


The Formation of β-Strand Nine (β9) in the Folding and Insertion of BamA from an Unfolded Form into Lipid Bilayers


Abstract
1. Introduction
2. Materials and Methods
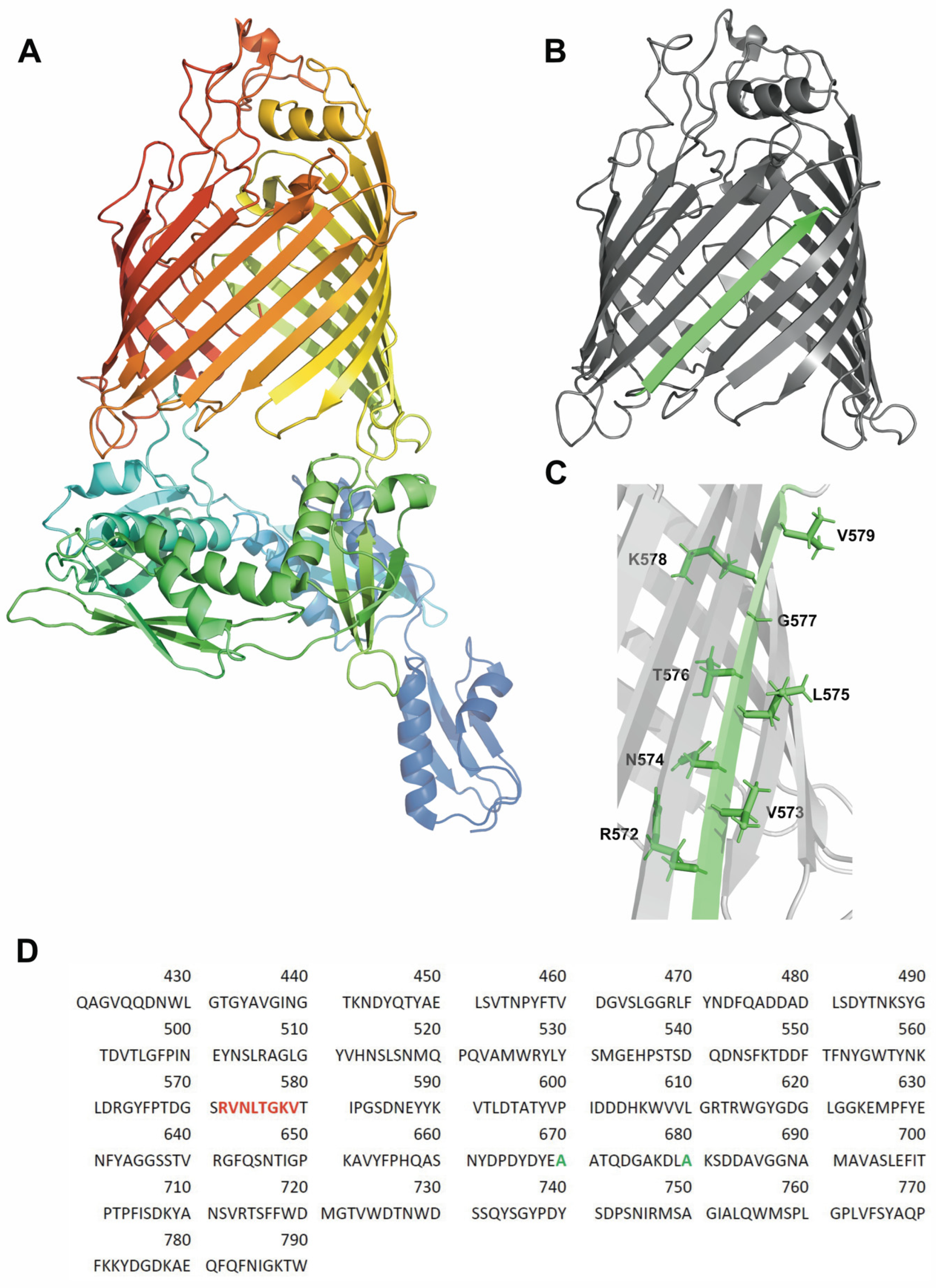
2.1. Single-Cysteine Mutants of BamA
2.2. Isolation of wt-BamA and XnC-BamA Mutants
2.3. Preparation of Lipid Bilayers
2.4. Labeling of Cysteine Residues
2.5. Folding of BamA and Trypsin Digestion
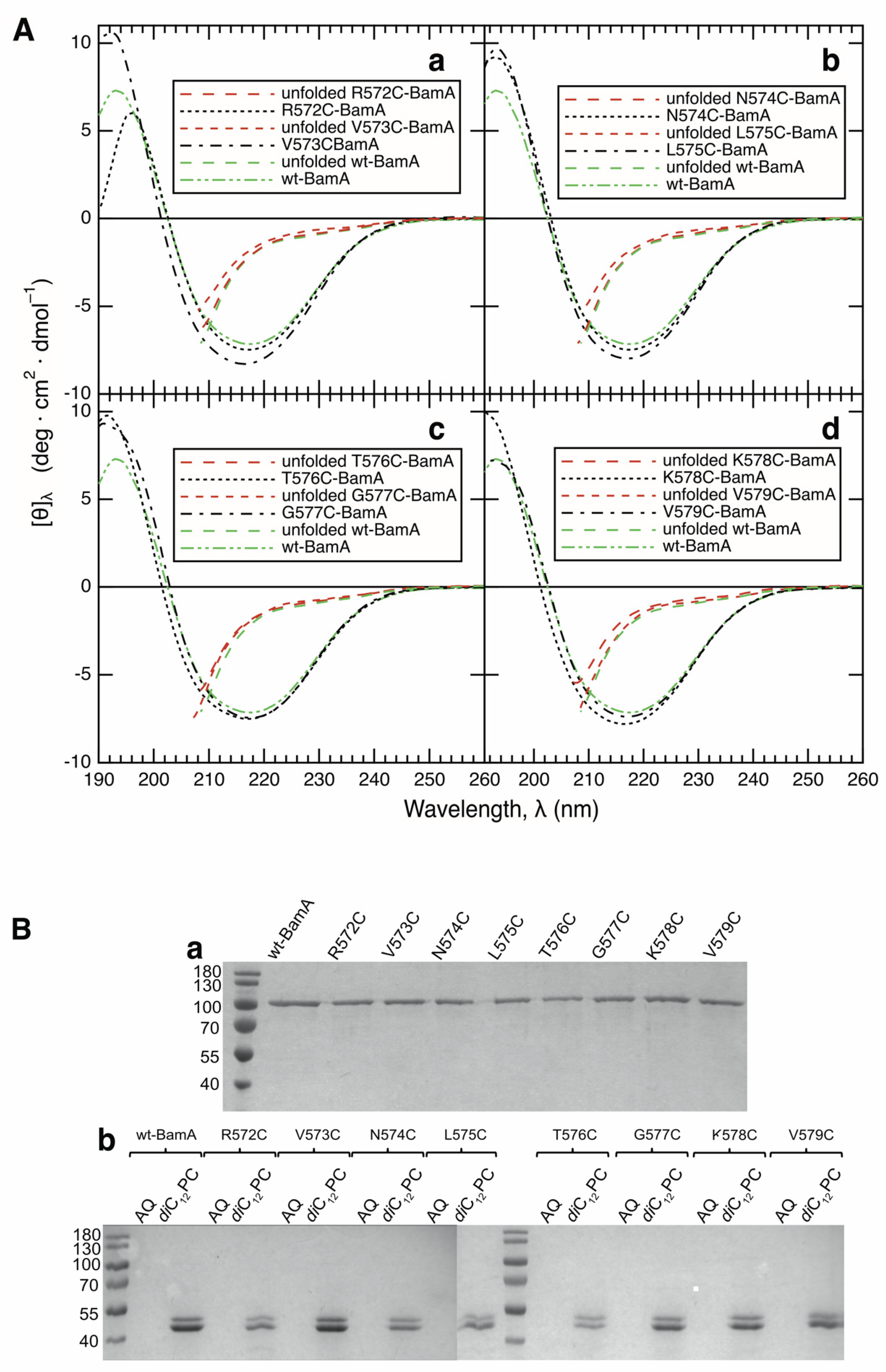
2.6. Circular Dichroism Spectroscopy
2.7. Fluorescence Spectroscopy
3. Results
3.1. All Examined Single-Cysteine Mutants of BamA Fold and Insert into Lipid Bilayers of diC12PC
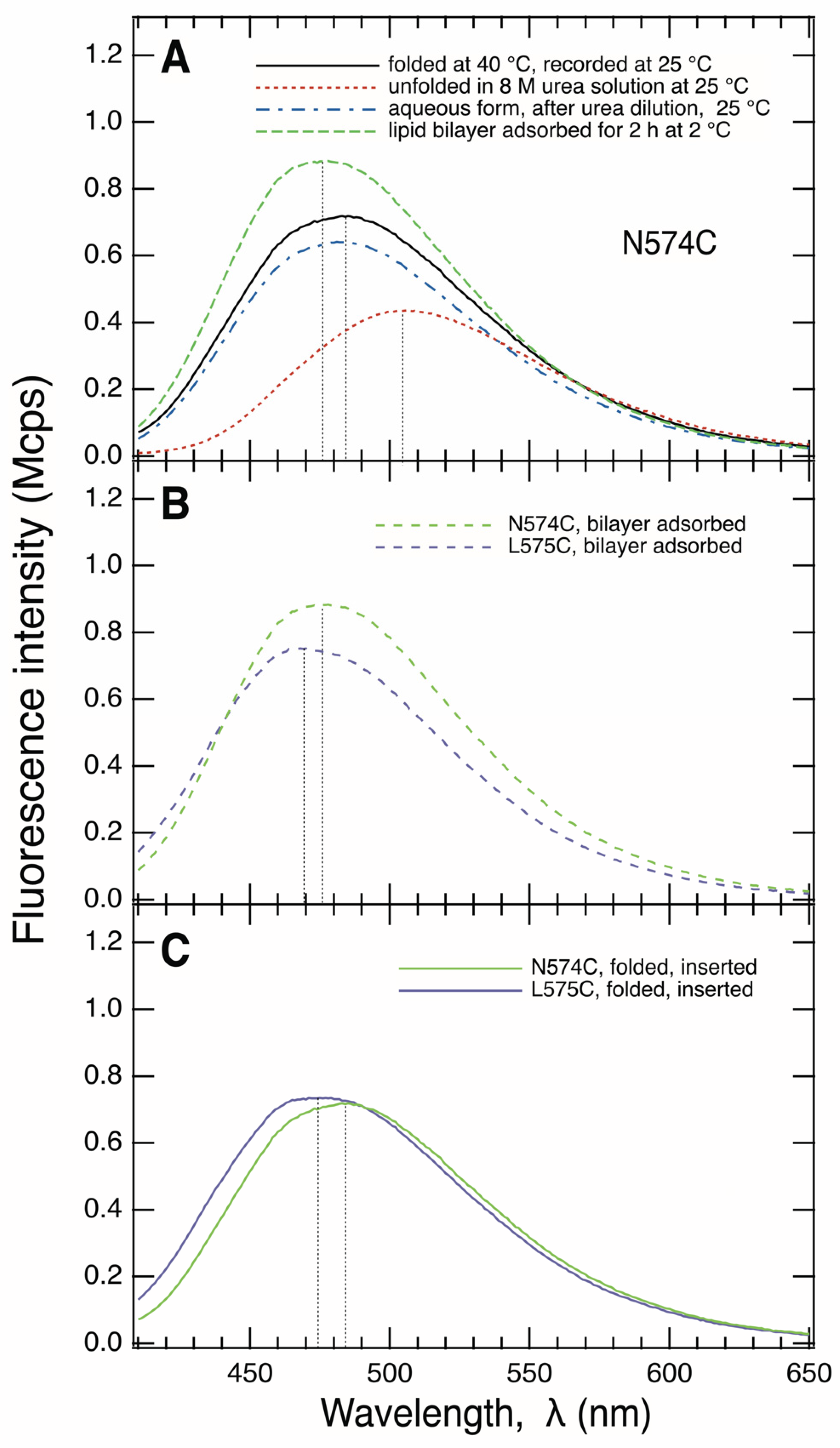
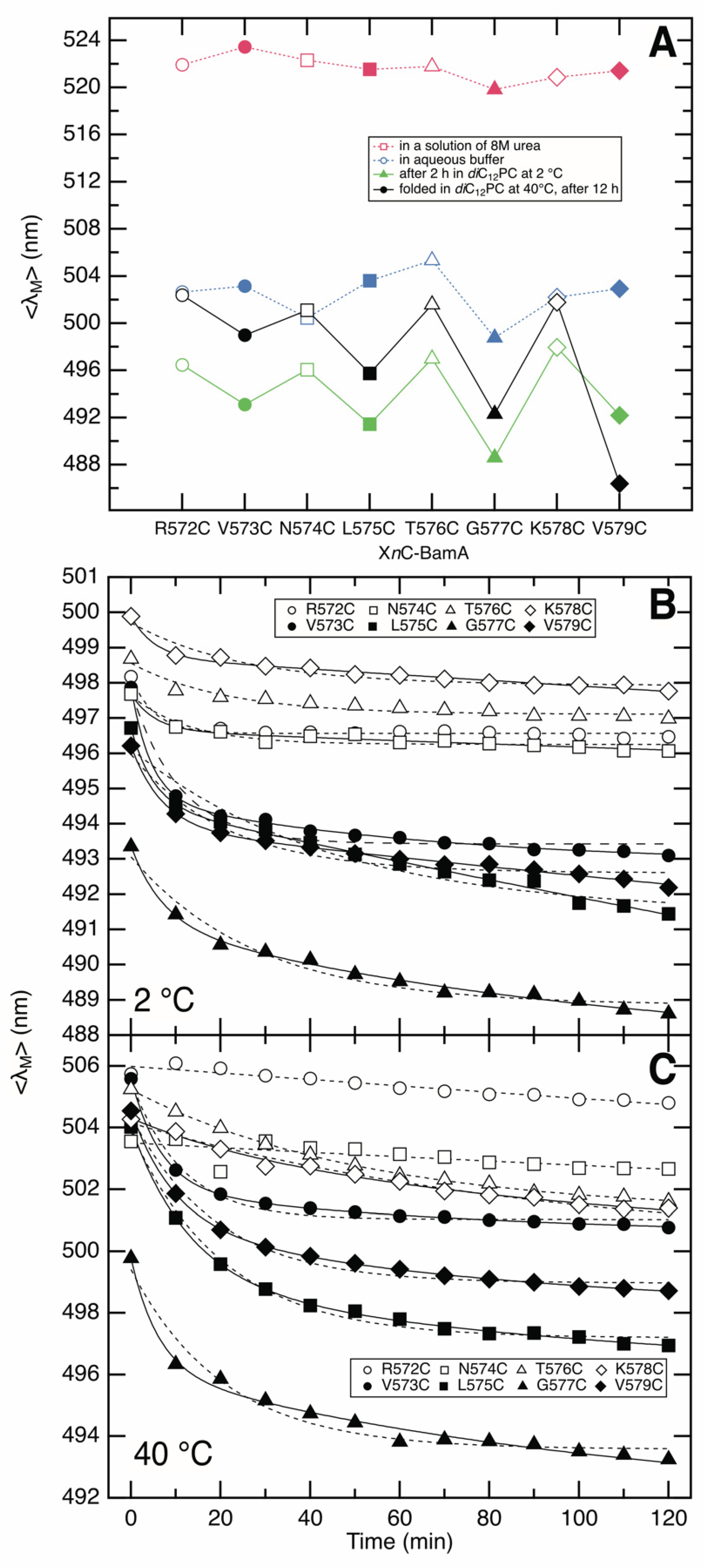
3.2. Fluorescence Spectra of IAEDANS-Labeled XnC-BamA Mutants Indicate BamA Folding
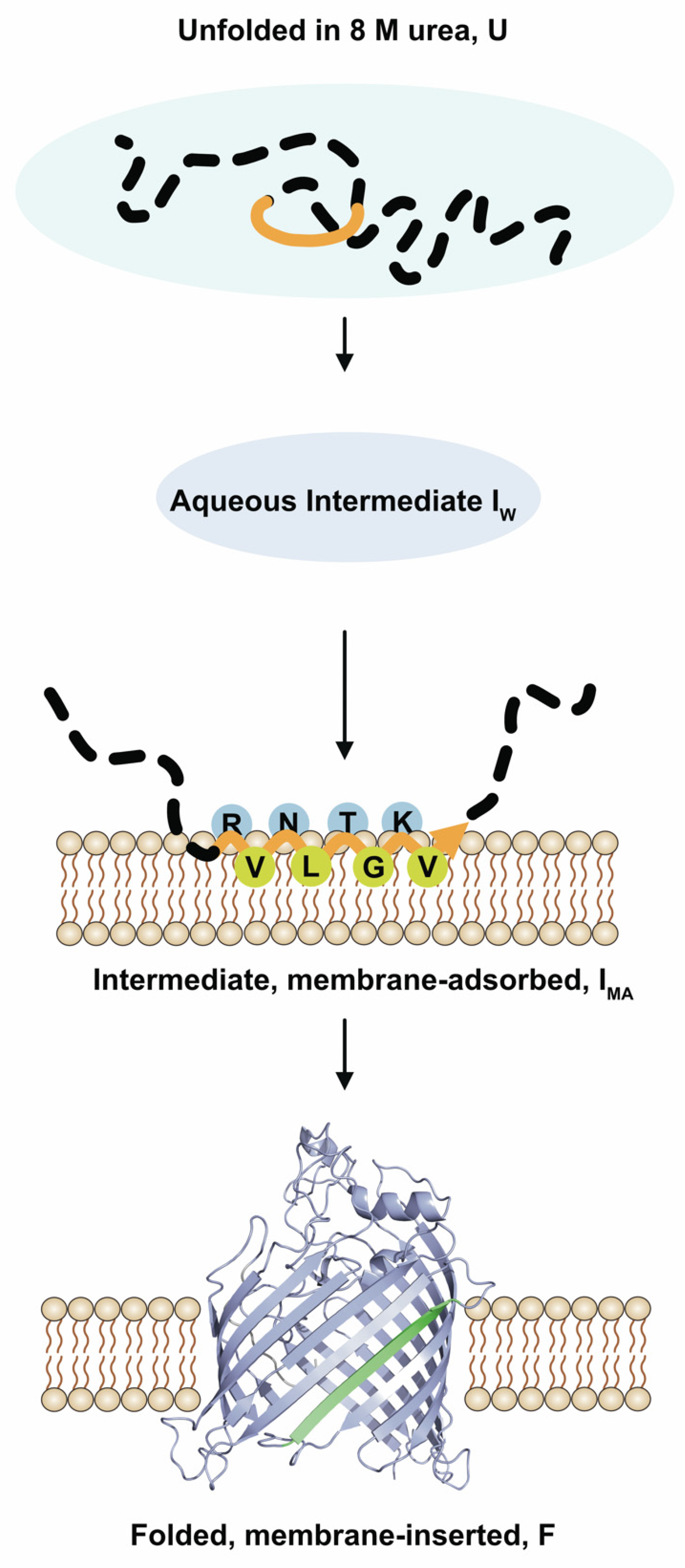
3.3. BamA Folds via a Membrane-Adsorbed Folding Intermediate
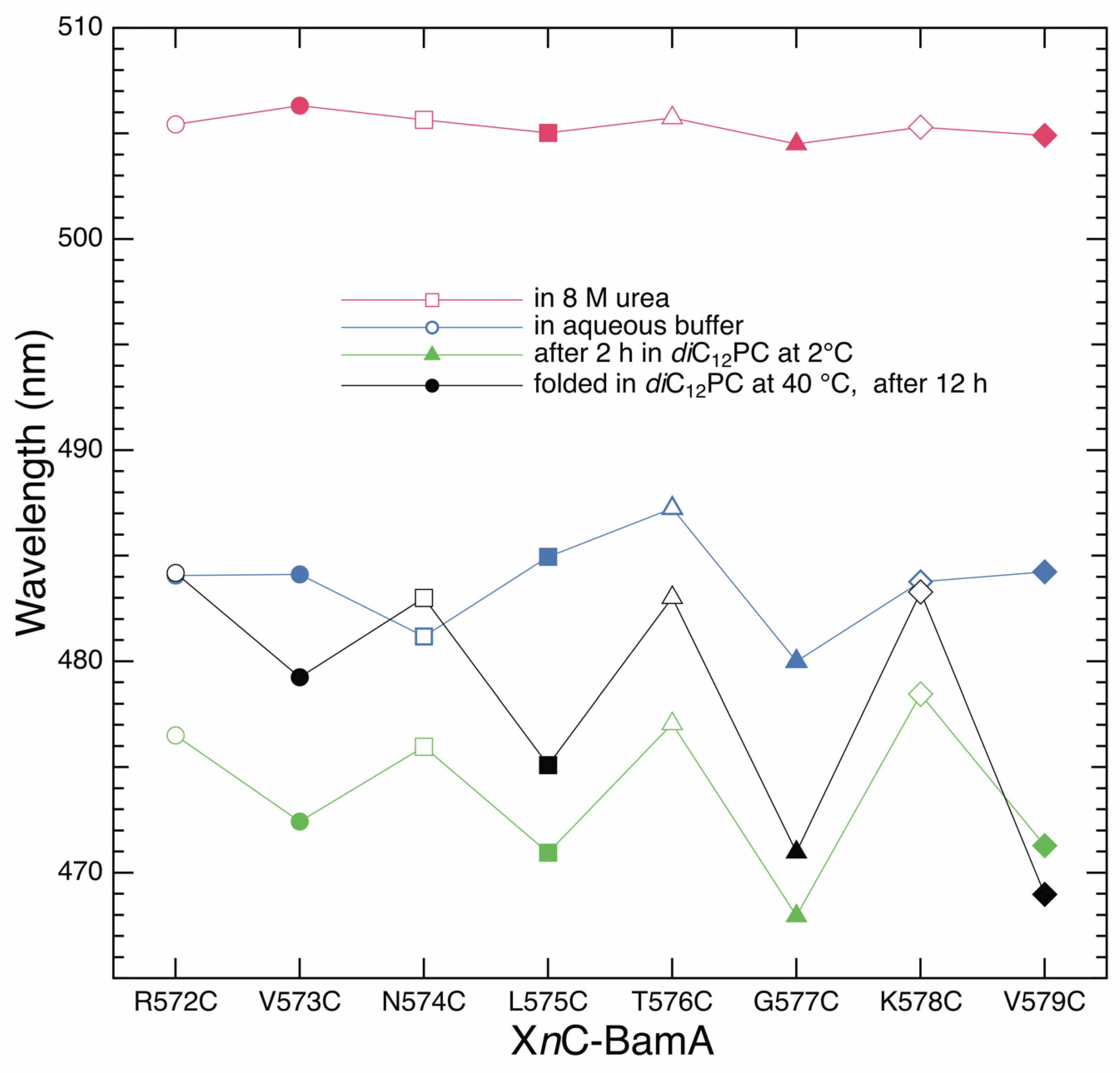
3.4. Strand β9 of BamA Forms Prior to Insertion upon Adsorption to the Lipid Bilayer
3.5. Strand β9 of BamA Forms Rapidly after Adsorption to the Lipid Bilayer
4. Discussion
4.1. BamA Folding in an Aqueous Environment
4.2. β-Strand Formation in Polypeptide Chains of Transmembrane β-Barrel Domains Precedes Membrane Insertion for Energetic Reasons
4.3. Not All β-Sheets of Transmembrane β-Barrels Form in the Polar/Apolar Interface
4.4. BamA Folds and Inserts Slower Than the Smaller OmpA and Does Not Catalyze Its Own Folding
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Patel, G.; Kleinschmidt, J.H. The lipid-bilayer inserted membrane protein BamA of Escherichia coli facilitates insertion and folding of outer membrane protein A from its complex with Skp. Biochemistry 2013, 52, 3974–3986. [Google Scholar] [CrossRef]
- Bos, M.P.; Robert, V.; Tommassen, J. Functioning of outer membrane protein assembly factor Omp85 requires a single POTRA domain. EMBO Rep. 2007, 8, 1149–1154. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Li, H.; Dong, H.; Zeng, Y.; Zhang, Z.; Paterson, N.G.; Stansfeld, P.J.; Wang, Z.; Zhang, Y.; Wang, W.; et al. Structural basis of outer membrane protein insertion by the BAM complex. Nature 2016, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Delano, W.L. The PyMOL Molecular Graphics System; DeLano Scientific: San Carlos, CA, USA, 2002. [Google Scholar]
- Schweizer, M.; Hindennach, I.; Garten, W.; Henning, U. Major proteins of the Escherichia coli outer cell envelope membrane. Interaction of protein II with lipopolysaccharide. Eur. J. Biochem. 1978, 82, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Eisele, J.L.; Rosenbusch, J.P. In vitro folding and oligomerization of a membrane protein. Transition of bacterial porin from random coil to native conformation. J. Biol. Chem. 1990, 265, 10217–10220. [Google Scholar] [CrossRef] [PubMed]
- Kleinschmidt, J.H.; Wiener, M.C.; Tamm, L.K. Outer membrane protein A of E. coli folds into detergent micelles, but not in the presence of monomeric detergent. Protein Sci. 1999, 8, 2065–2071. [Google Scholar] [CrossRef] [PubMed]
- Visudtiphole, V.; Thomas, M.B.; Chalton, D.A.; Lakey, J.H. Refolding of Escherichia coli outer membrane protein F in detergent creates LPS-free trimers and asymmetric dimers. Biochem. J. 2005, 392, 375–381. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Surrey, T.; Jähnig, F. Refolding and oriented insertion of a membrane protein into a lipid bilayer. Proc. Natl. Acad. Sci. USA 1992, 89, 7457–7461. [Google Scholar] [CrossRef]
- Pocanschi, C.L.; Dahmane, T.; Gohon, Y.; Rappaport, F.; Apell, H.-J.; Kleinschmidt, J.H.; Popot, J.-L. Amphipathic polymers: Tools to fold integral membrane proteins to their active form. Biochemistry 2006, 45, 13954–13961. [Google Scholar] [CrossRef]
- Horne, J.E.; Brockwell, D.J.; Radford, S.E. Role of the lipid bilayer in outer membrane protein folding in Gram-negative bacteria. J. Biol. Chem. 2020, 295, 10340–10367. [Google Scholar] [CrossRef]
- Kleinschmidt, J.H. Folding of β-barrel membrane proteins in lipid bilayers - unassisted and assisted folding and insertion. Biochim. Biophys. Acta 2015, 1848, 1927–1943. [Google Scholar] [CrossRef] [PubMed]
- Otzen, D.E.; Andersen, K.K. Folding of outer membrane proteins. Arch. Biochem. Biophys. 2013, 531, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Kleinschmidt, J.H.; Bulieris, P.V.; Qu, J.; Dogterom, M.; den Blaauwen, T. Association of neighboring β-strands of outer membrane protein A in lipid bilayers revealed by site directed fluorescence quenching. J. Mol. Biol. 2011, 407, 316–332. [Google Scholar] [CrossRef]
- Kleinschmidt, J.H.; Tamm, L.K. Secondary and tertiary structure formation of the β-barrel membrane protein OmpA is synchronized and depends on membrane thickness. J. Mol. Biol. 2002, 324, 319–330. [Google Scholar] [CrossRef]
- Kleinschmidt, J.H.; den Blaauwen, T.; Driessen, A.; Tamm, L.K. Outer membrane protein A of E. coli inserts and folds into lipid bilayers by a concerted mechanism. Biochemistry 1999, 38, 5006–5016. [Google Scholar] [CrossRef]
- Kleinschmidt, J.H.; Tamm, L.K. Folding intermediates of a β-barrel membrane protein. Kinetic evidence for a multi-step membrane insertion mechanism. Biochemistry 1996, 35, 12993–13000. [Google Scholar] [CrossRef][Green Version]
- Pocanschi, C.L.; Patel, G.J.; Marsh, D.; Kleinschmidt, J.H. Curvature Elasticity and Refolding of OmpA in Large Unilamellar Vesicles. Biophys. J. 2006, 91, L75–L78. [Google Scholar] [CrossRef][Green Version]
- Marsh, D.; Shanmugavadivu, B.; Kleinschmidt, J.H. Membrane elastic fluctuations and the insertion and tilt of β-barrel proteins. Biophys. J. 2006, 91, 227–232. [Google Scholar] [CrossRef]
- Asamoto, D.K.; Kang, G.; Kim, J.E. Folding of the β-Barrel Membrane Protein OmpA into Nanodiscs. Biophys. J. 2020, 118, 403–414. [Google Scholar] [CrossRef] [PubMed]
- Danoff, E.J.; Fleming, K.G. Novel Kinetic Intermediates Populated along the Folding Pathway of the Transmembrane β-Barrel OmpA. Biochemistry 2017, 56, 47–60. [Google Scholar] [CrossRef]
- Huysmans, G.H.; Radford, S.E.; Baldwin, S.A.; Brockwell, D.J. Malleability of the folding mechanism of the outer membrane protein PagP: Parallel pathways and the effect of membrane elasticity. J. Mol. Biol. 2012, 416, 453–464. [Google Scholar] [CrossRef][Green Version]
- Huysmans, G.H.; Radford, S.E.; Brockwell, D.J.; Baldwin, S.A. The N-terminal helix is a post-assembly clamp in the bacterial outer membrane protein PagP. J. Mol. Biol. 2007, 373, 529–540. [Google Scholar] [CrossRef]
- Huysmans, G.H.; Baldwin, S.A.; Brockwell, D.J.; Radford, S.E. The transition state for folding of an outer membrane protein. Proc. Natl. Acad. Sci. USA 2010, 107, 4099–4104. [Google Scholar] [CrossRef]
- Surrey, T.; Schmid, A.; Jähnig, F. Folding and membrane insertion of the trimeric β-barrel protein OmpF. Biochemistry 1996, 35, 2283–2288. [Google Scholar] [CrossRef]
- Conlan, S.; Bayley, H. Folding of a monomeric porin, OmpG, in detergent solution. Biochemistry 2003, 42, 9453–9465. [Google Scholar] [CrossRef]
- Pocanschi, C.L.; Apell, H.-J.; Puntervoll, P.; Høgh, B.T.; Jensen, H.B.; Welte, W.; Kleinschmidt, J.H. The major outer membrane protein of Fusobacterium nucleatum (FomA) folds and inserts into lipid bilayers via parallel folding pathways. J. Mol. Biol. 2006, 355, 548–561. [Google Scholar] [CrossRef] [PubMed]
- Shanmugavadivu, B.; Apell, H.J.; Meins, T.; Zeth, K.; Kleinschmidt, J.H. Correct folding of the β-barrel of the human membrane protein VDAC requires a lipid bilayer. J. Mol. Biol. 2007, 368, 66–78. [Google Scholar] [CrossRef] [PubMed]
- Burgess, N.K.; Dao, T.P.; Stanley, A.M.; Fleming, K.G. β-barrel proteins that reside in the Escherichia coli outer membrane in vivo demonstrate varied folding behavior in vitro. J. Biol. Chem. 2008, 283, 26748–26758. [Google Scholar] [CrossRef] [PubMed]
- Surrey, T.; Jähnig, F. Kinetics of folding and membrane insertion of a β-barrel membrane protein. J. Biol. Chem. 1995, 270, 28199–28203. [Google Scholar] [CrossRef]
- Gessmann, D.; Chung, Y.H.; Danoff, E.J.; Plummer, A.M.; Sandlin, C.W.; Zaccai, N.R.; Fleming, K.G. Outer membrane beta-barrel protein folding is physically controlled by periplasmic lipid head groups and BamA. Proc. Natl. Acad. Sci. USA 2014, 111, 5878–5883. [Google Scholar] [CrossRef]
- Rodionova, N.A.; Tatulian, S.A.; Surrey, T.; Jähnig, F.; Tamm, L.K. Characterization of two membrane-bound forms of OmpA. Biochemistry 1995, 34, 1921–1929. [Google Scholar] [CrossRef]
- Kleinschmidt, J.H.; Tamm, L.K. Time-resolved distance determination by tryptophan fluorescence quenching: Probing intermediates in membrane protein folding. Biochemistry 1999, 38, 4996–5005. [Google Scholar] [CrossRef]
- Prilipov, A.; Phale, P.S.; Van Gelder, P.; Rosenbusch, J.P.; Koebnik, R. Coupling site-directed mutagenesis with high-level expression: Large scale production of mutant porins from E. coli. FEMS Microbiol. Lett. 1998, 163, 65–72. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Lapinski, M.M.; Castro-Forero, A.; Greiner, A.J.; Ofoli, R.Y.; Blanchard, G.J. Comparison of liposomes formed by sonication and extrusion: Rotational and translational diffusion of an embedded chromophore. Langmuir 2007, 23, 11677–11683. [Google Scholar] [CrossRef]
- Qiu, C.; Blanchard, G.J. Phospholipid vesicle stability and temporal variations in acyl chain organization. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2013, 110, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, L.; Gholami, O.; Schürmann, N.; Kleinschmidt, J.H. Folding of β-Barrel Membrane Proteins into Lipid Membranes by Site-Directed Fluorescence Spectroscopy. Methods Mol. Biol. 2019, 2003, 465–492. [Google Scholar] [CrossRef] [PubMed]
- Owenius, R.; Österlund, M.; Lindgren, M.; Svensson, M.; Olsen, O.H.; Persson, E.; Freskgård, P.O.; Carlsson, U. Properties of spin and fluorescent labels at a receptor-ligand interface. Biophys. J. 1999, 77, 2237–2250. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Riddles, P.W.; Blakeley, R.L.; Zerner, B. Reassessment of Ellman’s reagent. Methods Enzymol. 1983, 91, 49–60. [Google Scholar] [CrossRef]
- Weber, K.; Osborne, M. The reliability of molecular weight determinations by dodecyl sulfate-polyacrylamide gel electrophoresis. J. Biol. Chem. 1964, 244, 4406–4412. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Provencher, S.W.; Glöckner, J. Estimation of globular protein secondary structure from circular dichroism. Biochemistry 1981, 20, 33–37. [Google Scholar] [CrossRef]
- Compton, L.A.; Johnson, W.C., Jr. Analysis of protein circular dichroism spectra for secondary structure using a simple matrix multiplication. Anal. Biochem. 1986, 155, 155–167. [Google Scholar] [CrossRef]
- Whitmore, L.; Wallace, B.A. DICHROWEB, an online server for protein secondary structure analyses from circular dichroism spectroscopic data. Nucleic Acids Res. 2004, 32, W668–W673. [Google Scholar] [CrossRef]
- Kim, S.; Malinverni, J.C.; Sliz, P.; Silhavy, T.J.; Harrison, S.C.; Kahne, D. Structure and function of an essential component of the outer membrane protein assembly machine. Science 2007, 317, 961–964. [Google Scholar] [CrossRef] [PubMed]
- Gatzeva-Topalova, P.Z.; Walton, T.A.; Sousa, M.C. Crystal structure of YaeT: Conformational flexibility and substrate recognition. Structure 2008, 16, 1873–1881. [Google Scholar] [CrossRef]
- Pocanschi, C.L.; Kleinschmidt, J.H. The Thermodynamic Stability of Membrane Proteins in Micelles and Lipid Bilayers Investigated with the Ferrichrom Receptor FhuA. J. Membrane Biol. 2022, 255, 485–502. [Google Scholar] [CrossRef]
- Frillingos, S.; Sahin-Toth, M.; Wu, J.; Kaback, H.R. Cys-scanning mutagenesis: A novel approach to structure function relationships in polytopic membrane proteins. FASEB J. 1998, 12, 1281–1299. [Google Scholar] [CrossRef]
- Nesper, J.; Brosig, A.; Ringler, P.; Patel, G.; Müller, S.; Kleinschmidt, J.H.; Boos, W.; Diederichs, K.; Welte, W. TtOmp85 from Thermus thermophilus HB 27: An ancestral type of the Omp85 protein family. J. Bacteriol. 2008, 190, 4568–4575. [Google Scholar] [CrossRef] [PubMed]
- Bakelar, J.; Buchanan, S.K.; Noinaj, N. The structure of the β-barrel assembly machinery complex. Science 2016, 351, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Zheng, J.; Wang, Y.; Yang, X.; Liu, Y.; Sun, C.; Cao, B.; Zhou, H.; Ni, D.; Lou, J.; et al. Structure of the BAM complex and its implications for biogenesis of outer-membrane proteins. Nat. Struct. Mol. Biol. 2016. [Google Scholar] [CrossRef]
- Hammarström, P.; Owenius, R.; Mårtensson, L.G.; Carlsson, U.; Lindgren, M. High-resolution probing of local conformational changes in proteins by the use of multiple labeling: Unfolding and self-assembly of human carbonic anhydrase II monitored by spin, fluorescent, and chemical reactivity probes. Biophys. J. 2001, 80, 2867–2885. [Google Scholar] [CrossRef] [PubMed]
- Schiffrin, B.; Brockwell, D.J.; Radford, S.E. Outer membrane protein folding from an energy landscape perspective. BMC Biol. 2017, 15, 123. [Google Scholar] [CrossRef]
- Horne, J.E.; Radford, S.E. A growing toolbox of techniques for studying β-barrel outer membrane protein folding and biogenesis. Biochem. Soc. Trans. 2016, 44, 802–809. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kleinschmidt, J.H. Folding kinetics of the outer membrane proteins OmpA and FomA into phospholipid bilayers. Chem. Phys. Lipids 2006, 141, 30–47. [Google Scholar] [CrossRef] [PubMed]
- Royer, C.A.; Mann, C.J.; Matthews, C.R. Resolution of the fluorescence equilibrium unfolding profile of trp aporepressor using single tryptophan mutants. Protein Sci. 1993, 2, 1844–1852. [Google Scholar] [CrossRef]
- Roumestand, C.; Boyer, M.; Guignard, L.; Barthe, P.; Royer, C.A. Characterization of the Folding and Unfolding Reactions of a Small β-barrel Protein of Novel Topology, the MTCP1 Oncogene Product P13. J. Mol. Biol. 2001, 312, 247–259. [Google Scholar] [CrossRef]
- Asamoto, D.K.; Kozachenko, I.A.; Lopez-Pena, I.; Kim, J.E. Bimolecular quenching of tryptophan fluorescence in a membrane protein: Evolution of local solvation and environment during folding into a bilayer. Spectrochim. Acta. Part A Mol. Biomol. Spectrosc. 2021, 260, 119919. [Google Scholar] [CrossRef]
- Qu, J.; Mayer, C.; Behrens, S.; Holst, O.; Kleinschmidt, J.H. The trimeric periplasmic chaperone Skp of Escherichia coli forms 1:1 complexes with outer membrane proteins via hydrophobic and electrostatic interactions. J. Mol. Biol. 2007, 374, 91–105. [Google Scholar] [CrossRef]
- McMorran, L.M.; Bartlett, A.I.; Huysmans, G.H.; Radford, S.E.; Brockwell, D.J. Dissecting the effects of periplasmic chaperones on the in vitro folding of the outer membrane protein PagP. J. Mol. Biol. 2013, 425, 3178–3191. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wimley, W.C.; White, S.H. Experimentally determined hydrophobicity scale for proteins at membrane interfaces. Nat. Struct. Biol. 1996, 3, 842–848. [Google Scholar] [CrossRef] [PubMed]
- Roseman, M.A. Hydrophobicity of the peptide C=O...H-N hydrogen-bonded group. J. Mol. Biol. 1988, 201, 621–623. [Google Scholar] [CrossRef] [PubMed]
- White, S.H.; Wimley, W.C. Membrane protein folding and stability: Physical principles. Annu. Rev. Biophys. Biomol. Struct. 1999, 28, 319–365. [Google Scholar] [CrossRef] [PubMed]
- Wimley, W.C.; Hristova, K.; Ladokhin, A.S.; Silvestro, L.; Axelsen, P.H.; White, S.H. Folding of β-sheet membrane proteins: A hydrophobic hexapeptide model. J. Mol. Biol. 1998, 277, 1091–1110. [Google Scholar] [CrossRef]
- Lewis, B.A.; Engelman, D.M. Lipid bilayer thickness varies linearly with acyl chain length in fluid phosphatidylcholine vesicles. J. Mol. Biol. 1983, 166, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Anbazhagan, V.; Vijay, N.; Kleinschmidt, J.H.; Marsh, D. Protein-Lipid Interactions with Fusobacterium nucleatum Major Outer Membrane Protein FomA: Spin-Label EPR and Polarized Infrared Spectroscopy. Biochemistry 2008, 47, 8414–8423. [Google Scholar] [CrossRef] [PubMed]
- Puntervoll, P.; Ruud, M.; Bruseth, L.J.; Kleivdal, H.; Høgh, B.T.; Benz, R.; Jensen, H.B. Structural characterization of the fusobacterial non-specific porin FomA suggests a 14-stranded topology, unlike the classical porins. Microbiology 2002, 148, 3395–3403. [Google Scholar] [CrossRef]
- Hagan, C.L.; Westwood, D.B.; Kahne, D. Bam Lipoproteins Assemble BamA in Vitro. Biochemistry 2013, 52, 6108–6113. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (A) 2 °C | |||||
| Mutant a | <λ0> | Afc | kfb (min−1) | Afd | ksb (min−1) |
| R572C | 496 ± 0.0 | 1.61 ± 0.07 | 0.19 ± 0.03 | - | - |
| V573C | 492.8 ± 0.2 | 3.1 ± 0.1 | 0.23 ± 0.03 | 1.9 ± 0.1 | 0.016 ± 0.004 |
| N574C | 494.0 ± 19 | 1.0 ± 0.2 | 0.19 ± 0.11 | 2 ± 19 | 0.002 ± 0.02 |
| L575C | 484 ± 8 | 2.0 ± 0.2 | 0.29 ± 0.16 | 10 ± 7 | 0.003 ± 0.003 |
| T576C | 497.1 ± 0.5 | - | - | 1.4 ± 0.1 | |
| G577C | 488 ± 1 | 2.1 ± 0.4 | 0.14 ± 0.04 | 3.7 ± 0.7 | 0.010 ± 0.006 |
| K578C | 482 ± 238 | 1.16 ± 0.17 | 0.18 ± 0.08 | 16 ± 238 | 0.0005 ± 0.007 |
| V579C | 485 ± 17 | 2.3 ± 0.2 | 0.14 ± 0.02 | 8 ± 16 | 0.002 ± 0.004 |
| (B) 40 °C | |||||
| Mutant a | <λ0> | Afc | kfb (min−1) | Afd | ksb (min−1) |
| R572C | 500.1 | - | - | 5.90 ± 0.06 | 0.0019 ± 0.0002 |
| V573C | 500.5 ± 0.1 | 3.58 ± 0.09 | 0.148 ± 0.006 | 1.50 ± 0.05 | 0.013 ± 0.003 |
| N574C | 499.1 | - | - | 4.4 ± 0.2 | 0.0017 ± 0.0005 |
| L575C | 495.9 ± 1.2 | 5.0 ± 0.7 | 0.076 ± 0.009 | 3.0 ± 0.6 | 0.009 ± 0.008 |
| T576C | 501.2 | - | - | 3.99 ± 0.03 | 0.0191 ± 0.0003 |
| G577C | 492.0 ± 1.4 | 3.4 ± 0.7 | 0.18 ± 0.07 | 4.4 ± 0.9 | 0.011 ± 0.008 |
| K578C | 500.8 | - | - | 3.43 ± 0.07 | 0.0157 ± 0.0006 |
| V579C | 498.1 ± 0.2 | 3.7 ± 0.2 | 0.102 ± 0.005 | 2.74 ± 0.07 | 0.013 ± 0.002 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herwig, S.; Kleinschmidt, J.H. The Formation of β-Strand Nine (β9) in the Folding and Insertion of BamA from an Unfolded Form into Lipid Bilayers. Membranes 2023, 13, 247. https://doi.org/10.3390/membranes13020247
Herwig S, Kleinschmidt JH. The Formation of β-Strand Nine (β9) in the Folding and Insertion of BamA from an Unfolded Form into Lipid Bilayers. Membranes. 2023; 13(2):247. https://doi.org/10.3390/membranes13020247
Chicago/Turabian StyleHerwig, Sascha, and Jörg H. Kleinschmidt. 2023. "The Formation of β-Strand Nine (β9) in the Folding and Insertion of BamA from an Unfolded Form into Lipid Bilayers" Membranes 13, no. 2: 247. https://doi.org/10.3390/membranes13020247
APA StyleHerwig, S., & Kleinschmidt, J. H. (2023). The Formation of β-Strand Nine (β9) in the Folding and Insertion of BamA from an Unfolded Form into Lipid Bilayers. Membranes, 13(2), 247. https://doi.org/10.3390/membranes13020247