Experimental PCEP-Adjuvanted Swine Influenza H1N1 Vaccine Induced Strong Immune Responses but Did Not Protect Piglets against Heterologous H3N2 Virus Challenge




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Swine Influenza Virus Adsorption and Purification
2.2. Formalin Inactivation of Influenza Virus
2.3. Adjuvant and Vaccine Preparation
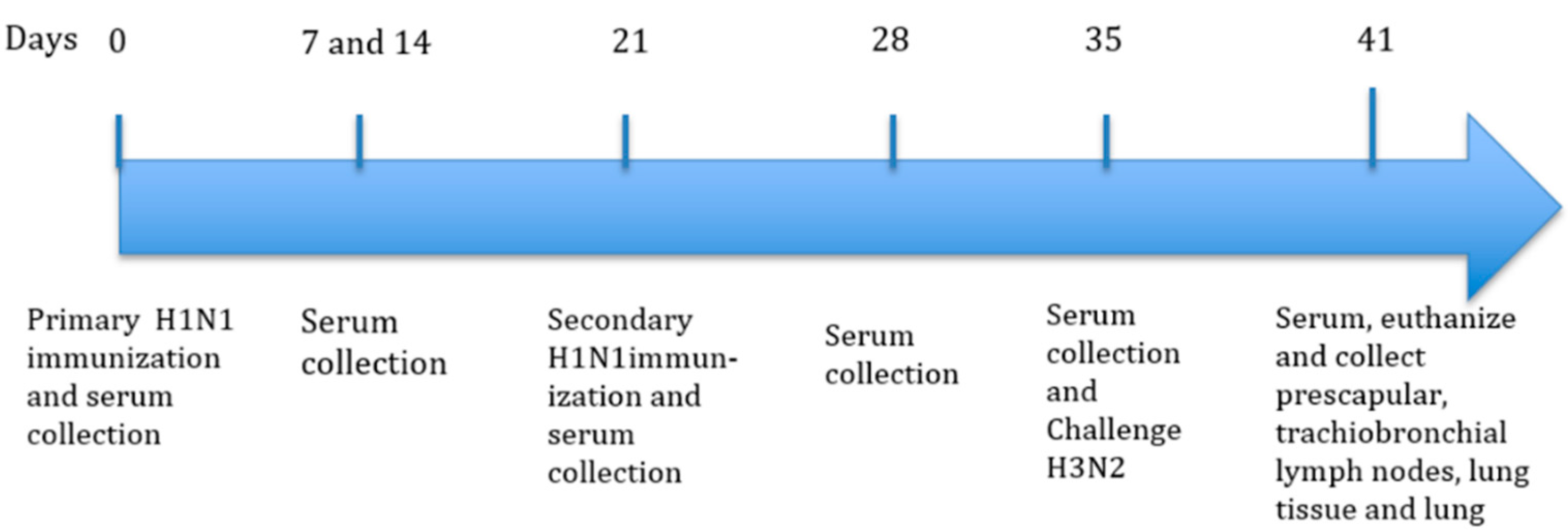
2.4. Immunization with Inactivated H1N1 and Challenge with H3N2 Virus Experimental Design
2.5. Necropsy and Macroscopic Examination of the Lungs
2.6. Detection of Swine Influenza Virus H1N1 and H3N2 Antibodies in Porcine Serum by Enzyme-Linked Immunosorbent Assay (ELISA)
2.7. Enzyme-Linked Immunosorbent SPOT Assay (ELISPOT) for IFN-γ, IL-13 and IL-17A
2.8. Swine Influenza Virus Neutralization Assay
2.9. Statistical Analysis
3. Results
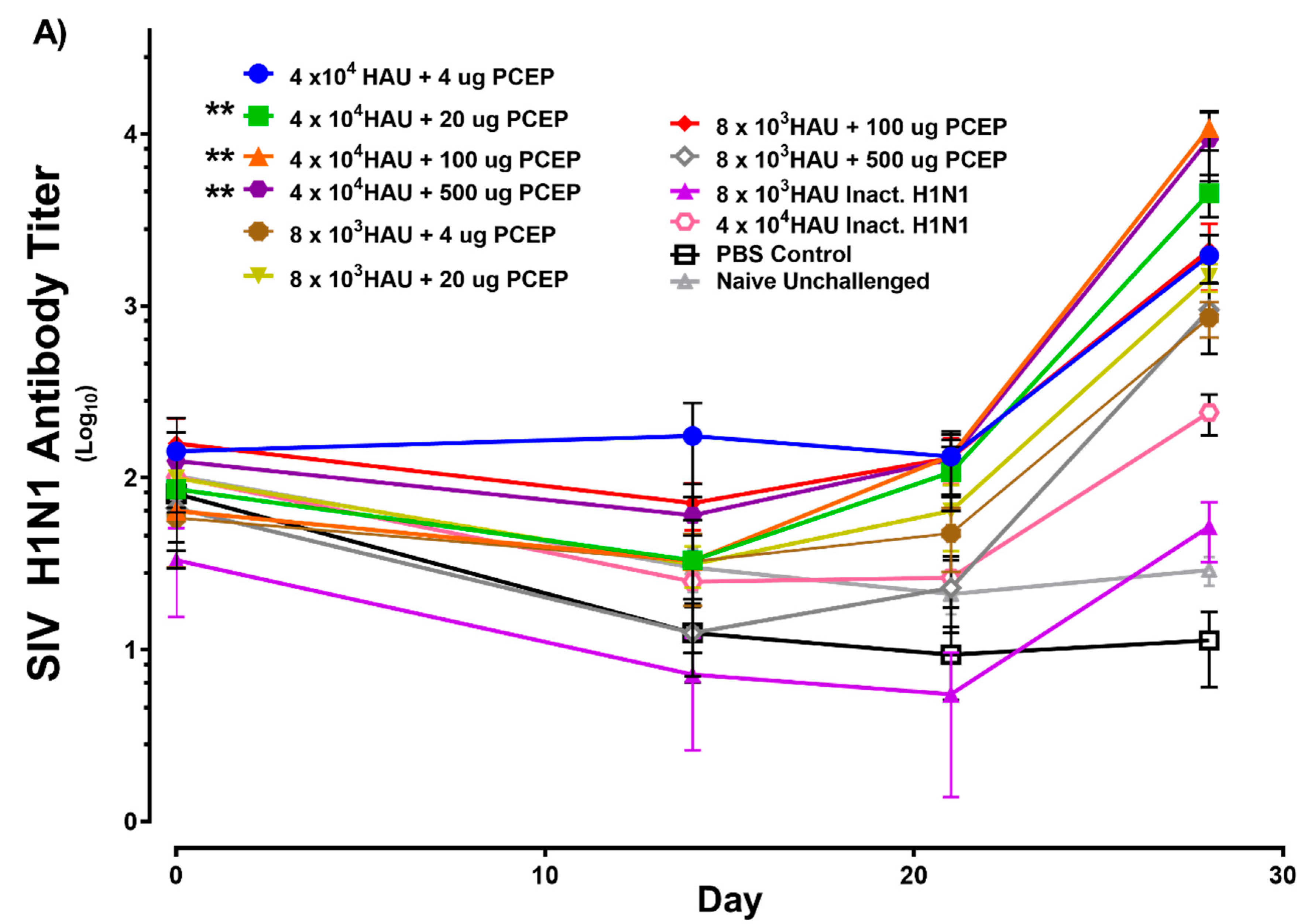
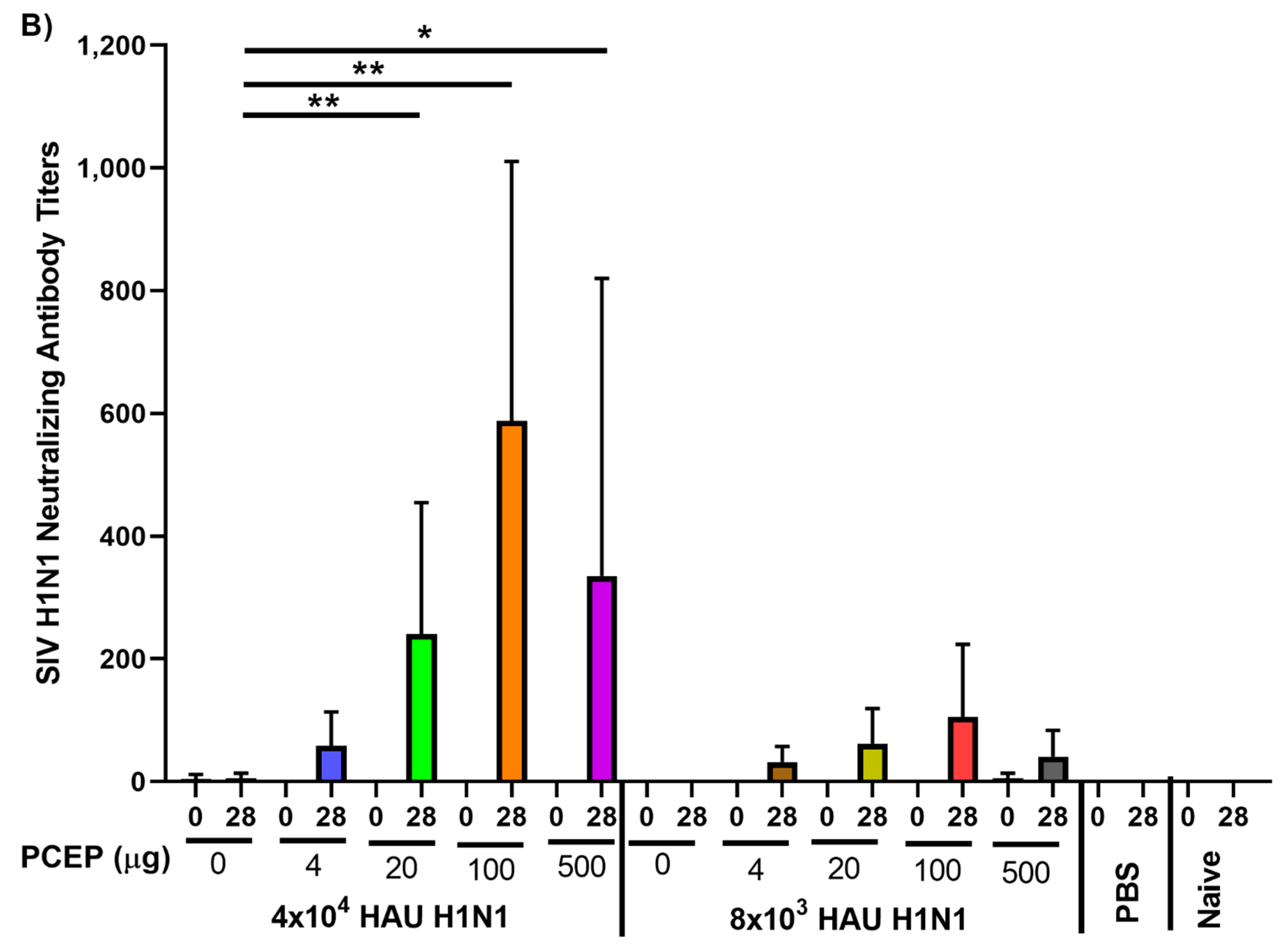
3.1. Immune Responses after Intradermal Vaccination of Pigs with PCEP-Adjuvanted Inactivated SIV H1N1 Swine Influenza Virus Vaccine and Viral Protection Study
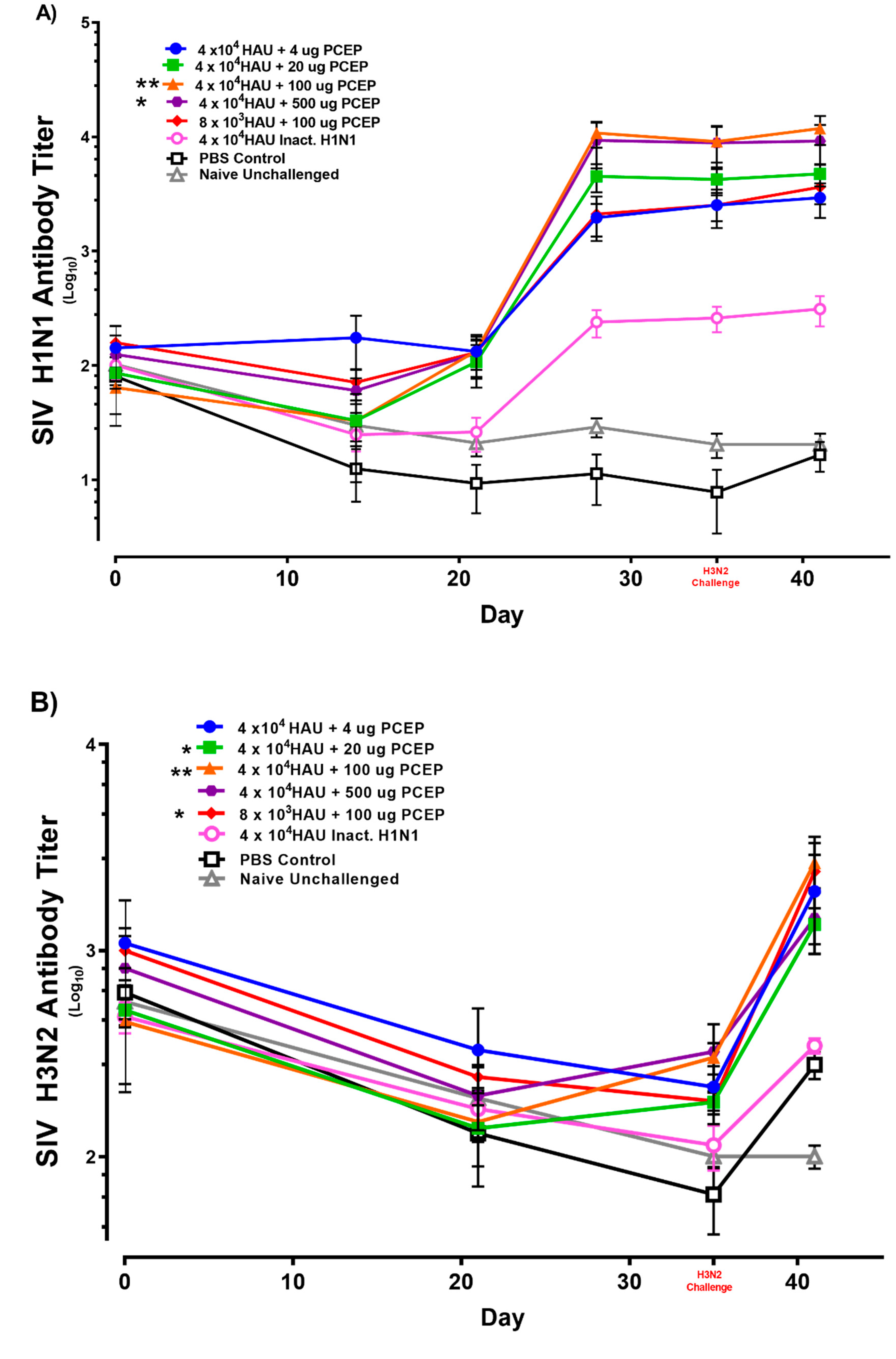
3.2. Immune Responses after Vaccination with Inactivated SIV H1N1 Swine Influenza Virus with PCEP Adjuvant after Challenge with Live H3N2
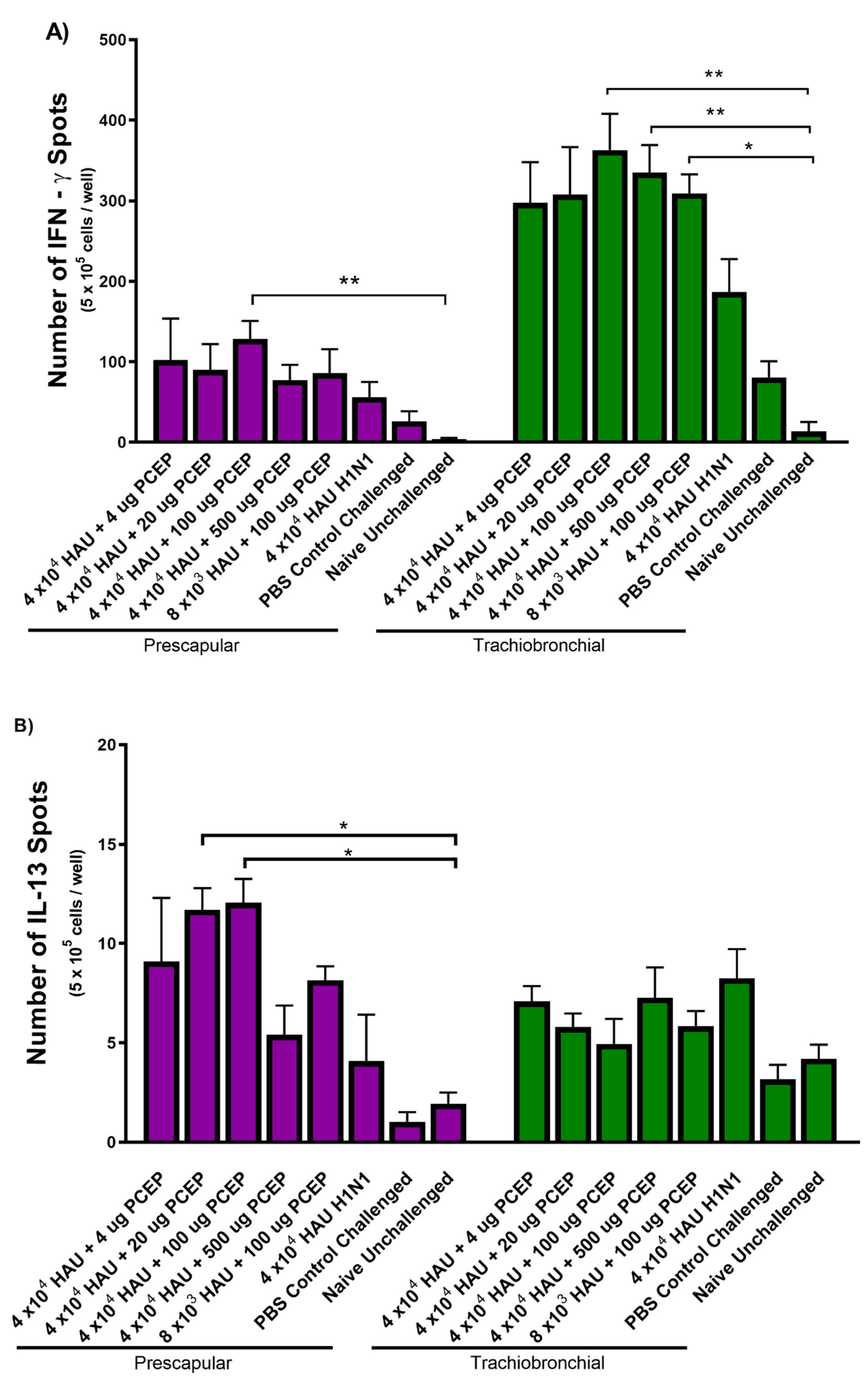
3.3. Cytokine Production from Lymph Nodes Collected from the Injection and Vaccination Site at Time of Termination
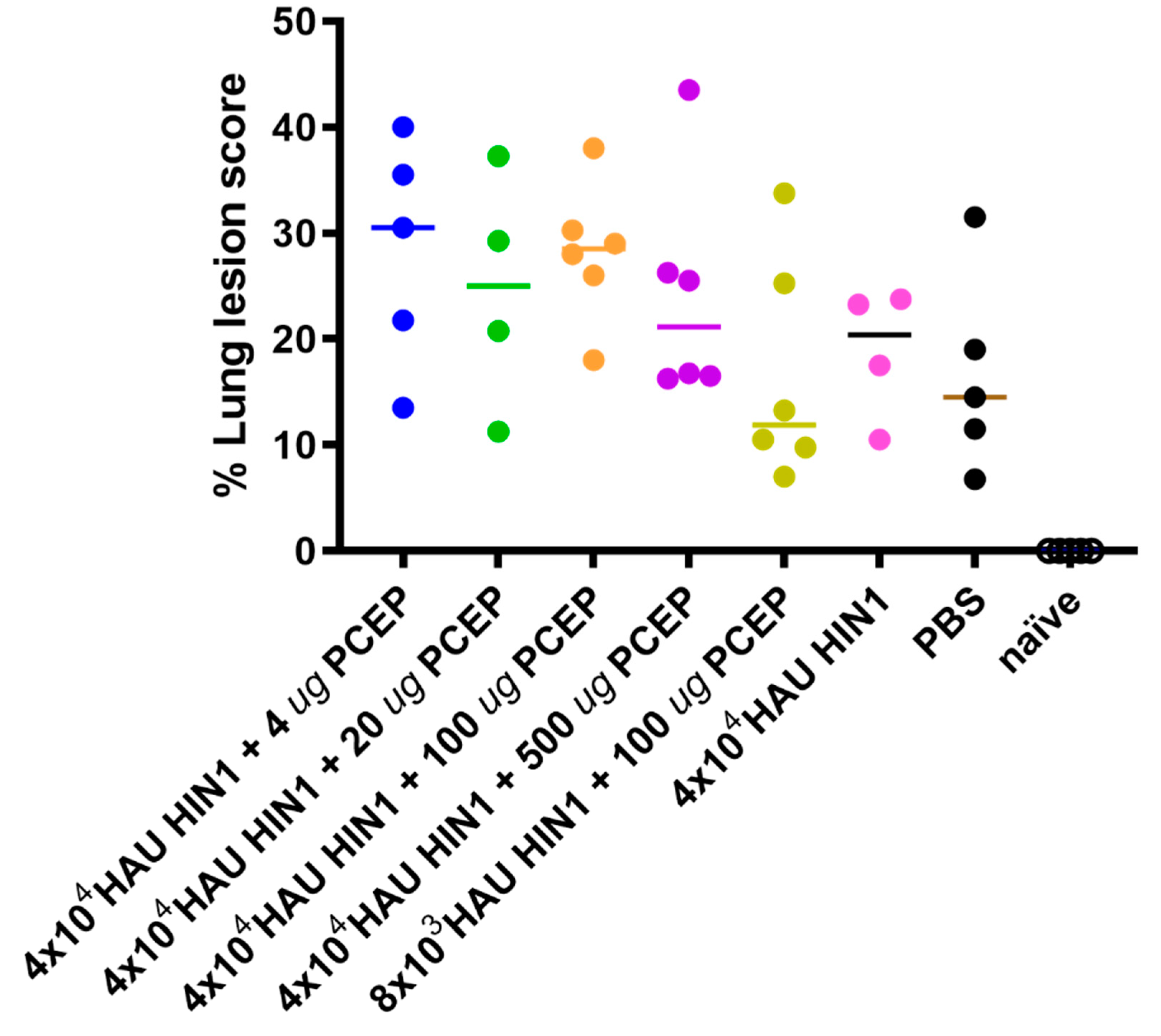
3.4. Lung Lesion Score
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dandagi, G.L.; Byahatti, S.M. An insight into the swine-influenza A (H1N1) virus infection in humans. Lung India Off. Organ Indian Chest Soc. 2011, 28, 34. [Google Scholar] [CrossRef] [PubMed]
- Vincent, A.L.; Ma, W.; Lager, K.M.; Janke, B.H.; Richt, J.A. Swine influenza viruses: A North American perspective. Adv. Virus Res. 2008, 72, 127–154. [Google Scholar] [PubMed]
- Anderson, T.K.; Nelson, M.I.; Kitikoon, P.; Swenson, S.L.; Korslund, J.A.; Vincent, A.L. Population dynamics of cocirculating swine influenza A viruses in the United States from 2009 to 2012. Influenza Other Resp. Virus. 2013, 7, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Brown, I.H. History and Epidemiology of Swine Influenza in Europe. In Swine Influenza. Current Topics in Microbiology and Immunology; Richt, J., Webby, R., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; Volume 370. [Google Scholar]
- Smith, G.J.; Vijaykrishna, D.; Bahl, J.; Lycett, S.J.; Worobey, M.; Pybus, O.G.; Ma, S.K.; Cheung, C.L.; Raghwani, J.; Bhatt, S.; et al. Origins and evolutionary genomics of the 2009 swine-origin H1N1 influenza A epidemic. Nature 2009, 459, 1122–1125. [Google Scholar] [CrossRef] [PubMed]
- Rahn, J.; Hoffmann, D.; Harder, T.; Beer, M. Vaccines against influenza a viruses in poultry and swine: Status and future developments. Vaccine 2015, 33, 2414–2424. [Google Scholar] [CrossRef] [PubMed]
- Clegg, C.H.; Roque, R.; Van Hoeven, N.; Perrone, L.; Baldwin, S.L.; Rininger, J.A.; Reed, S.G. Adjuvant solution for pandemic influenza vaccine production. Proc. Natl. Acad. Sci. USA 2012, 109, 17585–17590. [Google Scholar] [CrossRef]
- McNeal, M.M.; Rae, M.N.; Ward, R.L. Effects of different adjuvants on rotavirus antibody responses and protection in mice following intramuscular immunization with inactivated rotavirus. Vaccine 1999, 17, 1573–1580. [Google Scholar] [CrossRef]
- Mutwiri, G.; Benjamin, P.; Soita, H.; Townsend, H.; Yost, R.; Roberts, B.; Andrianov, A.K.; Babiuk, L.A. Poly[di(sodium carboxylatoethylphenoxy)phosphazene] (PCEP) is a potent enhancer of mixed Th1/Th2 immune responses in mice immunized with influenza virus antigens. Vaccine 2007, 25, 1204–1213. [Google Scholar] [CrossRef]
- Mutwiri, G.; Benjamin, P.; Soita, H.; Babiuk, L.A. Co-administration of polyphosphazenes with CpG oligodeoxynucleotides strongly enhances immune responses in mice immunized with Hepatitis B virus surface antigen. Vaccine 2008, 26, 2680–2688. [Google Scholar] [CrossRef]
- Eng, N.F.; Garlapati, S.; Gerdts, V.; Potter, A.; Babiuk, L.A.; Mutwiri, G.K. The potential of polyphosphazenes for delivery of vaccine antigens and immunotherapeutic agents. Curr. Drug Deliv. 2010, 7, 13–20. [Google Scholar] [CrossRef]
- Awate, S.; Wilson, H.L.; Lai, K.; Babiuk, L.A.; Mutwiri, G. Activation of adjuvant core response genes by the novel adjuvant PCEP. Mol. Immunol. 2012, 51, 292–303. [Google Scholar] [CrossRef] [PubMed]
- Magiri, R.; Lai, K.; Chaffey, A.; Zhou, Y.; Pyo, H.-M.; Gerdts, V.; Wilson, H.L.; Mutwiri, G. Intradermal immunization with inactivated swine influenza virus and adjuvant polydi (sodium carboxylatoethylphenoxy) phosphazene (PCEP) induced humoral and cell-mediated immunity and reduced lung viral titres in pigs. Vaccine 2018, 36, 1606–1613. [Google Scholar] [CrossRef] [PubMed]
- Mutwiri, G.; Gerdts, V.; Lopez, M.; Babiuk, L.A. Innate immunity and new adjuvants. Rev. Sci. Technol. 2007, 26, 147–156. [Google Scholar] [CrossRef]
- Magiri, R.; Lai, K.; Huang, Y.; Mutwiri, G.; Wilson, H.L. Innate immune response profiles in pigs injected with vaccine adjuvants polydi (sodium carboxylatoethylphenoxy) phosphazene (PCEP) and Emulsigen. Vet. Immunol. Immunopathol. 2019, 209, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Andrianov, A.K.; Svirkin, Y.Y.; LeGolvan, M.P. Synthesis and biologically relevant properties of polyphosphazene polyacids. Biomacromolecules 2004, 5, 1999–2006. [Google Scholar] [CrossRef]
- Dar, A.; Lai, K.; Dent, D.; Potter, A.; Gerdts, V.; Babiuk, L.A.; Mutwiri, G.K. Administration of poly[di(sodium carboxylatoethylphenoxy)]phosphazene (PCEP) as adjuvant activated mixed Th1/Th2 immune responses in pigs. Vet. Immunol. Immunopathol. 2012, 146, 289–295. [Google Scholar] [CrossRef]
- Magiri, R.; Lai, K.; Chaffey, A.; Wilson, H.; Berry, W.; Szafron, M.; Mutwiri, G. Response of immune response genes to adjuvants poly [di (sodium carboxylatoethylphenoxy) phosphazene](PCEP), CpG oligodeoxynucleotide and emulsigen at intradermal injection site in pigs. Vet. Immunol. Immunopathol. 2016, 175, 57–63. [Google Scholar] [CrossRef]
- Bikour, M.H.; Cornaglia, E.; Elazhary, Y. Evaluation of a protective immunity induced by an inactivated influenza H3N2 vaccine after an intratracheal challenge of pigs. Can. J. Vet. Res. 1996, 60, 312–314. [Google Scholar]
- Ma, W.; Vincent, A.L.; Gramer, M.R.; Brockwell, C.B.; Lager, K.M.; Janke, B.H.; Gauger, P.C.; Patnayak, D.P.; Webby, R.J.; Richt, J.A. Identification of H2N3 influenza A viruses from swine in the United States. Proc. Natl. Acad. Sci. USA 2007, 104, 20949–20954. [Google Scholar] [CrossRef]
- Richt, J.A.; Lager, K.M.; Janke, B.H.; Woods, R.D.; Webster, R.G.; Webby, R.J. Pathogenic and antigenic properties of phylogenetically distinct reassortant H3N2 swine influenza viruses cocirculating in the United States. J. Clin. Microbiol. 2003, 41, 3198–3205. [Google Scholar] [CrossRef]
- Bordet, E.; Frétaud, M.; Crisci, E.; Bouguyon, E.; Rault, S.; Pezant, J.; Pleau, A.; Renson, P.; Giuffra, E.; Larcher, T.; et al. Macrophage-B cell interactions in the inverted porcine lymph node and their response to Porcine Reproductive and Respiratory Syndrome Virus. Front. Immunol. 2019, 10, 953. [Google Scholar] [CrossRef] [PubMed]
- Brown, I.H. The epidemiology and evolution of influenza viruses in pigs. Vet. Microbiol. 2000, 74, 29–46. [Google Scholar] [CrossRef]
- Ma, W.; Gramer, M.; Rossow, K.; Yoon, K.-J. Isolation and genetic characterization of new reassortant H3N1 swine influenza virus from pigs in the midwestern United States. J. Virol. 2006, 80, 5092–5096. [Google Scholar] [CrossRef] [PubMed]
- Simon, G.; Larsen, L.E.; Dürrwald, R.; Foni, E.; Harder, T.; van Reeth, K.; Markowska-Daniel, I.; Reid, S.M.; Dan, A.; Maldonado, J. European surveillance network for influenza in pigs: Surveillance programs, diagnostic tools and Swine influenza virus subtypes identified in 14 European countries from 2010 to 2013. PLoS ONE 2014, 9, e115815. [Google Scholar] [CrossRef] [PubMed]
- Thacker, E.; Janke, B. Swine influenza virus: Zoonotic potential and vaccination strategies for the control of avian and swine influenzas. J. Infect Dis. 2008, 197 (Suppl. 1), S19–S24. [Google Scholar] [CrossRef]
- Ma, W.; Kahn, R.E.; Richt, J.A. The pig as a mixing vessel for influenza viruses: Human and veterinary implications. J. Mol. Genet. Med. 2009, 3, 158. [Google Scholar] [CrossRef]
- Nichol, K.L.; Treanor, J.J. Vaccines for seasonal and pandemic influenza. J. Infect Dis. 2006, 194 (Suppl. 2), S111–S118. [Google Scholar] [CrossRef]
- Lorusso, A.; Vincent, A.L.; Harland, M.L.; Alt, D.; Bayles, D.O.; Swenson, S.L.; Gramer, M.R.; Russell, C.A.; Smith, D.J.; Lager, K.M.; et al. Genetic and antigenic characterization of H1 influenza viruses from United States swine from 2008. J. Gen. Virol. 2011, 92, 919–930. [Google Scholar] [CrossRef]
- Lee, J.H.; Gramer, M.R.; Joo, H.S. Efficacy of swine influenza A virus vaccines against an H3N2 virus variant. Can. J. Vet. Res. 2007, 71, 207. [Google Scholar]
- Schnitzler, S.U.; Schnitzler, P. An update on swine-origin influenza virus A/H1N1: A review. Virus Genes 2009, 39, 279. [Google Scholar] [CrossRef]
- Girard, M.P.; Tam, J.S.; Assossou, O.M.; Kieny, M.P. The 2009 A (H1N1) influenza virus pandemic: A review. Vaccine 2010, 28, 4895–4902. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, H.J.; Müller, M.; Oh, H.M.; Tambyah, P.A.; Joukhadar, C.; Montomoli, E.; Fisher, D.; Berezuk, G.; Fritsch, S.; Löw-Baselli, A.; et al. A clinical trial of a whole-virus H5N1 vaccine derived from cell culture. N. Engl. J. Med. 2008, 358, 2573–2584. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.C.; Wang, H.; Fang, H.H.; Yang, J.G.; Lin, X.J.; Liang, X.F.; Zhang, X.F.; Pan, H.X.; Meng, F.Y.; Hu, Y.M.; et al. A novel influenza A (H1N1) vaccine in various age groups. N. Engl. J. Med. 2009, 361, 2414–2423. [Google Scholar] [CrossRef] [PubMed]
- Kappes, M.A.; Sandbulte, M.R.; Platt, R.; Wang, C.; Lager, K.M.; Henningson, J.N.; Lorusso, A.; Vincent, A.L.; Loving, C.L.; Roth, J.A. Vaccination with NS1-truncated H3N2 swine influenza virus primes T cells and confers cross-protection against an H1N1 heterosubtypic challenge in pigs. Vaccine 2012, 30, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, A.; Pillai, P.S. Innate immunity to influenza virus infection. Nat. Rev. Immunol. 2014, 14, 315. [Google Scholar] [CrossRef]
- Loving, C.L.; Vincent, A.L.; Pena, L.; Perez, D.R. Heightened adaptive immune responses following vaccination with a temperature-sensitive, live-attenuated influenza virus compared to adjuvanted, whole-inactivated virus in pigs. Vaccine 2012, 30, 5830–5838. [Google Scholar] [CrossRef][Green Version]
- Hoft, D.F.; Lottenbach, K.R.; Blazevic, A.; Turan, A.; Blevins, T.P.; Pacatte, T.P.; Yu, Y.; Mitchell, M.C.; Hoft, S.G.; Belshe, R.B. Comparisons of the humoral and cellular immune responses induced by live attenuated influenza vaccine and inactivated influenza vaccine in adults. Clin. Vaccine Immunol. 2017, 24, e00414–e00416. [Google Scholar] [CrossRef]
- O’Donnell, H.; Pham, O.H.; Li, L.-X.; Atif, S.M.; Lee, S.-J.; Ravesloot, M.M.; Stolfi, J.L.; Nuccio, S.-P.; Broz, P.; Monack, D.M. Toll-like receptor and inflammasome signals converge to amplify the innate bactericidal capacity of T helper 1 cells. Immunity 2014, 40, 213–224. [Google Scholar] [CrossRef]
- Baylor, N.W.; Houn, F. Considerations for Licensure of Influenza Vaccines with Pandemic and Prepandemic Indications; Vaccines for Pandemic Influenza Springer: Berlin, Heidelberg, 2009; pp. 453–470. [Google Scholar]
- Wilkinson, T.M.; Li, C.K.; Chui, C.S.; Huang, A.K.; Perkins, M.; Liebner, J.C.; Lambkin-Williams, R.; Gilbert, A.; Oxford, J.; Nicholas, B. Preexisting influenza-specific CD4+ T cells correlate with disease protection against influenza challenge in humans. Nat. Med. 2012, 18, 274. [Google Scholar] [CrossRef]
- Sridhar, S.; Begom, S.; Bermingham, A.; Hoschler, K.; Adamson, W.; Carman, W.; Bean, T.; Barclay, W.; Deeks, J.J.; Lalvani, A. Cellular immune correlates of protection against symptomatic pandemic influenza. Nat. Med. 2013, 19, 1305. [Google Scholar] [CrossRef]
- Khurana, S.; Loving, C.L.; Manischewitz, J.; King, L.R.; Gauger, P.C.; Henningson, J.; Golding, H. Vaccine-induced anti-HA2 antibodies promote virus fusion and enhance influenza virus respiratory disease. Sci. Transl. Med. 2013, 5, 200ra114. [Google Scholar] [CrossRef] [PubMed]
- Co, M.D.T.; Terajima, M.; Thomas, S.J.; Jarman, R.G.; Rungrojcharoenkit, K.; Fernandez, S.; Ennis, F.A. Relationship of preexisting influenza hemagglutination inhibition, complement-dependent lytic, and antibody-dependent cellular cytotoxicity antibodies to the development of clinical illness in a prospective study of A (H1N1) pdm09 Influenza in children. Viral. Immunol. 2014, 27, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Winarski, K.L.; Tang, J.; Klenow, L.; Lee, J.; Coyle, E.M.; Manischewitz, J.; Turner, H.L.; Takeda, K.; Ward, A.B.; Golding, H.; et al. Antibody-dependent enhancement of influenza disease promoted by increase in hemagglutinin stem flexibility and virus fusion kinetics. Proc. Natl. Acad. Sci. USA 2019, 116, 15194–15199. [Google Scholar] [CrossRef] [PubMed]
- Monsalvo, A.C.; Batalle, J.P.; Lopez, M.F.; Krause, J.C.; Klemenc, J.; Hernandez, J.Z.; Maskin, B.; Bugna, J.; Rubinstein, C. Severe pandemic 2009 H1N1 influenza disease due to pathogenic immune complexes. Nat. Med. 2011, 17, 195–199. [Google Scholar] [CrossRef]
- Crowe, J.E. Universal flu vaccines: Primum non nocere. Sci. Transl. Med. 2013, 5, 200fs34. [Google Scholar] [CrossRef][Green Version]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magiri, R.B.; Lai, K.J.; Mutwiri, G.K.; Wilson, H.L. Experimental PCEP-Adjuvanted Swine Influenza H1N1 Vaccine Induced Strong Immune Responses but Did Not Protect Piglets against Heterologous H3N2 Virus Challenge. Vaccines 2020, 8, 235. https://doi.org/10.3390/vaccines8020235
Magiri RB, Lai KJ, Mutwiri GK, Wilson HL. Experimental PCEP-Adjuvanted Swine Influenza H1N1 Vaccine Induced Strong Immune Responses but Did Not Protect Piglets against Heterologous H3N2 Virus Challenge. Vaccines. 2020; 8(2):235. https://doi.org/10.3390/vaccines8020235
Chicago/Turabian StyleMagiri, Royford Bundi, Ken John Lai, George Kiremu Mutwiri, and Heather Lynne Wilson. 2020. "Experimental PCEP-Adjuvanted Swine Influenza H1N1 Vaccine Induced Strong Immune Responses but Did Not Protect Piglets against Heterologous H3N2 Virus Challenge" Vaccines 8, no. 2: 235. https://doi.org/10.3390/vaccines8020235
APA StyleMagiri, R. B., Lai, K. J., Mutwiri, G. K., & Wilson, H. L. (2020). Experimental PCEP-Adjuvanted Swine Influenza H1N1 Vaccine Induced Strong Immune Responses but Did Not Protect Piglets against Heterologous H3N2 Virus Challenge. Vaccines, 8(2), 235. https://doi.org/10.3390/vaccines8020235