
A Combined Adjuvant TF–Al Consisting of TFPR1 and Aluminum Hydroxide Augments Strong Humoral and Cellular Immune Responses in Both C57BL/6 and BALB/c Mice

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Mouse Immunization and Sample Collection
2.3. ELISA
2.4. ELISPOT Assay for IFN-γ and IL-4
2.5. Stimulation of Mouse Splenocytes with TFPR1 In Vitro
2.6. Detection of Cytokine Levels In Vivo Immunized with TFPR1
2.7. Statistical Analysis
3. Results
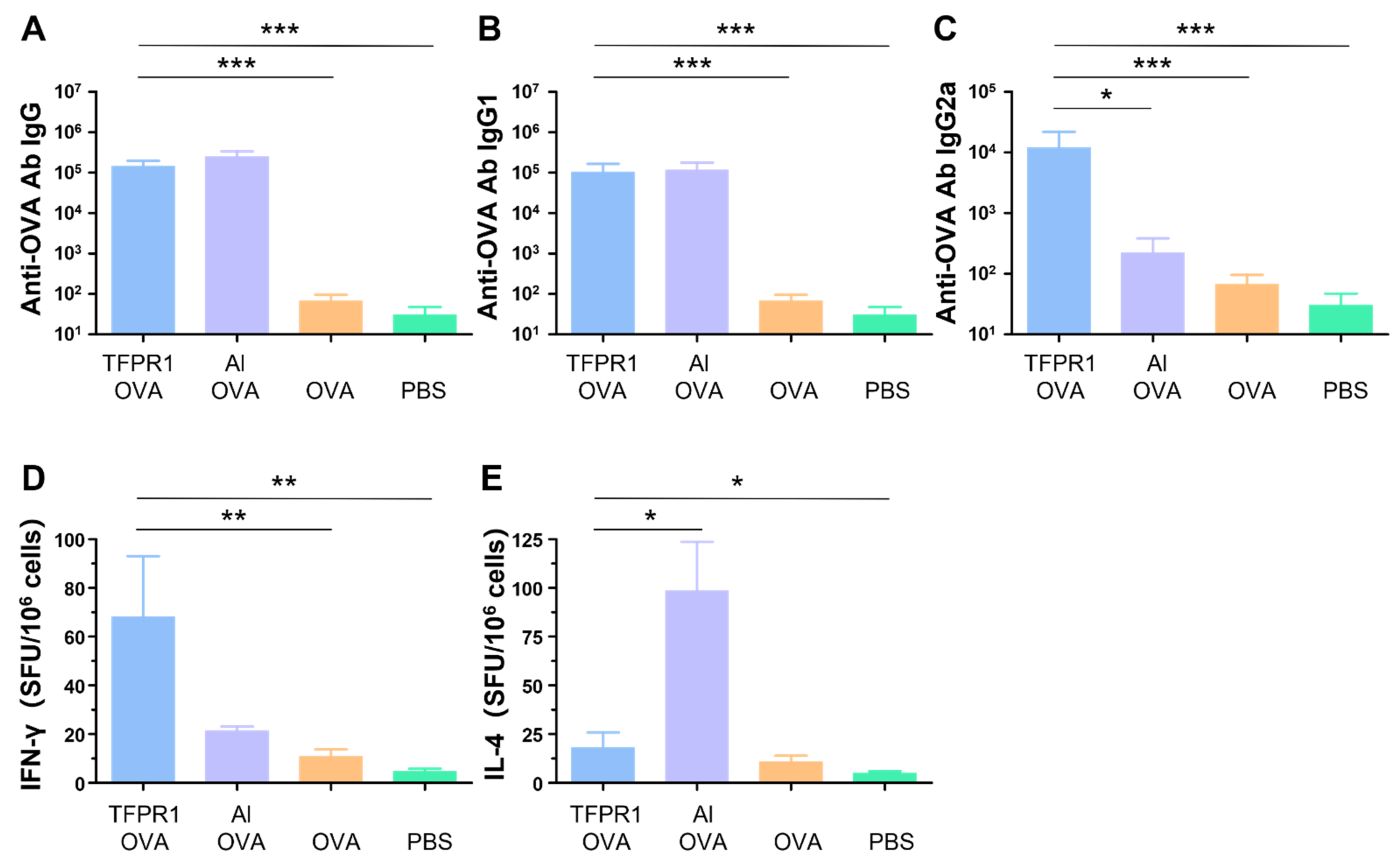
3.1. TFPR1 Acts as an Effective Adjuvant for the Model Antigen OVA in C57BL/6 Mice
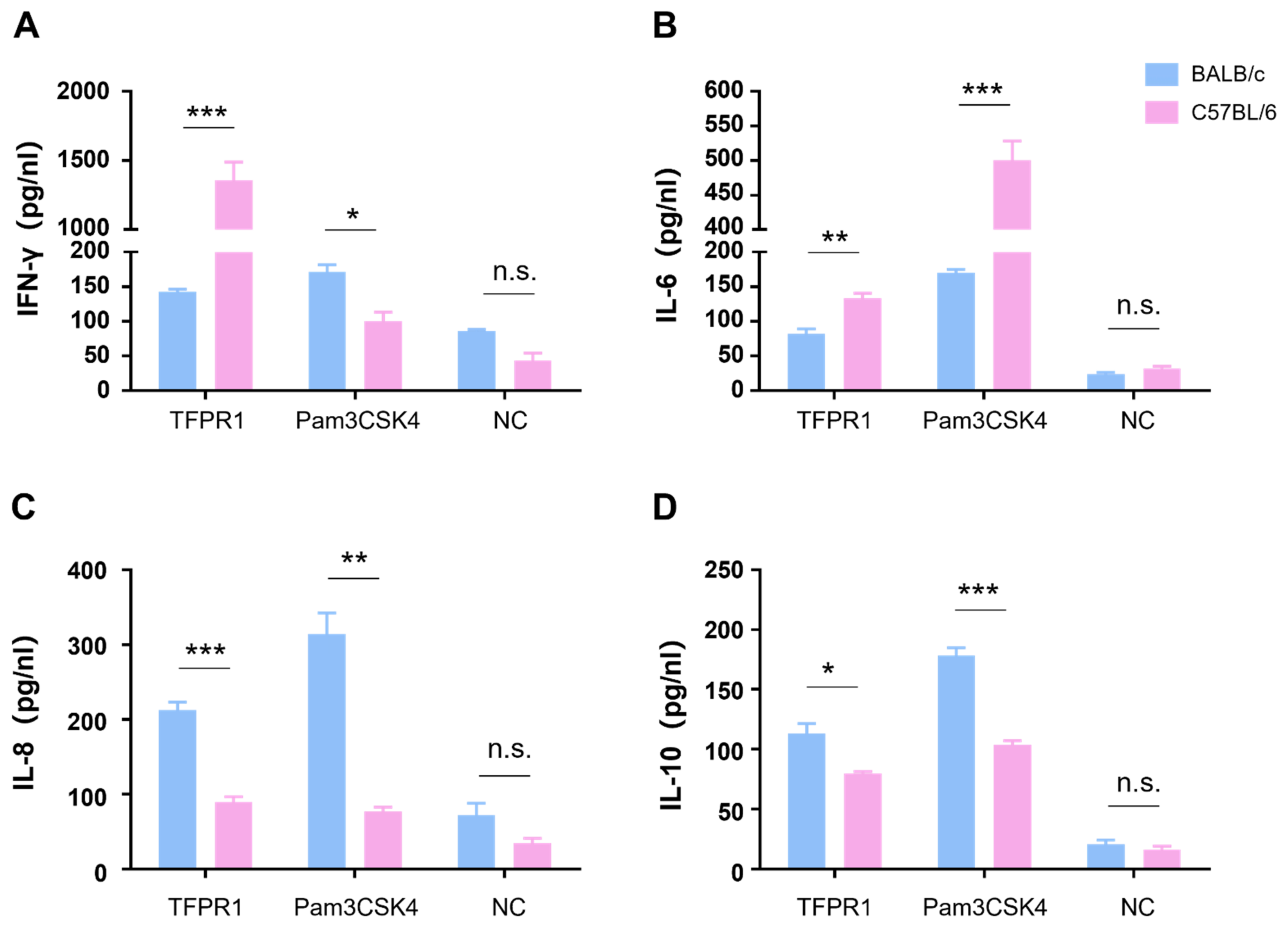
3.2. TFPR1 Activates Splenocytes from BALB/c Mice or C57BL/6 Mice to Produce Cytokines with Different Profiles
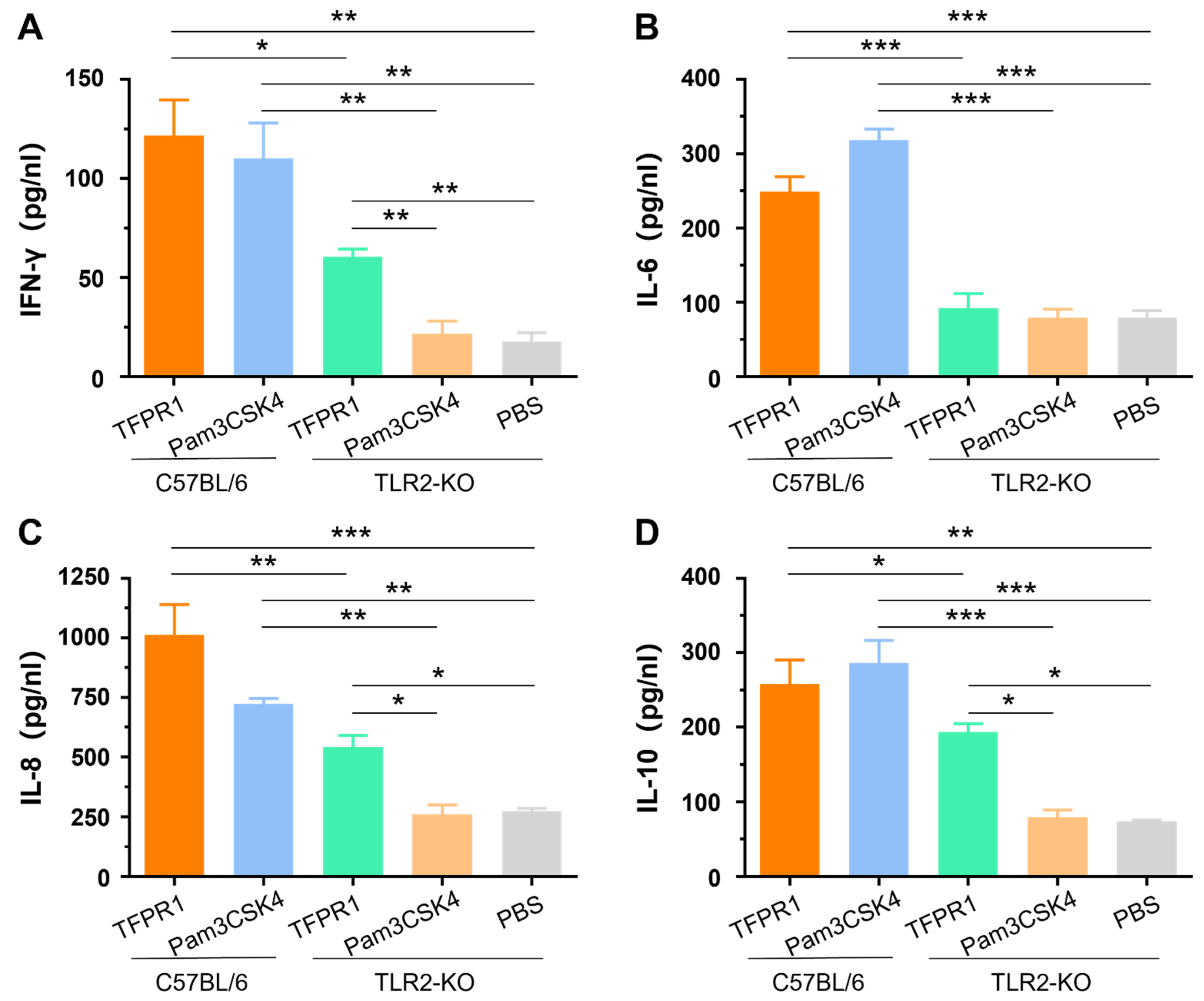
3.3. TFPR1 Activates TLR2-KO Mice to Generate Lower Levels of Cytokines than Those in C57BL/6 Mice
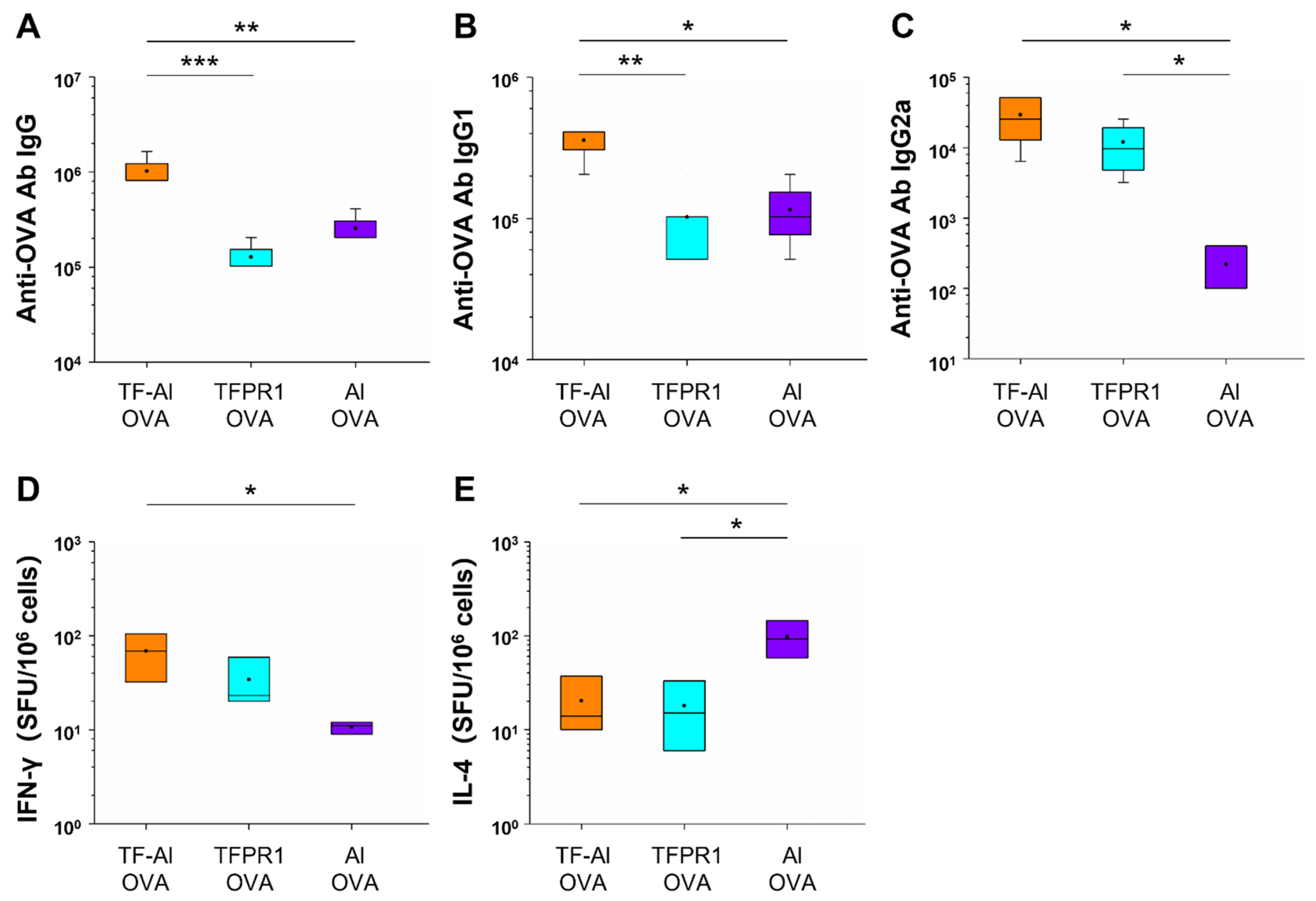
3.4. TF–Al Augments Anti-OVA-Specific Humoral and Cellular Immune Responses in C57BL/6 Mice
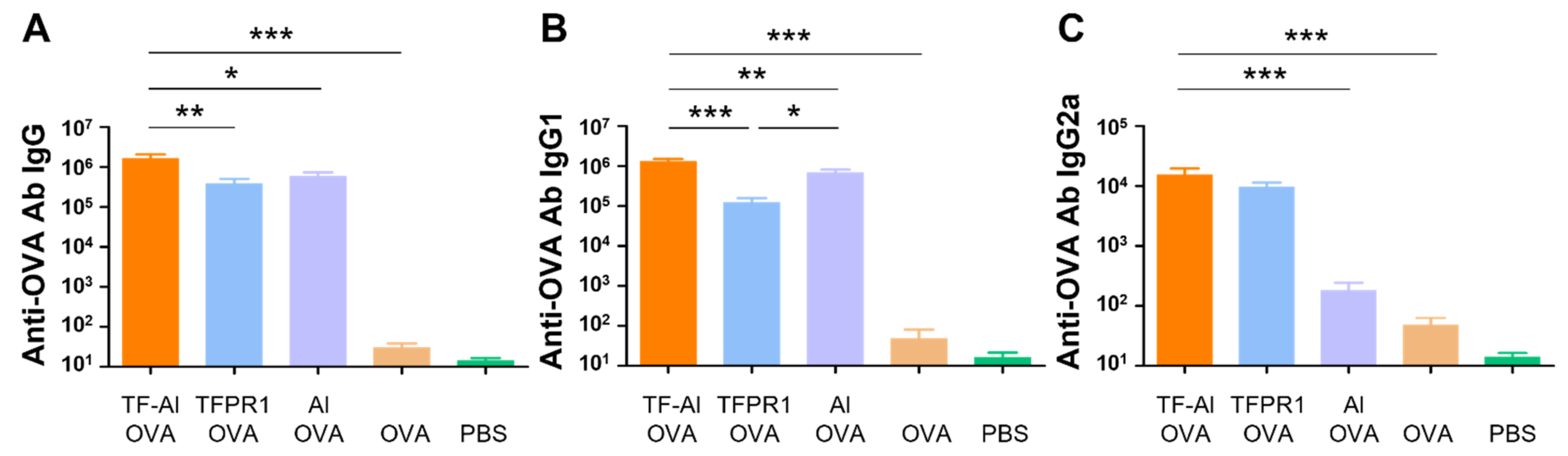
3.5. TF–Al Acts as an Effective Adjuvant for the Model Antigen OVA in BALB/c Mice
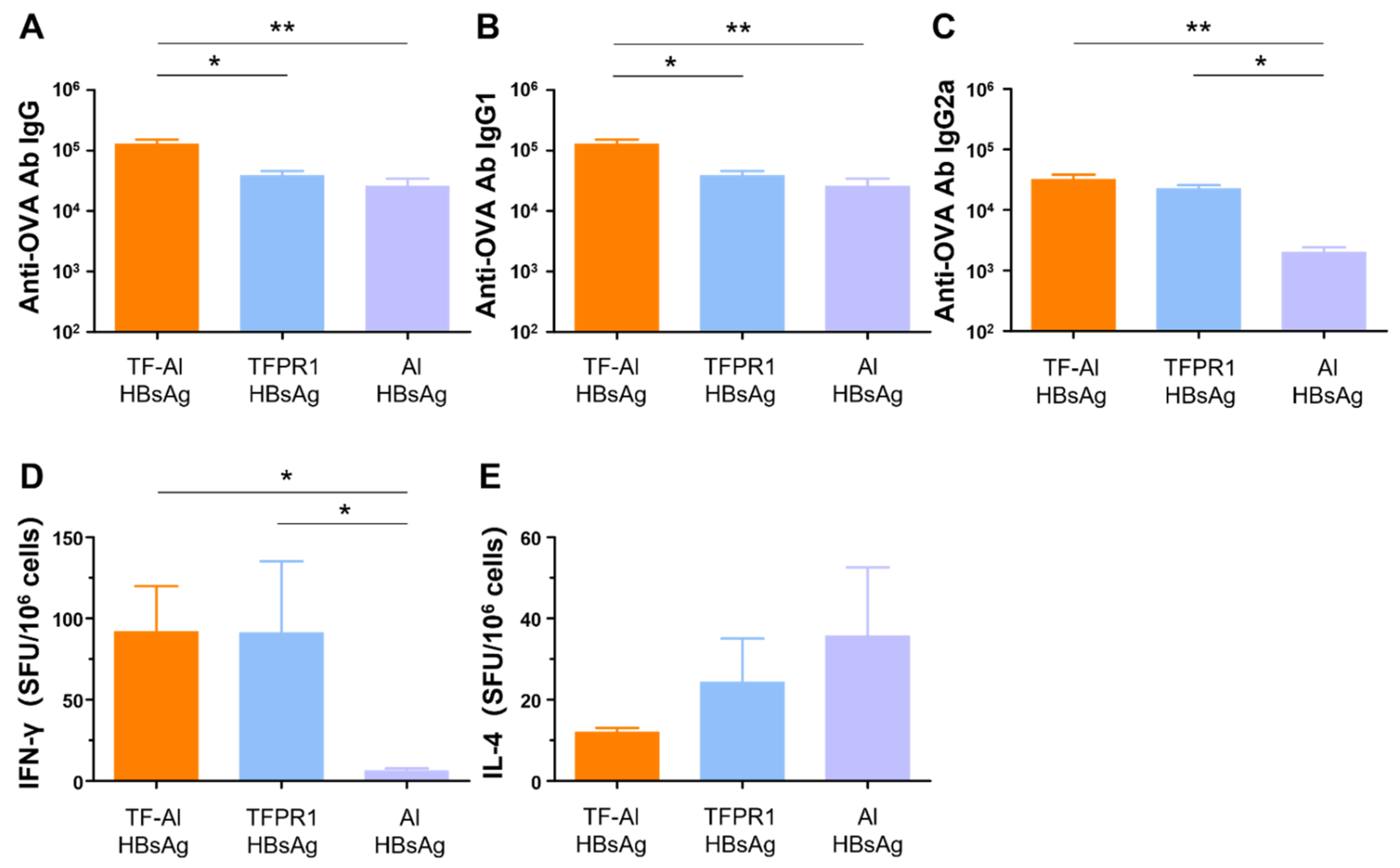
3.6. TF–Al Augments Anti-HBsAg-Specific Humoral and Cellular Immune Responses in BALB/c Mice
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- O’Hagan, D.T.; Lodaya, R.N.; Lofano, G. The continued advance of vaccine adjuvants—‘We can work it out’. Semin. Immunol. 2020, 50, 101426. [Google Scholar] [CrossRef] [PubMed]
- Carter, D.; Duthie, M.S.; Reed, S.G. Adjuvants. Curr. Top. Microbiol. Immunol. 2020, 428, 103–127. [Google Scholar]
- Shi, S.; Zhu, H.; Xia, X.; Liang, Z.; Ma, X.; Sun, B. Vaccine adjuvants: Understanding the structure and mechanism of adjuvanticity. Vaccine 2019, 37, 3167–3178. [Google Scholar] [CrossRef] [PubMed]
- Hardt, K.; Bonanni, P.; King, S.; Santos, J.I.; El-Hodhod, M.; Zimet, G.D.; Preiss, S. Vaccine strategies: Optimising outcomes. Vaccine 2016, 34, 6691–6699. [Google Scholar] [CrossRef] [Green Version]
- Danielsson, R.; Eriksson, H. Aluminium adjuvants in vaccines—A way to modulate the immune response. Semin. Cell Dev. Biol. 2021, 115, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Cerofolini, L.; Giuntini, S.; Ravera, E.; Luchinat, C.; Berti, F.; Fragai, M. Structural characterization of a protein adsorbed on aluminum hydroxide adjuvant in vaccine formulation. NPJ Vaccines 2019, 4, 20. [Google Scholar] [CrossRef] [Green Version]
- Montomoli, E.; Piccirella, S.; Khadang, B.; Mennitto, E.; Camerini, R.; De Rosa, A. Current adjuvants and new perspectives in vaccine formulation. Expert Rev. Vaccines 2011, 10, 1053–1061. [Google Scholar] [CrossRef]
- Martinon, S.; Cisneros, A.; Villicana, S.; Hernandez-Miramontes, R.; Mixcoha, E.; Calderon-Vargas, P. Chemical and Immunological Characteristics of Aluminum-Based, Oil-Water Emulsion, and Bacterial-Origin Adjuvants. J. Immunol. Res. 2019, 2019, 3974127. [Google Scholar] [CrossRef] [Green Version]
- Domnich, A.; Arata, L.; Amicizia, D.; Puig-Barbera, J.; Gasparini, R.; Panatto, D. Effectiveness of MF59-adjuvanted seasonal influenza vaccine in the elderly: A systematic review and meta-analysis. Vaccine 2017, 35, 513–520. [Google Scholar] [CrossRef]
- Schijns, V.E.; Lavelle, E.C. Trends in vaccine adjuvants. Expert Rev. Vaccines 2011, 10, 539–550. [Google Scholar] [CrossRef]
- Paudel, Y.N.; Angelopoulou, E.; Akyuz, E.; Piperi, C.; Othman, I.; Shaikh, M.F. Role of Innate Immune Receptor TLR4 and its endogenous ligands in epileptogenesis. Pharmacol. Res. 2020, 160, 105172. [Google Scholar] [CrossRef] [PubMed]
- Luchner, M.; Reinke, S.; Milicic, A. TLR Agonists as Vaccine Adjuvants Targeting Cancer and Infectious Diseases. Pharmaceutics 2021, 13, 142. [Google Scholar] [CrossRef] [PubMed]
- Kayraklioglu, N.; Horuluoglu, B.; Klinman, D.M. CpG Oligonucleotides as Vaccine Adjuvants. Methods Mol. Biol. 2021, 2197, 51–85. [Google Scholar] [PubMed]
- Pahlavanneshan, S.; Sayadmanesh, A.; Ebrahimiyan, H.; Basiri, M. Toll-Like Receptor-Based Strategies for Cancer Immunotherapy. J. Immunol. Res. 2021, 2021, 9912188. [Google Scholar] [CrossRef]
- McKee, A.S.; Marrack, P. Old and new adjuvants. Curr. Opin. Immunol. 2017, 47, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Lindemann, M.; Zaslavskaya, M.; Fiedler, M.; Wilde, B.; Heinemann, F.M.; Heinold, A.; Horn, P.A.; Witzke, O. Humoral and Cellular Responses to a Single Dose of Fendrix in Renal Transplant Recipients with Non-response to Previous Hepatitis B Vaccination. Scand. J. Immunol. 2017, 85, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.M.; Puthanakit, T.; Cheng-Hsun, C.; Ren-Bin, T.; Schwarz, T.; Pellegrino, A.; Esposito, S.; Frenette, L.; McNeil, S.; Durando, P.; et al. Sustained Immunogenicity of 2-dose Human Papillomavirus 16/18 AS04-adjuvanted Vaccine Schedules in Girls Aged 9-14 Years: A Randomized Trial. J. Infect. Dis. 2017, 215, 1711–1719. [Google Scholar] [CrossRef]
- Rivera, L.; Chanthavanich, P.; Poder, A.; Suryakiran, P.V.; Jastorff, A.; van der Wielen, M. MenACWY-TT is immunogenic when co-administered with Tdap and AS04-HPV16/18 in girls and young women: Results from a phase III randomized trial. Vaccine 2018, 36, 3967–3975. [Google Scholar] [CrossRef]
- Garcon, N.; Di Pasquale, A. From discovery to licensure, the Adjuvant System story. Hum. Vaccin. Immunother. 2017, 13, 19–33. [Google Scholar] [CrossRef]
- Richmond, P.; Hatchuel, L.; Dong, M.; Ma, B.; Hu, B.; Smolenov, I.; Li, P.; Liang, P.; Han, H.H.; Liang, J.; et al. Safety and immunogenicity of S-Trimer (SCB-2019), a protein subunit vaccine candidate for COVID-19 in healthy adults: A phase 1, randomised, double-blind, placebo-controlled trial. Lancet 2021, 397, 682–694. [Google Scholar] [CrossRef]
- Sun, W.; Li, Q.; Ning, X.; Yang, Y.; Guo, J.; Zhu, Q.; Guo, Y.; Li, H.; Wang, Y.; Zhou, Y.; et al. TFPR1 acts as an immune regulator and an efficient adjuvant for proteins and peptides by activating immune cells, primarily through TLR2. Vaccine 2020, 38, 288–297. [Google Scholar] [CrossRef]
- Li, Q.; Wang, Y.; Zhu, Q.; Ning, X.; Sun, W.; Zhou, Y.; Kou, Z. TFPR1 acts as a novel adjuvant by activating Dendritic cells and promoting its maturation. Mil. Med. Sci. 2018, 42, 101–104. [Google Scholar]
- Tam, W.Y.; Cheung, K.K. Phenotypic characteristics of commonly used inbred mouse strains. J. Mol. Med. 2020, 98, 1215–1234. [Google Scholar] [CrossRef]
- Beck, J.A.; Lloyd, S.; Hafezparast, M.; Lennon-Pierce, M.; Eppig, J.T.; Festing, M.F.; Fisher, E.M.C. Genealogies of mouse inbred strains. Nat. Genet. 2000, 24, 23–25. [Google Scholar] [CrossRef] [PubMed]
- Mosley, Y.C.; Lu, F.; HogenEsch, H. Differences in innate IFNγ and IL-17 responses to Bordetella pertussis between BALB/c and C57BL/6 mice: Role of γδT cells, NK cells, and dendritic cells. Immunol. Res. 2017, 65, 1139–1149. [Google Scholar] [CrossRef]
- Watanabe, H.; Numata, K.; Ito, T.; Takagi, K.; Matsukawa, A. Innate immune response in Th1- and Th2-dominant mouse strains. Shock 2004, 22, 460–466. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Matsuguchi, T.; Tsuboi, N.; Yajima, T.; Yoshikai, Y. Differences in expression of toll-like receptors and their reactivities in dendritic cells in BALB/c and C57BL/6 mice. Infect. Immun. 2002, 70, 6638–6645. [Google Scholar] [CrossRef] [Green Version]
- Shan, W.; Zheng, H.; Fu, G.; Liu, C.; Li, Z.; Ye, Y.; Zhao, J.; Xu, D.; Sun, L.; Wang, X.; et al. Bioengineered Nanocage from HBc Protein for Combination Cancer Immunotherapy. Nano Lett. 2019, 19, 1719–1727. [Google Scholar] [CrossRef] [PubMed]
- Stevens, J.C.; Banks, G.T.; Festing, M.F.; Fisher, E.M. Quiet mutations in inbred strains of mice. Trends Mol. Med. 2007, 13, 512–519. [Google Scholar] [CrossRef] [PubMed]
- Locarnini, S.; Hatzakis, A.; Chen, D.S.; Lok, A. Strategies to control hepatitis B: Public policy, epidemiology, vaccine and drugs. J. Hepatol. 2015, 62, S76–S86. [Google Scholar] [CrossRef]
- Li, Q.; Ning, X.; Wang, Y.; Zhu, Q.; Guo, Y.; Li, H.; Zhou, Y.; Kou, Z. The integrity of α-β-α sandwich conformation is essential for a novel adjuvant TFPR1 to maintain its adjuvanticity. Biomolecules 2019, 9, 869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollard, A.J.; Bijker, E.M. A guide to vaccinology: From basic principles to new developments. Nat. Rev. Immunol. 2021, 21, 83–100. [Google Scholar] [CrossRef]
- Pulendran, B.; Davis, M.M. The science and medicine of human immunology. Science 2020, 369, 6511. [Google Scholar] [CrossRef]
- Mascola, J.R.; Fauci, A.S. Novel vaccine technologies for the 21st century. Nat. Rev. Immunol. 2020, 20, 87–88. [Google Scholar] [CrossRef] [PubMed]
- Rappuoli, R.; Hanon, E. Sustainable vaccine development: A vaccine manufacturer’s perspective. Curr. Opin. Immunol. 2018, 53, 111–118. [Google Scholar] [CrossRef]
- Pulendran, B.; Arunachalam, P.S.; O’Hagan, D.T. Emerging concepts in the science of vaccine adjuvants. Nat. Rev. Drug Discov. 2021, 20, 454–475. [Google Scholar] [CrossRef]
- Reed, S.G.; Tomai, M.; Gale, M.J., Jr. New horizons in adjuvants for vaccine development. Curr. Opin. Immunol. 2020, 65, 97–101. [Google Scholar] [CrossRef]
- Burakova, Y.; Madera, R.; McVey, S.; Schlup, J.R.; Shi, J. Adjuvants for Animal Vaccines. Viral Immunol. 2018, 31, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Vetter, V.; Denizer, G.; Friedland, L.R.; Krishnan, J.; Shapiro, M. Understanding modern-day vaccines: What you need to know. Ann. Med. 2018, 50, 110–120. [Google Scholar] [CrossRef]
- Reed, S.G.; Orr, M.T.; Fox, C.B. Key roles of adjuvants in modern vaccines. Nat. Med. 2013, 19, 1597–1608. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, S.; Li, H.; Wang, H.; Zhang, T.; Hutchinson, M.R.; Yin, H.; Wang, X. Small-Molecule Modulators of Toll-like Receptors. Acc. Chem. Res. 2020, 53, 1046–1055. [Google Scholar] [CrossRef]
- Dunne, A.; Marshall, N.A.; Mills, K.H. TLR based therapeutics. Curr. Opin. Pharmacol. 2011, 11, 404–411. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Chen, M.; Wang, T. Liposomes used as a vaccine adjuvant-delivery system: From basics to clinical immunization. J. Control. Release 2019, 303, 130–150. [Google Scholar] [CrossRef] [PubMed]
- Garcia, A.; De Sanctis, J.B. An overview of adjuvant formulations and delivery systems. APMIS 2014, 122, 257–267. [Google Scholar] [CrossRef] [PubMed]
- Bonam, S.R.; Partidos, C.D.; Halmuthur, S.K.M.; Muller, S. An Overview of Novel Adjuvants Designed for Improving Vaccine Efficacy. Trends Pharmacol. Sci. 2017, 38, 771–793. [Google Scholar] [CrossRef]
- Mount, A.; Koernig, S.; Silva, A.; Drane, D.; Maraskovsky, E.; Morelli, A.B. Combination of adjuvants: The future of vaccine design. Expert Rev. Vaccines 2013, 12, 733–746. [Google Scholar] [CrossRef] [PubMed]
- Guy, B. The perfect mix: Recent progress in adjuvant research. Nat. Rev. Microbiol. 2007, 5, 505–517. [Google Scholar] [CrossRef]
- Leroux-Roels, G.; Leroux-Roels, I.; Clement, F.; Ofori-Anyinam, O.; Lievens, M.; Jongert, E.; Moris, P.; Ballou, W.R.; Cohen, J. Evaluation of the immune response to RTS,S/AS01 and RTS,S/AS02 adjuvanted vaccines: Randomized, double-blind study in malaria-naive adults. Hum. Vaccines Immunother. 2014, 10, 2211–2219. [Google Scholar] [CrossRef] [Green Version]
- Lecrenier, N.; Beukelaers, P.; Colindres, R.; Curran, D.; De Kesel, C.; De Saegher, J.P.; Didierlaurent, A.M.; Ledent, E.Y.; Mols, J.F.; Mrkvan, T.; et al. Development of adjuvanted recombinant zoster vaccine and its implications for shingles prevention. Expert Rev. Vaccines 2018, 17, 619–634. [Google Scholar] [CrossRef]
- Del Giudice, G.; Rappuoli, R.; Didierlaurent, A.M. Correlates of adjuvanticity: A review on adjuvants in licensed vaccines. Semin. Immunol. 2018, 39, 14–21. [Google Scholar] [CrossRef]
- Didierlaurent, A.M.; Laupeze, B.; Di Pasquale, A.; Hergli, N.; Collignon, C.; Garcon, N. Adjuvant system AS01: Helping to overcome the challenges of modern vaccines. Expert Rev. Vaccines 2017, 16, 55–63. [Google Scholar] [CrossRef] [Green Version]
- Karandikar, S.H.; Sidney, J.; Sette, A.; Selby, M.J.; Korman, A.J.; Srivastava, P.K. New epitopes in ovalbumin provide insights for cancer neoepitopes. JCI Insight 2019, 5, 127882. [Google Scholar] [CrossRef] [PubMed]
- Sjaastad, F.V.; Kucaba, T.A.; Dileepan, T.; Swanson, W.; Dail, C.; Cabrera-Perez, J.; Murphy, K.A.; Badovinac, V.P.; Griffith, T.S. Polymicrobial Sepsis Impairs Antigen-Specific Memory CD4 T Cell-Mediated Immunity. Front. Immunol. 2020, 11, 1786. [Google Scholar] [CrossRef] [PubMed]
- Jarnagin, K.; Alvarez, O.; Shresta, S.; Webb, D.R. Animal models for SARS-Cov2/Covid19 research-A commentary. Biochem. Pharmacol. 2021, 188, 114543. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Q.; Liu, Z.; Liu, Y.; Liang, C.; Shu, J.; Jin, X.; Li, C.; Kou, Z. A Combined Adjuvant TF–Al Consisting of TFPR1 and Aluminum Hydroxide Augments Strong Humoral and Cellular Immune Responses in Both C57BL/6 and BALB/c Mice. Vaccines 2021, 9, 1408. https://doi.org/10.3390/vaccines9121408
Li Q, Liu Z, Liu Y, Liang C, Shu J, Jin X, Li C, Kou Z. A Combined Adjuvant TF–Al Consisting of TFPR1 and Aluminum Hydroxide Augments Strong Humoral and Cellular Immune Responses in Both C57BL/6 and BALB/c Mice. Vaccines. 2021; 9(12):1408. https://doi.org/10.3390/vaccines9121408
Chicago/Turabian StyleLi, Qiao, Zhihua Liu, Yi Liu, Chen Liang, Jiayi Shu, Xia Jin, Chuanyou Li, and Zhihua Kou. 2021. "A Combined Adjuvant TF–Al Consisting of TFPR1 and Aluminum Hydroxide Augments Strong Humoral and Cellular Immune Responses in Both C57BL/6 and BALB/c Mice" Vaccines 9, no. 12: 1408. https://doi.org/10.3390/vaccines9121408
APA StyleLi, Q., Liu, Z., Liu, Y., Liang, C., Shu, J., Jin, X., Li, C., & Kou, Z. (2021). A Combined Adjuvant TF–Al Consisting of TFPR1 and Aluminum Hydroxide Augments Strong Humoral and Cellular Immune Responses in Both C57BL/6 and BALB/c Mice. Vaccines, 9(12), 1408. https://doi.org/10.3390/vaccines9121408