

Effects of a Pseudomonas Strain on the Lipid Transfer Proteins, Appoplast Barriers and Activity of Aquaporins Associated with Hydraulic Conductance of Pea Plants
 , , ,
, , ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strain and Cultural Media
2.2. Plant Growth Conditions and Treatments
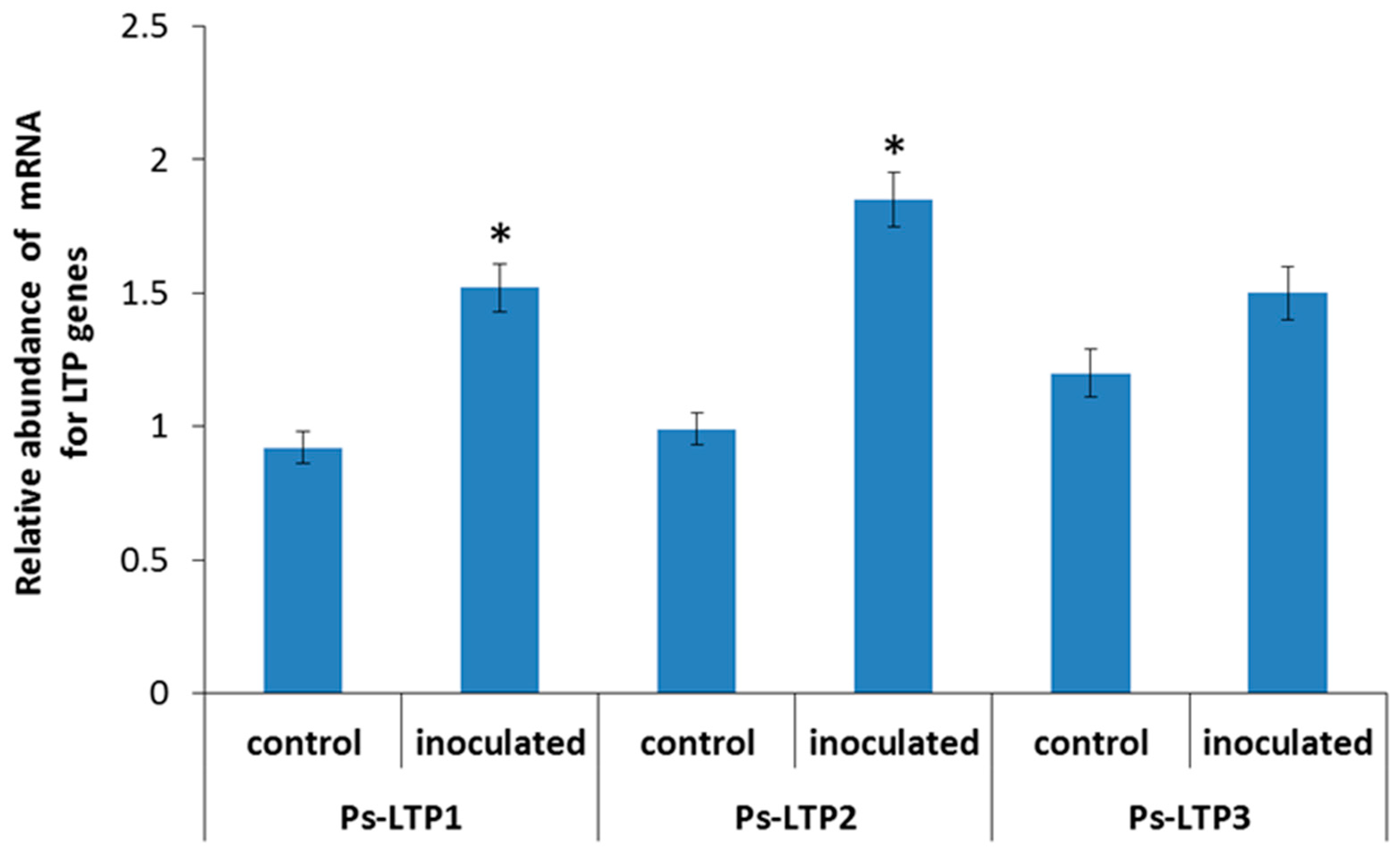
2.3. RNA Extraction and Analysis of the Abundance of LTP mRNA
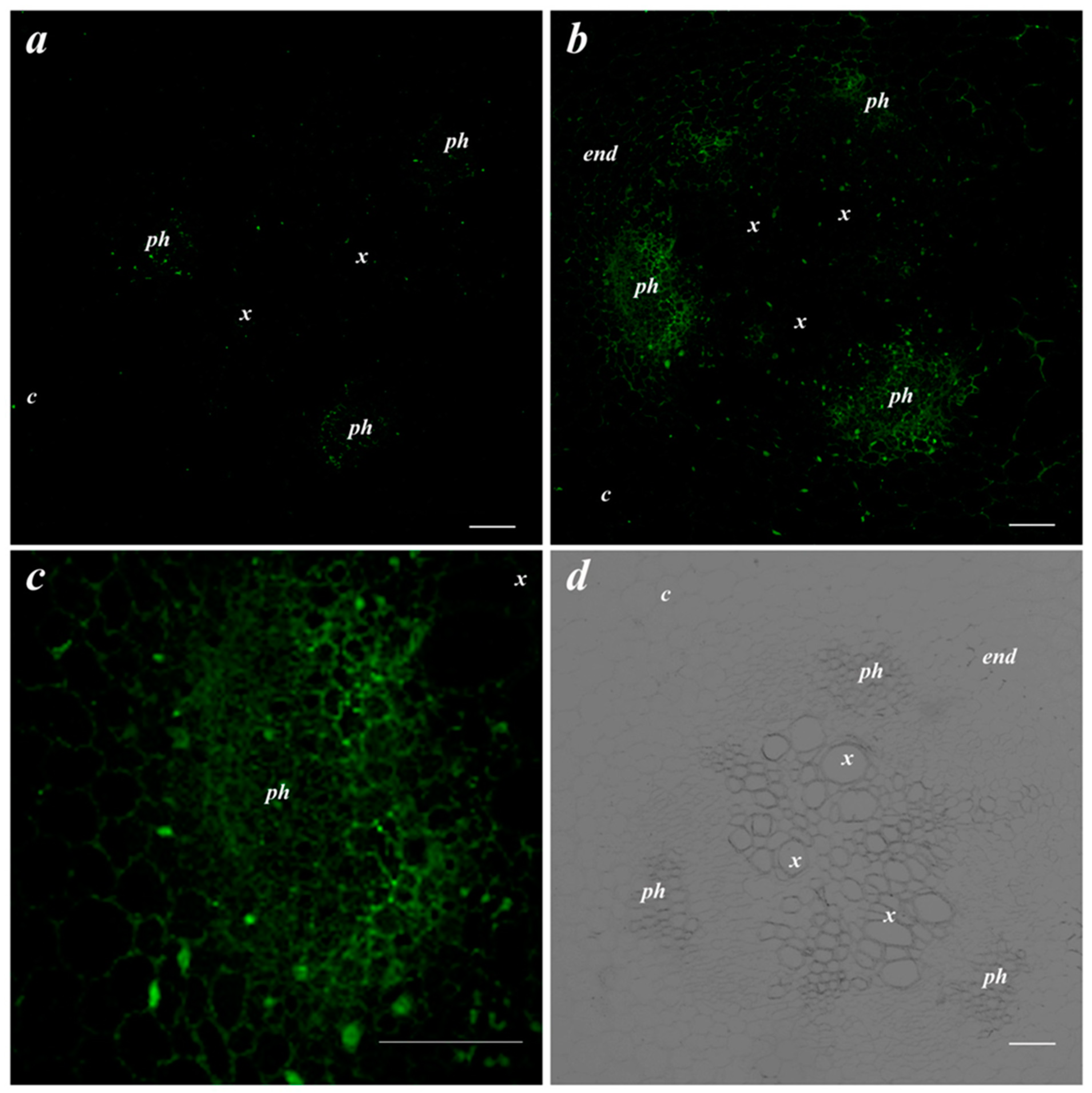
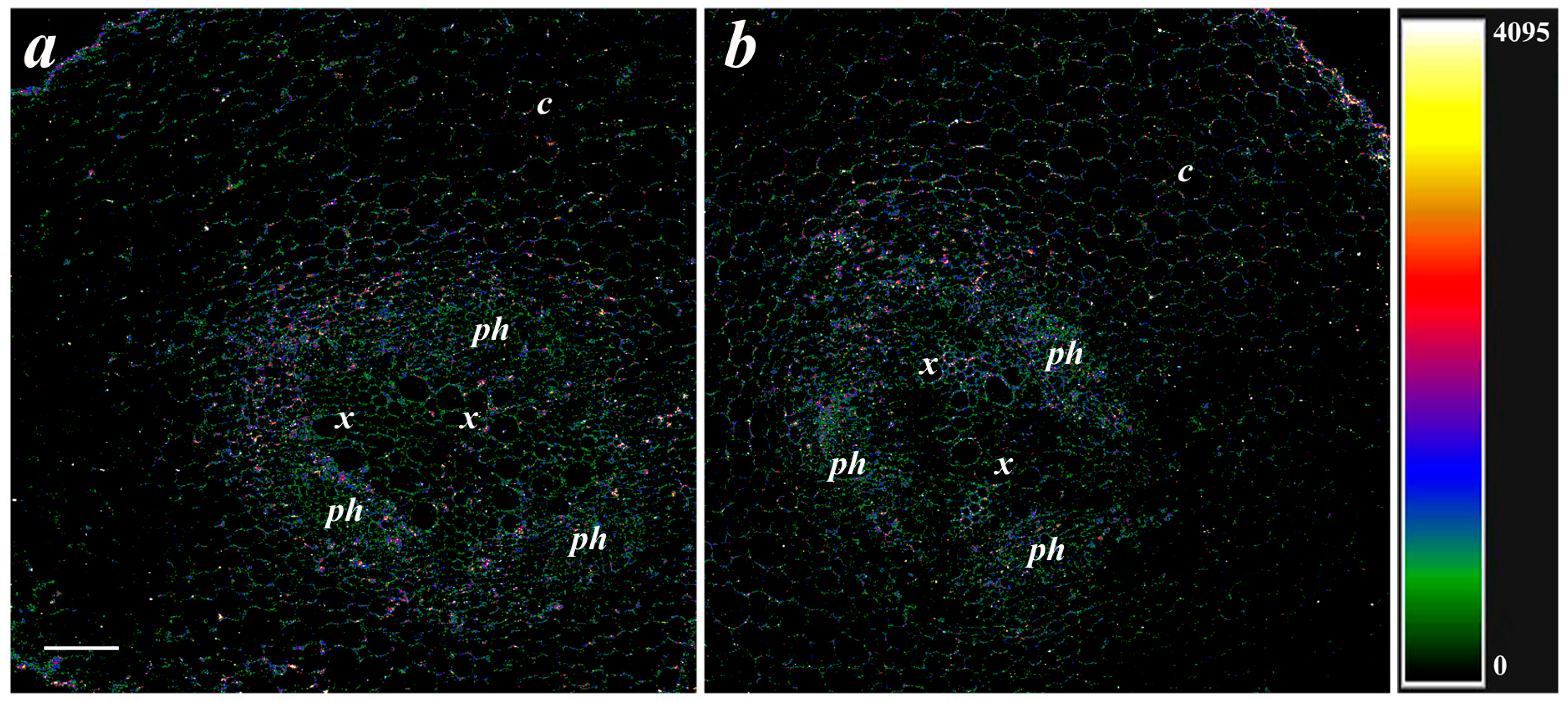
2.4. Immunolocalization of LTPs and the ABA
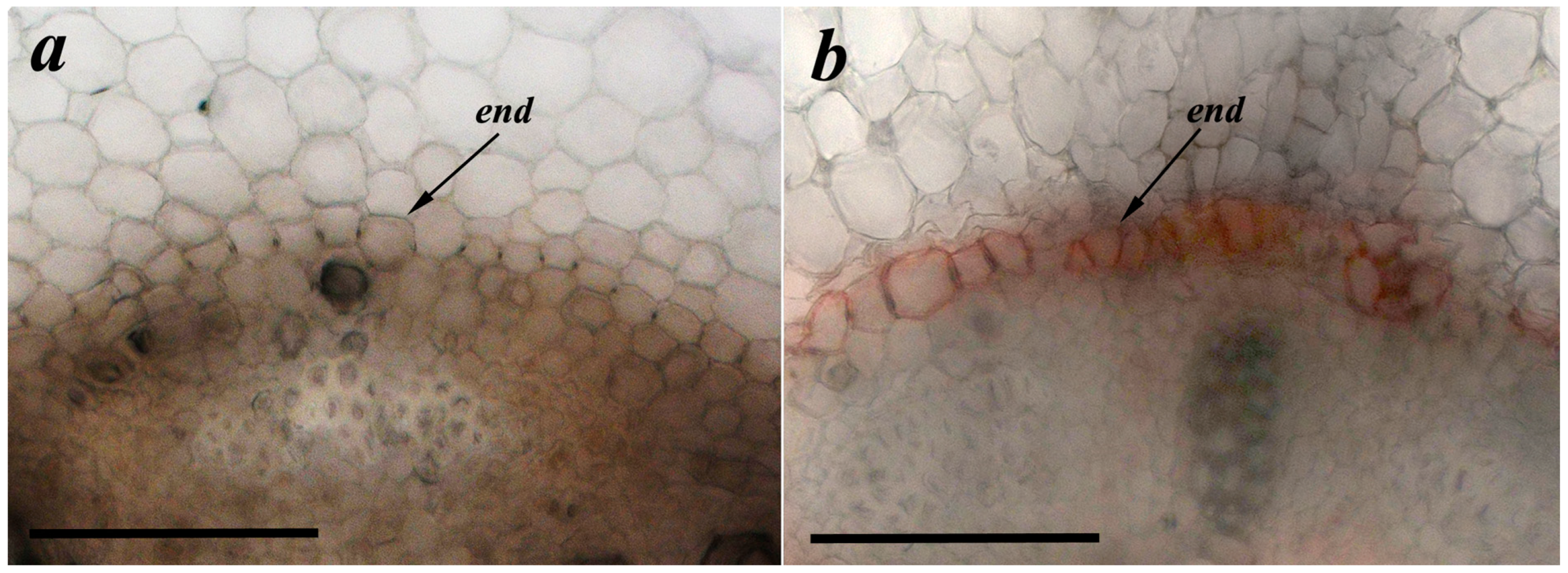
2.5. Suberin Detection
2.6. Parameters of Water Relations
2.7. Statistics
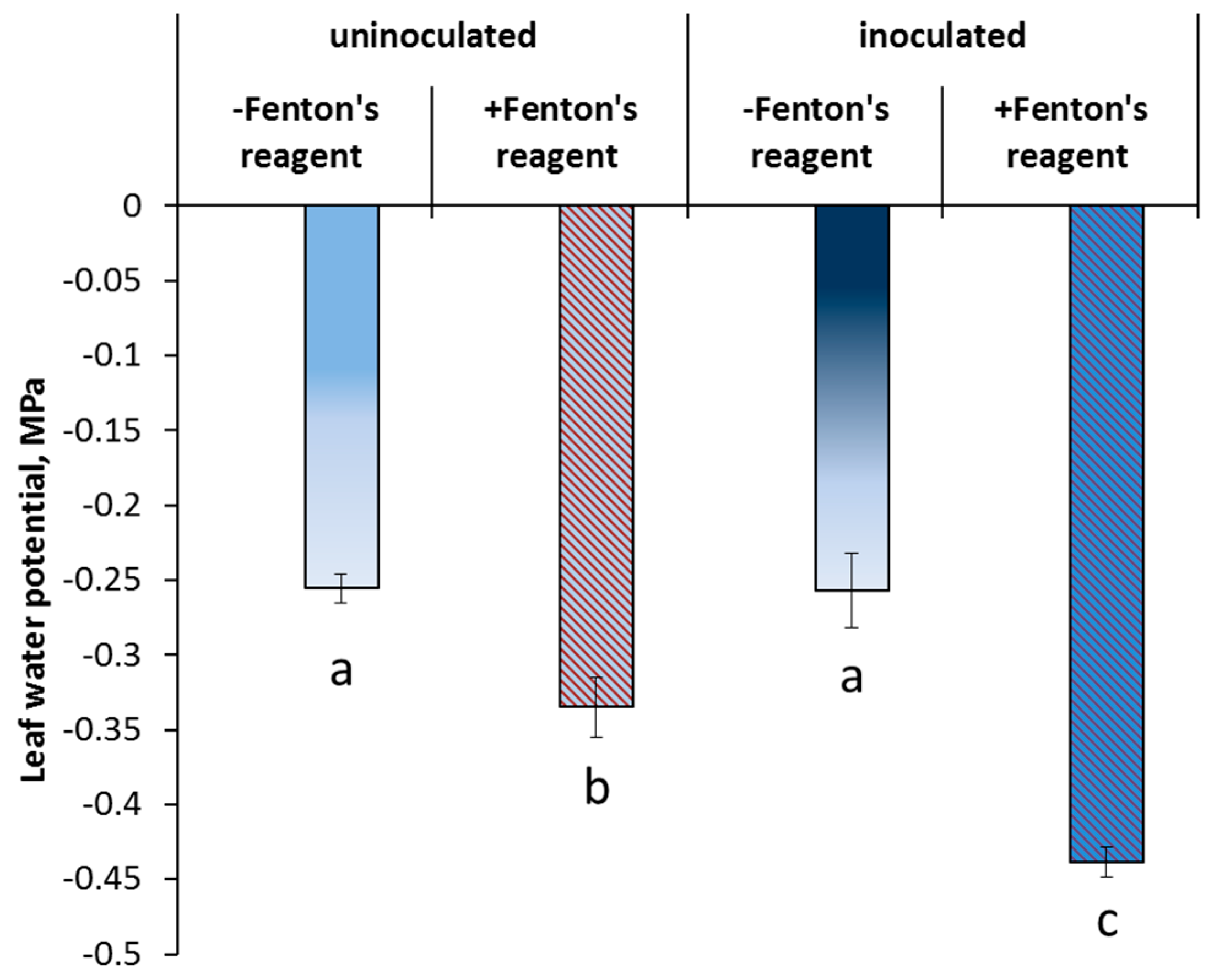
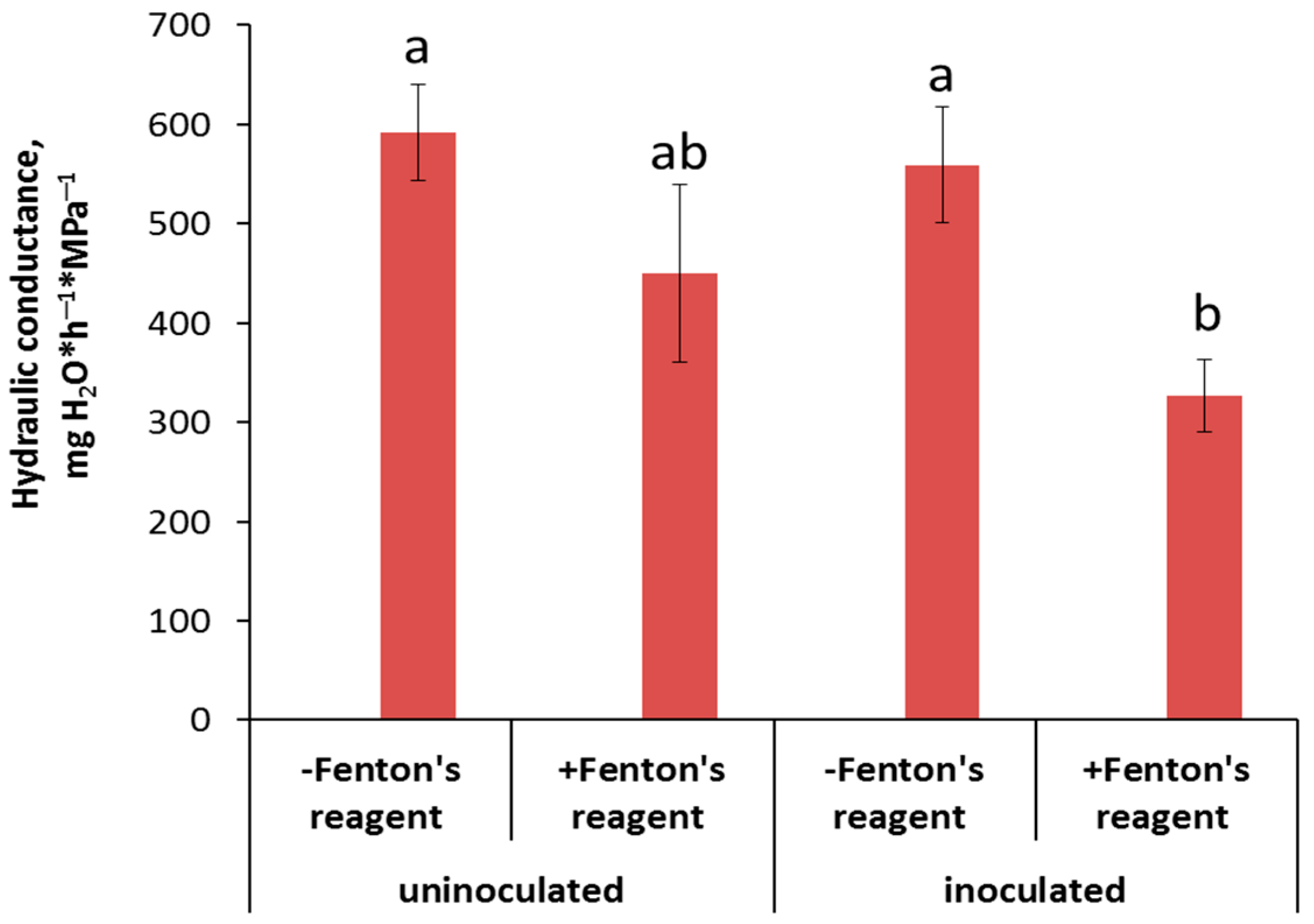
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kudoyarova, G.; Arkhipova, T.; Korshunova, T.; Bakaeva, M.; Loginov, O.; Dodd, I.C. Phytohormone mediation of interactions between plants and non-symbiotic growth promoting bacteria under edaphic stresses. Front. Plant Sci. 2019, 10, 1368. [Google Scholar] [CrossRef]
- Ruzzi, M.; Aroca, R. Plant growth-promoting rhizobacteria act as biostimulants in horticulture. Sci. Hortic. 2015, 196, 124–134. [Google Scholar] [CrossRef]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant growth- promoting rhizobacteria: Context, mechanisms of action, and roadmap to commercialization of biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9, 1473. [Google Scholar] [CrossRef]
- Petrillo, C.; Vitale, E.; Ambrosino, P.; Arena, C.; Isticato, R. Plant growth-promoting bacterial consortia as a strategy to alleviate drought stress in Spinacia oleracea. Microorganisms 2022, 10, 1798. [Google Scholar] [CrossRef]
- Boursiac, Y.; Protto, V.; Rishmawi, L.; Maurel, C. Experimental and conceptual approaches to root water transport. Plant Soil 2022, 478, 349–370. [Google Scholar] [CrossRef]
- Cui, B.; Liu, R.; Flowers, T.J.; Song, J. Casparian bands and suberin lamellae: Key targets for breeding salt tolerant crops? Environ. Exp. Bot. 2021, 191, 104600. [Google Scholar] [CrossRef]
- Lee, M.-H.; Jeon, H.S.; Kim, S.H.; Chung, J.H.; Roppolo, D.; Lee, H.-J.; Cho, H.J.; Tobimatsu, Y.; Ralph, J.; Park, O.K. Lignin based barrier restricts pathogens to the infection site and confers resistance in plants. EMBO J. 2019, 38, e101948. [Google Scholar] [CrossRef]
- Akhiyarova, G.R.; Finkina, E.I.; Ovchinnikova, T.N.; Veselov, D.S.; Kudoyarova, G.R. Role of pea LTPs and abscisic acid in salt-stressed roots. Biomolecules 2020, 10, 15. [Google Scholar] [CrossRef]
- Edqvist, J.; Blomqvist, K.; Nieuwland, J.; Salminen, T.A. Plant lipid transfer proteins: Are we finally closing in on the roles of these enigmatic proteins? J. Lipid Res. 2018, 59, 1374–1380. [Google Scholar] [CrossRef]
- Finkina, E.I.; Melnikova, D.N.; Bogdanov, I.V.; Ovchinnikova, T.V. Lipid transfer proteins as components of the plant innate immune system: Structure, functions, and applications. Acta Nat. 2016, 8, 47–61. [Google Scholar] [CrossRef]
- Gao, H.; Ma, K.; Ji, G.; Pan, L.; Zhou, Q. Lipid transfer proteins involved in plant–pathogen interactions and their molecular mechanisms. Mol. Plant Pathol. 2022, 23, 1815–1829. [Google Scholar] [CrossRef]
- Martynenko, E.; Arkhipova, T.; Safronova, V.; Seldimirova, O.; Galin, I.; Akhtyamova, Z.; Veselov, D.; Ivanov, R.; Kudoyarova, G. Effects of phytohormone-producing rhizobacteria on casparian band formation, ion homeostasis and salt tolerance of durum wheat. Biomolecules 2022, 12, 230. [Google Scholar] [CrossRef]
- Wang, P.; Wang, C.M.; Gao, L.; Cui, Y.-N.; Yang, H.-L.; De Silva, N.D.G.; Ma, Q.; Bao, A.-K.; Flowers, T.J.; Rowland, O.; et al. Aliphatic suberin confers salt tolerance to Arabidopsis by limiting Na+ influx, K+ efflux and water backflow. Plant Soil 2020, 448, 603–620. [Google Scholar] [CrossRef]
- Li, L.; Pan, S.; Melzer, R.; Fricke, W. Apoplastic barriers, aquaporin gene expression and root and cell hydraulic conductivity in phosphate-limited sheepgrass plants. Physiol Plant. 2020, 168, 118–132. [Google Scholar] [CrossRef]
- Flexas, J.; Carriquí, M.; Nadal, M. Gas exchange and hydraulics during drought in crops: Who drives whom? J. Exp. Bot. 2018, 69, 3791–3795. [Google Scholar] [CrossRef]
- Lawson, T.; Vialet-Chabrand, S. Speedy stomata, photosynthesis and plant water use efficiency. New Phytol. 2019, 221, 93–98. [Google Scholar] [CrossRef]
- Moshelion, M.; Halperin, O.; Wallach, R.; Oren, R.; Way, D.A. Role of aquaporins in determining transpiration and photosynthesis in water-stressed plants: Crop water-use efficiency, growth and yield. Plant Cell Environ. 2015, 38, 1785–1793. [Google Scholar] [CrossRef] [PubMed]
- Aroca, R.; Porcel, R.; Ruiz-Lozano, J.M. Regulation of root water uptake under abiotic stress conditions. J. Exp. Bot. 2012, 63, 43–57. [Google Scholar] [CrossRef] [PubMed]
- Arkhipova, T.; Sharipova, G.; Akhiyarova, G.; Kuzmina, L.; Galin, I.; Martynenko, E.; Seldimirova, O.; Nuzhnaya, T.; Feoktistova, A.; Timergalin, M.; et al. The effects of rhizosphere inoculation with Pseudomonas mandelii on formation of apoplast barriers, HvPIP2 aquaporin sand hydraulic conductance of barley. Microorganisms 2022, 10, 935. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Tan, H.-T.; Scharwies, J.; Levin, K.; Evans, J.R.; Tyerman, S.D. Association between water and carbon dioxide transport in leaf plasma membranes: Assessing the role of aquaporins. Plant Cell Environ. 2017, 40, 789–801. [Google Scholar] [CrossRef]
- Akhiyarova, G.R.; Ivanov, R.S.; Ivanov, I.I.; Finkina, E.I.; Melnikova, D.N.; Bogdanov, I.V.; Nuzhnaya, T.V.; Ovchinnikova, T.V.; Veselov, D.S.; Kudoyarova, G.R. Effects of salinity and abscisic acid on lipid transfer protein accumulation, suberin deposition and hydraulic conductance in pea roots. Membranes 2021, 11, 762. [Google Scholar] [CrossRef] [PubMed]
- Shiono, K.; Yoshikawa, M.; Kreszies, T.; Yamada, S.; Hojo, Y.; Matsuura, T.; Mori, I.C.; Schreiber, L.; Yoshioka, T. Abscisic acid is required for exodermal suberization to form a barrier to radial oxygen loss in the adventitious roots of rice (Oryza sativa). New Phytol. 2022, 233, 655–669. [Google Scholar] [CrossRef] [PubMed]
- Olaetxea, M.; Mora, V.; Bacaico, E.; Garnic, M.; Fuentes, M.; Casanova, E.; Zamarreco, A.M.; Iriarte, J.C.; Etayo, D.; Ederra, I.; et al. Abscisic acid regulation of root hydraulic conductivity and aquaporin gene expression is crucial to the plant shoot growth enhancement caused by rhizosphere humic acids. Plant Physiol. 2015, 169, 2587–2596. [Google Scholar] [CrossRef] [PubMed]
- Sharipova, G.; Veselov, D.; Kudoyarova, G.; Fricke, W.; Dodd, I.; Katsuhara, M.; Furuichi, T.; Ivanov, I.; Veselov, S. Exogenous application of abscisic acid (ABA) increases root and cell hydraulic conductivity and abundance of some aquaporin isoforms in the ABA deficient barley mutant Az34. Ann. Bot. 2016, 118, 777–785. [Google Scholar] [CrossRef]
- Bogdanov, I.V.; Finkina, E.I.; Balandin, S.V.; Melnikova, D.N.; Stukacheva, E.A.; Ovchinnikova, T.V. Structural and functional characterization of recombinant isoforms of the lentil lipid transfer protein. Acta Nat. 2015, 7, 65–73. [Google Scholar] [CrossRef]
- Bogdanov, I.V.; Shenkarev, Z.O.; Finkina, E.I.; Melnikova, D.N.; Rumynskiy, E.I.; Arseniev, A.S.; Ovchinnikova, T.V. A novel lipid transfer protein from the pea Pisum sativum: Isolation, recombinant expression, solution structure, antifungal activity, lipid binding, and allergenic properties. BMC Plant Biol. 2016, 16, 107. [Google Scholar] [CrossRef]
- Efetova, M.; Zeier, J.; Riederer, M.; Lee, C.W.; Stingl, N.; Mueller, M.; Hartung, W.; Hedrich, R.; Deeken, R. A central role of abscisic acid in drought stress protection of Agrobacterium-induced tumors on Arabidopsis. Plant Physiol. 2007, 145, 853–862. [Google Scholar] [CrossRef]
- Henzler, T.Y.; Steudle, Q.E. Oxidative gating of water channels (aquaporins) in Chara by hydroxyl radicals. Plant Cell Environ. 2004, 27, 1184–1195. [Google Scholar] [CrossRef]
- Bunce, J.A.; Ziska, L.H. Decreased hydraulic conductance in plants at elevated carbon dioxide. Plant Cell Environ. 1998, 21, 121–126. [Google Scholar] [CrossRef]
- Gonzalez-Klein, Z.; Cuevas-Zuviria, B.; Wangorsch, A.; Hernandez-Ramirez, G.; Pazos-Castro, D.; Oeo-Santos, C.; Romero-Sahagun, A.; Pacios, L.F.; Tome-Amat, J.; Scheurer, S.; et al. The key to the allergenicity of lipid transfer protein (LTP) ligands: A structural characterization. Biochim. Biophys. Acta. Mol. Cell Biol. Lipids 2021, 1866, 158928. [Google Scholar] [CrossRef]
- Scheurer, S.; Van Ree, R.; Vieths, S. The role of lipid transfer proteins as food and pollen allergens outside the mediterranean area. Curr. Allergy Asthma Rep. 2021, 21, 7. [Google Scholar] [CrossRef]
- Missaoui, K.; Gonzalez-Klein, Z.; Pazos-Castro, D.; Hernandez-Ramirez, G.; Garrido-Arandia, M.; Brini, F.; Perales, A.D.; Tome-Amat, J. Plant non-specific lipid transfer proteins. Plant Physiol. Biochem. 2022, 171, 115–127. [Google Scholar] [CrossRef]
- Megeressa, M.; Siraj, B.; Zarina, S.; Ahmed, A. Structural characterization and in vitro lipid binding studies of non-specific lipid transfer protein 1 (nsLTP1) from fennel (Foeniculum vulgare) seeds. Sci. Rep. 2020, 10, 21243. [Google Scholar] [CrossRef]
- Peretti, D.; Kim, S.H.; Tufi, R.; Lev, S. Lipid transfer proteins and membrane contact sites in human cancer. Front. Cell Dev. Biol. 2020, 7, 371. [Google Scholar] [CrossRef]
- Deeken, R.; Saupe, S.; Klinkenberg, J.; Riedel, M.; Leide, J.; Hedrich, R.; Mueller, T.D. The nonspecific lipid transfer protein atltpi-4 is involved in suberin formation of Arabidopsis thaliana crown galls. Plant Physiol. 2016, 172, 1911–1927. [Google Scholar] [CrossRef]
- Samuel, D.; Liu, Y.J.; Cheng, C.S.; Lyu, P.C. Solution structure of plant nonspecific lipid transfer protein-2 from rice (Oryza sativa). J. Biol. Chem. 2002, 277, 35267–35273. [Google Scholar] [CrossRef]
- Benning, U.F.; Tamot, B.; Guelette, B.S.; Hoffmann-Benning, S. New aspects of phloem-mediated long-distance lipid signaling inplants. Front. Plant Sci. 2012, 3, 53. [Google Scholar] [CrossRef]
- Kaluarachchi, M.; Boulangé, C.L.; Karaman, I.; Lindon, J.C.; Ebbels, T.M.D.; Elliott, P.; Tracy, R.P.; Olson, N.C. A comparison of human serum and plasma metabolites using untargeted 1H NMR spectroscopy and UPLC-MS. Metabolomics 2018, 14, 32. [Google Scholar] [CrossRef]
- Barbaglia, A.M.; Tamot, B.; Greve, V.; Hoffmann-Benning, S. Phloem proteomics reveals new lipid-binding proteins with a putative role in lipid-mediated signaling. Front. Plant Sci. 2016, 7, 563. [Google Scholar] [CrossRef]
- Guelette, B.S.; Benning, U.F.; Hoffmann-Benning, S. Identification of lipids and lipid-binding proteins in phloem exudates from Arabidopsis Thaliana. J. Exp. Bot. 2012, 63, 3603–3616. [Google Scholar] [CrossRef]
- Ogden, A.J.; Bhatt, J.J.; Brewer, H.M.; Kintigh, J.; Kariuki, S.M.; Rudrabhatla, S.; Adkins, J.N.; Curtis, W.R. Phloem exudate protein profiles during drought and recovery reveal abiotic stress responses in tomato vasculature. Int. J. Mol. Sci. 2020, 21, 4461. [Google Scholar] [CrossRef]
- Gao, S.; Guo, W.; Feng, W.; Liu, L.; Song, X.; Chen, J.; Hou, W.; Zhu, H.; Tang, S.; Hu, J. LTP3 contributes to disease susceptibility in Arabidopsis by enhancing abscisic acid (ABA) biosynthesis. Mol. Plant Pathol. 2016, 17, 412–426. [Google Scholar] [CrossRef]
- Xu, Y.; Zheng, X.; Song, Y.; Zhu, L.; Yu, Z.; Gan, L.; Zhou, S.; Liu, H.; Wen, F.; Zhu, C. NtLTP4, a lipid transfer protein that enhances salt and drought stresses tolerance in Nicotiana tabacum. Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef]
- Lee, S.B.; Go, Y.S.; Bae, H.J.; Park, J.H.; Cho, S.H.; Cho, H.J.; Lee, D.S.; Park, O.; Hwang, K.I.; Suh, M. Disruption of glycosylphosphatidylinositol-anchored lipid transfer protein gene altered cuticular lipid composition, increased plastoglobules, and enhanced susceptibility to infection by the fungal pathogen Alternaria brassicicola. Plant Physiol. 2009, 150, 42–54. [Google Scholar] [CrossRef]
- Fonseca-García, C.; Solis-Miranda, J.; Pacheco, R.; Quinto, C. Non-specific lipid transfer proteins in legumes and their participation during root-nodule symbiosis. Front. Agron. 2021, 3, 660100. [Google Scholar] [CrossRef]
- Amador, V.C.; Santos-Silva, C.A.d.; Vilela, L.M.B.; Oliveira-Lima, M.; de Santana Rêgo, M.; Roldan-Filho, R.S.; Oliveira-Silva, R.L.d.; Lemos, A.B.; de Oliveira, W.D.; Ferreira-Neto, J.R.C. Lipid transfer proteins (LTPs)—Structure, diversity and roles beyond antimicrobial activity. Antibiotics 2021, 10, 1281. [Google Scholar] [CrossRef]
- Wang, C.; Wang, H.; Li, P.; Li, H.; Xu, C.; Cohen, H.; Aharoni, A.; Wu, S. Developmental programs interact with abscisic acid to coordinate root suberization in Arabidopsis. Plant J. 2020, 104, 241–251. [Google Scholar] [CrossRef]
- Woolfson, K.N.; Esfandiari, M.; Bernards, M.A. Suberin biosynthesis, assembly and regulation. Plants 2022, 11, 555. [Google Scholar] [CrossRef]
- Sorrentino, G.; Haworth, M.; Wahbi, S.; Mahmood, T.; Zuomin, S.; Centritto, M. Abscisic acid induces rapid reductions in mesophyll conductance to carbon dioxide. PLoS ONE 2016, 11, e0148554. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Strand | 5′ to 3′ Primer Sequences | GenBank Accession Number |
|---|---|---|---|
| Ps-LTP1 | Forward | GCTGCCGGTTCTATTCCTAAA | KJ569141 |
| Reverse | GTTGGTGGAGGTACTGATCTTG | ||
| Ps-LTP2 | Forward | TGGTAGTTATTGCGCCTATGG | KJ569142 |
| Reverse | GGTGGAGGACTGGCATTATTAG | ||
| Ps-LTP3 | Forward | TGGCGATGTGCATGTTAGT | KJ569143 |
| Reverse | GGGTGAAGGACTGGCATTATTA | ||
| β-tubulin | Forward | GCTCCCAGCAGTACAGGACTCT | X54844.1 |
| Reverse | TGGCATCCCACATTTGTTGA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martynenko, E.; Arkhipova, T.; Akhiyarova, G.; Sharipova, G.; Galin, I.; Seldimirova, O.; Ivanov, R.; Nuzhnaya, T.; Finkina, E.; Ovchinnikova, T.; et al. Effects of a Pseudomonas Strain on the Lipid Transfer Proteins, Appoplast Barriers and Activity of Aquaporins Associated with Hydraulic Conductance of Pea Plants. Membranes 2023, 13, 208. https://doi.org/10.3390/membranes13020208
Martynenko E, Arkhipova T, Akhiyarova G, Sharipova G, Galin I, Seldimirova O, Ivanov R, Nuzhnaya T, Finkina E, Ovchinnikova T, et al. Effects of a Pseudomonas Strain on the Lipid Transfer Proteins, Appoplast Barriers and Activity of Aquaporins Associated with Hydraulic Conductance of Pea Plants. Membranes. 2023; 13(2):208. https://doi.org/10.3390/membranes13020208
Chicago/Turabian StyleMartynenko, Elena, Tatiana Arkhipova, Guzel Akhiyarova, Guzel Sharipova, Ilshat Galin, Oksana Seldimirova, Ruslan Ivanov, Tatiana Nuzhnaya, Ekaterina Finkina, Tatiana Ovchinnikova, and et al. 2023. "Effects of a Pseudomonas Strain on the Lipid Transfer Proteins, Appoplast Barriers and Activity of Aquaporins Associated with Hydraulic Conductance of Pea Plants" Membranes 13, no. 2: 208. https://doi.org/10.3390/membranes13020208
APA StyleMartynenko, E., Arkhipova, T., Akhiyarova, G., Sharipova, G., Galin, I., Seldimirova, O., Ivanov, R., Nuzhnaya, T., Finkina, E., Ovchinnikova, T., & Kudoyarova, G. (2023). Effects of a Pseudomonas Strain on the Lipid Transfer Proteins, Appoplast Barriers and Activity of Aquaporins Associated with Hydraulic Conductance of Pea Plants. Membranes, 13(2), 208. https://doi.org/10.3390/membranes13020208