Novel Inactivated Subtype B Avian Metapneumovirus Vaccine Induced Humoral and Cellular Immune Responses

 , ,
, , 
 add
Show full author list
add
Show full author list
Abstract
1. Introduction
2. Methods
2.1. Experimental Animals
2.2. Ethical and Regulatory Approval
2.3. Cell and Virus
2.4. Vaccine Preparation
2.5. Experimental Design
2.6. ELISA
2.7. Virus Neutralization Test
2.8. Virus and Cytokine Detection Using RT-qPCR
2.9. Lymphocyte Proliferation Assay
2.10. Clinical Signs
2.11. Histopathological Analysis
2.12. Immunity Duration of the Inactivated Vaccine
2.13. Statistical Analysis
3. Results
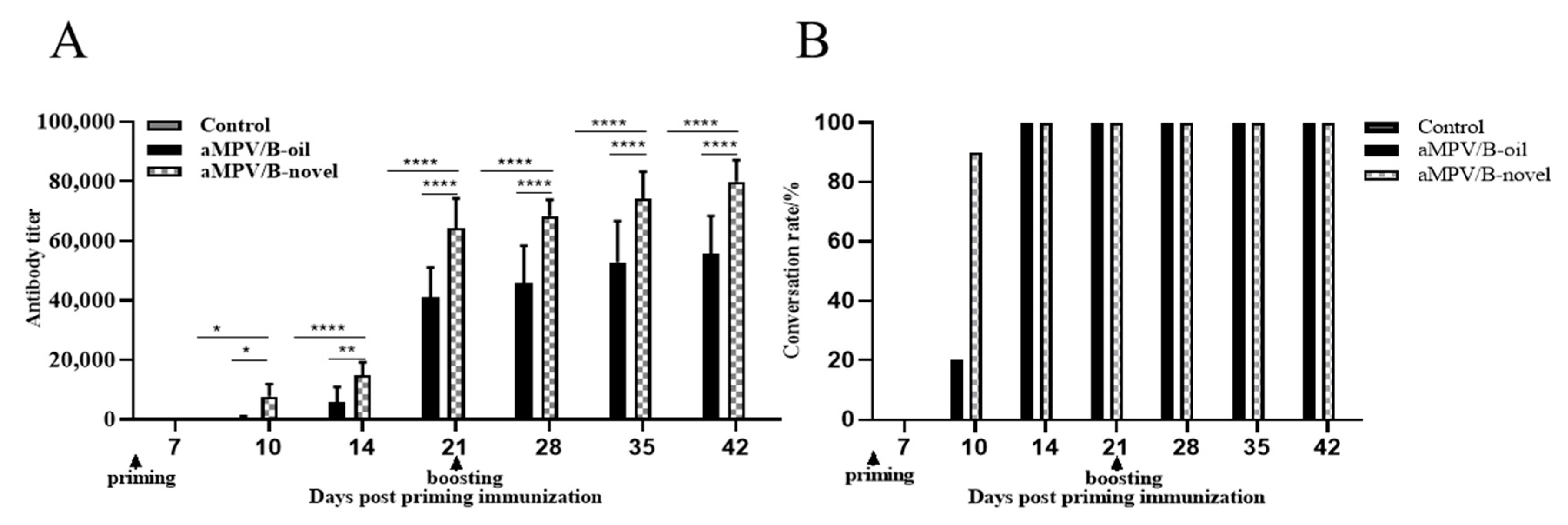
3.1. Serum IgG Antibody Levels
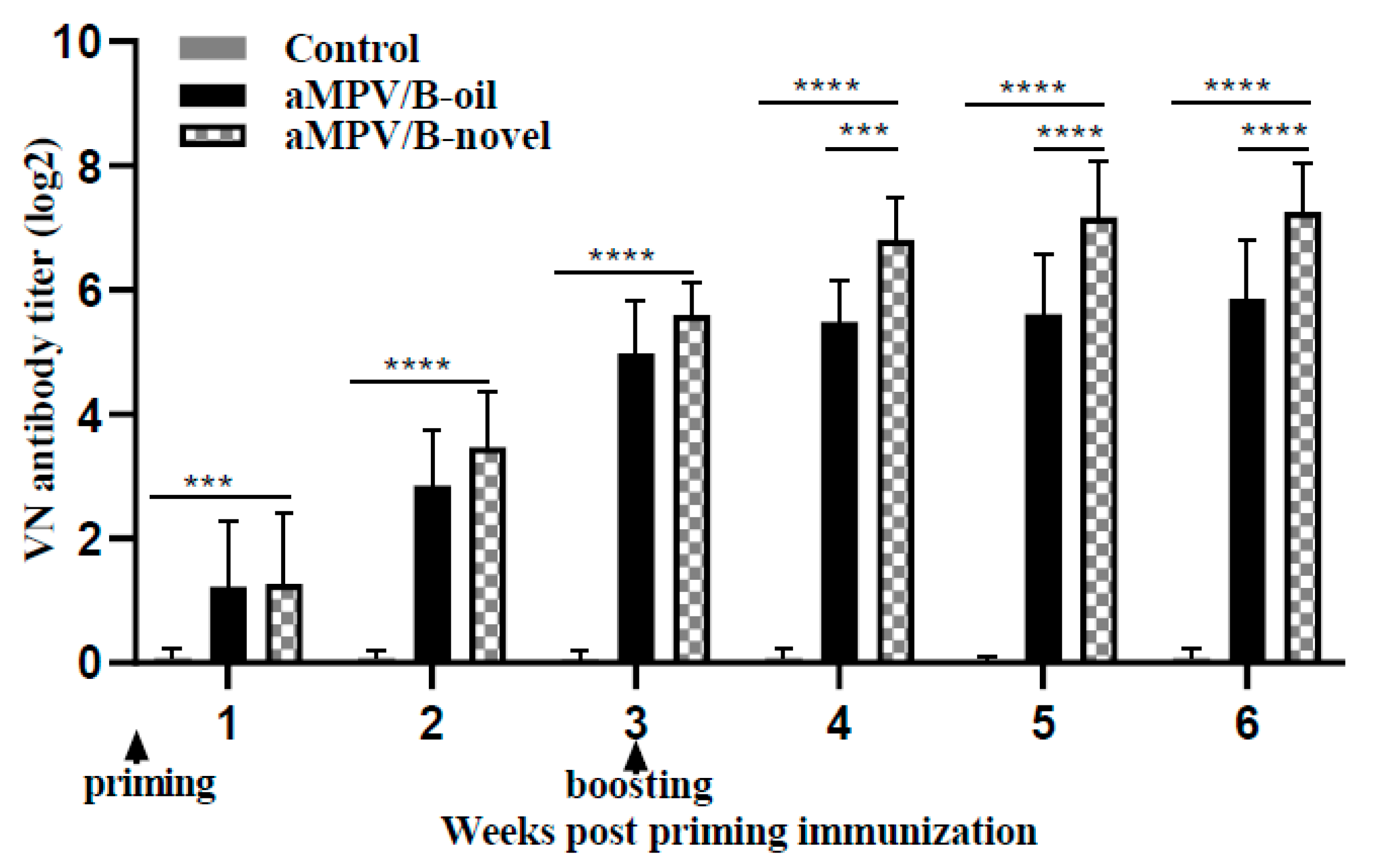
3.2. Virus Neutralizing Antibody Level
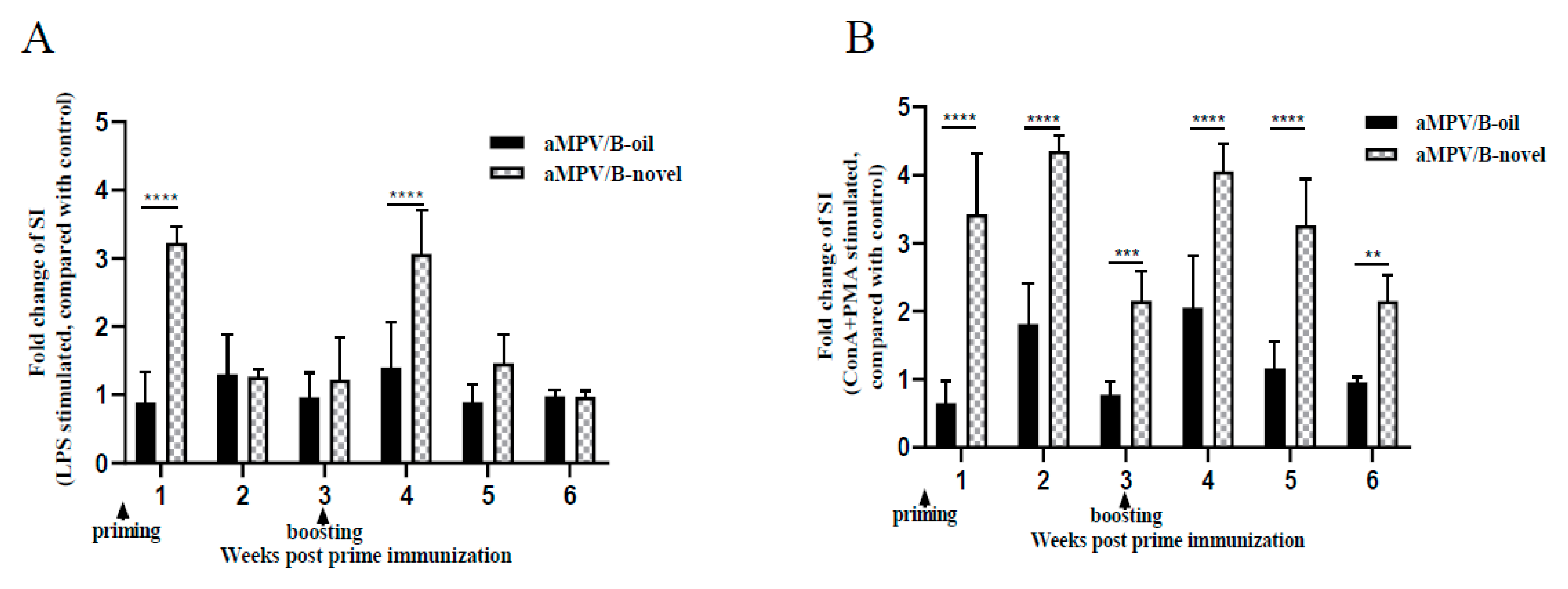
3.3. Lymphocyte Proliferation Activity in PBMCs
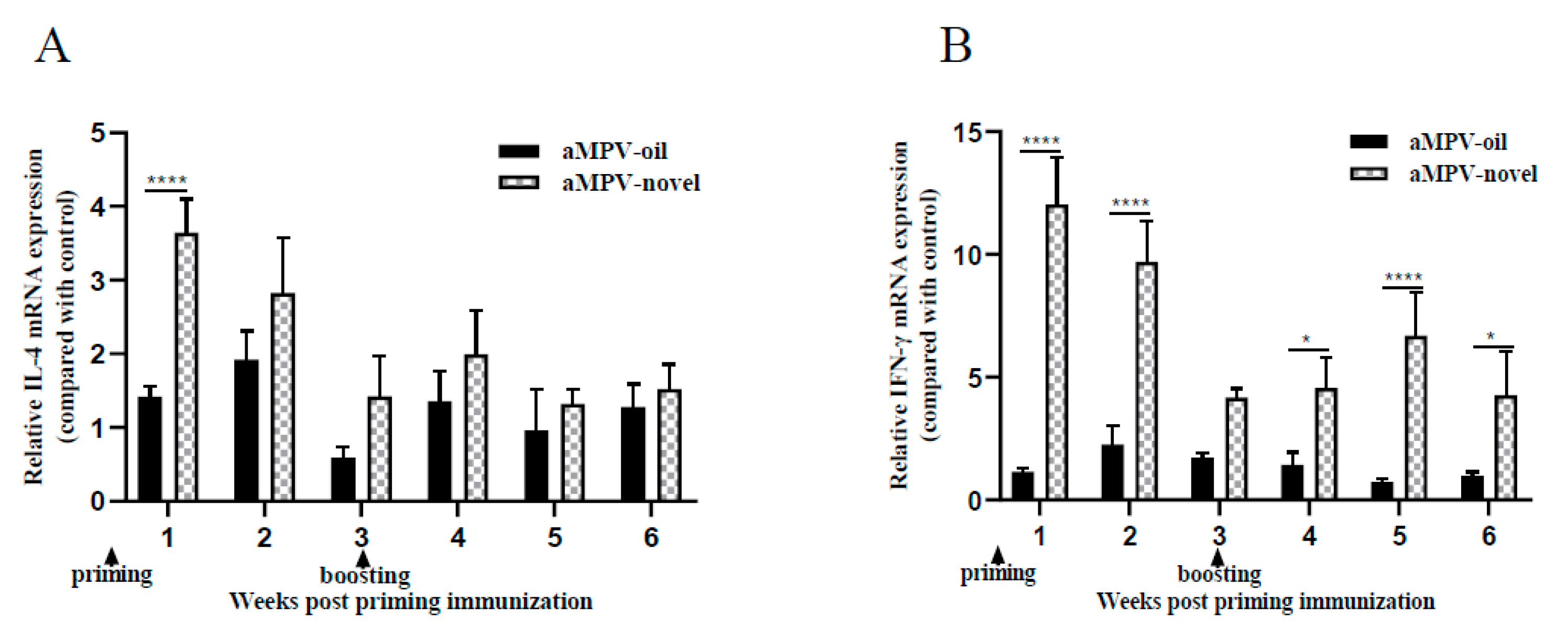
3.4. Th1- and Th2-Associated Cytokine Detection in Spleen
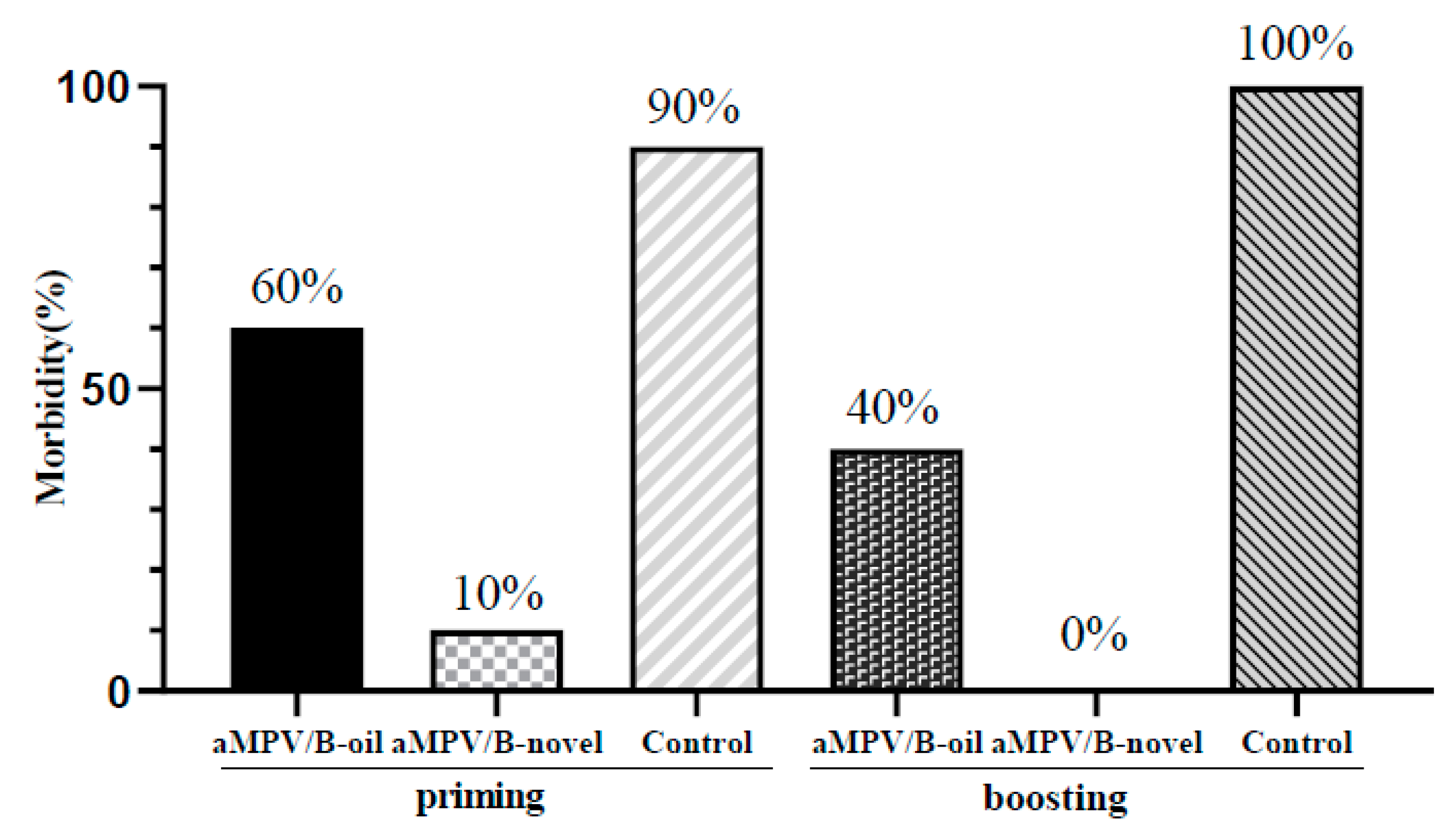
3.5. Clinical Symptoms
3.6. Virus Shedding
3.7. Histopathological Analysis
3.8. Duration of the Inactivated Vaccine
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Buys, S.B.; du Preez, J.H.; Els, H.J. Swollen head syndrome in chickens: A preliminary report on the isolation of a possible aetiological agent. J. S. Afr. Vet. Assoc. 1989, 60, 221–222. [Google Scholar] [PubMed]
- Zhu, Y.; Gong, X.; Guo, W.; Xu, B.; Li, L.; Lang, F.; Liu, H.; Liu, D.; Fan, H. Molecular epidemiological analysis of avian Metapneumovirus in some areas of China from 2012 to 2015. Prog. Vet. Med. 2016, 37, 30–34. (In Chinese) [Google Scholar]
- Giovanni, F.; Matteo, L.; Giulia, M.; Claudia, M.T.; Caterina, L.; Giulia, Q.; Elen, C.; Mattia, C. Avian Metapneumovirus subtype B around Europe: A phylodynamic reconstruction. Vet. Res. 2020, 51, 88. [Google Scholar]
- Seal, B.S.; Sellers, H.S.; Meinersmann, R.J. Fusion protein predicted amino acid sequence of the first US avian Pneumovirus isolate and lack of heterogeneity among other US isolates. Virus Res. 2000, 66, 139–147. [Google Scholar] [CrossRef]
- Cook, J.K.A.; Kinloch, S.; Ellis, M.M. In vitro and in vivo studies in chickens and turkeys on strains of turkey rhinotracheitis virus isolated from the two species. Avian Pathol. 1993, 22, 157–170. [Google Scholar] [CrossRef] [PubMed]
- Van de Zande, S.; Nauwynck, H.; De Jonghe, S.; Pensaert, M. Comparative pathogenesis of a subtype A with a subtype B avian Pneumovirus in turkeys. Avian Pathol. 1999, 28, 239–244. [Google Scholar] [CrossRef][Green Version]
- Jones, R.C. Avian Pneumovirus infection: Questions still unanswered. Avian Pathol. 1996, 25, 639–648. [Google Scholar] [CrossRef]
- Timms, L.M.; Jahans, K.L.; Marshall, R.N. Evidence of immunosuppression in turkey poults affected by rhinotracheitis. Vet. Rec. 1986, 119, 91–92. [Google Scholar] [CrossRef]
- Jones, R.C.; Naylor, C.J.; al-Afaleq, A.; Worthington, K.J.; Jones, R. Effect of cyclophosphamide immunosuppression on the immunity of turkeys to viral rhinotracheitis. Res. Vet. Sci. 1992, 53, 38–41. [Google Scholar] [CrossRef]
- Chary, P.; Rautenschlein, S.; Sharma, J.M. Reduced efficacy of hemorrhagic enteritis virus vaccine in turkeys exposed to avian Pneumovirus. Avian Dis. 2002, 46, 353–359. [Google Scholar] [CrossRef]
- Van de Zande, S.V.D.; Nauwynck, H.; Pensaert, M. The clinical, pathological and microbiological outcome of an Escherichia coli O2:K1 infection in avian Pneumovirus infected turkeys. Vet. Microbiol. 2001, 81, 353–365. [Google Scholar] [CrossRef]
- Abdel-Azeem, A.A.; Franzo, G.; Dalle Zotte, A.; Drigo, M.; Catelli, E.; Lupini, C. First evidence of avian Metapneumovirus subtype a infection in turkeys in Egypt. Trop. Anim. Health Prod. 2014, 46, 1093–1097. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Jiang, F.; Sun, F.; Wang, K.; Wang, S. Research progress on avian Metapneumovirus. Chin. Anim. Health Inspect. 2020, 37, 86–91. (In Chinese) [Google Scholar]
- Yu, M.; Xing, L.; Chang, F.; Bao, Y.; Wang, S.; He, X.; Wang, J.; Wang, S.; Liu, Y.; Muhammad, F.; et al. Genomic sequence and pathogenicity of the first avian Metapneumovirus subtype B isolated from chicken in China. Vet. Microbiol. 2019, 228, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.-K.; Chen, F.; Cao, S.; Liu, J.; Lei, W.; Li, G.; Song, Y.; Lu, J.; Liu, C.; Qin, J.; et al. Isolation and characterization of a subtype C avian Metapneumovirus circulating in Muscovy ducks in China. Vet. Res. 2014, 45, 74. [Google Scholar] [CrossRef]
- Turpin, E.A.; Stallknecht, D.E.; Slemons, R.D.; Zsak, L.; Swayne, D.E. Evidence of avian Metapneumovirus subtype C infection of wild birds in Georgia, South Carolina, Arkansas and Ohio, USA. Avian Pathol. 2008, 37, 343–351. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A simple method of estimating 50% endpoints. Am. J. Med. 1938, 27, 493–497. [Google Scholar]
- Bachmann, M.F.; Kündig, T.M.; Kalberer, C.P.; Hengartner, H.; Zinkernagel, R.M. Formalin inactivation of vesicular stomatitis virus impairs T-cell- but not T-help-independent B-cell responses. J. Virol. 1993, 67, 3917–3922. [Google Scholar] [CrossRef]
- Minna, M.H.; Virginia, M.S.; Amir-Babak, S.K.; Suvi, H.; Vesa, P.H. A comparative study of the effect of UV and formalin inactivation on the stability and immunogenicity of a Coxsackievirus B1 vaccine. Vaccine 2019, 37, 40. [Google Scholar]
- Bennett, R.S.; LaRue, R.; Shaw, D.; Yu, Q.; Nagaraja, K.V.; Halvorson, D.A.; Njenga, M.K. A wild goose Metapneumovirus containing a large attachment glycoprotein is avirulent but immunoprotective in domestic turkeys. J. Virol. 2005, 79, 14834–14842. [Google Scholar] [CrossRef]
- Hu, H.; Roth, J.P.; Zsak, L.; Yu, Q. Engineered Newcastle disease virus expressing the F and G proteins of AMPV-C confers protection against challenges in turkeys. Sci. Rep. 2017, 7, 4025. [Google Scholar] [CrossRef] [PubMed]
- Morel, S.; Didierlaurent, A.; Bourguignon, P.; Delhaye, S.; Baras, B.; Jacob, V.; Planty, C.; Elouahabi, A.; Harvengt, P.; Carlsen, H.; et al. Adjuvant System AS03 containing α-tocopherol modulates innate immune response and leads to improved adaptive immunity. Vaccine 2011, 29, 2461–2473. [Google Scholar] [CrossRef] [PubMed]
- Calabro, S.; Tortoli, M.; Baudner, B.C.; Pacitto, A.; Cortese, M.; O’Hagan, D.T.; De Gregorio, E.; Seubert, A.; Wack, A. Vaccine adjuvants alum and MF59 induce rapid recruitment of neutrophils and monocytes that participate in antigen transport to draining lymph nodes. Vaccine 2011, 29, 1812–1823. [Google Scholar] [CrossRef] [PubMed]
- Del Giudice, G.; Rappuoli, R.; Didierlaurent, A.M. Correlates of adjuvanticity: A review on adjuvants in licensed vaccines. Semin. Immunol. 2018, 39, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Jackson, S.T.; Zachary, L.B.; Irina, L.G. Signals 1, 2 and B cell fate or: Where, when and for how long? Immunol. Rev. 2020, 296, 9–23. [Google Scholar]
- Rubbenstroth, D.; Rautenschlein, S. Investigations on the protective role of passively transferred antibodies against avian Metapneumovirus infection in turkeys. Avian Pathol. 2009, 38, 427–436. [Google Scholar] [CrossRef]
- Kapczynski, D.R.; Perkins, L.L.; Sellers, H.S. Mucosal vaccination with formalin-inactivated avian Metapneumovirus subtype C does not protect turkeys following intranasal challenge. Avian Dis. 2008, 52, 28–33. [Google Scholar] [CrossRef]
- Robinson, S.R.; Li, J.; Nelson, E.A.; Murtaugh, M.P. Broadly neutralizing antibodies against the rapidly evolving porcine reproductive and respiratory syndrome virus. Virus Res. 2015, 203, 56–65. [Google Scholar] [CrossRef]
- Sedeik, M.E.; Elbestawy, A.R.; El-shall, N.A.; Abd El-Hack, M.E.; Saadeldin, I.M.; Swelum, A.A. Comparative efficacy of commercial inactivated Newcastle disease virus vaccines against Newcastle disease virus genotype VII in broiler chickens. Poult. Sci. 2019, 98, 2000–2007. [Google Scholar] [CrossRef]
- Kapczynski, D.R.; Afonso, C.L.; Miller, P.J. Immune responses of poultry to Newcastle disease virus. Dev. Comp. Immunol. 2013, 41, 447–453. [Google Scholar] [CrossRef]
- Gunjan, K.; Mohsin, R.; Brijendra, K.T. Interferon-gamma (IFN-γ): Exploring its implications in infectious diseases. Biomol. Concepts 2018, 9, 64–79. [Google Scholar]
- Chen, X.; Liu, H.; Peng, Y.; He, L.; Liu, F. Expression and correlation analysis of IL-4, IFN-γ and FcαRI in tonsillar mononuclear cells in patients with IgA nephropathy. Cell Immunol. 2014, 289, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wang, S.; Xing, H.; Sun, Y.; Ding, J.; He, N. Bulleyaconitine A inhibits the lung inflammation and airway remodeling through restoring Th1/Th2 balance in asthmatic model mice. Biosci. Biotechnol. Biochem. 2020, 84, 1409–1417. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Okada, H.; Itoh, T.; Tada, T.; Mase, M.; Nakamura, K.; Kubo, M.; Tsukamoto, K. Association of increased pathogenicity of Asian H5N1 highly pathogenic avian influenza viruses in chickens with highly efficient viral replication accompanied by early destruction of innate immune responses. J. Virol. 2009, 83, 7475–7486. [Google Scholar] [CrossRef] [PubMed]
- Susta, L.; Cornax, I.; Diel, D.G.; Garcia, S.C.; Miller, P.J.; Liu, X.; Hu, S.; Brown, C.C.; Afonso, C.L. Expression of interferon gamma by a highly virulent strain of Newcastle disease virus decreases its pathogenicity in chickens. Microb. Pathog. 2013, 61–62, 73–83. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
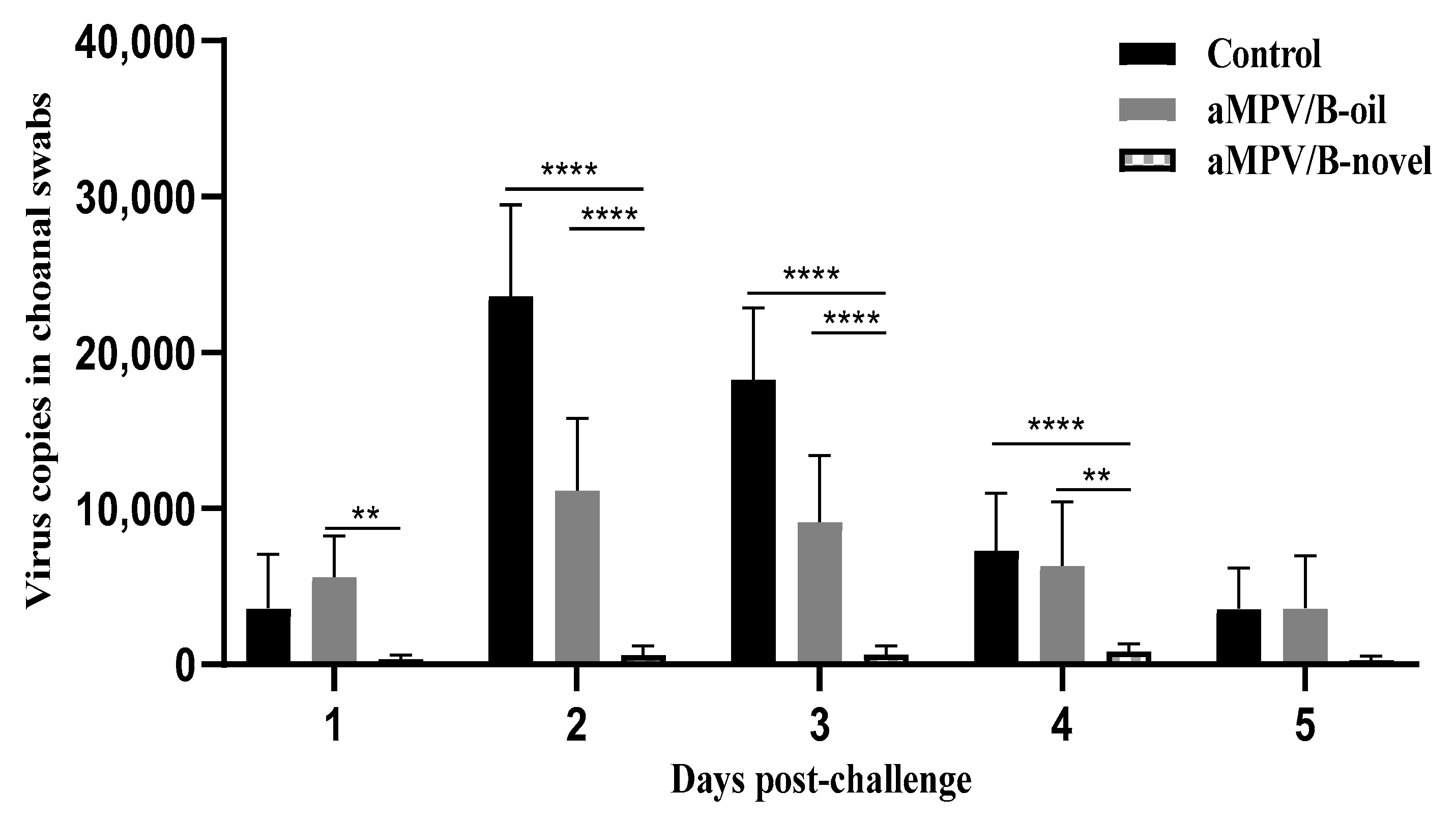{kind=link}
{kind=link}
| RNA Target | Probe/Primer Sequence (5′-3′) |
|---|---|
| aMPV/B | Probe CTGGTGTTATCAGCCTTAGGCTTGACGCT |
| F AATAGTCCTCAAGCAAGTCCTCAGA | |
| R CTGTTGTAATTTGACCTGTTCTA CACT | |
| 28s | Probe GCATGGCTTAATCTTTGAGACAA |
| F ATCCTGCCAGTAGCATATG | |
| R GCCGTGCGTACTTACACGT | |
| IL-4 | Probe CTCTTCCTCAACATGCGTCAGCTCCTGAATGCCAG |
| F TGCCACAAGAACCTGCAGGG | |
| R AAGTAGTGTTGCCTGCTGCC | |
| IFN-γ | Probe TGCCCTGCGTCAGGACTCTAA |
| F ATCTCGAAAAACAACCTTCCTGATGGC | |
| R TTCCAGCCCAAGTGGAGCCGG |
| Group | Geometric Mean VN Titers (log2) ± Standard Error at Weeks Post Immunization | |||||
|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | |
| aMPV/B-oil a | 1.27 ± 0.91 | 2.72 ± 0.85 | 4.91 ± 0.82 | 5.45 ± 0.62 | 5.54 ± 0.90 | 5.79 ± 0.89 |
| aMPV/B-novel b | 1.30 ± 0.98 | 3.35 ± 0.84 | 5.57 ± 0.50 | 6.78 ± 0.64 | 7.12 ± 0.87 | 7.21 ± 0.76 |
| Time after Boosting Vaccination/Months | |||||||
|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | ||
| VN titer | Vaccinated a | 7.09 ± 0.92 c | 6.59 ± 0.51 | 6.17 ± 1.17 | 5.57 ± 0.79 | 5.04 ± 1.39 | 5.28 ± 1.34 |
| Control b | <2 | <2 | <2 | <2 | <2 | <2 | |
| Morbidity | Vaccinated | 0% | 0% | 10% | 0% | 10% | 10% |
| Control | 90% | 100% | 100% | 90% | 100% | 90% | |
| Protective rate | 100% | 100% | 90% | 100% | 90% | 89% | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bao, Y.; Yu, M.; Liu, P.; Hou, F.; Muhammad, F.; Wang, Z.; Li, X.; Zhang, Z.; Wang, S.; Chen, Y.; et al. Novel Inactivated Subtype B Avian Metapneumovirus Vaccine Induced Humoral and Cellular Immune Responses. Vaccines 2020, 8, 762. https://doi.org/10.3390/vaccines8040762
Bao Y, Yu M, Liu P, Hou F, Muhammad F, Wang Z, Li X, Zhang Z, Wang S, Chen Y, et al. Novel Inactivated Subtype B Avian Metapneumovirus Vaccine Induced Humoral and Cellular Immune Responses. Vaccines. 2020; 8(4):762. https://doi.org/10.3390/vaccines8040762
Chicago/Turabian StyleBao, Yuanling, Mengmeng Yu, Peng Liu, Fujun Hou, Farooque Muhammad, Zhihao Wang, Xinyi Li, Zhuo Zhang, Suyan Wang, Yuntong Chen, and et al. 2020. "Novel Inactivated Subtype B Avian Metapneumovirus Vaccine Induced Humoral and Cellular Immune Responses" Vaccines 8, no. 4: 762. https://doi.org/10.3390/vaccines8040762
APA StyleBao, Y., Yu, M., Liu, P., Hou, F., Muhammad, F., Wang, Z., Li, X., Zhang, Z., Wang, S., Chen, Y., Cui, H., Liu, A., Qi, X., Pan, Q., Zhang, Y., Gao, L., Li, K., Liu, C., He, X., ... Gao, Y. (2020). Novel Inactivated Subtype B Avian Metapneumovirus Vaccine Induced Humoral and Cellular Immune Responses. Vaccines, 8(4), 762. https://doi.org/10.3390/vaccines8040762