
Chimeric Vaccines Based on Novel Insect-Specific Flaviviruses




Abstract
:1. Introduction
2. Flavivirus Vaccine Development
3. Genetically Engineered Live-Attenuated Vaccines
4. Challenges for Flavivirus Vaccine Development
Induction of Non-Neutralizing Cross-Reactive Antibodies and Antibody-Dependent Enhancement
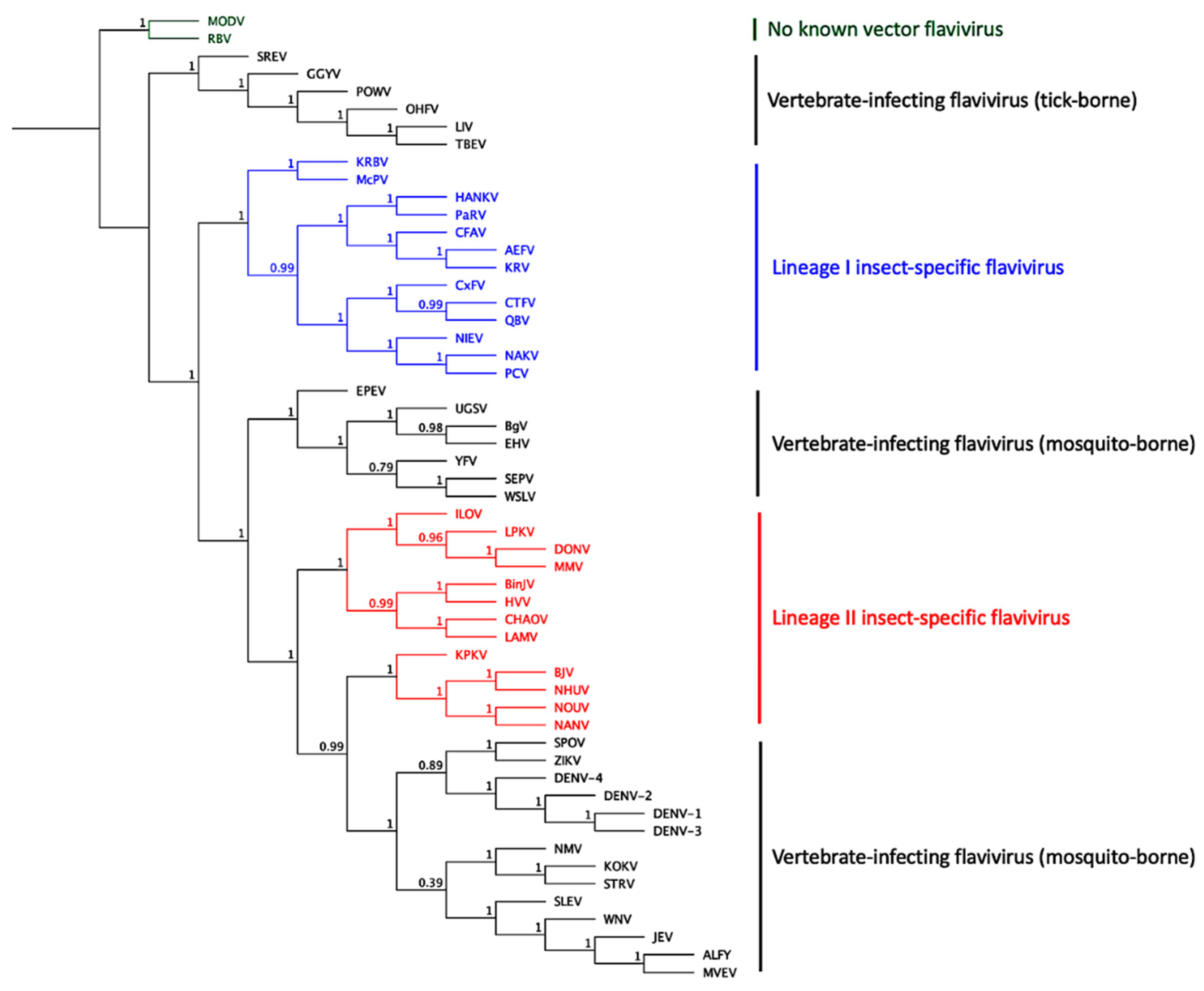
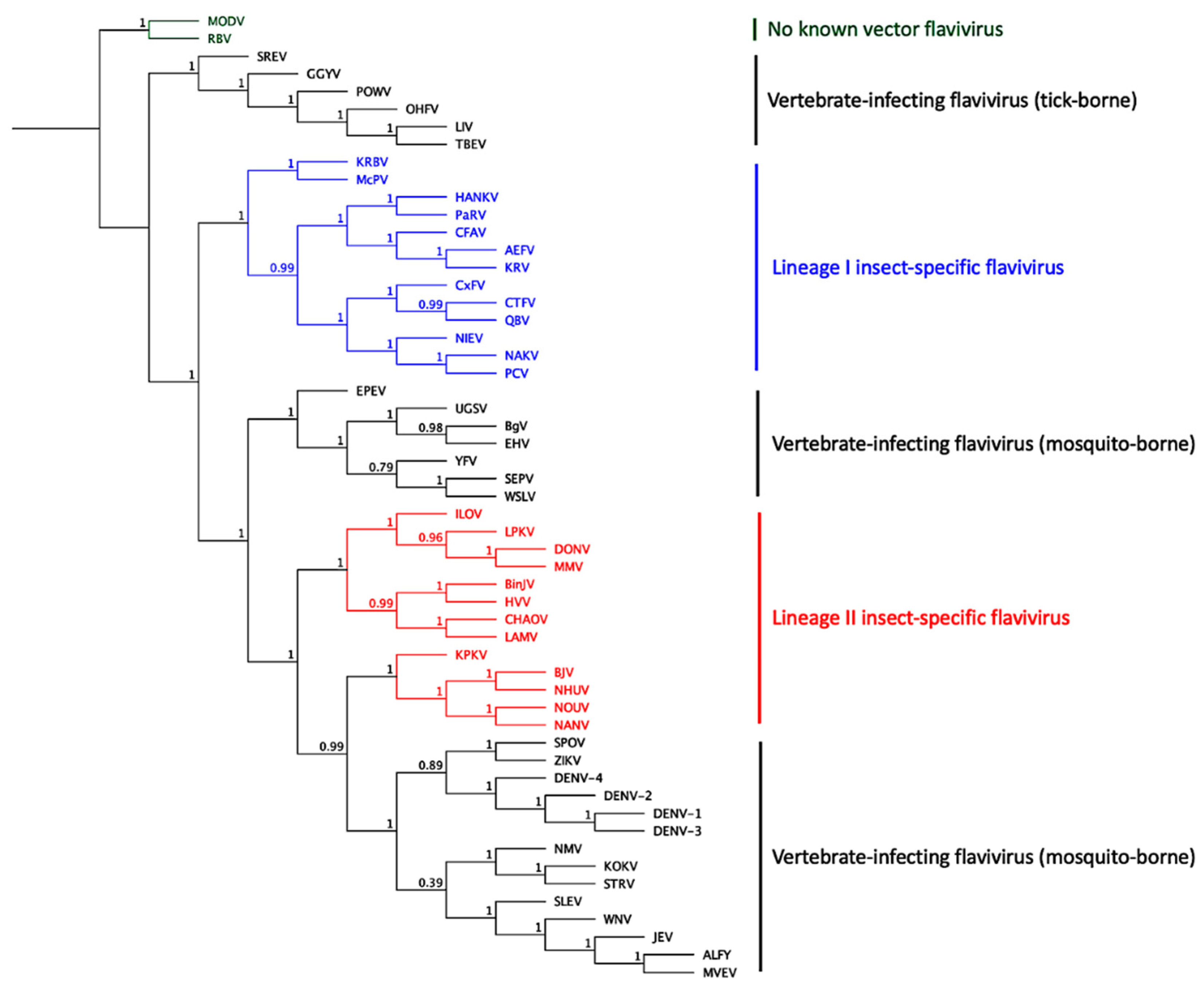
5. Insect-Specific Flaviviruses
6. Construction of Chimeric Flaviviruses Based on ISF Genomic Backbones
7. Host Range Restriction of ISFs
8. Antigenic and Structural Analysis of Chimeras
9. Use of Insect-Specific Flaviviruses as Vaccine Antigens for Pathogenic Flaviviruses
9.1. Vaccine Efficacy against ZIKV
9.2. Vaccine Efficacy against WNV
9.3. Vaccine Efficacy against DENV
9.4. Vaccine Efficacy against YFV
10. BinJ-VIF Chimeric Vaccines Are Highly Immunogenic
11. Safety and Regulatory Issues of ISF-Based Vaccines
12. Future Challenges for ISF-Based Vaccines
13. Application of ISF-Based Chimeric Viruses for Diagnostics
14. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Monath, T.P.; Cetron, M.S.; Teuwen, D.E. Yellow Fever Vaccine. In Vaccines; Plotkin, S.A., Orenstein, W.A., Offit, P.A., Eds.; Saunders, Elsevier: Philadelphia, PA, USA, 2008; pp. 959–1055. [Google Scholar]
- Halstead, S.B. Dengue. Lancet 2007, 370, 1644–1652. [Google Scholar] [CrossRef]
- Simon, L.V.; Kruse, B. Encephalitis, Japanese; Statpearls: Treasure Island, FL, USA, 2018. [Google Scholar]
- Mackenzie, J.S.; Gubler, D.J.; Petersen, L.R. Emerging Flaviviruses: The Spread and Resurgence of Japanese Encephelitis, West Nile and Dengue Viruses. Nat. Med. 2004, 10, 98–109. [Google Scholar] [CrossRef] [PubMed]
- World Health Organisation. Vaccines against Tick-Borne Encephalitis: Who Position Paper. Wkly. Epidemiol. Rec. 2011, 86, 241–256. [Google Scholar]
- Caminade, C.; Medlock, J.M.; Ducheyne, E.; McIntyre, K.M.; Leach, S.; Baylis, M.; Morse, A.P. Suitability of European Climate for the Asian Tiger Mosquito Aedes Albopictus: Recent Trends and Future Scenarios. J. R. Soc. Interface 2012, 9, 2708–2717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papa, A.; DAnis, K.; Baka, A.; Bakas, A.; Dougas, G.; Lytras, T.; Theocharopoulos, G.; Chrysagis, D.; Vassiliadou, E.; Kamaria, F.; et al. Ongoing Outbreak of West Nile Virus Infections in Geece, July–August 2010. Eurosurveillance 2010, 15, 19644. [Google Scholar] [CrossRef] [PubMed]
- Lanciotti, R.S.; Roehrig, J.T.; Deubel, V.; Smith, J.; Parker, M.; Steele, K.; Crise, B.; Volpe, K.E.; Crabtree, M.B.; Scherret, J.H.; et al. Origin of the West Nile Virus Responsible for an Outbreak of Encephalitis in the Northeastern United States. Science 1999, 286, 2333–2337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, K.O.; Mertens, E.; Desprès, P. West Nile Virus and Its Emergence in the United States of America. Vet. Res. 2010, 4, 41–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, T.; Hathaway, D.; Jennings, N.; Champ, D.; Chiang, Y.W.; Chu, H.J. Equine Vaccine for West Nile Virus. Dev. Biol. 2003, 114, 221–227. [Google Scholar]
- El Garch, H.; Minke, J.M.; Rehder, J.; Richard, S.; Toulemonde, C.E.; Dinic, S.; Andreoni, C.; Audonnet, J.C.; Nordgren, R.; Juillard, V. A West Nile Virus (Wnv) Recombinant Canarypox Virus Vaccine Elicits Wnv-Specific Neutralizing Antibodies and Cell-Mediated Immune Responses in the Horse. Vet. Immunol. Immunopathol. 2008, 123, 230–239. [Google Scholar] [CrossRef] [PubMed]
- Cortese, V.; Hankins, K.; Holland, R.; Syvrud, K. Serologis Responses of West Nile Virus Seronegative Mature Horses to West Nile Virus Vaccines. J. Equine. Vet. Sci. 2013, 33, 1101–1105. [Google Scholar] [CrossRef]
- Seino, K.K.; Long, M.T.; Gibbs, E.P.J.; Bowen, R.A.; Beachboard, S.E.; Humphrey, P.P.; Dixon, M.A.; Bourgeois, M.A. Comparative Efficacies of Three Commercially Available Vaccines against West Nile Virus (Wnv) in a Short-Duration Challenge Trial Involving a Equine Wnv Encephalitis Model. Clin. Vaccine Immunol. 2007, 14, 1465–1471. [Google Scholar] [CrossRef] [Green Version]
- Theiler, M.; Smith, H.H. The Use of Yellow Fever Virus Modified by in Vitro Cultivation for Human Immunization. J. Exp. Med. 1937, 65, 787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monath, T.P.; Lee, C.K.; Julander, J.G.; Brown, A.; Beasley, D.W.; Watts, D.M.; Hayman, E.; Guertin, P.; Makowiecki, J.; Crowell, J.; et al. Inactivated Yellow Fever 17d Vaccine: Development and Nonclinical Safety, Immunogenicity and Protective Activity. Vaccine 2010, 28, 3827–3840. [Google Scholar] [CrossRef] [PubMed]
- Galler, R.; Pugachev, K.; Santos, C.; Ocran, S.; Jabor, A.; Rodrigues, S.; Marchevsky, R.; Freire, M.; Almeida, L.; Cruz, A. Phenotypic and Molecular Analyses of Yellow Fever 17d Vaccine Viruses Associated with Serious Adverse Events in Brazil. Virology 2001, 290, 309–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Castro Ferreira, C.; Campi-Azevedo, A.C.; Peruhype-Magalhães, V.; Costa-Pereira, C.; Pires de Albuquerque, C.; Feitosa Muniz, L.; Yokoy de Souza, T.; Oliveira, A.C.V.; Martins-Filho, O.A.; Henrique da Mota, L.M. The 17d-204 and 17dd Yellow Fever Vaccines: An Overview of Major Similarities and Subtle Differences. Expert Rev. Vaccines 2018, 17, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Rajaiah, P. Kyasanur Forest Disease in India: Innovative Options for Intervention. Hum. Vaccines Immunother. 2019, 15, 2243–2248. [Google Scholar] [CrossRef]
- Yun, K.W.; Lee, H.J.; Kang, J.H.; Eun, B.W.; Kim, Y.J.; Kim, K.H.; Kim, N.H.; Hong, Y.J.; Kim, D.H.; Kim, H.M.; et al. Safety and Immunogenicty of a Freeze-Dried, Vero Cell-Culture Derived, Inactivated Japanese Encephalitis Vaccine (Kd-287, Encevac) Versus a Mouse Brain-Derived Inactivated Japanese Encephalitis Vaccine in Children: A Phase Iii, Multicenter, Double-Blinded, Randomized Trial. BMC Infect. Dis 2015, 15, 7. [Google Scholar]
- Schuller, E.; Jilma, B.; Voicu, V.; Golor, G.; Kollaritsch, H.; Kaltenböck, A.; Klade, C.; Tauber, E. Long-Term Immunogenicity of the New Vero Cell-Derived, Inactivated Japanese Encephalitis Virus Vaccine Ic51: Six and 12 Month Results of a Multicenter Follow-up Phase 3 Study. Vaccine 2008, 26, 4382–4386. [Google Scholar] [CrossRef]
- Ishikawa, T.; Yamanaka, A.; Konishi, E. A Review of Successful Flavivirus Vaccines and the Problems with Those Flaviviruses for Which Vaccines Are Not yet Available. Vaccine 2004, 32, 1326–1337. [Google Scholar] [CrossRef]
- Xin, Y.Y.; Ming, Z.G.; Peng, G.Y.; Jian, A.; Min, L.H. Safety of a Live-Attenuated Japanese Encephalitis Virus Vaccine (Sa14-14-2) for Children. Am. J. Trop. Med. Hyg. 1988, 319, 214–217. [Google Scholar] [CrossRef]
- Oliveira, A.; Selvaraj, K.; Tripathy, J.P.; Betodkar, U.; Cacodcar, J.; Wadkar, A. Kyasanur Forest Disease Vaccination Coverage and Its Perceived Barriers in Goa, India- a Mixed Methods Operational Research. PLoS ONE 2019, 14, e0226141. [Google Scholar] [CrossRef] [PubMed]
- Chambers, T.J.; Nestorowicz, A.; Mason, P.W.; Rice, C.M. Yellow Fever/Japanese Encephalitis Chimeric Viruses: Construction and Biological Properties. J. Virol. 1999, 73, 3095–3101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arroyo, J.; Miller, C.; Catalan, J.; Myers, G.A.; Ratterree, M.S.; Trent, D.W.; Monath, T.P. Chimerivax-West Nile Virus Live-Attenuated Vaccine:Preclinical Evaluation of Safety, Immunogenicity, and Afficacy. J. Virol. 2004, 78, 12497–12507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guy, B.; Barrere, B.; Malinowski, C.; Saville, M.; Teyssou, R.; Lang, J. From Research to Phase Iii: Preclinical, Industrial and Clinical Development of the Sanofi Pasteur Tetravalent Dengue Vaccine. Vaccine 2011, 29, 7229–7741. [Google Scholar] [CrossRef]
- Monath, T.P. Prospects for Development of a Vaccine against the West Nile Virus. Ann. N.Y. Acad 2001, 951, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Monath, T.P.; Guirakhoo, F.; Nichols, R.; Yoksan, S.; Schrader, R.; Murphy, C.; Blum, P.; Woodward, S.; McCarthy, K.; Mathis, D.; et al. Chimeric Live, Attenuated Vaccine against Japanese Encephalitis (Chimerivax-Je): Phase 2 Clinical Trials for Safety and Immunogenicity, Effect of Vaccine Dose and Schedule, and Memory Response to Challenge with Inactivated Japanese Encephalitis Antigen. J. Infect. Dis. 2003, 188, 1213–1230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monath, T.P.; McCarthy, K.; Bedford, P.; Johnson, C.T.; Nichols, R.; Yoksan, S.; Marchesani, R.; Knauber, M.; Wells, K.H.; Arroyo, J. Clinical Proof of Principle for Chimerivax Recombinant Live, Attenuated Vaccines against Flavivirus Infections. Vaccine 2002, 20, 1004–1018. [Google Scholar] [CrossRef]
- Guirakhoo, F.; Kitchener, S.; Morrison, D.; Forrat, R.; McCarthy, K.; Nichols, R.; Yoksan, S.; Duan, X.; Ermak, T.H.; Kanesa-Thasan, N. Live Attenuated Chimeric Yellow Fever Dengue Type 2 (Chimerivax-Den2) Vaccine: Phase I Clinical Trial for Safety and Immunogenicity: Effect of Yellow Fever Pre-Immunity in Induction of Cross Neutralising Antibody Responses to All. Hum. Vaccines 2006, 2, 60–67. [Google Scholar] [CrossRef] [Green Version]
- Guy, B.; Guirakhoo, F.; Barban, V.; Higgs, S.; Monath, T.P.; Lang, J. Preclinical and Clinical Development of Yfv 17d-Based Chimeric Vaccines against Dengue, West Nile and Japanese Encephaltis Viruses. Vaccine 2010, 28, 632–649. [Google Scholar] [CrossRef]
- Monath, T.; Soike, K.; Levenbook, I.; Zhang, Z.-X.; Arroyo, J.; Delagrave, S.; Myers, G.A.; Barrett, A.; Shope, R.; Ratterree, M. Recombinant, Chimeric Live, Attenuated Vaccine (Chimerivax) Incorporating the Envelope Genes of Japanese Encephalitis (Sa14-14-2) Virus and the Capsid and Nonstructural Genes of Yellow Fever (17d) Is Safe, Immunogenic and Protective in Non-Human Primates. Vaccine 1999, 17, 1869–1882. [Google Scholar] [CrossRef]
- Monath, T.; Soike, K.; Levenbook, I.; Zhang, Z.-X.; Ratterree, M.; Draper, K.; Barrett, A.; Nichols, R.; Weltzin, R.; Arroyo, J. Chimeric Yellow Fever Virus 17d-Japanese Encephalitis Virus Vaccine: Dose-Response Effectiveness and Extended Safety Testing in Rhesus Monkeys. J. Virol. 2000, 74, 1742–1751. [Google Scholar] [CrossRef] [Green Version]
- Pletnev, A.G.; St Claire, M.; Elkins, R.; Speicher, J.; Murphy, B.R.; Chanock, R.M. Molecularly Engineered Live-Attenuated Chimeric West Nile/Dengue Virus Vaccines Protect Rhesus Monkeys from West Nile Virus. Virology 2003, 314, 190–195. [Google Scholar] [CrossRef] [Green Version]
- Biswal, S.; Borja-Tabora, C.; Martinez Vargas, L.; Velásquez, H.; Alera, M.T.; Sierra, V.; Rodriguez-Arenales, E.J.; Yu, D.; Pujitha Wickramasinghe, V.; Duarte Moreira, J., Jr.; et al. Efficacy of a Tetravalent Dengue Vaccine in Healthy Children Aged 4–16 Years: A Randomised, Placebo-Controlled, Phase 3 Trial. Lancet 2020, 395, 1423–1433. [Google Scholar] [CrossRef]
- ClinicalTrials.gov. Efficacy, Safety and Immunogenicity of Takeda’s Tetravalent Dengue Vaccine (Tdv) in Healthy Children (Tides); NIH US National Library of Medicine. Available online: https://clinicaltrials.gov/ct2/show/NCT02747927?term=TIDES&draw=2&rank=7 (accessed on 21 July 2021).
- Driggers, R.W.; Ho, C.Y.; Korhonen, E.M.; Kuivanen, S.; Jaaskelainen, A.J.; Smura, T.; Rosenburg, A.; Hill, D.A.; DeBiasi, R.L.; Vezina, G.; et al. Zika Virus Infection with Prolonged Maternal Viremia and Fetal Brain Abnormalities. N. Engl. J. Med. 2016, 374, 2142–2151. [Google Scholar] [CrossRef] [PubMed]
- Mlakar, J.; Korva, M.; Tul, N.; Popović, M.; Poljšak-Prijatelj, M.; Mraz, J.; Kolenc, M.; Resman Rus, K.; Vipotnik, T.V.; Vodušek, V.F.; et al. Zika Virus Associated with Microcephaly. N. Engl. J. Med. 2016, 374, 951–958. [Google Scholar] [CrossRef]
- Sarno, M.; Sacramento, G.A.; Khouri, R.; do Rosário, M.S.; Costa, F.; Archanjo, G.; Santos, L.A.; Nery, N., Jr.; Vasilakis, N.; Ko, A.I.; et al. Zika Virus Infection and Stillbirths: A Case of Hydrops Fetalis, Hydranencephaly and Fetal Demise. PLoS Negl. Trop. Dis. 2016, 10, e0004517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ventura, C.V.; Maia, M.; Dias, N.; Ventura, L.O.; Belfort, R., Jr. Neurological and Ocular Findings in Infant without Microcephaly. Lancet 2016, 387, 2502. [Google Scholar] [CrossRef] [Green Version]
- Shan, C.; Xie, X.; Shi, P.-Y. Zika Virus Vaccine: Progress and Challenges. Cell Host Microbe 2018, 24, 12–17. [Google Scholar] [CrossRef] [Green Version]
- Belshe, R.B.; Edwards, K.M.; Vesikari, T.; Black, S.V.; Walker, R.E.; Hultquist, M.; Kemble, G.; Connor, E.M. Live Attenuated Versus Inactivated Influenza Vaccine in Infants and Young Children. N. Engl. J. Med. 2007, 356, 685–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavalcanti, D.P.; Salomão, M.A.; Lopez-Camelo, J.; Pessoto, M.A. Campinas Group of Yellow Fever Immunization during Pregnancy. Early Exposure to Yellow Fever Vaccine During Pregnancy. Trop. Med. Int. Health 2007, 12, 833–837. [Google Scholar] [CrossRef]
- Keller-Stanislawski, B.; Englund, J.A.; Kang, G.; Mangtani, P.; Neuzil, K.; Nohynek, H.; Pless, R.; Lambach, P.; Zuber, P. Safety of Immunization During Pregnanxy: A Review of the Evidence of Selected Inactivated and Live Attenuated Vaccines. Vaccine 2014, 32, 7057–7064. [Google Scholar] [CrossRef]
- Staples, J.E.; Gershman, M.; Fischer, M.; Centers for Disease Control and Prevention (CDC). Yellow Fever Vaccine: Recommendations of the Advisory Committee on Immunization Practices. MMWR Recomm. Rep. 2010, 59, 1–27. [Google Scholar] [PubMed]
- Laris-González, A.; Bernal-Serrano, D.; Jarde, A.; Kampmann, B. Safety of Administering Live Vaccines During Pregnancy: A Systematic Review of Meta-Analysis of Pregnancy Outcomes. Vaccine 2020, 8, 124. [Google Scholar] [CrossRef] [Green Version]
- Shan, C.; Muruato, A.E.; Nunes, B.T.; Luo, H.; Xie, X.; Medeiros, D.B.; Wakamiya, M.; Tesh, R.B.; Barrett, A.D.; Wang, T. A Live-Attenuated Zika Virus Vaccine Candidate Induces Sterilizing Immunity in Mouse Models. Nat. Med. 2017, 23, 763–767. [Google Scholar] [CrossRef]
- Xie, X.; Yang, Y.; Muruato, A.E.; Zou, J.; Shan, C.; Nunes, B.T.; Medeiros, D.B.; Vasconcelos, P.F.; Weaver, S.C.; Rossi, S.L.; et al. Understanding Zika Virus Stability and Developing a Chimeric Vaccine through Functional Analysis. mBio 2017, 8, e02134-16. [Google Scholar] [CrossRef] [Green Version]
- Aliota, M.T.; Bassit, L.; Bradrick, S.S.; Cox, B.; Garcia-Blanco, M.A.; Gavegnano, C.; Friedrich, T.C.; Golos, T.G.; Griffin, D.E.; Haddow, A.D.; et al. Zika in the Americas, Year 2: What Have We Learned? What Gaps Remain? A Report from the Global Virus Network. Antivir. Res. 2017, 144, 223–246. [Google Scholar] [CrossRef] [PubMed]
- Wilder-Smith, A.; Vannice, K.; Durbin, A.; Hombach, J.; Thomas, S.J.; Thevarjan, I.; Simmons, C.P. Zika Vaccines and Therpeutics: Landscape Alaysis and Challenges Ahead. BMC Med. 2018, 16, 84–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrett, A.D. Current Status of Zika Vaccine Development: Zika Vaccines Advance into Clinical Evaluation. NPJ Vaccines 2018, 3, 24. [Google Scholar] [CrossRef]
- Cornish, E.F.; Filipovic, I.; Åsenius, F.; Williams, D.J.; McDonnell, T. Innate Immune Responses to Acute Viral Infection during Pregnancy. Front. Immunol. 2020, 11, 2404. [Google Scholar] [CrossRef] [PubMed]
- Guabiraba, R.; Ryffel, B. Dengue Virus Infection: Current Concepts in Immune Mechanisms and Lessons from Murine Models. Immunol. Cell Biol. 2013, 141, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Katzelnick, L.C.; Gresh, L.; Halloran, M.E.; Mercado, J.C.; Kuan, G.; Gordon, A.; Balmaseda, A.; Harris, E. Antibody-Dependent Enhancement of Severe Dengue Disease in Humans. Science 2017, 358, 929–932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Men, R.; Bray, M.; Clark, D.C.; Chanock, R.M.; Lai, C.J. Dengue Type 4 Virus Mutants Containing Deletions in the 3′ Noncoding Region of the Rna Genome: Analysis of Growth Restriction in Cell Culture and Altered Viremia Pattern and Immunogenicity in Rhesus Monkeys. J. Virol. 1996, 70, 3930–3937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blaney, J.E.; Durbin, A.P.; Murphy, B.R.; Whitehead, S.S. Development of a Live Attenuated Dengue Virus Vaccine Using Reverse Genetics. Viral Immunol. 2006, 19, 10–32. [Google Scholar] [CrossRef]
- Kitchener, S.; Nissen, M.; Nasveld, P.; Forrat, R.; Yoksan, S.; Lang, J.; Saluzzo, J.F. Immunogenicity and Safety of Two Live-Attenuated Tetravalent Dengue Vaccine Formulations in Healthy Australian Adults. Vaccine 2006, 24, 1238–1241. [Google Scholar] [CrossRef]
- Sabchareon, A.; Lang, J.; Chanthavanich, P.; Yoksan, S.; Forrat, R.; Attanath, P.; Sirivichayakul, C.; Pengsaa, K.; Pojjaroen-Anant, C.; Chambonneau, L.; et al. Safety and Immunogenicity of a Three Dose Regimen of Two Tetravalent Live-Attenuated Dengue Vaccines in Five-to Twelve-Year-Old Thai Children. Pediatr. Infect. Dis. J. 2004, 23, 1238–1241. [Google Scholar] [CrossRef]
- FFatima, K.; Syed, N.I. Dengvaxia Controvery: Impact on Vaccine Hesitancy. J. Glob. Health 2018, 8, 020312. [Google Scholar]
- Dejnirattisai, W.; Supasa, P.; Wongwiwat, W.; Rouvinski, A.; Barba-Spaeth, G.; Duangchinda, T.; Sakuntabhai, A.; Cao-Lormeau, V.M.; Malasit, P.; Rey, F.A.; et al. Dengue Virus Sero-Cross-Reactivity Drives Antibody-Depenent Enhancement of Infection with Zika Virus. Nat. Immunol. 2016, 17, 1102–1108. [Google Scholar] [CrossRef]
- Bolling, B.G.; Olea-Popelka, F.J.; Eisen, L.; Moore, C.G.; Blair, C.D. Transmission Dynamics of an Insect-Specific Flavivirus in a Naturally Infected Culex Pipiens Laboratory Colony and Effects of Co-Infection on Vector Competence for West Nile Virus. Virology 2012, 427, 90–97. [Google Scholar] [CrossRef] [Green Version]
- Goenaga, S.; Kenney, J.L.; Duggal, N.K.; Delorey, M.; Ebel, G.D.; Zhang, B.; Levis, S.C.; Enria, D.A.; Brault, A.C. Potential for Co-Infection of a Mosquito-Specific Flavivirus, Nhumirim Virus, to Block West Nile Virus Transmission in Mosquitoes. Viruses 2015, 7, 5801–5812. [Google Scholar] [CrossRef] [Green Version]
- Hall-Mendelin, S.; McLean, B.J.; Bielefeldt-Ohmann, H.; Hobson-Peters, J.; Hall, R.A.; van den Hurk, A.F. The Insect-Specific Palm Creek Virus Modulates West Nile Virusinfection in and Transmission by Australian Mosquitoes. Parasites Vectors 2016, 9, 414–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hobson-Peters, J.; Yam, A.W.Y.; Lu, J.W.F.; Setoh, Y.X.; May, F.J.; Kurucz, N.; Walsh, S.; Prow, N.A.; Davis, S.S.; Weir, R.; et al. A New Insect-Specific Flavivirus from Northern Australia Suppresses Replication of West Nile Virus and Murray Valley Enephalitis Virus in Co-Infected Mosquito Cells. PLoS ONE 2013, 8, e56534. [Google Scholar]
- Newman, C.M.; Cerutti, F.; Anderson, T.K.; Hamer, G.L.; Walker, E.D.; Kitron, U.D.; Ruiz, M.O.; Brawn, J.D.; Goldberg, T.L. Culex Flavivirus and West Nile Virus Mosquito Coinfection and Positive Ecological Association in Chicago, United States. Vector Borne Zoonotic Dis. 2011, 11, 1099–1105. [Google Scholar] [CrossRef] [Green Version]
- Romo, H.; Kenney, J.L.; Blitvich, B.J.; Brault, A.C. Restriction of Zika Virus Infection and Transmission in Aedes Aegypti Mediated by an Insect-Specific Flavivirus. Emerg. Microbes Infect. 2018, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Choo, J.J.; Vet, L.J.; McMillan, C.L.; Harrison, J.J.; Scott, C.A.; Depelsenaire, A.C.; Fernando, G.J.; Watterson, D.; Hall, R.A.; Young, P.R.; et al. A Chimeric Dengue Virus Vaccine Candidate Delivered by High Density Microarray Patches Protects against Infection in Mice. NPJ Vaccines 2021, 7, 66–76. [Google Scholar] [CrossRef]
- Hazlewood, J.E.; Rawle, D.J.; Tang, B.; Yan, K.; Vet, L.J.; Nakayama, E.; Hobson-Peters, J.; Hall, R.A.; Suhrbier, A. A Zika Vaccine Generated Using the Chimeric Insect-Specific Binjari Virus Platform Protects against Fetal Brain Infection in Pregnant Mice. Vaccines 2020, 8, 496. [Google Scholar] [CrossRef] [PubMed]
- Hobson-Peters, J.; Harrison, J.J.; Watterson, D.; Hazlewood, J.E.; Vet, L.J.; Newton, N.D.; Warrilow, D.; Colmant, A.M.; Taylor, C.; Huang, B.; et al. A Recombinant Platform for Flavivirus Vaccines and Diagnostics Using Chimeras of a New Insect-Specific Virus. Sci. Transl. Med. 2019, 11, eaax7888. [Google Scholar] [CrossRef]
- Vet, L.J.; Setoh, Y.X.; Amarilla, A.A.; Habarugira, G.; Suen, W.W.; Newton, N.D.; Harrison, J.J.; Hobson-Peters, J.; Hall, R.A.; Bielefeldt-Ohmann, H. Protective Efficacy of a Chimeric Insect-Specific Flavivirus Vaccine against West Nile Virus. Vaccines 2020, 8, 258. [Google Scholar] [CrossRef] [PubMed]
- Yan, K.; Vet, L.J.; Tang, B.; Hobson-Peters, J.; Rawle, D.J.; Le, T.T.; Larcher, T.; Hall, R.A.; Suhrbier, A. A Yellow Fever Virus 17d Infection and Disease Mouse Model Used to Evaluate a Chimeric Binjari-Yellow Fever Virus Vaccine. Vaccines 2020, 8, 368. [Google Scholar] [CrossRef]
- Stollar, V.; Thomas, V.L. An Agent in the Aedes Aegypti Cell Line (Peleg) Which Causes Fusion of Aedes Albopictus Cells. Virology 1975, 64, 367–377. [Google Scholar] [CrossRef]
- Cammisa-Parks, H.; Cisar, L.A.; Kane, A.; Stollar, V. The Complete Nucleotide Sequence of Cell Fusing Agent (Cfa): Homology between the Nonstructural Proteins Encoded by Cfa and the Nonstructurakl Proteins Encoded by Arthropod-Borne Flaviviruses. Virology 1992, 189, 511–524. [Google Scholar] [CrossRef]
- Sang, R.C.; Gichogo, A.; Gachoya, J.; Dunster, M.D.; Ofula, V.; Hunt, A.R.; Crabtree, M.B.; Miller, B.R.; Dunster, L.M. Isolation of a New Flavivirus Related to Cell Fusing Agent Virus (Cfav) from Field-Collected Flood-Water Aedes Mosquitoes Sampled from a Dambo in Central Kenya. Arch. Virol. 2003, 148, 1085–1093. [Google Scholar] [CrossRef]
- Blitvich, B.J.; Firth, A.E. Insect-Specific Flaviviruses: A Systematic Review of Their Discovery, Host Range, Mode of Transmission, Superinfection Exclusion Potential and Genomic Organization. Viruses 2015, 7, 1927–1959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, R.A.; Bielefeldt-Ohmann, H.; McLean, B.J.; O’Brien, C.A.; Colmant, A.M.; Piyasena, T.B.; Harrison, J.J.; Newton, N.D.; Barnard, R.T.; Prow, N.A.; et al. Commensal Viruses of Mosquitoes: Host Restriction, Transmission, and Interaction with Arboviral Pathogens. Evol. Bioinform. Online 2016, 12, 35–44. [Google Scholar] [CrossRef] [PubMed]
- McLean, B.J.; Hobson-Peters, J.; Webb, C.E.; Watterson, D.; Prow, N.A.; Nguyen, H.D.; Hall-Mendelin, S.; Warrilow, D.; Johansen, C.A.; Jansen, C.C.; et al. A Novel Insect-Specific Flavivirus Replicates Only in Aedes-Derived Cells and Persists at High Prevalence in Wild Aedes Vigilax Populations in Syndey, Australia. Virology 2015, 486, 272–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colmant, A.M.G.; Hobson-Peters, J.; Bielefeldt-Ohmann, H.; van den Hurk, A.F.; Hall-Mendelin, S.; Chow, W.K.; Johansen, C.A.; Fros, J.; Simmonds, P.; Watterson, D.; et al. A New Clade of Insect-Specific Flaviviruses from Australian Anopheles Mosquitoes Displays Species-Specific Host Restriction. mSphere 2017, 2, e00262-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, J.J.; Hall-Mendelin, S.; Hall, R.A. Detection of Insect-Specific Flaviviruses in Australian Mosquitoes. 2021. Manuscript under Preparation. [Google Scholar]
- Zakrzewski, M.; Rašić, G.; Darbro, J.; Krause, L.; Poo, Y.S.; Filipović, I.; Parry, R.; Asgari, S.; Devine, G.; Suhrbier, A. Mapping the Virome in Wild-Caught Aedes Aegpyti from Cairns and Bangkok. Sci. Rep. 2018, 8, 4690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, J.J.; Hobson-Peters, J.; Colmant, A.M.G.; Koh, J.; Newton, N.D.; Warrilow, D.; Bielefeldt-Ohmann, H.; Piyasena, T.B.H.; O’Brien, C.A.; Vet, L.J.; et al. Antigenic Characterisation of New Lineage Ii Insect-Specific Flaviviruses in Australian Mosquitoes and Identification of Host Restriction Factors. mSphere 2020, 5, e00095-20. [Google Scholar] [CrossRef] [PubMed]
- Bolling, B.G.; Vasilakis, N.; Guzman, H.; Widen, S.G.; Wood, T.G.; Popov, V.L.; Thangamani, S.; Tesh, R.B. Insect-Specific Viruses Detected in Laboratory Mosquito Colonies and Their Potential Implications for Experiments Evaluating Arbovirus Vector Competence. Am. J. Trop. Med. Hyg. 2015, 92, 422–428. [Google Scholar] [CrossRef] [Green Version]
- Cook, S.; Bennett, S.N.; Holmes, E.C.; De Chesse, R.; Moureau, G.; De Lamballerie, X. Isolation of a New Strain of the Flavivirus Cell Fusing Agent Virus in a Natural Mosquito Population from Puerto Rico. J. Gen. Virol. 2006, 87, 735–748. [Google Scholar] [CrossRef] [PubMed]
- Espinoza-Gómez, F.; López-Lemus, A.U.; Rodriguez-Sanchez, I.P.; Martinez-Fierro, M.L.; Newton-Sánchez, O.A.; Chávez-Flores, E.; Delgado-Enciso, I. Detection of Sequences from a Potentially Novel Strain of Cell Fusing Agent Virus in Mexican Stegomyia (Aedes) Aegypti Mosquitoes. Arch. Virol. 2011, 156, 1263–1267. [Google Scholar] [CrossRef] [PubMed]
- Calzolari, M.; Zé-Zé, L.; Vázquez, A.; Seco, M.P.S.; Amaro, F.; Dottori, M. Insect-Specific Flaviviruses: A Worldwide Widespread Group of Viruses Only Detected in Insects. Infect. Genet. Evol. 2016, 40, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Grisenti, M.; Vazquez, A.; Herrero, L.; Cuevas, L.; Perez-Pastrana, E.; Arnoldi, D.; Rosa, R.; Capelli, G.; Tenorio, A.; Sanchez-Seco, M.P.; et al. Wide Spread Detection of Aedes Flavivirus in North-Eastern Italy, a European Hot-Spot of Emerging Mosquito-Borne Diseases. J. Gen. Virol. 2015, 96, 420–430. [Google Scholar] [CrossRef] [Green Version]
- Haddow, A.D.; Guzman, H.; Popov, V.L.; Wood, T.G.; Widen, S.G.; Haddow, A.D.; Tesh, R.B.; Weaver, S.C. First Isolation of Aedes Flavivirus in the Western Hemisphere and Evidence of Vertical Transmission in the Mosquito Aedes (Stegomyia) Albopictus (Diptera: Culcidae). Virology 2013, 440, 134–139. [Google Scholar] [CrossRef] [Green Version]
- Hoshino, K.; Isawa, H.; Tsuda, Y.; Sawabe, K.; Kobayashi, M. Isolation and Characterization of a New Insect Flavivirus from Aedes Albopictus and Aedes Flavopictus Mosquitoes in Japan. Virology 2009, 391, 119–129. [Google Scholar] [CrossRef] [Green Version]
- Roiz, D.; Vázquez, A.; Rosso, F.; Arnoldi, D.; Girardi, M.; Cuevas, L.; Perez-Pastrana, E.; Sánchez-Seco, M.P.; Tenorio, A.; Rizzoli, A. Detection of a New Insect-Specific Flavivirus and Isolation of Aedes Flavivirus in Northern Italy. Parasites Vectors 2012, 5, 223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolling, B.G.; Eisen, L.; Moore, C.G.; Blair, C.D. Insect-Specific Flaviviruses from Culex Mosquitoes in Colorado, with Evidence of Vertical Transmission. Am. J. Trop. Med. Hyg. 2011, 85, 169–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cook, S.; Moureau, G.; Harbach, R.E.; Mukwaya, L.; Goodger, K.; Ssenfuka, F.; Gould, E.; Holmes, E.C.; de Lamballerie, X. Isolation of a Novel Species of Flavivirus and a New Strain of Culex Flavivirus (Flaviviridae) from a Natural Mosquito Population in Uganda. J. Gen. Virol. 2009, 90, 2669–2678. [Google Scholar] [CrossRef]
- Farfan-Ale, J.A.; Loroño-Pino, M.A.; Garcia-Rejon, J.E.; Hovav, E.; Powers, A.M.; Lin, M.; Dorman, K.S.; Platt, K.B.; Bartholomay, L.C.; Soto, V.; et al. Detection of Rna from a Novel West Nile-Like Virus and High Prevalence of an Insect-Specific Flavivirus in Mosquitoes in the Yucatan Peninsula of Mexico. Am. J. Trop. Med. Hyg. 2009, 80, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Goenaga, S.; Fabbri, C.M.; García, J.B.; Rondán, J.C.; Gardenal, N.; Calderón, G.E.; Enria, D.A.; Levis, S.M. New Strains of Culex Flavivirus Isolated in Argentina. J. Med. Entomol. 2014, 51, 900–906. [Google Scholar] [CrossRef]
- Huanyu, W.; Haiyan, W.; Shihong, F.; Guifang, L.; Hong, L.; Xiaoyan, G.; Lizhi, S.; Rayner, S.; Aiqiang, X.; Guodong, L. Isolation and Identification of a Distinct Strain of Culex Flavivirus from Mosquitoes Collected in Mainland China. Virology 2012, 359, 405–414. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.Y.; Guzman, H.; Bueno, R., Jr.; Dennett, J.A.; Auguste, A.J.; Carrington, C.V.; Popov, V.L.; Weaver, S.C.; Beasley, D.W.; Tesh, R.B. Characterisation of Culex Flavivirus (Flaviviridae) Strains Isolated from Mosquitoes in the United States and Trinidad. Virology 2009, 386, 154–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales-Betoulle, M.E.; Pineda, M.M.; Sosa, S.M.; Panella, N.; Cordon-Rosales, C.; Komar, N.; Powers, A.; Johnson, B.W. Culex Flavivirus Isolates from Mosquitoes in Guatemala. J. Med. Entomol. 2008, 45, 1187–1190. [Google Scholar] [CrossRef]
- Saiyasombat, R.; Dorman, K.S.; Garcia-Rejon, J.E.; Loroño-Pino, M.A.; Farfan-Ale, J.A.; Blitvich, B.J. Isolation and Sequence Analysis of Culex Flavivirus from Culex Interrogator and Culex Quinquefasciatus in the Yucatan Peninsula of Mexico. Arch. Virol. 2010, 155, 983–986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Junglen, S.; Korries, M.; Grasse, W.; Wieseler, J.; Kopp, A.; Hermanns, K.; León-Juárez, M.; Drosten, C.; Kümmerer, B.M. Host Range Restriction of Insect-Specific Flaviviruses Occurs at Several Levels of the Viral Life Cycle. mSphere 2017, 2, e00375-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolodziejek, J.; Pachler, K.; Bin, H.; Mendelson, E.; Shulman, L.; Orshan, L.; Nowotny, N. Barkedji Virus, a Novel Mosquito-Borne Flavivirus Identified in Culex perexiguus Mosquitoes, Israek, 2011. J. Gen. Virol. 2013, 94, 2449–2457. [Google Scholar] [CrossRef]
- Huhtamo, E.; Cook, S.; Moureau, G.; Uzcátegui, N.Y.; Sironen, T.; Kuivanen, S.; Putkuri, N.; Kurkela, S.; Harbach, R.E.; Firth, A.E.; et al. Novel Flaviviruses from Mosquitoes: Mosquito-Specific Evolutionary Lineages within the Phylogenetic Group of Mosquito-Borne Flaviviruses. Virology 2014, 464, 320–329. [Google Scholar] [CrossRef] [Green Version]
- Huhtamo, E.; Putkuri, N.; Kurkela, S.; Manni, T.; Vaheri, A.; Vapalahti, O.; Uzcátegui, N.Y. Characterization of a Novel Flavivirus from Mosquitoes in Northern Europe That Is Related to Mosquito-Borne Flaviviruses of the Tropics. J. Virol. 2009, 83, 9532–9540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charles, J.; Tangudu, C.S.; Firth, A.E.; Blitvich, B.J. Complete Genome Sequences of Two Insect-Specific Flaviviruses. Arch. Virol. 2017, 162, 3913–3917. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, F.; Cerutti, F.; Ballardini, M.; Mosca, A.; Vitale, N.; Radaelli, M.C.; Desiato, R.; Prearo, M.; Pautasso, A.; Casalone, C.; et al. Molecular Characterisaton of Flaviviruses from Field-Collected Mosquitoes in Northernwestern Italy, 2011–2012. Parasites Vectors 2014, 7, 395–406. [Google Scholar] [CrossRef] [Green Version]
- Vázquez, A.; Sánchez-Seco, M.P.; Palacios, G.; Molero, F.; Reyes, N.; Ruiz, S.; Aranda, C.; Marqués, E.; Escosa, R.; Moreno, J.; et al. Novel Flaviviruses Detected in Different Species of Mosquitoes in Spain. Vector Borne Zoonotic Dis. 2012, 12, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Guzman, H.; Contreras-Gutierrez, M.A.; Travassos da Rosa, A.P.A.; Nunes, M.R.T.; Cardoso, J.F.; Popov, V.L.; Young, K.I.; Savit, C.; Wood, T.G.; Widen, S.G.; et al. Characterization of Three New Insect-Specific Flaviviruses: Their Relationship to the Mosquito-Borne Flavivirus Pathogens. Am. J. Trop. Med. Hyg. 2018, 98, 410–419. [Google Scholar] [CrossRef]
- Junglen, S.; Drosten, C. Virus Discovery and Recent Insights into Virus Diversity in Arthropods. Curr. Opin. Microbiol. 2013, 16, 507–513. [Google Scholar] [CrossRef] [PubMed]
- Piyasena, T.B.; Setoh, Y.X.; Hobson-Peters, J.; Newton, N.D.; Bielefeldt-Ohmann, H.; McLean, B.J.; Vet, L.J.; Khromykh, A.A.; Hall, R.A. Infectious Dnas Derived from Insect-Specific Flavivirus Genomes Enable Identification of Pre- and Post-Entry Mechanisms of Host Restriction in Vertebrate Cells. Sci. Rep. 2017, 7, 2940. [Google Scholar] [CrossRef]
- Piyasena, T.B.; Newton, N.D.; Hobson-Peters, J.; Vet, L.J.; Setoh, Y.X.; Bielefeldt-Ohmann, H.; Khromykh, A.A.; Hall, R.A. Chimeric Viruses of the Insect-Specific Flavivirus Palm Creek with Structural Proteins of Vertebrate-Infecting Flaviviruses Identify Barriers to Replication of Insect-Specific Flaviviruses in Vertebrate Cells. J. Gen. Virol. 2019, 100, 1580–1586. [Google Scholar] [CrossRef] [PubMed]
- Amarilla, A.A.; Sng, J.D.J.; Parry, R.; Deerain, J.M.; Potter, J.R.; Setoh, Y.X.; Rawle, D.J.; Le, T.T.; Modhiran, N.; Wang, X.; et al. A Versatile Reverse Genetics Platform for Sars-Cov-2 and Other Positive-Strand Rna Viruses. Nat. Commun. 2021, 12, 3431. [Google Scholar] [CrossRef]
- Colmant, A.M.; Hobson-Peters, J.; Slijkerman, T.A.; Harrison, J.J.; Pijlman, G.P.; van Oers, M.M.; Simmonds, P.; Hall, R.A.; Fros, J.J. Insect-Specific Flavivirus Replication in Mammalian Cells Is Inhibited by Physiological Temperature and the Zinc-Finger Antiviral Protein. Viruses 2021, 13, 573. [Google Scholar] [CrossRef]
- Tangudu, C.S.; Charles, J.; Nunez-Avellaneda, D.; Hargett, A.M.; Brault, A.C.; Blitvich, B.J. Chimeric Zika Viruses Containing Structural Protein Genes of Insect-Specific Flaviviruses Cannot Replicate in Vertebrate Cells Due to Entry and Post-Translational Restrictions. Virology 2021, 559, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Aliota, M.T.; Kramer, L.D. Replication of West Nile Virus, Rabensburg Lineage in Mammalian Cells Is Restricted by Temperature. Parasites Vectors 2012, 5, 293. [Google Scholar] [CrossRef] [Green Version]
- Colmant, A.M.; Bielefeldt-Ohmann, H.; Hobson-Peters, J.; Suen, W.W.; O’Brien, C.A.; van den Hurk, A.F.; Hall, R.A. A Newly Discovered Flavivrus in the Yellow Fever Virus Group Displays Restricted Replication in Vertebrates. J. Gen. Virol. 2016, 97, 1087–1093. [Google Scholar] [CrossRef] [PubMed]
- Colmant, A.M.G.; Bielefeldt-Ohmann, H.; Vet, L.J.; O’Brien, C.A.; Bowen, R.A.; Hartwig, A.E.; Davis, S.; Piyasena, T.B.H.; Habarugira, G.; Harrison, J.J.; et al. NS4/5 Mutations Enhance Flavivirus Bamaga Virus Infectivity and Pathogenicity in Vitro and in Vivo. PLoS Negl. Trop. Dis. 2020, 14, e0008166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prow, N.A.; Tang, B.; Gardner, J.; Le, T.T.; Taylor, A.; Poo, Y.S.; Nakayama, E.; Hirata, T.D.; Nakaya, H.I.; Slonchak, A.; et al. Lower Temperatures Reduce Type I Interferon Activity and Promote Alphaviral Arthritis. PLoS Pathog. 2017, 13, e1006788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardy, J.M.; Newton, N.D.; Modhiran, N.; Scott, C.A.; Venugopal, H.; Vet, L.J.; Young, P.R.; Hall, R.A.; Hobson-Peters, J.; Coulibaly, F.; et al. A Unified Route for Flavivirus Structures Uncovers Essential Pocket Factors Conserved across Pathogenic Viruses. Nat. Commun. 2021, 12, 3266. [Google Scholar] [CrossRef]
- Erasmus, J.H.; Seymour, R.L.; Kaelber, J.T.; Kim, D.Y.; Leal, G.; Sherman, M.B.; Frolov, I.; Chiu, W.; Weaver, S.C.; Nasar, F. Novel Insect-Specific Eilat Virus-Based Chimeric Vaccine Candidates Provide Durable, Mono- and Multivalent, Sinlge-Dose Protection against Lethal Alphavirus Challenge. J. Virol. 2018, 92, e01274-17. [Google Scholar] [CrossRef] [Green Version]
- Erasmus, J.H.; Auguste, A.J.; Kaelber, J.T.; Luo, H.; Rossi, S.L.; Fenton, K.; Leal, G.; Kim, D.Y.; Chiu, W.; Wang, T.; et al. A Chikungunya Fever Vaccine Utilizing an Insect-Specific Virus Platform. Nat. Med. 2017, 23, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Forster, A.H.; Witham, K.; Depelsenaire, A.C.; Veitch, M.; Wells, J.W.; Wheatley, A.; Pryor, M.; Lickliter, J.D.; Francis, B.; Rockman, S.; et al. Safety, Tolerability and Immunogenicity of Influenza Vaccination with a High-Density Microarray Patch: Results from a Randomized, Controlled Phase I Clinical Trial. PLoS Med. 2020, 17, e1003024. [Google Scholar] [CrossRef] [PubMed]
- Muller, D.A.; Depelsenaire, A.C.; Shannon, A.E.; Watterson, D.; Corrie, S.R.; Owens, N.S.; Agyei-Yeboah, C.; Cheung, S.; Zhang, J.; Fernando, G.J.; et al. Efficient Delivery of Dengue Virus Subunit Vaccines to the Skin by Microprojection Arrays. Vaccine 2019, 7, 189. [Google Scholar] [CrossRef] [Green Version]
- Muller, D.A.; Fernando, G.J.; Owens, N.S.; Agyei-Yeboah, C.; Wei, J.C.; Depelsenaire, A.C.; Forster, A.; Fahey, P.; Weldon, W.C.; Oberste, M.S.; et al. High-Density Microprojection Array Delivery to Rat Skin of Low Doses of Trivalent Inactivated Poliovirus Vaccine Elicits Potent Neutralising Antibody Responses. Sci. Rep. 2017, 7, 12644. [Google Scholar] [CrossRef]
- Muller, D.A.; Pearson, F.E.; Fernando, G.J.; Agyei-Yeboah, C.; Owens, N.S.; Corrie, S.R.; Crichton, M.L.; Wei, J.C.; Weldon, W.C.; Oberste, M.S.; et al. Inactivated Poliovirus Type 2 Vaccine Delivered to Rat Skin Via High Density Microprojection Array Elicits Potent Neutralising Antibody Responses. Sci. Rep. 2016, 6, 22094. [Google Scholar] [CrossRef]
- Dalsgaard, K. Saponin Adjuvants. 3. Isolation of a Substance from Quillaja Saponaria Molina with Adjuvant Activiyt in Food- and-Mouth Disease Vaccines. Arch. Gesamte. Virusforsch. 1974, 44, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Metz, S.W.; Thomas, A.; White, L.; Stoops, M.; Corten, M.; Hannemann, H.; De Silva, A.M. Dengue Virus-Like Particles Mimic the Antigenic Properties of the Infectious Dengue Envelope. Virology 2018, 15, 60. [Google Scholar] [CrossRef] [PubMed]
- Suphatrakul, A.; Yasanga, T.; Keelapang, P.; Sriburi, R.; Roytrakul, T.; Pulmanausahakul, R.; Utaipat, U.; Kawilapan, Y.; Puttikhunt, C.; Kasinrerk, W.; et al. Generation and Preclinical Immunogenicity Study of Dengue Type 2 Virus-Like Particles Derived from Stably Transfected Mosquito Cells. Vaccine 2015, 33, 5613–5622. [Google Scholar] [CrossRef]
- Chang, T.-H.; Liao, C.-L.; Lin, Y.-L. Flavivirus Induces Interferon-Beta Geneexpression through a Pathway Involving Rig-I-Dependent Irf-3 and Pi3k-Dependent Nf-Kb Activation. Microbes Infect 2006, 8, 157–171. [Google Scholar] [CrossRef] [PubMed]
- Kell, A.M.; Gale, M. Rig-I in Rna Virus Recognition. Virology 2015, 479–480, 110–121. [Google Scholar] [CrossRef] [Green Version]
- Hacker, K.; White, L.; de Silva, A.M. N-Linked Glycans on Dengue Viruses Grown in Mammalian and Insect Cells. J. Gen. Virol. 2009, 90, 2097–2106. [Google Scholar] [CrossRef] [PubMed]
- Mason, P.W. Maturation of Japanese Encephalitis Virus Glycoproteins Produced by Infected Mammalian and Mosquito Cells. Virology 1989, 169, 354–364. [Google Scholar] [CrossRef]
- Bhatt, T.R.; Crabtree, M.B.; Guirakhoo, F.; Monath, T.P.; Miller, B.R. Growth Characteristics of the Chimeric Japanese Encephalitis Virus Vaccine Candidate, Chimerivax-Je (Yf/Je Sa14-14-2) in Culex Tritaeniorhynchus, Aedes Albopictus and Aedes Aegypti Mosquitoes. Am. J. Trop. Med. Hyg. 2000, 62, 480–484. [Google Scholar] [CrossRef] [Green Version]
- Higgs, S.; Vanlandingham, D.L.; Klingler, K.A.; McElroy, K.L.; McGee, C.E.; Harrington, L.; Lang, J.; Monath, T.P.; Guirakhoo, F. Growth Characteristics of Chimerivax-Den Vaccine Viruses in Aedes Aegypti and Aedes Albopictus from Thailand. Am. J. Trop. Med. Hyg. 2006, 75, 986–993. [Google Scholar] [CrossRef] [PubMed]
- Whitman, L. Failure of Aedes Aegypti to Trasmit Yellow Fever Cultured Virus (17d). Am. J. Trop. Med. Hyg. 1939, 1, 19–26. [Google Scholar] [CrossRef]
- Whitman, L.; Antunes, P. Studies on Aedes Aegypti Infected in the Larval Stage with the Virus of Yellow Fever. Proc. Soc. Exp. Biol. Med. 1938, 37, 664–666. [Google Scholar] [CrossRef]
- Sun, P.; Nie, K.; Zhu, Y.; Liu, Y.; Wu, P.; Liu, Z.; Du, S.; Fan, H.; Chen, C.H.; Zhang, R.; et al. A Mosquito Salivary Protein Promotes Flavivirus Transmission by Activation of Autophagy. Nat. Commun. 2020, 11, 260. [Google Scholar] [CrossRef] [Green Version]
- Arlian, L.G. Arthropod Allergens and Human Health. Annu. Rev. Entomol. 2002, 47, 395–433. [Google Scholar] [CrossRef] [PubMed]
- Galindo, P.; Gómez, E.; Feo, F.; Borja, J.; Garcia, R.; Lombardero, M.; Barber, D. Anaphylaxis from Mosquito Bite. Allergy 1998, 26, 251–254. [Google Scholar]
- McCormack, D.R.; Salata, K.F.; Hershey, J.N.; Carpenter, G.B.; Engler, R.J. Mosquito Bite Anaphylaxis: Immunotherapy with Whole Body Extracts. Ann. Allergy Asthma Immunol. 1995, 74, 39–44. [Google Scholar] [PubMed]
- Adam, A.; Luo, H.; Osman, S.R.; Wang, B.; Roundy, C.M.; Auguste, A.J.; Plante, K.S.; Peng, B.-H.; Thangamani, S.; Frolova, E.I.; et al. Optimized Production and Immunogenicity of an Insect Virus-Based Chikungunya Virus Candidate Vaccine in Cell Culture and Animal Models. Emerg. Microbes Infect. 2021, 10, 305–316. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Disease | Vaccine | Type of Vaccine | Where Licensed |
|---|---|---|---|
| yellow fever [17] | 17D-204 and 17DD | Live attenuated | globally |
| tick-borne encephalitis [18] | FSME-IMMUNE, Encepur, TBE-MOSCOW, EnceVir, CIBP | Formalin inactivated | European countries |
| Japanese encephalitis [19,20,21,22] | SA14-14-2 | Live-attenuated | China and some Asian countries |
| IXIARO/IC51 | Formalin inactivated | globally | |
| JEBIK V | Inactivated | Japan | |
| ENCEVAC | Inactivated | Japan | |
| JEEV | Inactivated | India | |
| Kyasanur forest disease [23] | - | Formalin inactivated | India and nearby regions |
| Disease | Parental Viruses | Vaccine | Comments |
|---|---|---|---|
| JEV | YFV 17DD genomic backbone; prM-E of JEV (SA14-14-2) | ChimeriVax-JE (IMOJEV, THAIJEV) | Showed robust protection against virulent JEV challenge in mice and Rhesus macaques [24,30,31,32,33]. Approved for use in humans in Australia, New Zealand, Thailand, Korea, Malaysia, Philippines and Vietnam |
| WNV | YFV 17DD genomic backbone; prM-E of WNV (NY99) | ChimeriVax-WN | Highly immunogenic and protective in preclinical animal studies [25,27], progressed to Phase II human clinical trial |
| YFV 17DD genomic backbone; prM-E of WNV (NY99) | ChimeriVax-WN02 (PreveNile) | Induced high levels of neutralising antibodies after a single dose during clinical trials and was licensed for veterinary use in 2006 but was recalled in 2010 | |
| Dengue 4 (Caribbean 814669) genomic backbone; prM-E of WNV (NY99) | WN-DEN4 | Shown to be attenuated in monkeys, induced moderate-to-high neutralising antibody titres and prevented viraemia after WNV challenge [34], progressed to Phase II human clinical trial | |
| DENV | YFV 17DD genomic backbone; prM-E of each DENV serotype (tetravalent) | Dengvaxia, CYD-TDV | Induced long-lasting, cross-neutralising antibody response to all 4 DENV serotypes [30]. Licensed for use over the age of 9 in DENV-endemic areas. |
| DENV2 genomic backbone; prM-E of each DENV serotype (tetravalent) | TAK-003 (Takeda) | Shown to be well tolerated and immunogenic against all four serotypes in Phase I and Phase II clinical trials [35]. Ongoing Phase III clinical trials [35,36]. |
| Virus | Lineage | Geographic Distribution | Host Range |
|---|---|---|---|
| Cell fusing agent virus—CFAV [82,83,84] | I | USA 2, Puerto Rico, Australia 2, Indonesia, Mexico, Thailand | Aedes aegypti, Aedes albopictus, Culex species |
| Kamiti River virus—KRV [74] | I | Kenya | Aedes macintoshi |
| Aedes flavivirus—AeFV [82,85,86,87,88,89] | I | Japan, Italy, USA, Thailand 2 | Aedes albopictus, Aedes flavopictus |
| Culex flavivirus—CxFV [88,90,91,92,93,94,95,96,97] | I | Japan, Indonesia, China, Guatemala, USA, Mexico, Trinidad, Uganda, Argentina, Australia | Culex species |
| Nienokoune virus—NIEV [98] | I | Cotê d’Ivoire | Culex species |
| Barkedji virus—BJV [99] | II | Senegal, Israel | Culex perexiguus |
| Lammi virus—LAMV [100,101] | II | Finland | Aedes cinereus |
| Marisma mosquito virus—MMV [102,103,104] | II | Spain, Italy | Aedes caspius |
| Ilomantsi virus—ILOV [100] | II | Finland | Ochlerotatus riparious, Anopheles species |
| Long Pine Key virus—LPKV [105] | II | USA | Anopheles crucians, Aedes atlanticus, Culex nigripalpus |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harrison, J.J.; Hobson-Peters, J.; Bielefeldt-Ohmann, H.; Hall, R.A. Chimeric Vaccines Based on Novel Insect-Specific Flaviviruses. Vaccines 2021, 9, 1230. https://doi.org/10.3390/vaccines9111230
Harrison JJ, Hobson-Peters J, Bielefeldt-Ohmann H, Hall RA. Chimeric Vaccines Based on Novel Insect-Specific Flaviviruses. Vaccines. 2021; 9(11):1230. https://doi.org/10.3390/vaccines9111230
Chicago/Turabian StyleHarrison, Jessica J., Jody Hobson-Peters, Helle Bielefeldt-Ohmann, and Roy A. Hall. 2021. "Chimeric Vaccines Based on Novel Insect-Specific Flaviviruses" Vaccines 9, no. 11: 1230. https://doi.org/10.3390/vaccines9111230
APA StyleHarrison, J. J., Hobson-Peters, J., Bielefeldt-Ohmann, H., & Hall, R. A. (2021). Chimeric Vaccines Based on Novel Insect-Specific Flaviviruses. Vaccines, 9(11), 1230. https://doi.org/10.3390/vaccines9111230