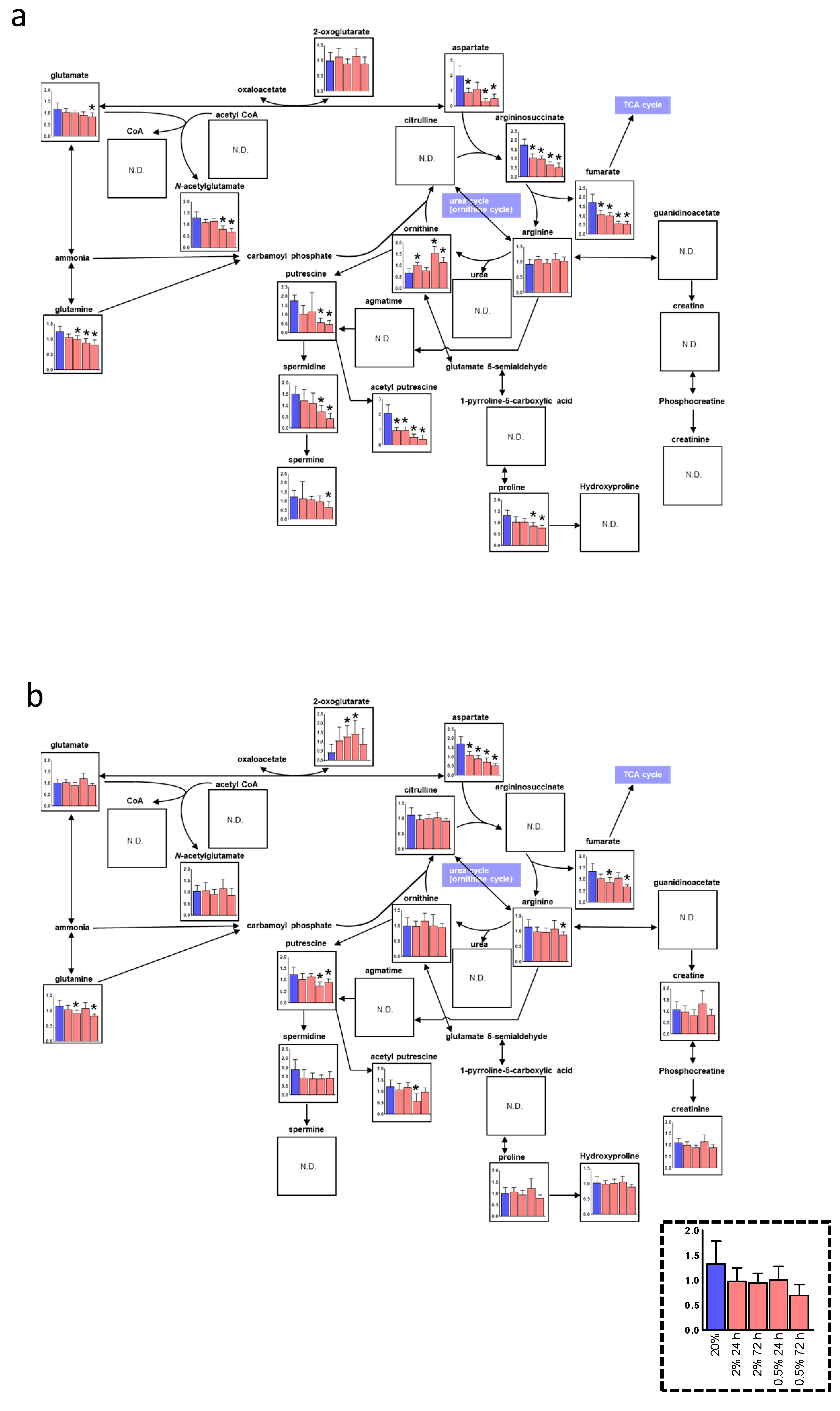
3. Results
3.1. The Metabolomic Concentration of Oral Keratinocytes and Oral Fibroblasts
3.2. Alteration of the Pentose Phosphate Pathway (PPP) and Urea Cycle under Hypoxia
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mohyeldin, A.; Garzón-Muvdi, T.; Quiñones-Hinojosa, A. Oxygen in Stem Cell Biology: A Critical Component of the Stem Cell Niche. Cell Stem Cell 2010, 7, 150–161. [Google Scholar] [CrossRef]
- Evans, S.M.; Schrlau, A.E.; Chalian, A.A.; Zhang, P.; Koch, C.J. Oxygen levels in normal and previously irradiated human skin as as-sessed by EF5 binding. J. Investig. Dermatol. 2006, 126, 2596–2606. [Google Scholar] [CrossRef]
- Rezvani, H.R.; Ali, N.; Serrano-Sanchez, M.; Dubus, P.; Varon, C.; Ged, C.; Pain, C.; Cario-André, M.; Seneschal, J.; Taïeb, A.; et al. Loss of epidermal hypoxia-inducible factor-1α accelerates epidermal aging and affects re-epithelialization in human and mouse. J. Cell Sci. 2011, 124, 4172–4183. [Google Scholar] [CrossRef]
- Ngo, M.A.; Sinitsyna, N.N.; Qin, Q.; Rice, R.H. Oxygen-Dependent Differentiation of Human Keratinocytes. J. Investig. Dermatol. 2007, 127, 354–361. [Google Scholar] [CrossRef]
- Steinbrech, D.S.; Longaker, M.T.; Mehrara, B.J.; Saadeh, P.B.; Chin, G.S.; Gerrets, R.P.; Chau, D.C.; Rowe, N.M.; Gittes, G.K. Fibroblast response to hypoxia: The relationship between angio-genesis and matrix regulation. J. Surg. Res. 1999, 84, 127–133. [Google Scholar] [CrossRef]
- Yoshida, Y.; Takahashi, K.; Okita, K.; Ichisaka, T.; Yamanaka, S. Hypoxia Enhances the Generation of Induced Pluripotent Stem Cells. Cell Stem Cell 2009, 5, 237–241. [Google Scholar] [CrossRef]
- Santilli, G.; Lamorte, G.; Carlessi, L.; Ferrari, D.; Nodari, L.R.; Binda, E.; Delia, D.; Vescovi, A.L.; De Filippis, L. Mild Hypoxia Enhances Proliferation and Multipotency of Human Neural Stem Cells. PLoS ONE 2010, 5, e8575. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Fujita, M.; Tanaka, Y.; Kojima, I.; Kanatani, Y.; Ishihara, M.; Tachibana, S. Low oxygen tension enhances proliferation and maintains stemness of ad-ipose tissue-derived stromal cells. Biores. Open Access 2013, 2, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Goda, N.; Kanai, M. Hypoxia-inducible factors and their roles in energy metabolism. Int. J. Hematol. 2012, 95, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Altman, B.J.; Stine, Z.E.; Dang, B.J.A.Z.E.S.C.V. From Krebs to clinic: Glutamine metabolism to cancer therapy. Nat. Rev. Cancer 2016, 16, 619–634. [Google Scholar] [CrossRef]
- Izumi, K.; Kato, H.; Feinberg, S.E. Three-dimensional reconstruction of oral mucosa. In Tissue Engineering Strategies. Stem Cell Biology and Tissue Engineering in Dental Science; Vishwakarma, A., Sharpe, P., Shi, S., Wang, X.-P., Ramalingam, M., Eds.; Academic Press/Elsevier: Waltham, MA, USA, 2015; Chapter 53; pp. 721–731. [Google Scholar]
- Thorn, J.J.; Kallehave, F.; Westergaard, P.; Hansen, E.H.; Gottrup, F. The effect of hyperbaric oxygen on irradiated oral tissues: Transmuco-sal oxygen tension measurements. J. Oral Maxillofac. Surg. 1997, 55, 1103–1107. [Google Scholar] [CrossRef]
- Chen, L.; Gajendrareddy, P.K.; DiPietro, L.A. Differential Expression of HIF-1α in Skin and Mucosal Wounds. J. Dent. Res. 2012, 91, 871–876. [Google Scholar] [CrossRef]
- Kato, H.; Izumi, K.; Uenoyama, A.; Shiomi, A.; Kuo, S.; Feinberg, S.E. Hypoxia induces an undifferentiated phenotype of oral keratinocytes in vitro. Cells Tissues Organs 2014, 199, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Kato, H.; Izumi, K.; Saito, T.; Ohnuki, H.; Terada, M.; Kawano, Y.; Nozawa-Inoue, K.; Saito, C.; Maeda, T. Distinct expression patterns and roles of aldehyde dehydrogenases in normal oral mucosa keratinocytes: Differential inhibitory effects of a pharmacological inhibitor and RNAi-mediated knockdown on cellu-lar phenotype and epithelial morphology. Histochem. Cell Biol. 2013, 139, 847–862. [Google Scholar] [CrossRef] [PubMed]
- Uenoyama, A.; Kakizaki, I.; Shiomi, A.; Saito, N.; Hara, Y.; Saito, T.; Ohnuki, H.; Kato, H.; Takagi, R.; Maeda, T.; et al. Effects of C-xylopyranoside derivative on epithelial regeneration in an in vitro 3D oral mucosa model. Biosci. Biotechnol. Biochem. 2016, 80, 1344–1355. [Google Scholar] [CrossRef][Green Version]
- Kato, H.; Marcelo, C.L.; Washington, J.B.; Bingham, E.L.; Feinberg, S.E. Fabrication of Large Size Ex Vivo-Produced Oral Mucosal Equivalents for Clinical Application. Tissue Eng. Part C Methods 2015, 21, 872–880. [Google Scholar] [CrossRef] [PubMed]
- Sakagami, H.; Sugimoto, M.; Kanda, Y.; Murakami, Y.; Amano, O.; Saitoh, J.; Kochi, A. Changes in Metabolic Profiles of Human Oral Cells by Benzylidene Ascorbates and Eugenol. Medicines 2018, 5, 116. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, M.; Sakagami, H.; Yokote, Y.; Onuma, H.; Kaneko, M.; Mori, M.; Sakaguchi, Y.; Soga, T.; Tomita, M. Non-targeted metabolite profiling in activated macrophage secretion. Metabolomics 2011, 8, 624–633. [Google Scholar] [CrossRef]
- Sugimoto, M.; Wong, D.T.; Hirayama, A.; Soga, T.; Tomita, M. Capillary electrophoresis mass spectrometry-based saliva metabolomics identified oral, breast and pancreatic cancer-specific profiles. Metabolomics 2009, 6, 78–95. [Google Scholar] [CrossRef]
- Sugimoto, M. Metabolomic pathway visualization tool outsourcing editing function. In Proceedings of the 2015 37th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), Milan, Italy, 25–29 August 2015; Volume 2015, pp. 7659–7662. [Google Scholar]
- Kato, H.; Niigata University, Niigata, Japan. Unpublished work. 2021.
- Semenza, G.L. Hypoxia-inducible factors: Coupling glucose metabolism and redox regulation with induction of the breast cancer stem cell phenotype. EMBO J. 2017, 36, 252–259. [Google Scholar] [CrossRef]
- Meléndez-Rodríguez, F.; Urrutia, A.A.; Lorendeau, D.; Rinaldi, G.; Roche, O.; Böğürcü-Seidel, N.; Muelas, M.O.; Mesa-Ciller, C.; Turiel, G.; Bouthelier, A.; et al. HIF1α Suppresses Tumor Cell Proliferation through Inhi-bition of Aspartate Biosynthesis. Cell Rep. 2019, 26, 2257–2265. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.P.; Fishel, R.S.; Efron, D.T.; Williams, J.Z.; Fishel, M.H.; Barbul, A. Effect of Supplemental Ornithine on Wound Healing. J. Surg. Res. 2002, 106, 299–302. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, M.; Sricholpech, M. Lysine post-translational modifications of collagen. Essays Biochem. 2012, 52, 113–133. [Google Scholar] [CrossRef]
- Roseeuw, D.I.; Marcelo, C.L.; Rhodes, L.M.; Voorhees, J.J. Epidermal keratinocytes actively maintain their intracellular polyamine levels. Cell Tissue Kinet. 1983, 16, 493–504. [Google Scholar] [PubMed]
- Kuehne, A.; Emmert, H.; Soehle, J.; Winnefeld, M.; Fischer, F.; Wenck, H.; Gallinat, S.; Terstegen, L.; Lucius, R.; Hildebrand, J.; et al. Acute Activation of Oxidative Pentose Phosphate Pathway as First-Line Response to Oxidative Stress in Human Skin Cells. Mol. Cell 2015, 59, 359–371. [Google Scholar] [CrossRef] [PubMed]
- Takubo, K.; Nagamatsu, G.; Kobayashi, C.I.; Nakamura-Ishizu, A.; Kobayashi, H.; Ikeda, E.; Goda, N.; Rahimi, Y.; Johnson, R.S.; Soga, T.; et al. Regulation of Glycolysis by Pdk Functions as a Metabolic Checkpoint for Cell Cycle Quiescence in Hematopoietic Stem Cells. Cell Stem Cell 2013, 12, 49–61. [Google Scholar] [CrossRef]
- Hawkins, K.E.; Joy, S.; Delhove, J.M.; Kotiadis, V.N.; Fernandez, E.; Fitzpatrick, L.M.; Whiteford, J.R.; King, P.J.; Bolanos, J.P.; Duchen, M.R.; et al. NRF2 Orchestrates the Metabolic Shift during Induced Pluripotent Stem Cell Reprogramming. Cell Rep. 2016, 14, 1883–1891. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Yu, S.-S.; Zong, M.; Fan, S.-S.; Lu, T.-B.; Gong, R.-H.; Sun, L.-S.; Fan, L.-Y. Glucose-6-Phosphate Isomerase (G6PI) Mediates Hypoxia-Induced Angiogenesis in Rheumatoid Arthritis. Sci. Rep. 2017, 7, 40274. [Google Scholar] [CrossRef] [PubMed]
- Pavlides, S.; Whitaker-Menezes, D.; Castello-Cros, R.; Flomenberg, N.; Witkiewicz, A.K.; Frank, P.G.; Casimiro, M.C.; Wang, C.; Fortina, P.; Addya, S.; et al. The reverse Warburg effect: Aerobic glycolysis in cancer associated fibroblasts and the tumor stroma. Cell Cycle 2009, 8, 3984–4001. [Google Scholar] [CrossRef]
- Alfarouk, K.O.; Ahmed, S.; Elliott, R.L.; Benoit, A.; Alqahtani, S.S.; Ibrahim, M.E.; Bashir, A.H.H.; Alhoufie, S.T.S.; Elhassan, G.O.; Wales, C.C.; et al. The pentose phosphate pathway dynamics in cancer and its de-pendency on intracellular pH. Metabolites 2020, 10, 285. [Google Scholar] [CrossRef]
- Sørensen, B.S.; Busk, M.; Overgaard, J.; Horsman, M.R.; Alsner, J. Simultaneous hypoxia and low extracellular pH suppress overall metabol-ic rate and protein synthesis in vitro. PLoS ONE 2015, 10, 1–14. [Google Scholar] [CrossRef]
- Hamanaka, R.B.; Mutlu, G.M. PFKFB3, a Direct Target of p63, Is Required for Proliferation and Inhibits Differentiation in Epidermal Keratinocytes. J. Investig. Dermatol. 2017, 137, 1267–1276. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Xu, Q.; Wan, H.; Hu, Y.; Xing, S.; Yang, H.; Gao, Y.; He, Z. PI3K-Akt-mTOR/PFKFB3 pathway mediated lung fibroblast aerobic glycolysis and col-lagen synthesis in lipopolysaccharide-induced pulmonary fibrosis. Lab. Investig. 2020, 100, 801–811. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, L.B.; Luengo, A.; Danai, L.V.; Bush, L.N.; Diehl, F.F.; Hosios, A.M.; Lau, A.N.; Elmiligy, S.; Malstrom, S.; Lewis, C.A.; et al. Aspartate is an endogenous metabolic limitation for tumour growth. Nat. Cell Biol. 2018, 20, 782–788. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Bermudez, J.; Baudrier, L.; La, K.; Zhu, X.G.; Fidelin, J.; Sviderskiy, V.O.; Papagiannakopoulos, T.; Molina, H.; Snuderl, M.; Lewis, C.A.; et al. Aspartate is a limiting metabolite for cancer cell proliferation under hy-poxia and in tumours. Nat. Cell Biol. 2018, 20, 775–781. [Google Scholar] [CrossRef]
- Lee, P.; Chandel, N.S.; Simon, M.C. Cellular adaptation to hypoxia through hypoxia inducible factors and beyond. Nat. Rev. Mol. Cell Biol. 2020, 21, 268–283. [Google Scholar] [CrossRef]
- Pietilä, M.; Pirinen, E.; Keskitalo, S.; Juutinen, S.; Pasonen-Seppänen, S.; Keinänen, T.; Alhonen, L.; Jänne, J. Disturbed keratinocyte differentiation in transgenic mice and organotypic keratinocyte cultures as a result of spermidme/spermine N1-acetyltransferase overexpression. J. Investig. Dermatol. 2005, 124, 596–601. [Google Scholar] [CrossRef]
- Zdzisińska, B.; Żurek, A.; Kandefer-Szerszeń, M. Alpha-Ketoglutarate as a Molecule with Pleiotropic Activity: Well-Known and Novel Possibilities of Therapeutic Use. Arch. Immunol. Ther. Exp. 2017, 65, 21–36. [Google Scholar] [CrossRef]
- Lee, W.D.; Mukha, D.; Aizenshtein, E.; Shlomi, T. Spatial-fluxomics provides a subcellular-compartmentalized view of reductive glutamine metabolism in cancer cells. Nat. Commun. 2019, 10, 1–14. [Google Scholar] [CrossRef]
- Wang, Y.; Bai, C.; Ruan, Y.; Liu, M.; Chu, Q.; Qiu, L.; Yang, C.; Li, B. Coordinative metabolism of glutamine carbon and nitrogen in proliferating cancer cells under hypoxia. Nat. Commun. 2019, 10, 1–14. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kato, H.; Sugimoto, M.; Enomoto, A.; Kaneko, M.; Hara, Y.; Saito, N.; Shiomi, A.; Ohnuki, H.; Izumi, K. Metabolomic Alteration of Oral Keratinocytes and Fibroblasts in Hypoxia. J. Clin. Med. 2021, 10, 1156. https://doi.org/10.3390/jcm10061156
Kato H, Sugimoto M, Enomoto A, Kaneko M, Hara Y, Saito N, Shiomi A, Ohnuki H, Izumi K. Metabolomic Alteration of Oral Keratinocytes and Fibroblasts in Hypoxia. Journal of Clinical Medicine. 2021; 10(6):1156. https://doi.org/10.3390/jcm10061156
Chicago/Turabian StyleKato, Hiroko, Masahiro Sugimoto, Ayame Enomoto, Miku Kaneko, Yuko Hara, Naoaki Saito, Aki Shiomi, Hisashi Ohnuki, and Kenji Izumi. 2021. "Metabolomic Alteration of Oral Keratinocytes and Fibroblasts in Hypoxia" Journal of Clinical Medicine 10, no. 6: 1156. https://doi.org/10.3390/jcm10061156
APA StyleKato, H., Sugimoto, M., Enomoto, A., Kaneko, M., Hara, Y., Saito, N., Shiomi, A., Ohnuki, H., & Izumi, K. (2021). Metabolomic Alteration of Oral Keratinocytes and Fibroblasts in Hypoxia. Journal of Clinical Medicine, 10(6), 1156. https://doi.org/10.3390/jcm10061156