
Exploring Nanofiltration for Transport of Small Molecular Species for Application in Artificial Kidney Devices to Treat End-Stage Kidney Disease




Abstract
1. Introduction
2. Materials and Methods
2.1. Commercial NF Membranes and Their Characteristics
2.2. Feed Conditions of Commercial NF Membrane Experiments
2.3. Bench-Scale Unmodified Commercial Membrane Experimental Set-Up and Procedure
2.4. Chemical Analysis
2.5. Rejection
2.6. Statistical Analysis
3. Results and Discussion
3.1. Permeate Flux of NF Commercial Membranes Based on Feed Conditions
3.2. Urea Rejection of Commercial NF Membranes at Varying Feed Conditions
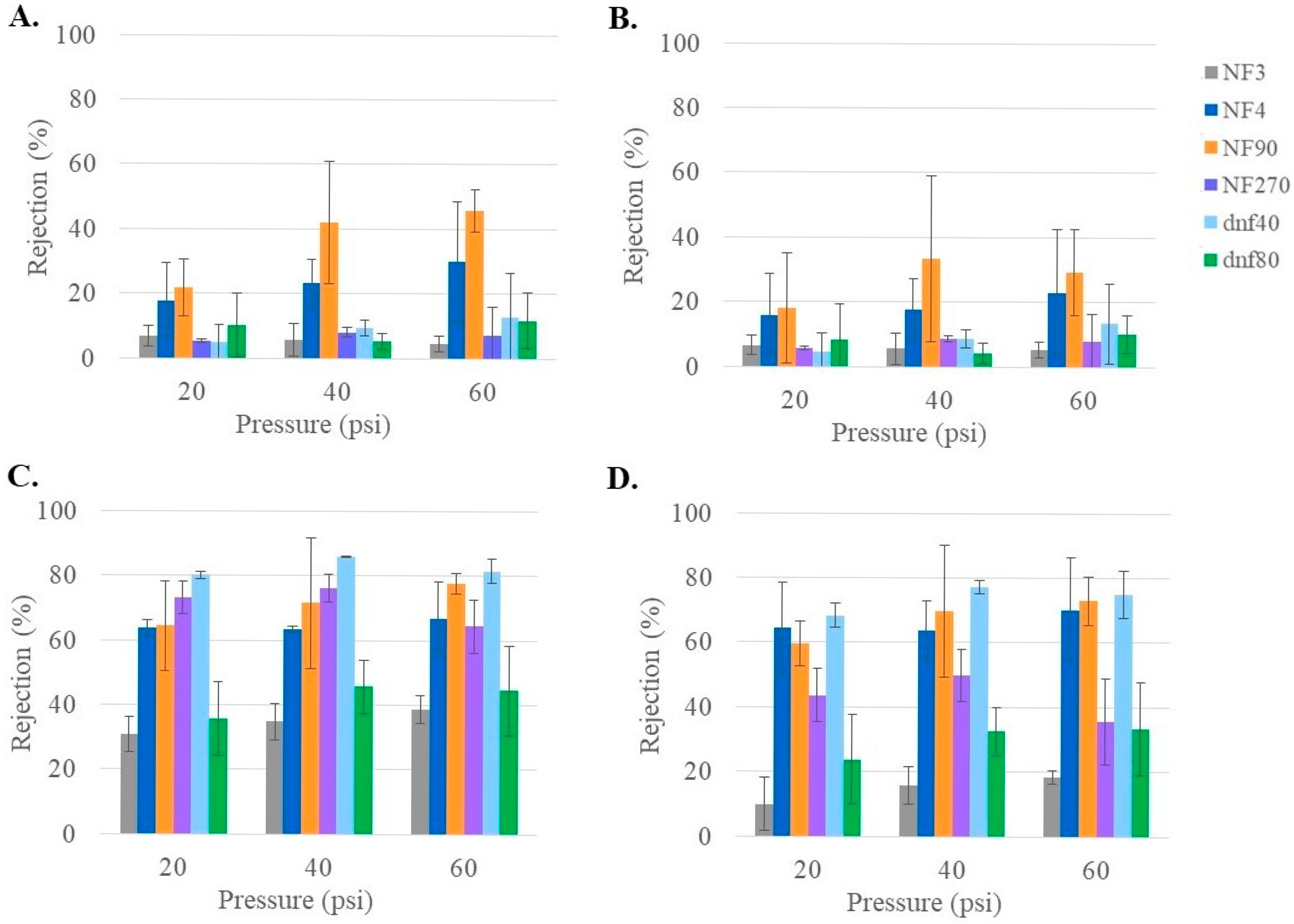
3.3. Glucose Rejection of Commercial NF Membranes at Varying Feed Conditions
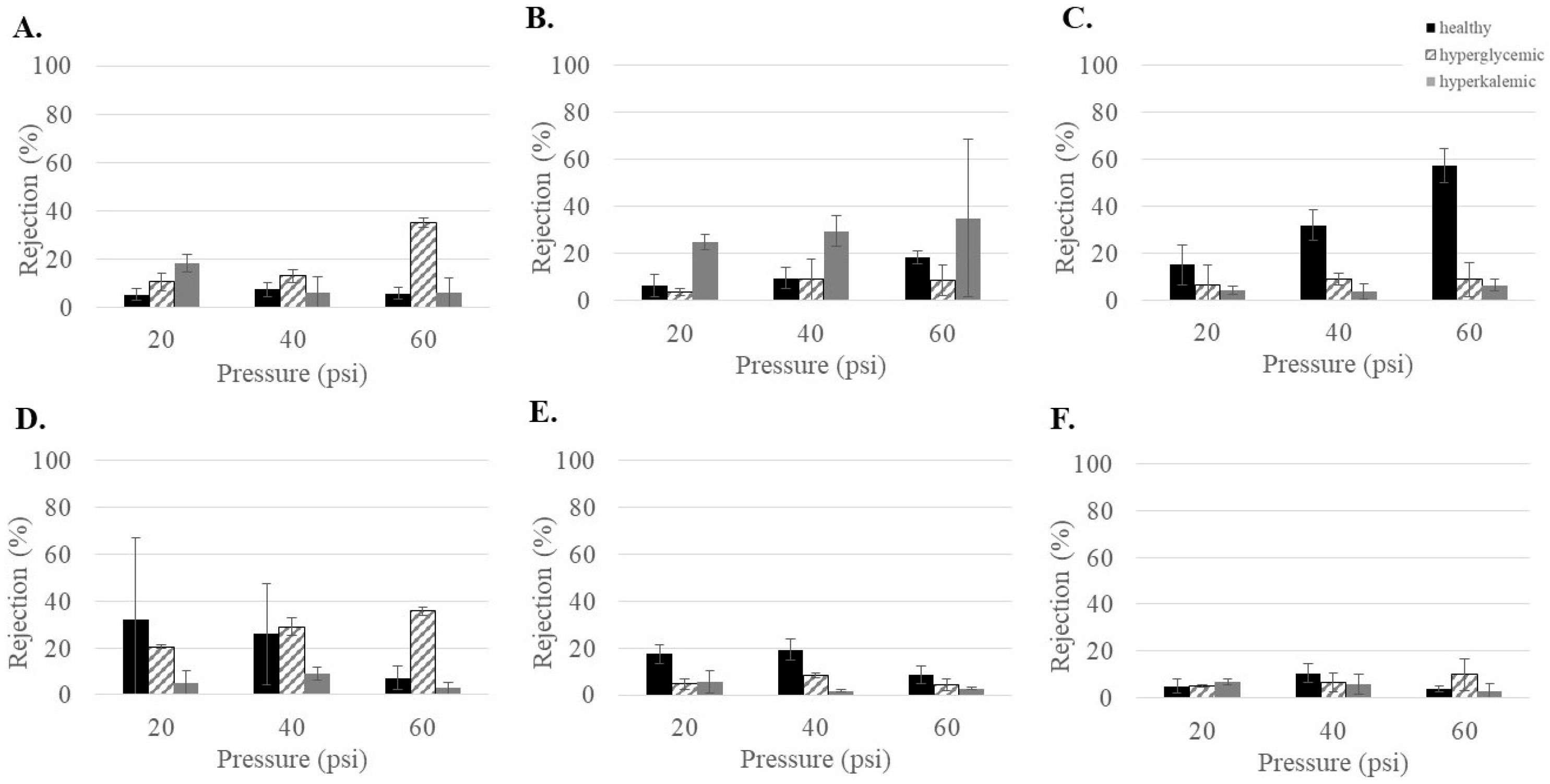
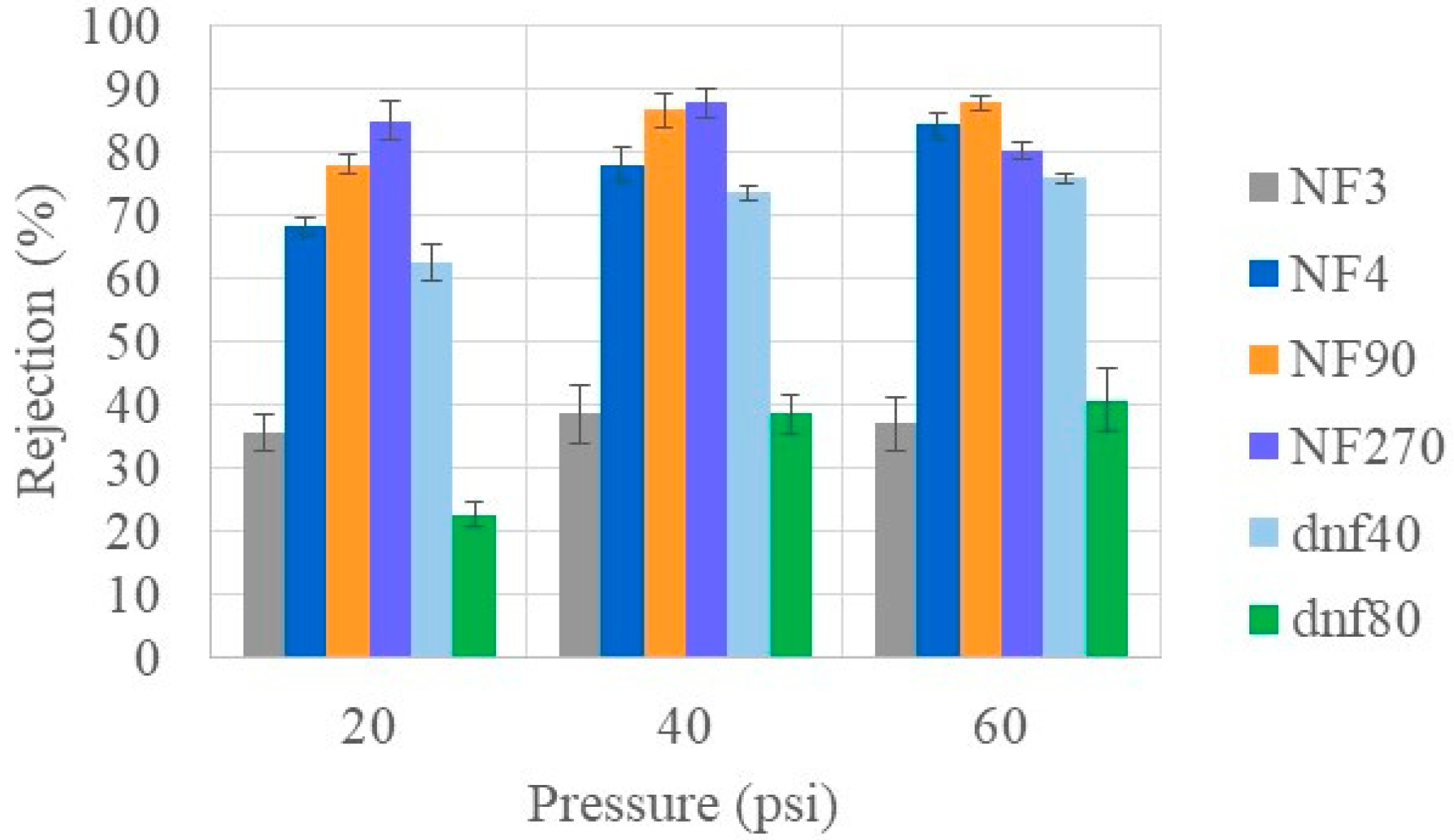
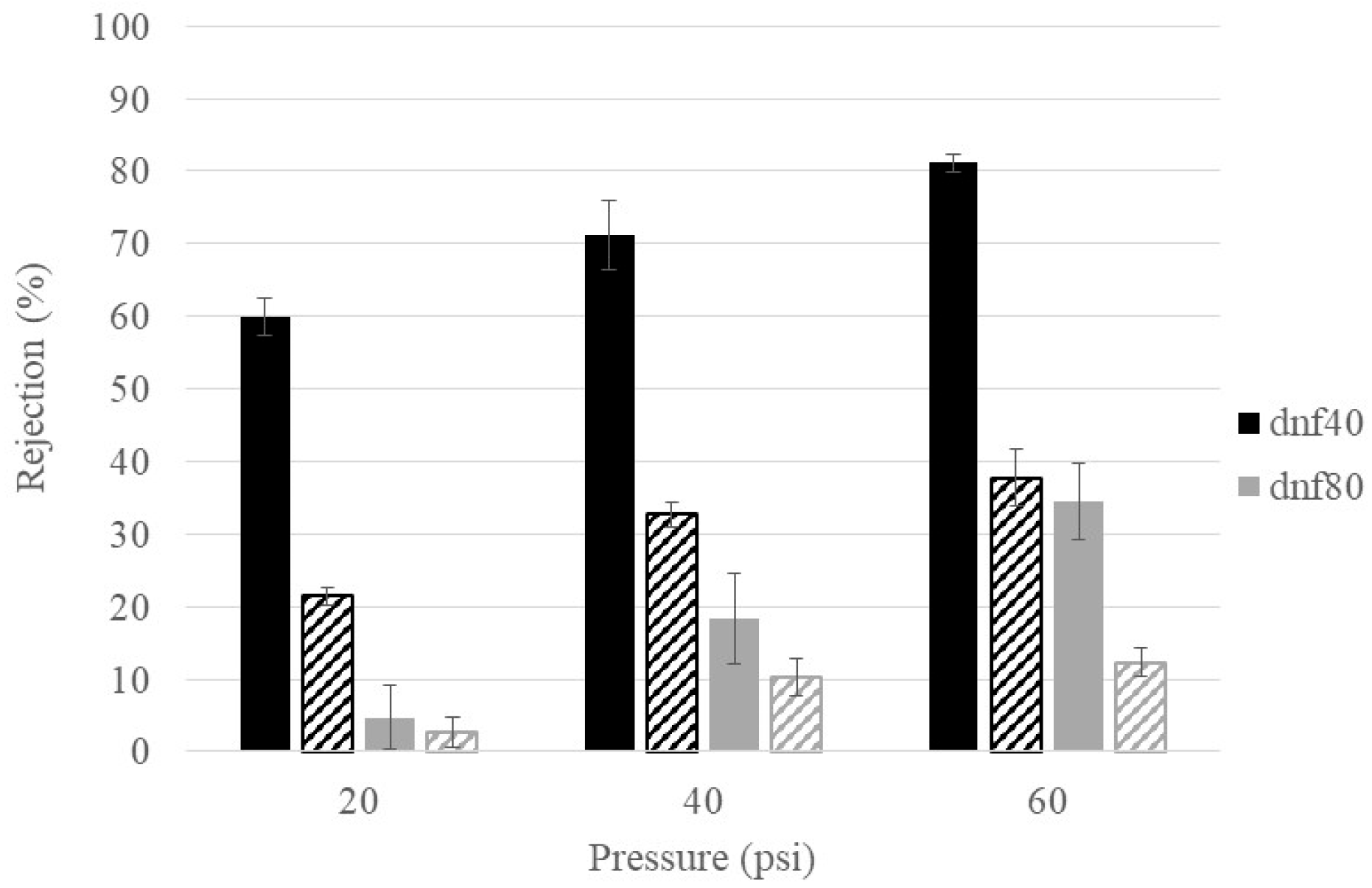
3.4. Ion Rejections of Commercial NF Membranes at Varying Feed Conditions
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ESRD | End-stage renal disease |
| MWCO | Molecular weight cutoff |
| NF | Nanofiltration |
| TFC | thin-film composite |
References
- Quick Kidney Disease Facts and Stats. Available online: www.kidneyfund.org/all-about-kidneys/quick-kidney-disease-facts-and-stats#:~:text=CKD%20in%20the%20United%20States,557%2C000%20Americans%20are%20on%20dialysis (accessed on 7 June 2024).
- Reutens, A.T.; Atkins, R.C. Chromic kidney disease (CKD): The scope of the global problem. In Chronic Kidney Disease: A Practical Guide to Understanding and Management, 1st ed.; El Nahas, M., Levin, A., Eds.; Oxford University Press: Oxford, UK, 2009; pp. 39–76. [Google Scholar]
- Pereira, B.J.G.; Sayegh, M.H.; Blake, P. Chronic Kidney Disease, Dialysis, and Transplantation: A Companion to Brenner and Rector’s the Kidney, 2nd ed.; Saunders: Philadelphia, PA, USA, 2005. [Google Scholar]
- Chronic Kidney Disease: Common, Serious, and Costly. Available online: www.cdc.gov/kidney-disease/ckd-facts/index.html (accessed on 8 August 2024).
- Demystifying Dialysis: Understanding Treatment Options and Life Changes. Available online: www.kidney.org/newsletter/demystifying-dialysis-understanding-treatment-options-and-life-changes#:~:text=The%20average%20life%20expectancy%20for,20%20or%20even%2030%20years (accessed on 7 June 2024).
- Himmelfarb, J.; Vanholder, R.; Mehrotra, R.; Tonelli, M. The current and future landscape of dialysis. Nat. Rev. Nephrol. 2020, 16, 573–585. [Google Scholar] [CrossRef] [PubMed]
- Walker, S.; Beckett, G.; Rae, P.; Ashby, P. Clinical Biochemistry, 9th ed.; John Wiley & Sons, Incorporated: West Sussex, UK, 2013. [Google Scholar]
- Shao, G.; Himmelfarb, J.; Hinds, B. Strategies for optimizing urea removal to enable portable kidney dialysis: A reappraisal. Artif. Organs 2022, 46, 997–1011. [Google Scholar] [CrossRef] [PubMed]
- Uragami, T. Comparison of pressure-driven membrane separation processes. In Science and Technology of Separation Membranes; John Wiley & Sons, Ltd.: West Sussex, UK, 2013; pp. 379–383. [Google Scholar] [CrossRef]
- Kim, J.C.; Garzotto, F.; Nalesso, F.; Cruz, D.; Kim, J.H.; Kang, E.; Kim, H.C.; Ronco, C. A wearable artificial kidney: Technical requirements and potential solutions. Expert Rev. Med. Devices 2011, 8, 567–579. [Google Scholar] [CrossRef]
- Ghezzi, P.M.; Canepari, G.; Ronco, C. Behaviour of clearances and diffusive permeability during hemodialysis with PMMA dialyzers: Clinical study. In Polymethylmethacrylate: A Flexible Membrane for a Tailored Dialysis; Berlyne, G.M., Ronco, C., Eds.; S. Karger: Basel, Switzerland, 1998; pp. 53–64. [Google Scholar] [CrossRef]
- Yoon, Y.; Lueptow, R.M. Removal of organic contaminants by RO and NF membranes. J. Membr. Sci. 2005, 261, 76–86. [Google Scholar] [CrossRef]
- Hestekin, C.N.; Hestekin, J.A.; Paracha, S.; Morrison, G.; Pakkaner, E.; Moore, J.; Santos de Souza, L.; Stephen, S.; Atchley, C.; Kurtz, I. Simulating nephron ion transport function using activated wafer electrodeionization. Commun. Mater. 2020, 1, 20. [Google Scholar] [CrossRef]
- Eriksson, P. Nanofiltration extends the range of membrane filtration. Environ. Prog. Sustain. 1988, 7, 58–62. [Google Scholar] [CrossRef]
- Van der Bruggen, B.; Schaep, J.; Wilms, D.; Vandecasteele, C. Influence of molecular size, polarity and charge on the retention of organic molecules by nanofiltration. J. Membr. Sci. 1998, 156, 29–41. [Google Scholar] [CrossRef]
- Kiso, Y.; Sugiura, Y.; Kitao, T.; Nishimura, K. Effects of hydrophobicity and molecular size on rejection of aromatic pesticides with nanofiltration membranes. J. Membr. Sci. 2001, 192, 1–10. [Google Scholar] [CrossRef]
- Xu, Y.; Lebrun, R.E. Investigation of the solute separation by charged nanofiltration membrane: Effect of pH, ionic strength and solute type. J. Membr. Sci. 1999, 158, 93–104. [Google Scholar] [CrossRef]
- Teixeira, M.R.; Rosa, M.J.; Nystrom, M. The role of membrane charge on nanofiltration performance. J. Membr. Sci. 2005, 265, 160–166. [Google Scholar] [CrossRef]
- Childress, A.E.; Elimelech, M. Relating nanofiltration membrane performance to membrane charge (electrokinetic) characteristics. Environ. Sci. Technol. 2000, 34, 3710–3716. [Google Scholar] [CrossRef]
- Mohammad, A.W.; Teow, Y.H.; Ang, W.L.; Chung, Y.T.; Oatley-Radcliffe, D.L.; Hilal, N. Nanofiltration membranes review: Recent advances and future prospects. Desalination 2013, 356, 226–254. [Google Scholar] [CrossRef]
- Musbah, I.; Ciceron, D.; Saboni, A.; Alexandrova, S. Retention of pesticides and metabolites by nanofiltration by effects of size and dipole moment. Desalination 2013, 313, 51–56. [Google Scholar] [CrossRef]
- Berg, P.; Hagmeyer, G.; Gimbel, R. Removal of pesticides and other micropollutants by nanofiltration. Desalination 1997, 113, 205–208. [Google Scholar] [CrossRef]
- Vergili, I. Application of nanofiltration for the removal of carbamazepine, diclofenac and ibuprofen from drinking water sources. J. Environ. Manag. 2013, 127, 177–187. [Google Scholar] [CrossRef]
- Salehi, F. Current and future applications for nanofiltration technology in the Food Processing. Food Bioprod. Process. 2013, 92, 161–177. [Google Scholar] [CrossRef]
- Umpierrez, G.E.; Davis, G.M.; ElSayed, N.A.; Fadini, G.P.; Galindo, R.J.; Hirsch, I.B.; Klonoff, D.C.; McCoy, R.G.; Misra, S.; Gabbay, R.A.; et al. Hyperglycemic Crises in Adults With Diabetes: A Consensus Report. Diabetes Care 2024, 47, 1257–1275. [Google Scholar] [CrossRef]
- Gupta, N.; Prasad, P. Hyperkalemia in diabetes: Newer insights into mechanism and treatment. Postgrad. Med. J. 2022, 98, e45. [Google Scholar] [CrossRef]
- Lopez-Munoz, M.J.; Sotto, A.; Arsuaga, J.M.; Van der Bruggen, B. Influence of membrane, solute, and solution properties on the retention of phenolic compounds in aqueous solution by nanofiltration membranes. Sep. Purif. Technol. 2009, 66, 194–201. [Google Scholar] [CrossRef]
- Van der Bruggen, B.; Vandecasteele, C. Flux decline during nanofiltration of organic components in aqueous solution. Environ. Sci. Technol. 2001, 35, 3535–3540. [Google Scholar] [CrossRef]
- Wang, R.; Li, Y.; Wang, J.; You, G.; Cai, C.; Chen, B.H. Modeling the permeate flux and rejection of nanofiltration membrane separation with high concentration uncharged aqueous solutions. Desalination 2012, 299, 44–49. [Google Scholar] [CrossRef]
- Roy, Y.; Warsinger, D.M.; Lienhard, J.H. Effect of temperature on ion transport in nanofiltration membranes: Diffusion, convection and electromigration. Desalination 2017, 420, 241–257. [Google Scholar] [CrossRef]
- Tsuru, T.; Izumi, S.; Yoshioka, T.; Asaeda, M. Temperature effect on transport performance by inorganic nanofiltration membranes. AIChE J. 2000, 46, 565–574. [Google Scholar] [CrossRef]
- Ray, H.; Perreault, F.; Boyer, T.H. Rejection of nitrogen species in real fresh and hydrolyzed human urine by reverse osmosis and nanofiltration. J. Environ. Chem. Eng. 2020, 8, 103993. [Google Scholar] [CrossRef]
- Courtney, C.; Randall, D.G. A hybrid nanofiltration and reverse osmosis process for urine treatment: Effect on urea recovery and purity. Water. Res. 2022, 222, 118851. [Google Scholar] [CrossRef]
- Mohammad, A.W.; Basha, R.K.; Leo, C.P. Nanofiltration of glucose solution containing salts: Effects of membrane characteristics, organic component and salts on retention. J. Food Eng. 2009, 97, 510–518. [Google Scholar] [CrossRef]
- Almazán, J.E.; Romero-Dondiz, E.M.; Rajal, V.B.; Castro-Vidaurre, E.F. Nanofiltration of glucose: Analysis of parameters and membrane characterization. Chem. Eng. Res. Des. 2014, 94, 485–493. [Google Scholar] [CrossRef]
- Sjöman, E.; Mänttäri, M.; Nyström, M.; Koivikko, H.; Heikkilä, H. Separation of xylose from glucose by nanofiltration from concentrated monosaccharide solutions. J. Membr. Sci. 2007, 292, 106–115. [Google Scholar] [CrossRef]
- Vellenga, E.; Trägårdh, G. Nanofiltration of combined salt and sugar solutions: Coupling between retentions. Desalination 1998, 120, 211–220. [Google Scholar] [CrossRef]
- Bargeman, G.; Westerink, J.B.; Miguez, O.G.; Wessling, M. The effect of NaCl and glucose concentration on retentions for nanofiltration membranes processing concentrated solutions. Sep Purif. Technol. 2014, 134, 46–57. [Google Scholar] [CrossRef]
- Bouchoux, A.; Roux-de Balmann, H.; Lutin, F. Nanofiltration of glucose and sodium lactate solutions: Variations of retention between single- and mixed-solute solutions. J. Membr. Sci. 2005, 258, 123–132. [Google Scholar] [CrossRef]
- Nilsson, M.; Trägårdh, G.; Östergren, K. The influence of pH, salt and temperature on nanofiltration performance. J. Membr. Sci. 2008, 312, 97–106. [Google Scholar] [CrossRef]
- Freger, V.; Arnot, T.C.; Howell, J.A. Separation of concentrated organic/inorganic salt mixtures by nanofiltration. J. Membr. Sci. 2000, 178, 185–193. [Google Scholar] [CrossRef]
- Bandini, S.; Morelli, V. Effect of temperature, pH and composition on nanofiltration of mono/disaccharides: Experiments and modeling assessment. J. Membr. Sci. 2017, 533, 57–74. [Google Scholar] [CrossRef]
- Li, Y.; Wang, S.; Li, H.; Kang, G.; Sun, Y.; Yu, H.; Jin, Y.; Cao, Y. Preparation of highly selective nanofiltration membranes by moderately increasing pore size and optimizing microstructure of polyamide layer. J. Membr. Sci. 2022, 643, 120056. [Google Scholar] [CrossRef]
- Wang, E.; Lv, X.; Liu, S.; Dong, Q.; Li, J.; Li, H.; Su, B. A Selective Separation Mechanism for Mono/divalent Cations and Properties of a Hollow-Fiber Composite Nanofiltration Membrane Having a Positively Charged Surface. Membranes 2024, 14, 1. [Google Scholar] [CrossRef]
- Pagès, N.; Reig, M.; Gibert, O.; Cortina, J.L. Trace ions rejection tunning in NF by selecting solution composition: Ion permeances estimation. Chem. Eng. J. 2017, 308, 126–134. [Google Scholar] [CrossRef]
- Ben Amar, N.; Saidani, H.; Deratani, A.; Paimeri, J. Effect of temperature on the transport of water and neutral solutes across nanofiltration membranes. Langmuir 2006, 23, 2937–2952. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Membrane Name | Membrane Composition | MWCO (Da) | Active Area (m2) | Nominal Salt Rejections | Manufacturer |
|---|---|---|---|---|---|
| NF3 | polyamide | ND | 1.3 | ~45% | Axeon |
| NF4 | polyamide | ND | 1.3 | ~85% | Axeon |
| NF90 | polyamide | 180 Da | 2.6 | 97.0% | Dupont |
| NF270 | polypiperazine | 340 Da | 2.6 | >97.0% | Dupont |
| dnf40 | polysulfone | 400 Da | 0.05 | 91% | NX Filtration |
| dnf80 | polysulfone | 800 Da | 0.05 | 76% | NX Filtration |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duncan, H.; Newton, C.; Hestekin, J.; Hestekin, C.; Kurtz, I. Exploring Nanofiltration for Transport of Small Molecular Species for Application in Artificial Kidney Devices to Treat End-Stage Kidney Disease. Membranes 2025, 15, 168. https://doi.org/10.3390/membranes15060168
Duncan H, Newton C, Hestekin J, Hestekin C, Kurtz I. Exploring Nanofiltration for Transport of Small Molecular Species for Application in Artificial Kidney Devices to Treat End-Stage Kidney Disease. Membranes. 2025; 15(6):168. https://doi.org/10.3390/membranes15060168
Chicago/Turabian StyleDuncan, Haley, Christopher Newton, Jamie Hestekin, Christa Hestekin, and Ira Kurtz. 2025. "Exploring Nanofiltration for Transport of Small Molecular Species for Application in Artificial Kidney Devices to Treat End-Stage Kidney Disease" Membranes 15, no. 6: 168. https://doi.org/10.3390/membranes15060168
APA StyleDuncan, H., Newton, C., Hestekin, J., Hestekin, C., & Kurtz, I. (2025). Exploring Nanofiltration for Transport of Small Molecular Species for Application in Artificial Kidney Devices to Treat End-Stage Kidney Disease. Membranes, 15(6), 168. https://doi.org/10.3390/membranes15060168