
Rapid Production and Purification of Dye-Loaded Liposomes by Electrodialysis-Driven Depletion



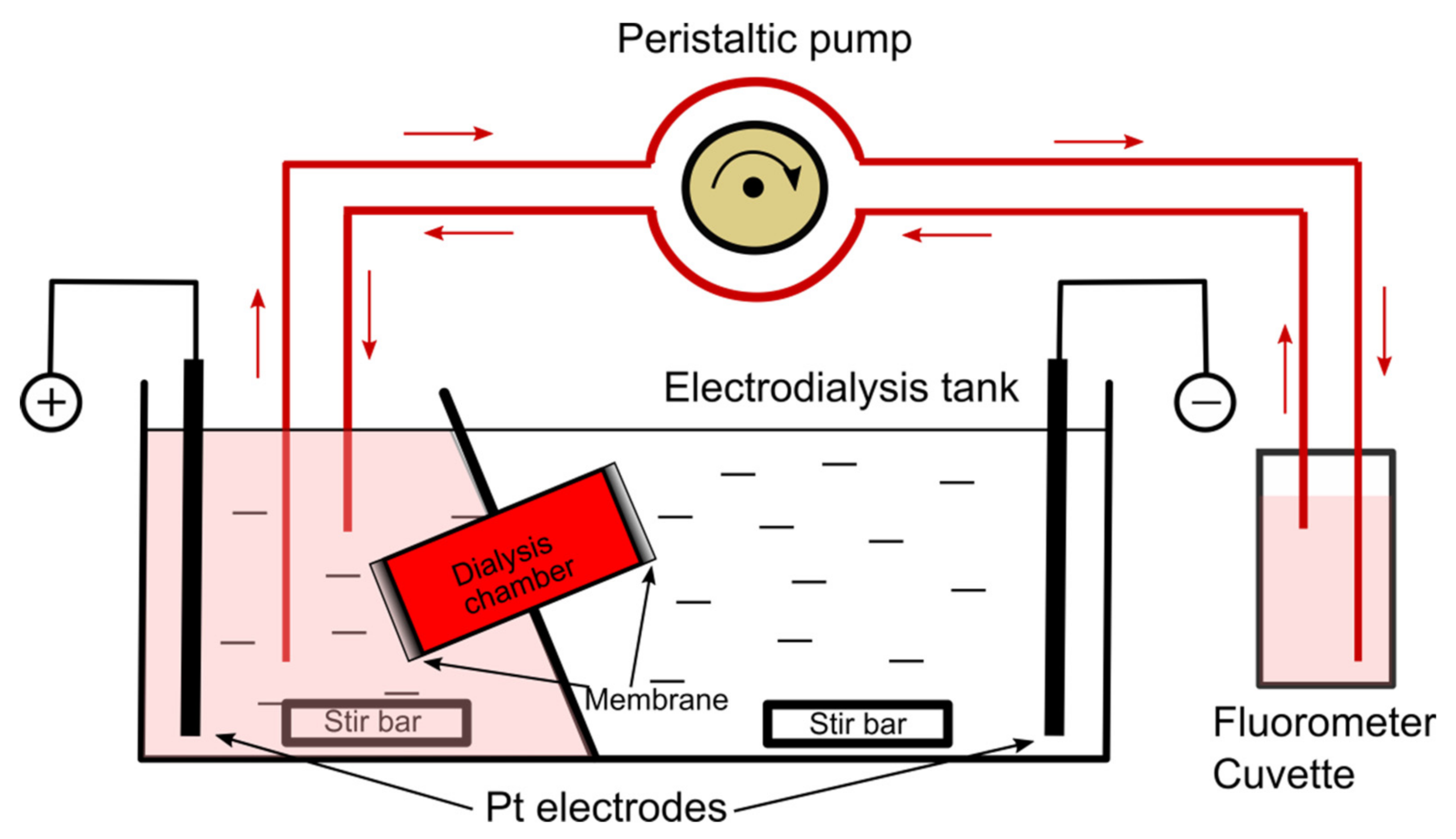{kind=link}
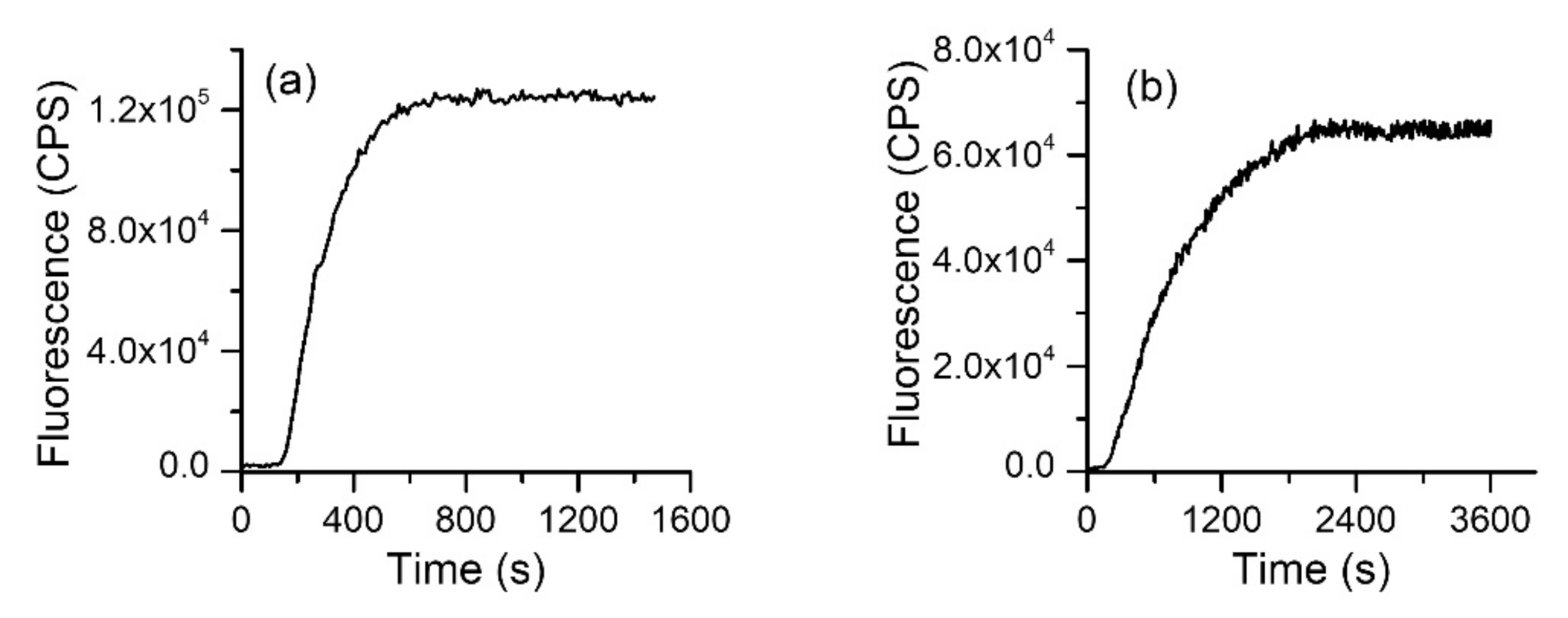{kind=link}
{kind=link}
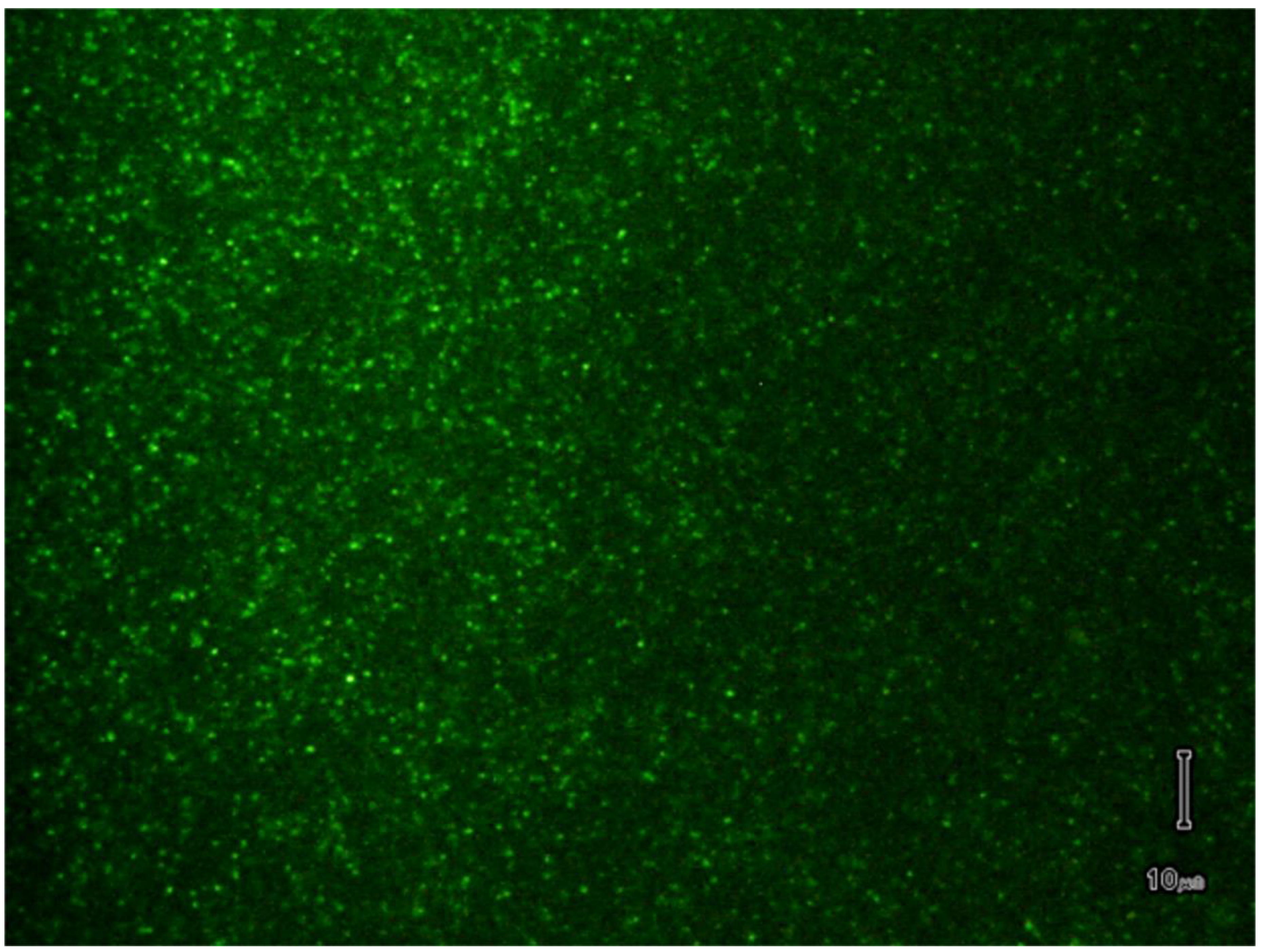{kind=link}
{kind=link}
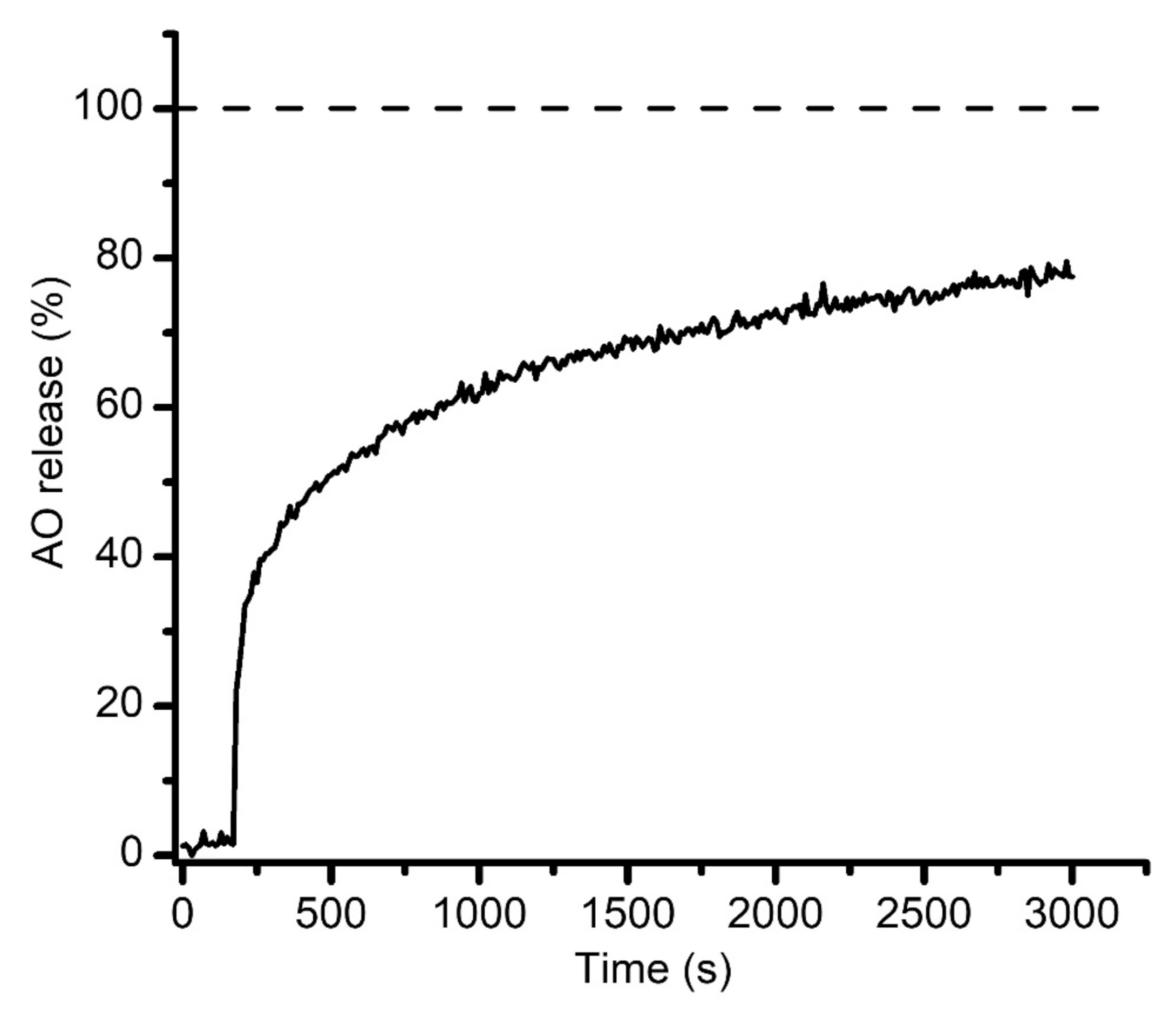{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
3. Results and Discussions
3.1. Dye separation by Electrodialysis
3.2. Simultaneous Liposome Formation, Loading and Purification by EDD
3.3. Production of Long-Circulating Liposomes by Electrodialysis
3.4. Verification of Dye Loading
3.5. Unilamellar or Multilamellar?
3.6. EDD Comparison to Extrusion and Sonication
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ajeeshkumar, K.K.; Aneesh, P.A.; Raju, N.; Suseela, M.; Ravishankar, C.N.; Benjakul, S. Advancements in liposome technology: Preparation techniques and applications in food, functional foods, and bioactive delivery: A review. Compr. Rev. Food Sci. Food Saf. 2021, 20, 1280–1306. [Google Scholar] [CrossRef] [PubMed]
- Bozzuto, G.; Molinari, A. Liposomes as nanomedical devices. Int. J. Nanomed. 2015, 10, 975–999. [Google Scholar] [CrossRef] [PubMed]
- Laouini, A.; Jaafar-Maalej, C.; Limayem-Blouza, I.; Sfar, S.; Charcosset, C.; Fessi, H. Preparation, Characterization and Applications of Liposomes: State of the Art. J. Colloid Sci. Biotechnol. 2012, 1, 147–168. [Google Scholar] [CrossRef]
- Martinez-Crespo, L.; Sun-Wang, J.L.; Sierra, A.F.; Aragay, G.; Errasti-Murugarren, E.; Bartoccioni, P.; Palacin, M.; Ballester, P. Facilitated Diffusion of Proline across Membranes of Liposomes and Living Cells by a Calix[4]pyrrole Cavitand. Chem 2020, 6, 3054–3070. [Google Scholar] [CrossRef]
- Pick, H.; Alves, A.C.; Vogel, H. Single-Vesicle Assays Using Liposomes and Cell-Derived Vesicles: From Modeling Complex Membrane Processes to Synthetic Biology and Biomedical Applications. Chem. Rev. 2018, 118, 8598–8654. [Google Scholar] [CrossRef]
- Routledge, S.J.; Linney, J.A.; Goddard, A.D. Liposomes as models for membrane integrity. Biochem. Soc. Trans. 2019, 47, 919–932. [Google Scholar] [CrossRef]
- Srivastava, A.; Eisenthal, K.B. Kinetics of molecular transport across a liposome bilayer. Chem. Phys. Lett. 1998, 292, 345–351. [Google Scholar] [CrossRef]
- Aronson, M.R.; Medina, S.; Mitchell, M.J. Peptide functionalized liposomes for receptor targeted cancer therapy. APL Bioeng. 2021, 5, 011501. [Google Scholar] [CrossRef]
- Brea, R.J.; Cole, C.M.; Lyda, B.R.; Ye, L.; Prosser, R.S.; Sunahara, R.K.; Devaraj, N.K. In Situ Reconstitution of the Adenosine A2A Receptor in Spontaneously Formed Synthetic Liposomes. J. Am. Chem. Soc. 2017, 139, 3607–3610. [Google Scholar] [CrossRef]
- Zhang, X.; Qi, J.; Lu, Y.; He, W.; Li, X.; Wu, W. Biotinylated liposomes as potential carriers for the oral delivery of insulin. Nanomedicine 2014, 10, 167–176. [Google Scholar] [CrossRef]
- Gabizon, A.; Horowitz, A.T.; Goren, D.; Tzemach, D.; Mandelbaum-Shavit, F.; Qazen, M.M.; Zalipsky, S. Targeting Folate Receptor with Folate Linked to Extremities of Poly(ethylene glycol)-Grafted Liposomes: In Vitro Studies. Bioconjug. Chem. 1999, 10, 289–298. [Google Scholar] [CrossRef]
- Soe, Z.C.; Thapa, R.K.; Ou, W.; Gautam, M.; Nguyen, H.T.; Jin, S.G.; Ku, S.K.; Oh, K.T.; Choi, H.-G.; Yong, C.S.; et al. Folate receptor-mediated celastrol and irinotecan combination delivery using liposomes for effective chemotherapy. Colloids Surf. B 2018, 170, 718–728. [Google Scholar] [CrossRef]
- Yingchoncharoen, P.; Kalinowski, D.S.; Richardson, D.R. Lipid-Based Drug Delivery Systems in Cancer Therapy: What Is Available and What Is Yet to Come. Pharmacol. Rev. 2016, 68, 701–787. [Google Scholar] [CrossRef]
- Dawidczyk, C.M.; Kim, C.; Park, J.H.; Russell, L.M.; Lee, K.H.; Pomper, M.G.; Searson, P.C. State-of-the-art in design rules for drug delivery platforms: Lessons learned from FDA-approved nanomedicines. J. Control Release 2014, 187, 133–144. [Google Scholar] [CrossRef]
- Bulbake, U.; Doppalapudi, S.; Kommineni, N.; Khan, W. Liposomal Formulations in Clinical Use: An Updated Review. Pharmaceutics 2017, 9, 12. [Google Scholar] [CrossRef]
- Beltran-Gracia, E.; Lopez-Camacho, A.; Higuera-Ciapara, I.; Velazquez-Fernandez, J.B.; Vallejo-Cardona, A.A. Nanomedicine review: Clinical developments in liposomal applications. Cancer Nanotechnol. 2019, 10, 11. [Google Scholar] [CrossRef]
- Klibanov, A.L.; Maruyama, K.; Torchilin, V.P.; Huang, L. Amphipathic polyethyleneglycols effectively prolong the circulation time of liposomes. FEBS Lett. 1990, 268, 235–237. [Google Scholar] [CrossRef]
- Najlah, M.; Said Suliman, A.; Tolaymat, I.; Kurusamy, S.; Kannappan, V.; Elhissi, A.M.A.; Wang, W. Development of Injectable PEGylated Liposome Encapsulating Disulfiram for Colorectal Cancer Treatment. Pharmaceutics 2019, 11, 610. [Google Scholar] [CrossRef]
- Hobbs, S.K.; Monsky, W.L.; Yuan, F.; Roberts, W.G.; Griffith, L.; Torchilin, V.P.; Jain, R.K. Regulation of transport pathways in tumor vessels: Role of tumor type and microenvironment. Proc. Natl. Acad. Sci. USA 1998, 95, 4607–4612. [Google Scholar] [CrossRef]
- Golombek, S.K.; May, J.-N.; Theek, B.; Appold, L.; Drude, N.; Kiessling, F.; Lammers, T. Tumor targeting via EPR: Strategies to enhance patient responses. Adv. Drug Deliv. Rev. 2018, 130, 17–38. [Google Scholar] [CrossRef]
- Rosenblum, D.; Joshi, N.; Tao, W.; Karp, J.M.; Peer, D. Progress and challenges towards targeted delivery of cancer therapeutics. Nat. Commun. 2018, 9, 1410. [Google Scholar] [CrossRef]
- Gabizon, A.; Catane, R.; Uziely, B.; Kaufman, B.; Safra, T.; Cohen, R.; Martin, F.; Huang, A.; Barenholz, Y. Prolonged Circulation Time and Enhanced Accumulation in Malignant Exudates of Doxorubicin Encapsulated in Polyethylene-glycol Coated Liposomes. Cancer Res. 1994, 54, 987. [Google Scholar]
- Cho, N.-J.; Hwang, L.Y.; Solandt, J.J.R.; Frank, C.W. Comparison of Extruded and Sonicated Vesicles for Planar Bilayer Self-Assembly. Materials 2013, 6, 3294. [Google Scholar] [CrossRef]
- Johnson, S.M.; Bangham, A.D.; Hill, M.W.; Korn, E.D. Single bilayer liposomes. Biochim. Biophys. Acta Biomembr. 1971, 233, 820–826. [Google Scholar] [CrossRef]
- Olson, F.; Hunt, C.A.; Szoka, F.C.; Vail, W.J.; Papahadjopoulos, D. Preparation of liposomes of defined size distribution by extrusion through polycarbonate membranes. Biochim. Biophys. Acta Biomembr. 1979, 557, 9–23. [Google Scholar] [CrossRef]
- Ong, S.G.; Chitneni, M.; Lee, K.S.; Ming, L.C.; Yuen, K.H. Evaluation of Extrusion Technique for Nanosizing Liposomes. Pharmaceutics 2016, 8, 36. [Google Scholar] [CrossRef] [PubMed]
- Abraham, S.A.; Waterhouse, D.N.; Mayer, L.D.; Cullis, P.R.; Madden, T.D.; Bally, M.B. The liposomal formulation of doxorubicin. Methods Enzymol. 2005, 391, 71–97. [Google Scholar] [CrossRef] [PubMed]
- Cullis, P.R.; Mayer, L.D.; Bally, M.B.; Madden, T.D.; Hope, M.J. Generating and loading of liposomal systems for drug-delivery applications. Adv. Drug Deliv. Rev. 1989, 3, 267–282. [Google Scholar] [CrossRef]
- Batzri, S.; Korn, E.D. Single bilayer liposomes prepared without sonication. Biochim. Biophys. Acta Biomembr. 1973, 298, 1015–1019. [Google Scholar] [CrossRef]
- Deamer, D.W. Preparation and properties of ether-injection liposomes. Ann. N Y Acad. Sci. 1978, 308, 250–258. [Google Scholar] [CrossRef]
- Sala, M.; Miladi, K.; Agusti, G.; Elaissari, A.; Fessi, H. Preparation of liposomes: A comparative study between the double solvent displacement and the conventional ethanol injection—From laboratory scale to large scale. Colloids Surf. Physicochem. Eng. Aspects 2017, 524, 71–78. [Google Scholar] [CrossRef]
- Liu, D.; Huang, L. Role of cholesterol in the stability of pH-sensitive, large unilamellar liposomes prepared by the detergent-dialysis method. Biochim. Biophys. Acta Biomembr. 1989, 981, 254–260. [Google Scholar] [CrossRef]
- Milsmann, M.H.; Schwendener, R.A.; Weder, H.G. The preparation of large single bilayer liposomes by a fast and controlled dialysis. Biochim. Biophys. Acta Biomembr. 1978, 512, 147–155. [Google Scholar] [CrossRef]
- Ollivon, M.; Lesieur, S.; Grabielle-Madelmont, C.C.; Paternostre, M.Ì.t. Vesicle reconstitution from lipid-detergent mixed micelles. Biochim. Biophys. Acta Biomembr. 2000, 1508, 34–50. [Google Scholar] [CrossRef]
- Schubert, R. Liposome preparation by detergent removal. Methods Enzymol. 2003, 367, 46–70. [Google Scholar] [CrossRef]
- Yao, X.; Fan, X.; Yan, N. Cryo-EM analysis of a membrane protein embedded in the liposome. Proc. Natl. Acad. Sci. USA 2020, 117, 18497. [Google Scholar] [CrossRef]
- Helenius, A.; Simons, K. Solubilization of membranes by detergents. Biochim. Biophys. Acta Rev. Biomembr. 1975, 415, 29–79. [Google Scholar] [CrossRef]
- Schubert, R.; Beyer, K.; Wolburg, H.; Schmidt, K.H. Structural changes in membranes of large unilamellar vesicles after binding of sodium cholate. Biochemistry 1986, 25, 5263–5269. [Google Scholar] [CrossRef]
- Lasic, D.D. A molecular model for vesicle formation. Biochim. Biophys. Acta Biomembr. 1982, 692, 501–502. [Google Scholar] [CrossRef]
- Lichtenberg, D.; Ahyayauch, H.; Goñi, F.M. The Mechanism of Detergent Solubilization of Lipid Bilayers. Biophys. J. 2013, 105, 289–299. [Google Scholar] [CrossRef]
- Jimah, J.R.; Schlesinger, P.H.; Tolia, N.H. Liposome Disruption Assay to Examine Lytic Properties of Biomolecules. Bio-Protocol 2017, 7, e2433. [Google Scholar] [CrossRef]
- Zagana, P.; Mourtas, S.; Basta, A.; Antimisiaris, S.G. Preparation, Physicochemical Properties, and In Vitro Toxicity towards Cancer Cells of Novel Types of Arsonoliposomes. Pharmaceutics 2020, 12, 327. [Google Scholar] [CrossRef]
- Cabral, D.J.; Hamilton, J.A.; Small, D.M. The ionization behavior of bile acids in different aqueous environments. J. Lipid Res. 1986, 27, 334–343. [Google Scholar] [CrossRef]
- Rigaud, J.-L.; Lévy, D. Reconstitution of Membrane Proteins into Liposomes. Methods Enzymol. 2003, 372, 65–86. [Google Scholar] [CrossRef]
- Agrawal, A.K.; Gupta, C.M. Tuftsin-bearing liposomes in treatment of macrophage-based infections. Adv. Drug Del. Rev. 2000, 41, 135–146. [Google Scholar] [CrossRef]
- Caron, W.P.; Lay, J.C.; Fong, A.M.; La-Beck, N.M.; Kumar, P.; Newman, S.E.; Zhou, H.; Monaco, J.H.; Clarke-Pearson, D.L.; Brewster, W.R.; et al. Translational Studies of Phenotypic Probes for the Mononuclear Phagocyte System and Liposomal Pharmacology. J. Pharmacol. Exp. Ther. 2013, 347, 599. [Google Scholar] [CrossRef]
- Kao, Y.J.; Juliano, R.L. Interactions of liposomes with the reticuloendothelial system. Effects of reticuloendothelial blockade on the clearance of large unilamellar vesicles. Biochim. Biophys. Acta Gen. Subj. 1981, 677, 453–461. [Google Scholar] [CrossRef]
- McSweeney, M.D.; Shen, L.; DeWalle, A.C.; Joiner, J.B.; Ciociola, E.C.; Raghuwanshi, D.; Macauley, M.S.; Lai, S.K. Pre-treatment with high molecular weight free PEG effectively suppresses anti-PEG antibody induction by PEG-liposomes in mice. J. Control Release 2021, 329, 774–781. [Google Scholar] [CrossRef]
- Papahadjopoulos, D.; Allen, T.M.; Gabizon, A.; Mayhew, E.; Matthay, K.; Huang, S.K.; Lee, K.D.; Woodle, M.C.; Lasic, D.D.; Redemann, C. Sterically stabilized liposomes: Improvements in pharmacokinetics and antitumor therapeutic efficacy. Proc. Nat. Acad. Sci. USA 1991, 88, 11460. [Google Scholar] [CrossRef]
- Amado, A.M.; Ramos, A.P.; Silva, E.R.; Borissevitch, I.E. Quenching of acridine orange fluorescence by salts in aqueous solutions: Effects of aggregation and charge transfer. J. Lumin. 2016, 178, 288–294. [Google Scholar] [CrossRef]
- Bae, W.; Yoon, T.-Y.; Jeong, C. Direct evaluation of self-quenching behavior of fluorophores at high concentrations using an evanescent field. PLoS ONE 2021, 16, e0247326. [Google Scholar] [CrossRef] [PubMed]
- Hamann, S.; Kiilgaard, J.F.; Litman, T.; Alvarez-Leefmans, F.J.; Winther, B.R.; Zeuthen, T. Measurement of Cell Volume Changes by Fluorescence Self-Quenching. J. Fluoresc. 2002, 12, 139–145. [Google Scholar] [CrossRef]
- Needham, D.; Anyarambhatla, G.; Kong, G.; Dewhirst, M.W. A New Temperature-sensitive Liposome for Use with Mild Hyperthermia: Characterization and Testing in a Human Tumor Xenograft Model. Cancer Res. 2000, 60, 1197. [Google Scholar] [PubMed]
- Wang, T.; Deng, Y.; Geng, Y.; Gao, Z.; Zou, J.; Wang, Z. Preparation of submicron unilamellar liposomes by freeze-drying double emulsions. Biochim. Biophys. Acta Biomembr. 2006, 1758, 222–231. [Google Scholar] [CrossRef]
- Tejera-Garcia, R.; Ranjan, S.; Zamotin, V.; Sood, R.; Kinnunen, P.K.J. Making Unilamellar Liposomes Using Focused Ultrasound. Langmuir 2011, 27, 10088–10097. [Google Scholar] [CrossRef]
- Ide, T.; Aoki, T.; Takeuchi, Y.; Yanagida, T. Lysenin forms a voltage-dependent channel in artificial lipid bilayer membranes. Biochem. Biophys. Res. Commun. 2006, 346, 288–292. [Google Scholar] [CrossRef]
- Ishitsuka, R.; Kobayashi, T. Lysenin: A new tool for investigating membrane lipid organization. Anat. Sci. Int. 2004, 79, 184–190. [Google Scholar] [CrossRef]
- Kulma, M.; Hereć, M.; Grudziński, W.; Anderluh, G.; Gruszecki, W.I.; Kwiatkowska, K.; Sobota, A. Sphingomyelin-rich domains are sites of lysenin oligomerization: Implications for raft studies. Biochim. Biophys. Acta Biomembr. 2010, 1798, 471–481. [Google Scholar] [CrossRef]
- Yamaji-Hasegawa, A.; Makino, A.; Baba, T.; Senoh, Y.; Kimura-Suda, H.; Sato, S.B.; Terada, N.; Ohno, S.; Kiyokawa, E.; Umeda, M.; et al. Oligomerization and pore formation of a sphingomyelin-specific toxin, lysenin. J. Biol. Chem. 2003, 278, 22762–22770. [Google Scholar] [CrossRef]
- Shrestha, N.; Thomas, C.A.; Richtsmeier, D.; Bogard, A.; Hermann, R.; Walker, M.; Abatchev, G.; Brown, R.J.; Fologea, D. Temporary Membrane Permeabilization via the Pore-Forming Toxin Lysenin. Toxins 2020, 12, 343. [Google Scholar] [CrossRef]
- Lapinski, M.M.; Castro-Forero, A.; Greiner, A.J.; Ofoli, R.Y.; Blanchard, G.J. Comparison of Liposomes Formed by Sonication and Extrusion: Rotational and Translational Diffusion of an Embedded Chromophore. Langmuir 2007, 23, 11677–11683. [Google Scholar] [CrossRef]
- Danaei, M.; Dehghankhold, M.; Ataei, S.; Hasanzadeh Davarani, F.; Javanmard, R.; Dokhani, A.; Khorasani, S.; Mozafari, M.R. Impact of Particle Size and Polydispersity Index on the Clinical Applications of Lipidic Nanocarrier Systems. Pharmaceutics 2018, 10, 57. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abatchev, G.; Bogard, A.; Hutchinson, Z.; Ward, J.; Fologea, D. Rapid Production and Purification of Dye-Loaded Liposomes by Electrodialysis-Driven Depletion. Membranes 2021, 11, 417. https://doi.org/10.3390/membranes11060417
Abatchev G, Bogard A, Hutchinson Z, Ward J, Fologea D. Rapid Production and Purification of Dye-Loaded Liposomes by Electrodialysis-Driven Depletion. Membranes. 2021; 11(6):417. https://doi.org/10.3390/membranes11060417
Chicago/Turabian StyleAbatchev, Gamid, Andrew Bogard, Zoe Hutchinson, Jason Ward, and Daniel Fologea. 2021. "Rapid Production and Purification of Dye-Loaded Liposomes by Electrodialysis-Driven Depletion" Membranes 11, no. 6: 417. https://doi.org/10.3390/membranes11060417
APA StyleAbatchev, G., Bogard, A., Hutchinson, Z., Ward, J., & Fologea, D. (2021). Rapid Production and Purification of Dye-Loaded Liposomes by Electrodialysis-Driven Depletion. Membranes, 11(6), 417. https://doi.org/10.3390/membranes11060417