

Bacterial Outer Membrane Vesicles and Immune Modulation of the Host

 ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
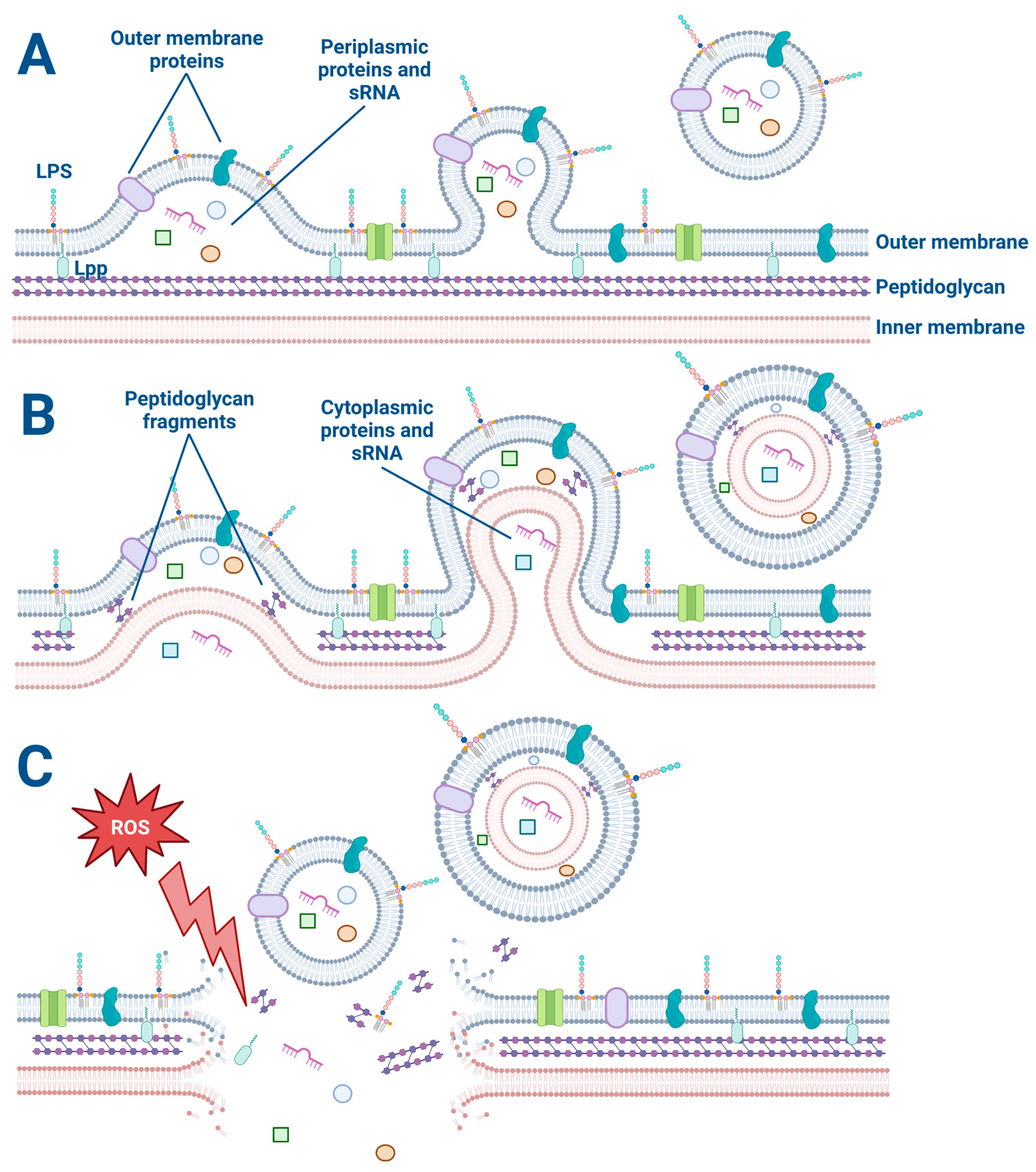
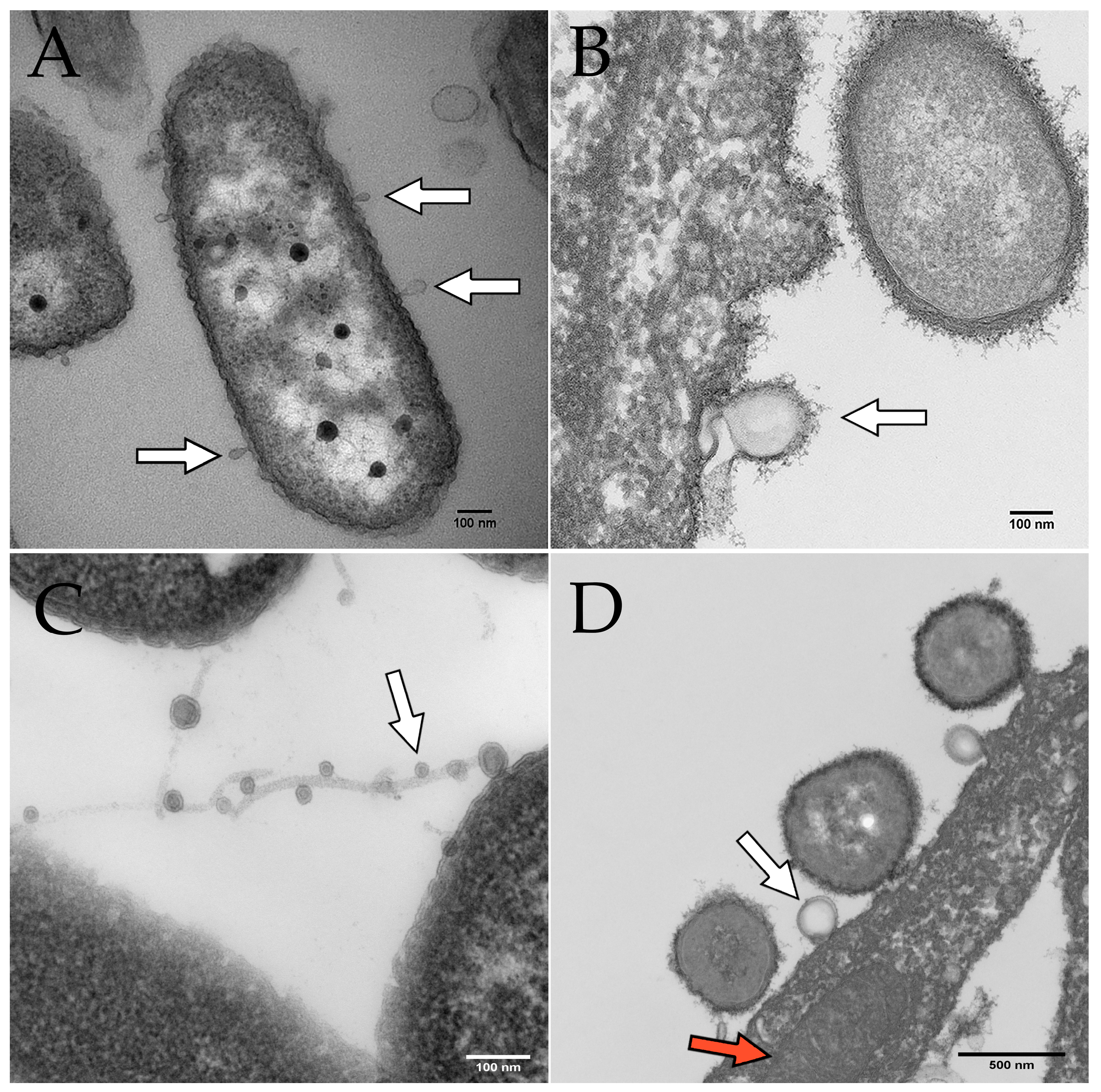
2. OMV Biogenesis
3. OMV Factors Modulate the Host Immune Response to Infection
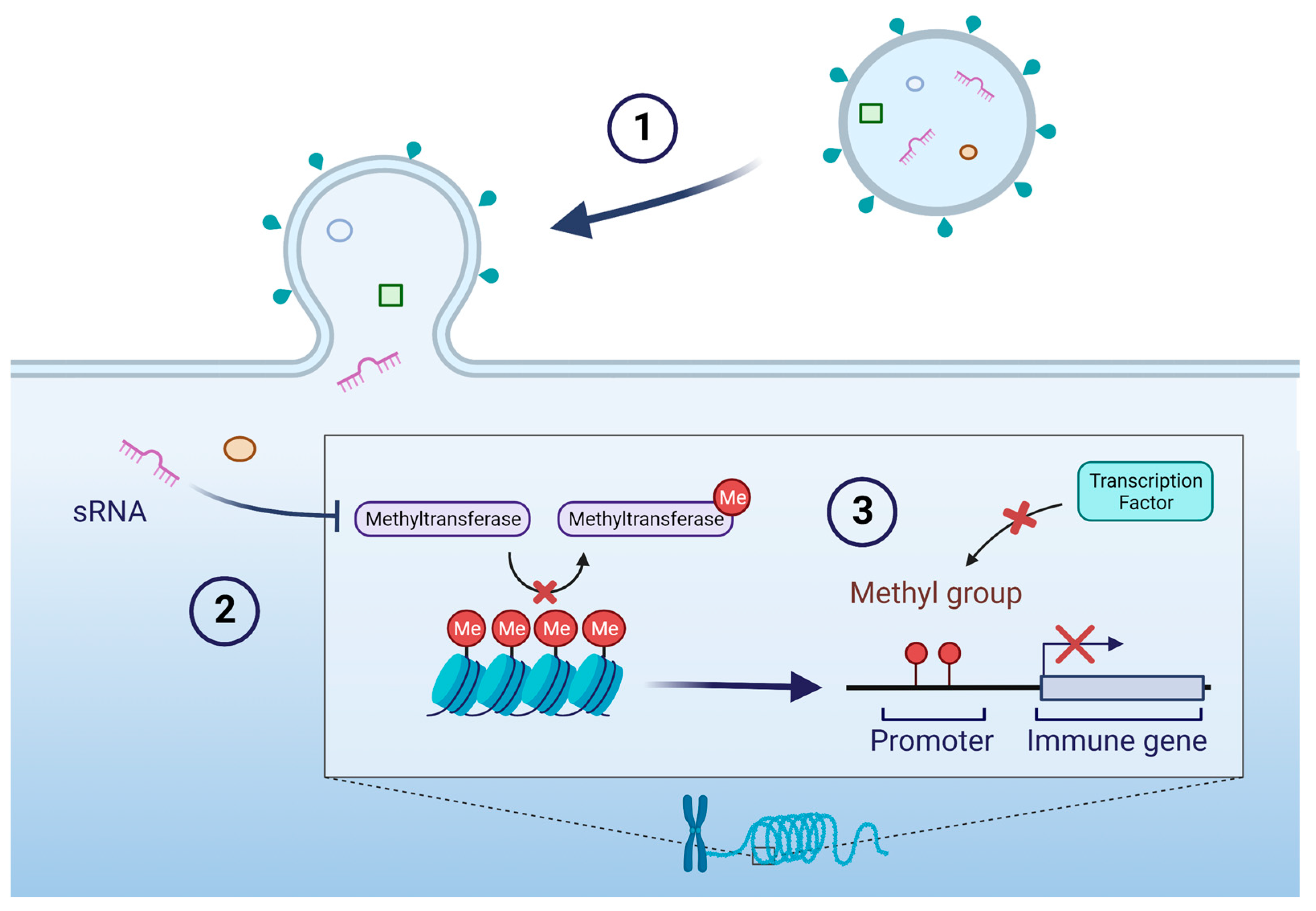
4. OMVs Alter the Host Immune Response to Subsequent Bacterial Infections through Alternative Methylation of Immune Genes
5. OMV Characterization
6. Conclusions and Future Research Prospects
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yáñez-Mó, M.; Siljander, P.R.M.; Andreu, Z.; Zavec, A.B.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological Properties of Extracellular Vesicles and Their Physiological Functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef]
- Koeppen, K.; Hampton, T.H.; Jarek, M.; Scharfe, M.; Gerber, S.A.; Mielcarz, D.W.; Demers, E.G.; Dolben, E.L.; Hammond, J.H.; Hogan, D.A.; et al. A Novel Mechanism of Host-Pathogen Interaction through SRNA in Bacterial Outer Membrane Vesicles. PLoS Pathog. 2016, 12, e1005672. [Google Scholar] [CrossRef]
- Kikuchi, Y.; Obana, N.; Toyofuku, M.; Kodera, N.; Soma, T.; Ando, T.; Fukumori, Y.; Nomura, N.; Taoka, A. Diversity of Physical Properties of Bacterial Extracellular Membrane Vesicles Revealed through Atomic Force Microscopy Phase Imaging. Nanoscale 2020, 12, 7950–7959. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, A.; Zechmann, B.; Sayes, C.M.; Taube, J.H.; Greathouse, K.L. A Preparation of Bacterial Outer Membrane with Osmium Tetroxide and Uranyl Acetate Co-Stain Enables Improved Structural Determination by Transmission Electron Microscopy. Microscopy 2023, dfad027. [Google Scholar] [CrossRef] [PubMed]
- Kolling, G.L.; Matthews, K.R. Export of Virulence Genes and Shiga Toxin by Membrane Vesicles of Escherichia coli O157:H7. Appl. Environ. Microbiol. 1999, 65, 1843–1848. [Google Scholar] [CrossRef]
- Grenier, D.; Mayrand, D. Functional Characterization of Extracellular Vesicles Produced by Bacteroides Gingivalis. Infect. Immun. 1987, 55, 111. [Google Scholar] [CrossRef]
- Wang, Y.F.; Fu, J. Secretory and Circulating Bacterial Small RNAs: A Mini-Review of the Literature. ExRNA 2019, 1, 14. [Google Scholar] [CrossRef]
- Dorward, D.W.; Garon, C.F. DNA Is Packaged within Membrane-Derived Vesicles of Gram-Negative but Not Gram-Positive Bacteria. Appl. Environ. Microbiol. 1990, 56, 1960. [Google Scholar] [CrossRef] [PubMed]
- Kuehn, M.J.; Kesty, N.C. Bacterial Outer Membrane Vesicles and the Host-Pathogen Interaction. Genes Dev. 2005, 19, 2645–2655. [Google Scholar] [CrossRef] [PubMed]
- Diallo, I.; Provost, P. RNA-Sequencing Analyses of Small Bacterial RNAs and Their Emergence as Virulence Factors in Host-Pathogen Interactions. Int. J. Mol. Sci. 2020, 21, 1627. [Google Scholar] [CrossRef]
- Chatterjee, S.N.; Das, J. Electron Microscopic Observations on the Excretion of Cell-Wall Material by Vibrio Cholerae. J. Gen. Microbiol. 1967, 49, 1–11. [Google Scholar] [CrossRef]
- Pérez-Cruz, C.; Delgado, L.; López-Iglesias, C.; Mercade, E. Outer-Inner Membrane Vesicles Naturally Secreted by Gram-Negative Pathogenic Bacteria. PLoS ONE 2015, 10, e0116896. [Google Scholar] [CrossRef] [PubMed]
- Kaparakis-Liaskos, M.; Ferrero, R.L. Immune Modulation by Bacterial Outer Membrane Vesicles. Nat. Rev. Immunol. 2015, 15, 375–387. [Google Scholar] [CrossRef]
- Anand, D.; Chaudhuri, A. Bacterial Outer Membrane Vesicles: New Insights and Applications. Mol. Membr. Biol. 2016, 33, 125–137. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.A.Q.; Besio, R.; Xiao, L.; Forlino, A. Outer Membrane Vesicles (OMVs) as Biomedical Tools and Their Relevance as Immune-Modulating Agents against H. Pylori Infections: Current Status and Future Prospects. Int. J. Mol. Sci. 2023, 24, 8542. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Lei, Q.; Zou, X.; Ma, D. The Role and Mechanisms of Gram-Negative Bacterial Outer Membrane Vesicles in Inflammatory Diseases. Front. Immunol. 2023, 14, 1157813. [Google Scholar] [CrossRef]
- Tiku, V.; Tan, M.W. Host Immunity and Cellular Responses to Bacterial Outer Membrane Vesicles. Trends Immunol. 2021, 42, 1024–1036. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Suh, J.W.; Kang, J.S.; Kim, S.B.; Yoon, Y.K.; Sohn, J.W. Gram-Negative Bacteria’s Outer Membrane Vesicles. Infect. Chemother. 2023, 55, 1. [Google Scholar] [CrossRef]
- Zhao, G.; Jones, M.K. Role of Bacterial Extracellular Vesicles in Manipulating Infection. Infect. Immun. 2023, 91, e0043922. [Google Scholar] [CrossRef]
- Stanton, B.A. Extracellular Vesicles and Host–Pathogen Interactions: A Review of Inter-Kingdom Signaling by Small Noncoding RNA. Genes 2021, 12, 1010. [Google Scholar] [CrossRef]
- Gilmore, W.J.; Johnston, E.L.; Zavan, L.; Bitto, N.J.; Kaparakis-Liaskos, M. Immunomodulatory Roles and Novel Applications of Bacterial Membrane Vesicles. Mol. Immunol. 2021, 134, 72–85. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.K.; Armstrong, D.A.; Hazlett, H.F.; Dessaint, J.A.; Mellinger, D.L.; Aridgides, D.S.; Christensen, B.C.; Ashare, A. Exposure to Extracellular Vesicles from Pseudomonas aeruginosa Result in Loss of DNA Methylation at Enhancer and DNase Hypersensitive Site Regions in Lung Macrophages. Epigenetics 2021, 16, 1187. [Google Scholar] [CrossRef]
- Fonseca, S.; Carvalho, A.L.; Miquel-Clopés, A.; Jones, E.J.; Juodeikis, R.; Stentz, R.; Carding, S.R. Extracellular Vesicles Produced by the Human Gut Commensal Bacterium Bacteroides Thetaiotaomicron Elicit Anti-Inflammatory Responses from Innate Immune Cells. Front. Microbiol. 2022, 13, 1050271. [Google Scholar] [CrossRef] [PubMed]
- Turnbull, L.; Toyofuku, M.; Hynen, A.L.; Kurosawa, M.; Pessi, G.; Petty, N.K.; Osvath, S.R.; Cárcamo-Oyarce, G.; Gloag, E.S.; Shimoni, R.; et al. Explosive Cell Lysis as a Mechanism for the Biogenesis of Bacterial Membrane Vesicles and Biofilms. Nat. Commun. 2016, 7, 11220. [Google Scholar] [CrossRef] [PubMed]
- Kulp, A.; Kuehn, M.J. Biological Functions and Biogenesis of Secreted Bacterial Outer Membrane Vesicles. Annu. Rev. Microbiol. 2010, 64, 163. [Google Scholar] [CrossRef] [PubMed]
- McBroom, A.J.; Kuehn, M.J. Release of Outer Membrane Vesicles by Gram-Negative Bacteria Is a Novel Envelope Stress Response. Mol. Microbiol. 2007, 63, 545. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, B.V.; Kuehn, M.J. Staphylococcus Aureus Secretes Immunomodulatory RNA and DNA via Membrane Vesicles. Sci. Rep. 2020, 10, 18293. [Google Scholar] [CrossRef] [PubMed]
- Melo, J.; Pinto, V.; Fernandes, T.; Malheiro, A.R.; Osório, H.; Figueiredo, C.; Leite, M. Isolation Method and Characterization of Outer Membranes Vesicles of Helicobacter Pylori Grown in a Chemically Defined Medium. Front. Microbiol. 2021, 12, 1253. [Google Scholar] [CrossRef]
- Schwechheimer, C.; Kuehn, M.J. Outer-Membrane Vesicles from Gram-Negative Bacteria: Biogenesis and Functions. Nat. Rev. Microbiol. 2015, 13, 605. [Google Scholar] [CrossRef]
- Haurat, M.F.; Elhenawy, W.; Feldman, M.F. Prokaryotic Membrane Vesicles: New Insights on Biogenesis and Biological Roles. Biol. Chem. 2015, 396, 95–109. [Google Scholar] [CrossRef]
- Zhou, L.; Srisatjaluk, R.; Justus, D.E.; Doyle, R.J. On the Origin of Membrane Vesicles in Gram-Negative Bacteria. FEMS Microbiol. Lett. 1998, 163, 223–228. [Google Scholar] [CrossRef]
- Zavan, L.; Fang, H.; Johnston, E.L.; Whitchurch, C.; Greening, D.W.; Hill, A.F.; Kaparakis-Liaskos, M. The Mechanism of Pseudomonas aeruginosa Outer Membrane Vesicle Biogenesis Determines Their Protein Composition. Proteomics 2023, 23, 2200464. [Google Scholar] [CrossRef] [PubMed]
- O’Donoghue, E.J.; Krachler, A.M. Mechanisms of Outer Membrane Vesicle Entry into Host Cells. Cell Microbiol. 2016, 18, 1508. [Google Scholar] [CrossRef] [PubMed]
- Amano, A.; Takeuchi, H.; Furuta, N. Outer Membrane Vesicles Function as Offensive Weapons in Host-Parasite Interactions. Microbes Infect. 2010, 12, 791–798. [Google Scholar] [CrossRef]
- Bierwagen, J.; Wiegand, M.; Laakmann, K.; Danov, O.; Limburg, H.; Herbel, S.M.; Heimerl, T.; Dorna, J.; Jonigk, D.; Preußer, C.; et al. Bacterial Vesicles Block Viral Replication in Macrophages via TLR4-TRIF-Axis. Cell Commun. Signal 2023, 21, 65. [Google Scholar] [CrossRef] [PubMed]
- Mancini, F.; Rossi, O.; Necchi, F.; Micoli, F. OMV Vaccines and the Role of TLR Agonists in Immune Response. Int. J. Mol. Sci. 2020, 21, 4416. [Google Scholar] [CrossRef] [PubMed]
- Balhuizen, M.D.; Versluis, C.M.; van Grondelle, M.O.; Veldhuizen, E.J.A.; Haagsman, H.P. Modulation of Outer Membrane Vesicle-Based Immune Responses by Cathelicidins. Vaccine 2022, 40, 2399–2408. [Google Scholar] [CrossRef]
- Feuillet, V.; Medjane, S.; Mondor, I.; Demaria, O.; Pagni, P.P.; Galán, J.E.; Flavell, R.A.; Alexopoulou, L. Involvement of Toll-like Receptor 5 in the Recognition of Flagellated Bacteria. Proc. Natl. Acad. Sci. USA 2006, 103, 12487–12492. [Google Scholar] [CrossRef]
- Poltorak, A.; He, X.; Smirnova, I.; Liu, M.Y.; Van Huffel, C.; Du, X.; Birdwell, D.; Alejos, E.; Silva, M.; Galanos, C.; et al. Defective LPS Signaling in C3H/HeJ and C57BL/10ScCr Mice: Mutations in Tlr4 Gene. Science 1998, 282, 2085–2088. [Google Scholar] [CrossRef]
- Hasan, A.; Akhter, N.; Al-Roub, A.; Thomas, R.; Kochumon, S.; Wilson, A.; Koshy, M.; Al-Ozairi, E.; Al-Mulla, F.; Ahmad, R. TNF-α in Combination with Palmitate Enhances IL-8 Production via The MyD88- Independent TLR4 Signaling Pathway: Potential Relevance to Metabolic Inflammation. Int. J. Mol. Sci. 2019, 20, 4112. [Google Scholar] [CrossRef]
- Baker, S.M.; Settles, E.W.; Davitt, C.; Gellings, P.; Kikendall, N.; Hoffmann, J.; Wang, Y.; Bitoun, J.; Lodrigue, K.R.; Sahl, J.W.; et al. Burkholderia Pseudomallei OMVs Derived from Infection Mimicking Conditions Elicit Similar Protection to a Live-Attenuated Vaccine. npj Vaccines 2021, 6, 18. [Google Scholar] [CrossRef] [PubMed]
- Stevens, M.P.; Friebel, A.; Taylor, L.A.; Wood, M.W.; Brown, P.J.; Hardt, W.D.; Galyov, E.E. A Burkholderia Pseudomallei Type III Secreted Protein, BopE, Facilitates Bacterial Invasion of Epithelial Cells and Exhibits Guanine Nucleotide Exchange Factor Activity. J. Bacteriol. 2003, 185, 4992. [Google Scholar] [CrossRef]
- Anderson, G.G.; Moreau-Marquis, S.; Stanton, B.A.; O’Toole, G.A. In Vitro Analysis of Tobramycin-Treated Pseudomonas aeruginosa Biofilms on Cystic Fibrosis-Derived Airway Epithelial Cells. Infect. Immun. 2008, 76, 1423. [Google Scholar] [CrossRef] [PubMed]
- Hvorecny, K.L.; Dolben, E.; Moreau-Marquis, S.; Hampton, T.H.; Shabaneh, T.B.; Flitter, B.A.; Bahl, C.D.; Bomberger, J.M.; Levy, B.D.; Stanton, B.A.; et al. An Epoxide Hydrolase Secreted by Pseudomonas aeruginosa Decreases Mucociliary Transport and Hinders Bacterial Clearance from the Lung. Am. J. Physiol. Lung Cell Mol. Physiol. 2018, 314, L150–L156. [Google Scholar] [CrossRef]
- Hvorecny, K.L.; Bahl, C.D.; Kitamura, S.; Lee, K.S.S.; Hammock, B.D.; Morisseau, C.; Madden, D.R. Active-Site Flexibility and Substrate Specificity in a Bacterial Virulence Factor: Crystallographic Snapshots of an Epoxide Hydrolase. Structure 2017, 25, 697. [Google Scholar] [CrossRef]
- Bomberger, J.M.; MacEachran, D.P.; Coutermarsh, B.A.; Ye, S.; O’Toole, G.A.; Stanton, B.A. Long-Distance Delivery of Bacterial Virulence Factors by Pseudomonas aeruginosa Outer Membrane Vesicles. PLoS Pathog. 2009, 5, 1000382. [Google Scholar] [CrossRef]
- Ballok, A.E.; Filkins, L.M.; Bomberger, J.M.; Stanton, B.A.; O’Toole, G.A. Epoxide-Mediated Differential Packaging of Cif and Other Virulence Factors into Outer Membrane Vesicles. J. Bacteriol. 2014, 196, 3633. [Google Scholar] [CrossRef]
- MacEachran, D.P.; Ye, S.; Bomberger, J.M.; Hogan, D.A.; Swiatecka-Urban, A.; Stanton, B.A.; O’Toole, G.A. The Pseudomonas aeruginosa Secreted Protein PA2934 Decreases Apical Membrane Expression of the Cystic Fibrosis Transmembrane Conductance Regulator. Infect. Immun. 2007, 75, 3902. [Google Scholar] [CrossRef]
- Kitamura, S.; Hvorecny, K.L.; Niu, J.; Hammock, B.D.; Madden, D.R.; Morisseau, C. Rational Design of Potent and Selective Inhibitors of an Epoxide Hydrolase Virulence Factor from Pseudomonas aeruginosa. J. Med. Chem. 2016, 59, 4790. [Google Scholar] [CrossRef] [PubMed]
- Bomberger, J.M.; Barnaby, R.L.; Stanton, B.A. The Deubiquitinating Enzyme USP10 Regulates the Endocytic Recycling of CFTR in Airway Epithelial Cells. Channels 2010, 4, 150. [Google Scholar] [CrossRef]
- Ballok, A.E.; O’Toole, G.A. Pouring Salt on a Wound: Pseudomonas aeruginosa Virulence Factors Alter Na+ and Cl− Flux in the Lung. J. Bacteriol. 2013, 195, 4013. [Google Scholar] [CrossRef] [PubMed]
- Bahl, C.D.; Morisseau, C.; Bomberger, J.M.; Stanton, B.A.; Hammock, B.D.; O’Toole, G.A.; Madden, D.R. Crystal Structure of the Cystic Fibrosis Transmembrane Conductance Regulator Inhibitory Factor Cif Reveals Novel Active-Site Features of an Epoxide Hydrolase Virulence Factor. J. Bacteriol. 2010, 192, 1785. [Google Scholar] [CrossRef] [PubMed]
- Bahl, C.D.; Hvorecny, K.L.; Bomberger, J.M.; Stanton, B.A.; Hammock, B.D.; Morisseau, C.; Madden, D.R. Inhibiting an Epoxide Hydrolase Virulence Strategy Protects CFTR. Angew. Chem. Int. Ed. Engl. 2015, 54, 9881. [Google Scholar] [CrossRef]
- Bomberger, J.M.; Ye, S.; MacEachran, D.P.; Koeppen, K.; Barnaby, R.L.; O’Toole, G.A.; Stanton, B.A. A Pseudomonas aeruginosa Toxin That Hijacks the Host Ubiquitin Proteolytic System. PLoS Pathog. 2011, 7, e1001325. [Google Scholar] [CrossRef] [PubMed]
- Bomberger, J.M.; Ely, K.H.; Bangia, N.; Ye, S.; Green, K.A.; Green, W.R.; Enelow, R.I.; Stanton, B.A. Pseudomonas aeruginosa Cif Protein Enhances the Ubiquitination and Proteasomal Degradation of the Transporter Associated with Antigen Processing (TAP) and Reduces Major Histocompatibility Complex (MHC) Class I Antigen Presentation. J. Biol. Chem. 2014, 289, 152–162. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.M.; Carvalho, H.M.; Rasmussen, S.B.; O’Brien, A.D. Cytotoxic Necrotizing Factor Type 1 Delivered by Outer Membrane Vesicles of Uropathogenic Escherichia coli Attenuates Polymorphonuclear Leukocyte Antimicrobial Activity and Chemotaxis. Infect. Immun. 2006, 74, 4401. [Google Scholar] [CrossRef] [PubMed]
- Hatakeyama, M.; Higashi, H. Helicobacter Pylori CagA: A New Paradigm for Bacterial Carcinogenesis. Cancer Sci. 2005, 96, 835–843. [Google Scholar] [CrossRef] [PubMed]
- Turkina, M.V.; Olofsson, A.; Magnusson, K.E.; Arnqvist, A.; Vikström, E. Helicobacter Pylori Vesicles Carrying CagA Localize in the Vicinity of Cell-Cell Contacts and Induce Histone H1 Binding to ATP in Epithelial Cells. FEMS Microbiol. Lett. 2015, 362, fnv076. [Google Scholar] [CrossRef]
- Chmiela, M.; Walczak, N.; Rudnicka, K. Helicobacter Pylori Outer Membrane Vesicles Involvement in the Infection Development and Helicobacter Pylori-Related Diseases. J. Biomed. Sci. 2018, 25, 78. [Google Scholar] [CrossRef]
- Elhenawy, W.; Debelyy, M.O.; Feldman, M.F. Preferential Packing of Acidic Glycosidases and Proteases into Bacteroides Outer Membrane Vesicles. mBio 2014, 5, e00909-14. [Google Scholar] [CrossRef]
- Rompikuntal, P.K.; Vdovikova, S.; Duperthuy, M.; Johnson, T.L.; Åhlund, M.; Lundmark, R.; Oscarsson, J.; Sandkvist, M.; Uhlin, B.E.; Wai, S.N. Outer Membrane Vesicle-Mediated Export of Processed PrtV Protease from Vibrio cholerae. PLoS ONE 2015, 10, e0134098. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.W.; Kim, S.C.; Hong, S.H.; Lee, H.J. Secretable Small RNAs via Outer Membrane Vesicles in Periodontal Pathogens. J. Dent. Res. 2017, 96, 458–466. [Google Scholar] [CrossRef]
- Gu, H.; Zhao, C.; Zhang, T.; Liang, H.; Wang, X.M.; Pan, Y.; Chen, X.; Zhao, Q.; Li, D.; Liu, F.; et al. Salmonella Produce MicroRNA-like RNA Fragment Sal-1 in the Infected Cells to Facilitate Intracellular Survival. Sci. Rep. 2017, 7, 2392. [Google Scholar] [CrossRef]
- Furuse, Y.; Finethy, R.; Saka, H.A.; Xet-Mull, A.M.; Sisk, D.M.; Jurcic Smith, K.L.; Lee, S.; Coers, J.; Valdivia, R.H.; Tobin, D.M.; et al. Search for MicroRNAs Expressed by Intracellular Bacterial Pathogens in Infected Mammalian Cells. PLoS ONE 2014, 9, 106434. [Google Scholar] [CrossRef]
- Maute, R.L.; Schneider, C.; Sumazin, P.; Holmes, A.; Califano, A.; Basso, K.; Dalla-Favera, R. TRNA-Derived MicroRNA Modulates Proliferation and the DNA Damage Response and Is down-Regulated in B Cell Lymphoma. Proc. Natl. Acad. Sci. USA 2013, 110, 1404–1409. [Google Scholar] [CrossRef]
- Yamasaki, S.; Ivanov, P.; Hu, G.F.; Anderson, P. Angiogenin Cleaves TRNA and Promotes Stress-Induced Translational Repression. J. Cell Biol. 2009, 185, 35. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.; Wilson, B.; Kumar, P.; Dutta, A. Non-Canonical Roles of TRNAs: TRNA Fragments and Beyond. Annu. Rev. Genet. 2020, 54, 47. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Koeppen, K.; Ashare, A.; Hogan, D.A.; Gerber, S.A.; Stanton, A.; Stanton, B.A. Tobramycin-Induced Secretion of P. Aeruginosa 5′ TRNA-FMet Halves Suppresses Lung Inflammation via AGO2 Gene Silencing. bioRxiv 2021. bioRxiv:2021.09.23.461540. [Google Scholar] [CrossRef]
- Ochando, J.; Mulder, W.J.M.; Madsen, J.C.; Netea, M.G.; Duivenvoorden, R. Trained Immunity—Basic Concepts and Contributions to Immunopathology. Nat. Rev. Nephrol. 2022, 19, 23–37. [Google Scholar] [CrossRef]
- Netea, M.G.; Quintin, J.; Van Der Meer, J.W.M. Trained Immunity: A Memory for Innate Host Defense. Cell Host Microbe 2011, 9, 355–361. [Google Scholar] [CrossRef]
- Netea, M.G.; Joosten, L.A.B.; Latz, E.; Mills, K.H.G.; Natoli, G.; Stunnenberg, H.G.; O’Neill, L.A.J.; Xavier, R.J. Trained Immunity: A Program of Innate Immune Memory in Health and Disease. Science 2016, 352, 427. [Google Scholar] [CrossRef]
- Govan, J.R.W.; Nelson, J.W. Microbiology of Lung Infection in Cystic Fibrosis. Br. Med. Bull. 1992, 48, 912–930. [Google Scholar] [CrossRef] [PubMed]
- Lyczak, J.B.; Cannon, C.L.; Pier, G.B. Lung Infections Associated with Cystic Fibrosis. Clin. Microbiol. Rev. 2002, 15, 194–222. [Google Scholar] [CrossRef] [PubMed]
- Jurado-Martín, I.; Sainz-Mejías, M.; Mcclean, S. Molecular Sciences Pseudomonas aeruginosa: An Audacious Pathogen with an Adaptable Arsenal of Virulence Factors. Int. J. Mol. Sci. 2021, 22, 3128. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Armstrong, D.A.; Salas, L.A.; Hazlett, H.F.; Nymon, A.B.; Dessaint, J.A.; Aridgides, D.S.; Mellinger, D.L.; Liu, X.; Christensen, B.C.; et al. Genome-Wide DNA Methylation Profiling Shows a Distinct Epigenetic Signature Associated with Lung Macrophages in Cystic Fibrosis. Clin. Epigenet. 2018, 10, 152. [Google Scholar] [CrossRef]
- Magalhães, M.; Tost, J.; Pineau, F.; Rivals, I.; Busato, F.; Alary, N.; Mely, L.; Leroy, S.; Murris, M.; Caimmi, D.; et al. Dynamic Changes of DNA Methylation and Lung Disease in Cystic Fibrosis: Lessons from a Monogenic Disease. Epigenomics 2018, 10, 1131–1145. [Google Scholar] [CrossRef]
- Deaton, A.M.; Bird, A. CpG Islands and the Regulation of Transcription. Genes. Dev. 2011, 25, 1010–1022. [Google Scholar] [CrossRef]
- Jones, P.A. Functions of DNA Methylation: Islands, Start Sites, Gene Bodies and Beyond. Nat. Rev. Genet. 2012, 13, 484–492. [Google Scholar] [CrossRef]
- Gardner, K.E.; Allis, C.D.; Strahl, B.D. OPERating ON Chromatin, a Colorful Language Where Context Matters. J. Mol. Biol. 2011, 409, 36. [Google Scholar] [CrossRef]
- Crimi, E.; Benincasa, G.; Cirri, S.; Mutesi, R.; Faenza, M.; Napoli, C. Clinical Epigenetics and Multidrug-Resistant Bacterial Infections: Host Remodelling in Critical Illness. Epigenetics 2020, 15, 1021–1034. [Google Scholar] [CrossRef]
- Mackaness, G.B. The Immunological Basis of Acquired Cellular Resistance. J. Exp. Med. 1964, 120, 105. [Google Scholar] [CrossRef]
- Bigot, J.; Guillot, L.; Guitard, J.; Ruffin, M.; Corvol, H.; Chignard, M.; Hennequin, C.; Balloy, V. Respiratory Epithelial Cells Can Remember Infection: A Proof-of-Concept Study. J. Infect. Dis. 2020, 221, 1000–1005. [Google Scholar] [CrossRef] [PubMed]
- Brindisi, M.; Barone, S.; Rossi, A.; Cassese, E.; Del Gaudio, N.; Feliz Morel, Á.J.; Filocamo, G.; Alberico, A.; De Fino, I.; Gugliandolo, D.; et al. Efficacy of Selective Histone Deacetylase 6 Inhibition in Mouse Models of Pseudomonas aeruginosa Infection: A New Glimpse for Reducing Inflammation and Infection in Cystic Fibrosis. Eur. J. Pharmacol. 2022, 936, 175349. [Google Scholar] [CrossRef] [PubMed]
- Han, F.; Wang, W.; Shi, M.; Zhou, H.; Yao, Y.; Li, C.; Shang, A. Outer Membrane Vesicles from Bacteria: Role and Potential Value in the Pathogenesis of Chronic Respiratory Diseases. Front. Cell Infect. Microbiol. 2022, 12, 1093327. [Google Scholar] [CrossRef]
- Worlitzsch, D.; Tarran, R.; Ulrich, M.; Schwab, U.; Cekici, A.; Meyer, K.C.; Birrer, P.; Bellon, G.; Berger, J.; Weiss, T.; et al. Effects of Reduced Mucus Oxygen Concentration in Airway Pseudomonas Infections of Cystic Fibrosis Patients. J. Clin. Investig. 2002, 109, 317. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lin, S.; Wang, L.; Cao, Z.; Zhang, M.; Zhang, Y.; Liu, R.; Liu, J. Versatility of Bacterial Outer Membrane Vesicles in Regulating Intestinal Homeostasis. Sci. Adv. 2023, 9, eade5079. [Google Scholar] [CrossRef]
- Duncan, L.; Yoshioka, M.; Chandad, F.; Grenier, D. Loss of Lipopolysaccharide Receptor CD14 from the Surface of Human Macrophage-like Cells Mediated by Porphyromonas Gingivalis Outer Membrane Vesicles. Microb. Pathog. 2004, 36, 319–325. [Google Scholar] [CrossRef]
- Waller, T.; Kesper, L.; Hirschfeld, J.; Dommisch, H.; Kölpin, J.; Oldenburg, J.; Uebele, J.; Hoerauf, A.; Deschner, J.; Jepsen, S.; et al. Porphyromonas Gingivalis Outer Membrane Vesicles Induce Selective Tumor Necrosis Factor Tolerance in a Toll-Like Receptor 4- and MTOR-Dependent Manner. Infect. Immun. 2016, 84, 1194. [Google Scholar] [CrossRef]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal Information for Studies of Extracellular Vesicles 2018 (MISEV2018): A Position Statement of the International Society for Extracellular Vesicles and Update of the MISEV2014 Guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef]
- Gardiner, C.; Di Vizio, D.; Sahoo, S.; Théry, C.; Witwer, K.W.; Wauben, M.; Hill, A.F. Techniques Used for the Isolation and Characterization of Extracellular Vesicles: Results of a Worldwide Survey. J. Extracell. Vesicles 2016, 5, 32945. [Google Scholar] [CrossRef]
- Shi, R.; Dong, Z.; Ma, C.; Wu, R.; Lv, R.; Liu, S.; Ren, Y.; Liu, Z.; van der Mei, H.C.; Busscher, H.J.; et al. High-Yield, Magnetic Harvesting of Extracellular Outer-Membrane Vesicles from Escherichia coli. Small 2022, 18, e2204350. [Google Scholar] [CrossRef] [PubMed]
- Klimentová, J.; Stulík, J. Methods of Isolation and Purification of Outer Membrane Vesicles from Gram-Negative Bacteria. Microbiol. Res. 2015, 170, 1–9. [Google Scholar] [CrossRef]
- Witwer, K.W.; Buzás, E.I.; Bemis, L.T.; Bora, A.; Lässer, C.; Lötvall, J.; Nolte-’t Hoen, E.N.; Piper, M.G.; Sivaraman, S.; Skog, J.; et al. Standardization of Sample Collection, Isolation and Analysis Methods in Extracellular Vesicle Research. J. Extracell. Vesicles 2013, 2, 20360. [Google Scholar] [CrossRef]
- Reimer, S.L.; Beniac, D.R.; Hiebert, S.L.; Booth, T.F.; Chong, P.M.; Westmacott, G.R.; Zhanel, G.G.; Bay, D.C. Comparative Analysis of Outer Membrane Vesicle Isolation Methods with an Escherichia coli TolA Mutant Reveals a Hypervesiculating Phenotype with Outer-Inner Membrane Vesicle Content. Front. Microbiol. 2021, 12, 628801. [Google Scholar] [CrossRef]
- Filipe, V.; Hawe, A.; Jiskoot, W. Critical Evaluation of Nanoparticle Tracking Analysis (NTA) by NanoSight for the Measurement of Nanoparticles and Protein Aggregates. Pharm. Res. 2010, 27, 796. [Google Scholar] [CrossRef]
- Brennan, K.; Martin, K.; FitzGerald, S.P.; O’Sullivan, J.; Wu, Y.; Blanco, A.; Richardson, C.; Mc Gee, M.M. A Comparison of Methods for the Isolation and Separation of Extracellular Vesicles from Protein and Lipid Particles in Human Serum. Sci. Rep. 2020, 10, 1039. [Google Scholar] [CrossRef] [PubMed]
- Montis, C.; Zendrini, A.; Valle, F.; Busatto, S.; Paolini, L.; Radeghieri, A.; Salvatore, A.; Berti, D.; Bergese, P. Size Distribution of Extracellular Vesicles by Optical Correlation Techniques. Colloids Surf. B Biointerfaces 2017, 158, 331–338. [Google Scholar] [CrossRef]
- Vogel, R.; Coumans, F.A.W.; Maltesen, R.G.; Böing, A.N.; Bonnington, K.E.; Broekman, M.L.; Broom, M.F.; Buzás, E.I.; Christiansen, G.; Hajji, N.; et al. A Standardized Method to Determine the Concentration of Extracellular Vesicles Using Tunable Resistive Pulse Sensing. J. Extracell. Vesicles 2016, 5, 31242. [Google Scholar] [CrossRef] [PubMed]
- Ejjigu, N.; Abdelgadir, K.; Flaten, Z.; Hoff, C.; Li, C.Z.; Sun, D. Environmental Noise Reduction for Tunable Resistive Pulse Sensing of Extracellular Vesicles. Sens. Actuators A Phys. 2022, 346, 113832. [Google Scholar] [CrossRef]
- Nolan, J.P.; Duggan, E. Analysis of Individual Extracellular Vesicles by Flow Cytometry. Methods Mol. Biol. 2018, 1678, 79–92. [Google Scholar] [CrossRef]
- Wieser, A.; Storz, E.; Liegl, G.; Peter, A.; Pritsch, M.; Shock, J.; Wai, S.N.; Schubert, S. Efficient Quantification and Characterization of Bacterial Outer Membrane Derived Nano-Particles with Flow Cytometric Analysis. Int. J. Med. Microbiol. 2014, 304, 1032–1037. [Google Scholar] [CrossRef]
- Park, J.Y.; Choi, J.; Lee, Y.; Lee, J.E.; Lee, E.H.; Kwon, H.J.; Yang, J.; Jeong, B.R.; Kim, Y.K.; Han, P.L. Metagenome Analysis of Bodily Microbiota in a Mouse Model of Alzheimer Disease Using Bacteria-Derived Membrane Vesicles in Blood. Exp. Neurobiol. 2017, 26, 369–379. [Google Scholar] [CrossRef]
- Lee, Y.; Park, J.Y.; Lee, E.H.; Yang, J.; Jeong, B.R.; Kim, Y.K.; Seoh, J.Y.; Lee, S.H.; Han, P.L.; Kim, E.J. Rapid Assessment of Microbiota Changes in Individuals with Autism Spectrum Disorder Using Bacteria-Derived Membrane Vesicles in Urine. Exp. Neurobiol. 2017, 26, 307–317. [Google Scholar] [CrossRef]
- Yoo, J.Y.; Rho, M.; You, Y.-A.; Kwon, E.J.; Kim, M.-H.; Kym, S.; Jee, Y.-K.; Kim, Y.-K.; Kim, Y.J. 16S RRNA Gene-Based Metagenomic Analysis Reveals Differences in Bacteria-Derived Extracellular Vesicles in the Urine of Pregnant and Non-Pregnant Women. Exp. Mol. Med. 2016, 48, e208. [Google Scholar] [CrossRef] [PubMed]
- Schaack, B.; Hindré, T.; Quansah, N.; Hannani, D.; Mercier, C.; Laurin, D. Microbiota-Derived Extracellular Vesicles Detected in Human Blood from Healthy Donors. Int. J. Mol. Sci. 2022, 2022, 13787. [Google Scholar] [CrossRef] [PubMed]
- Jones, E.J.; Booth, C.; Fonseca, S.; Parker, A.; Cross, K.; Miquel-Clopés, A.; Hautefort, I.; Mayer, U.; Wileman, T.; Stentz, R.; et al. The Uptake, Trafficking, and Biodistribution of Bacteroides Thetaiotaomicron Generated Outer Membrane Vesicles. Front. Microbiol. 2020, 11, 57. [Google Scholar] [CrossRef] [PubMed]
- Sampath, V.; McCaig, W.D.; Thanassi, D.G. Amino Acid Deprivation and Central Carbon Metabolism Regulate the Production of Outer Membrane Vesicles and Tubes by Francisella. Mol. Microbiol. 2018, 107, 523–541. [Google Scholar] [CrossRef]
- Pierson, T.; Matrakas, D.; Taylor, Y.U.; Manyam, G.; Morozov, V.N.; Zhou, W.; Van Hoek, M.L. Proteomic Characterization and Functional Analysis of Outer Membrane Vesicles of Francisella Novicida Suggests Possible Role in Virulence and Use as a Vaccine. J. Proteome Res. 2011, 10, 954–967. [Google Scholar] [CrossRef]
- Santos, S.; Juncioni De Arauz, L.; Baruque-Ramos, J.; Lebrun, I.; Mendes Carneiro, S.; Alves Barreto, S.; Perazzini, R.; Schenkman, F. Outer Membrane Vesicles (OMV) Production of Neisseria Meningitidis Serogroup B in Batch Process. Vaccine 2012, 30, 6064–6069. [Google Scholar] [CrossRef]
- De Jonge, E.F.; Balhuizen, M.D.; Van Boxtel, R.; Wu, J.; Haagsman, H.P.; Tommassen, J. Heat Shock Enhances Outer-Membrane Vesicle Release in Bordetella spp. Curr. Res. Microb. Sci. 2021, 2, 100009. [Google Scholar] [CrossRef]
- Bonnington, K.E.; Kuehn, M.J. Outer Membrane Vesicle Production Facilitates LPS Remodeling and Outer Membrane Maintenance in Salmonella during Environmental Transitions. mBio 2017, 4, 1532–1548. [Google Scholar] [CrossRef] [PubMed]
- Palmer, K.L.; Aye, L.M.; Whiteley, M. Nutritional Cues Control Pseudomonas aeruginosa Multicellular Behavior in Cystic Fibrosis Sputum. J. Bacteriol. 2007, 189, 8079. [Google Scholar] [CrossRef] [PubMed]
- Palmer, K.L.; Mashburn, L.M.; Singh, P.K.; Whiteley, M. Cystic Fibrosis Sputum Supports Growth and Cues Key Aspects of Pseudomonas aeruginosa Physiology. J. Bacteriol. 2005, 187, 5267. [Google Scholar] [CrossRef]
- Turner, K.H.; Wessel, A.K.; Palmer, G.C.; Murray, J.L.; Whiteley, M. Essential Genome of Pseudomonas aeruginosa in Cystic Fibrosis Sputum. Proc. Natl. Acad. Sci. USA 2015, 112, 4110–4115. [Google Scholar] [CrossRef]
- Cooke, A.C.; Nello, A.V.; Ernst, R.K.; Schertzer, J.W. Analysis of Pseudomonas aeruginosa Biofilm Membrane Vesicles Supports Multiple Mechanisms of Biogenesis. PLoS ONE 2019, 14, e0212275. [Google Scholar] [CrossRef] [PubMed]
- Johnston, E.L.; Zavan, L.; Bitto, N.J.; Petrovski, S.; Hill, A.F.; Kaparakis-Liaskos, M. Planktonic and Biofilm-Derived Pseudomonas aeruginosa Outer Membrane Vesicles Facilitate Horizontal Gene Transfer of Plasmid DNA. Microbiol. Spectr. 2023, 11, e0517922. [Google Scholar] [CrossRef] [PubMed]
- Toyofuku, M.; Roschitzki, B.; Riedel, K.; Eberl, L. Identification of Proteins Associated with the Pseudomonas aeruginosa Biofilm Extracellular Matrix. J. Proteome Res. 2012, 11, 4906–4915. [Google Scholar] [CrossRef]
- Carriquiriborde, F.; Martin Aispuro, P.; Ambrosis, N.; Zurita, E.; Bottero, D.; Gaillard, M.E.; Castuma, C.; Rudi, E.; Lodeiro, A.; Hozbor, D.F. Pertussis Vaccine Candidate Based on Outer Membrane Vesicles Derived From Biofilm Culture. Front. Immunol. 2021, 12, 730434. [Google Scholar] [CrossRef]
- Lindemann, S.; Allen, L.-A.H.; Welch, R.D.; Marcantonio, D.M.; Grande, R.; Di Marcantonio, M.C.; Robuffo, I.; Pompilio, A.; Celia, C.; Di Marzio, L.; et al. Helicobacter Pylori ATCC 43629/NCTC 11639 Outer Membrane Vesicles (OMVs) from Biofilm and Planktonic Phase Associated with Extracellular DNA (EDNA). Front. Microbiol. 2015, 6, 1369. [Google Scholar] [CrossRef]
- Schooling, S.R.; Beveridge, T.J. Membrane Vesicles: An Overlooked Component of the Matrices of Biofilms. J. Bacteriol. 2006, 188, 5945–5957. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Charpentier, L.A.; Dolben, E.F.; Hendricks, M.R.; Hogan, D.A.; Bomberger, J.M.; Stanton, B.A. Bacterial Outer Membrane Vesicles and Immune Modulation of the Host. Membranes 2023, 13, 752. https://doi.org/10.3390/membranes13090752
Charpentier LA, Dolben EF, Hendricks MR, Hogan DA, Bomberger JM, Stanton BA. Bacterial Outer Membrane Vesicles and Immune Modulation of the Host. Membranes. 2023; 13(9):752. https://doi.org/10.3390/membranes13090752
Chicago/Turabian StyleCharpentier, Lily A., Emily F. Dolben, Matthew R. Hendricks, Deborah A. Hogan, Jennifer M. Bomberger, and Bruce A. Stanton. 2023. "Bacterial Outer Membrane Vesicles and Immune Modulation of the Host" Membranes 13, no. 9: 752. https://doi.org/10.3390/membranes13090752
APA StyleCharpentier, L. A., Dolben, E. F., Hendricks, M. R., Hogan, D. A., Bomberger, J. M., & Stanton, B. A. (2023). Bacterial Outer Membrane Vesicles and Immune Modulation of the Host. Membranes, 13(9), 752. https://doi.org/10.3390/membranes13090752