Arthritogenic Alphavirus Vaccines: Serogrouping Versus Cross-Protection in Mouse Models

 , ,
, ,  , ,
, ,  , , add
Show full author list
, , add
Show full author list
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Mice, Infection, Virus Titration and Disease Evaluation
2.3. Cell Culture
2.4. The Arthritogenic Alphaviruses
2.5. RRVTT Sequencing and Phylogenetic Tree Construction
2.6. GETV MM2021 Sequencing
2.7. Quantitative RT PCR of RRV Infected Feet
2.8. Histology and Immunohistochemistry
2.9. The Alphaviral Vaccines and Vaccination
2.10. Antibody ELISA and Neutralization Assays
2.11. E1/E2 Contact Residue Visualizations
2.12. Serum, IgM and Ribavirin Treatments of Rag-/- Mice
2.13. Statistics
3. Results
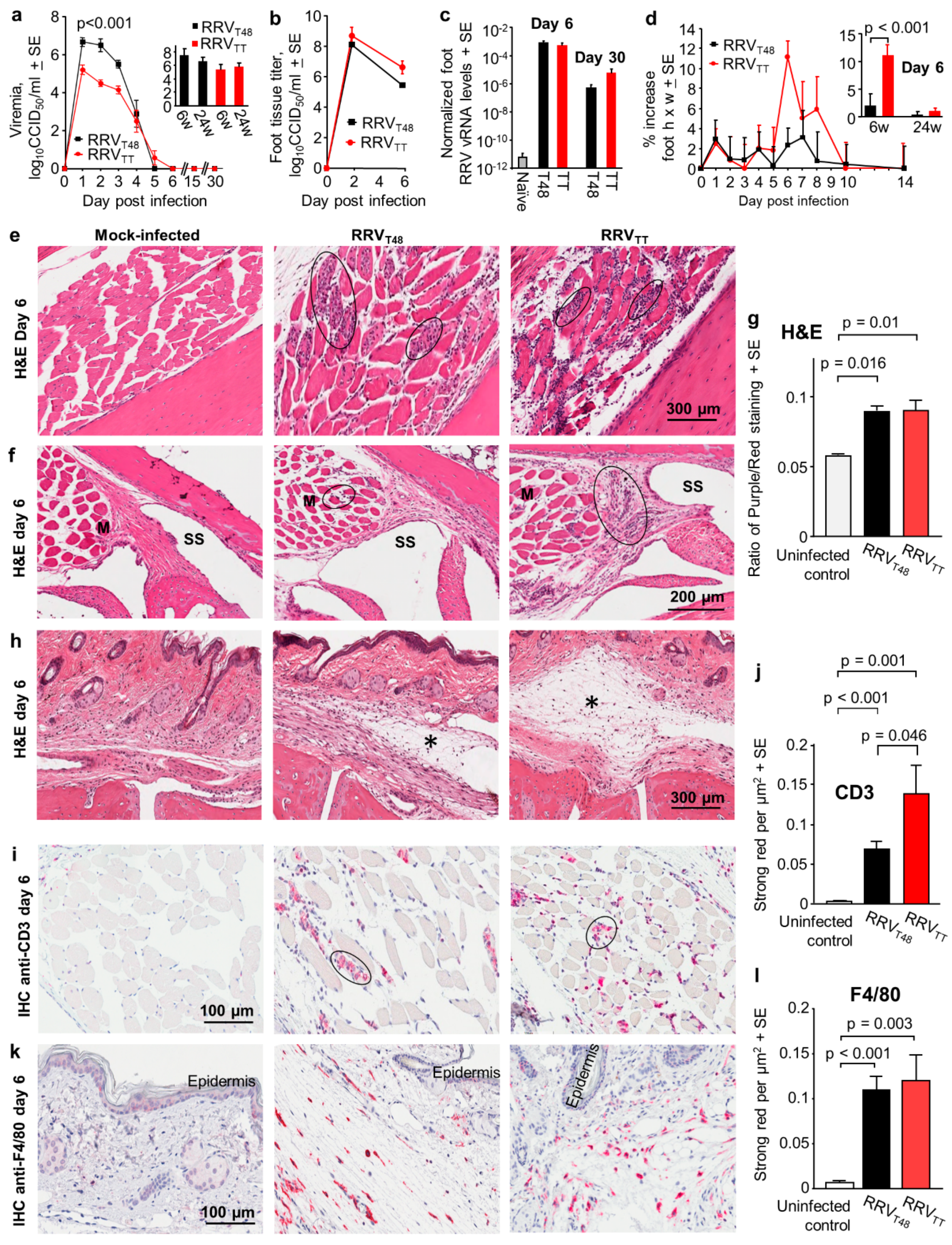
3.1. Characterisation of RRVTT, a Contemporary Human Isolate from an RRV Disease Patient, in Wild-Type Mice
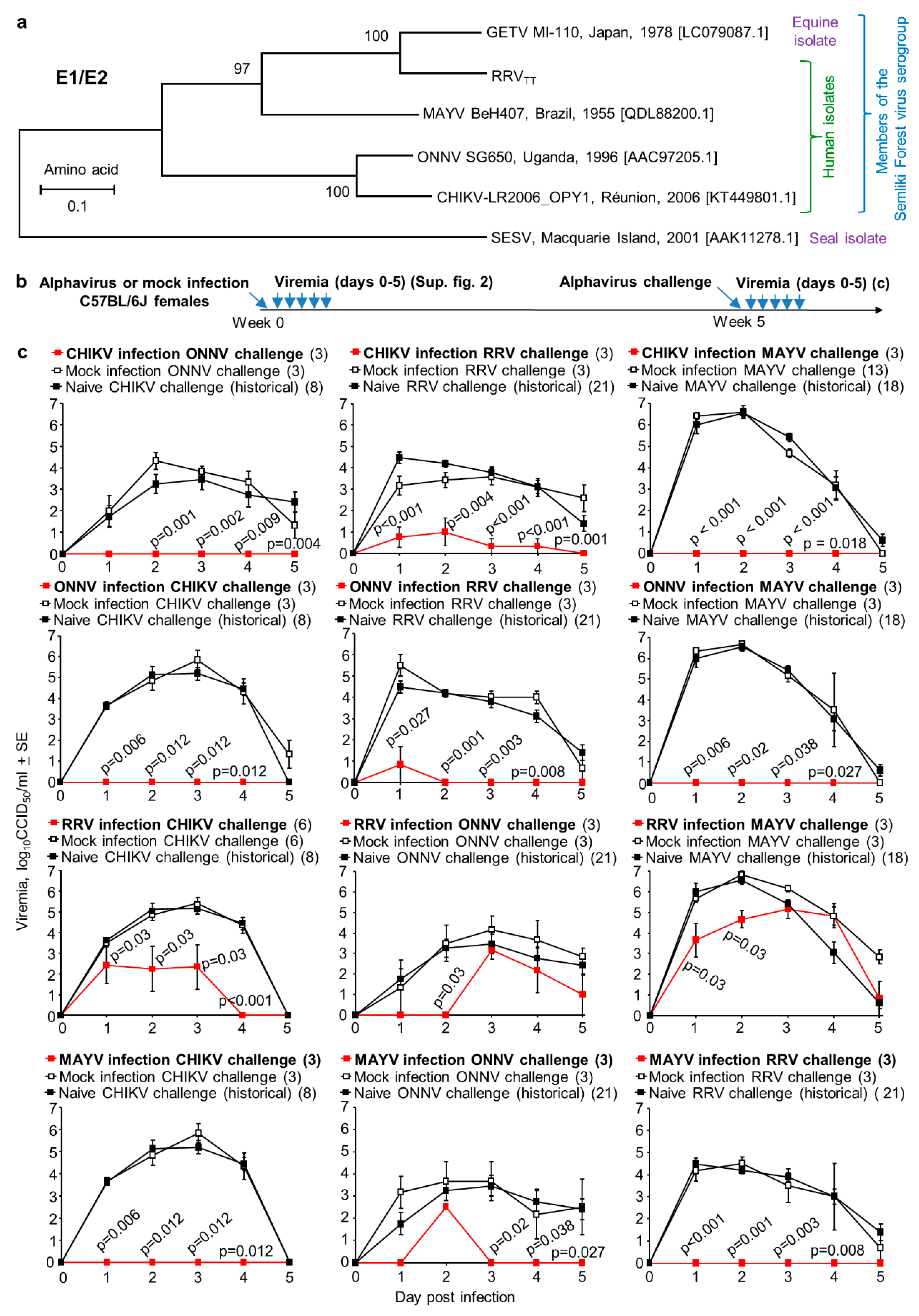
3.2. Protection and Cross-Protection Mediated by Arthritogenic Alphaviruses Infection
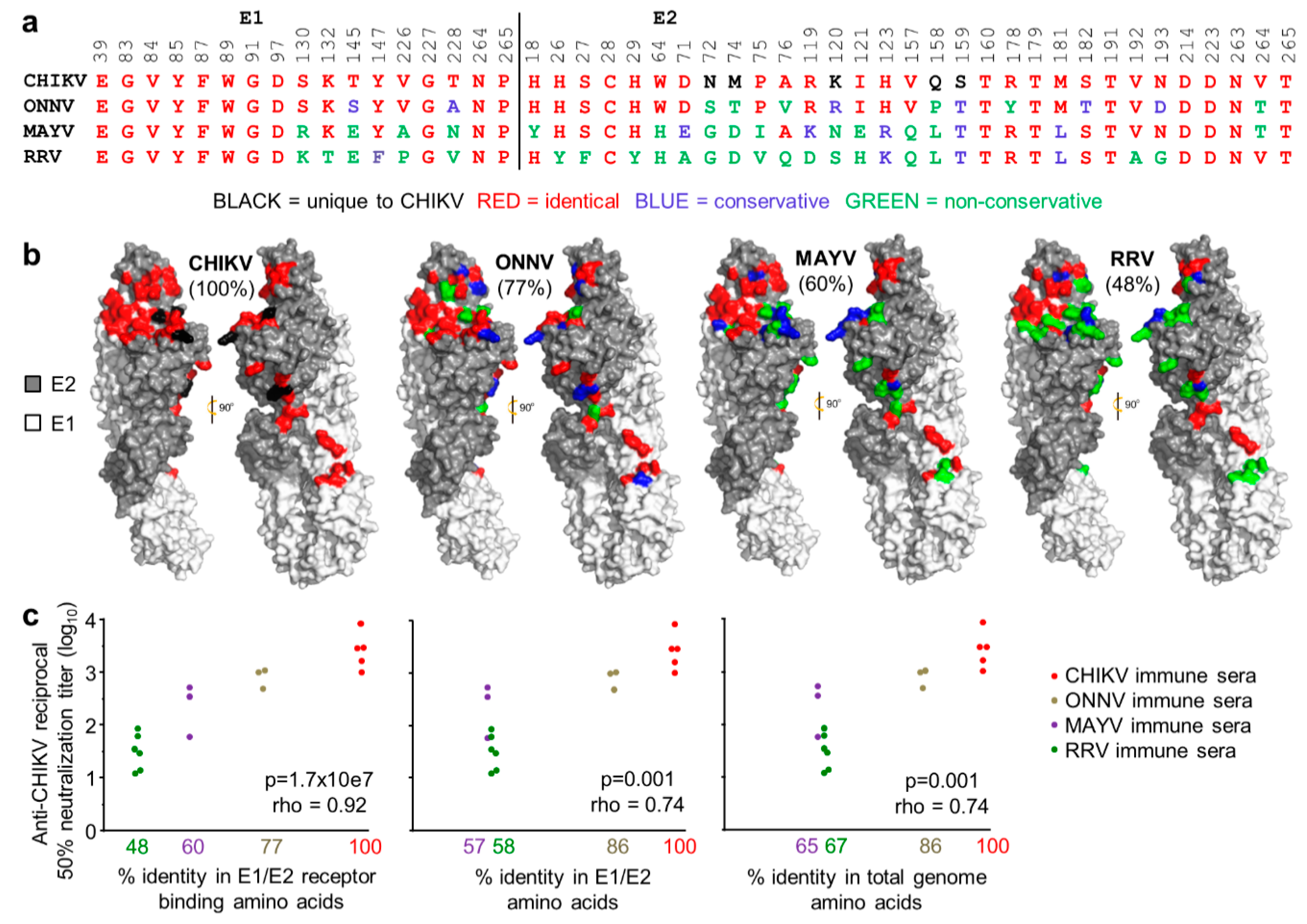
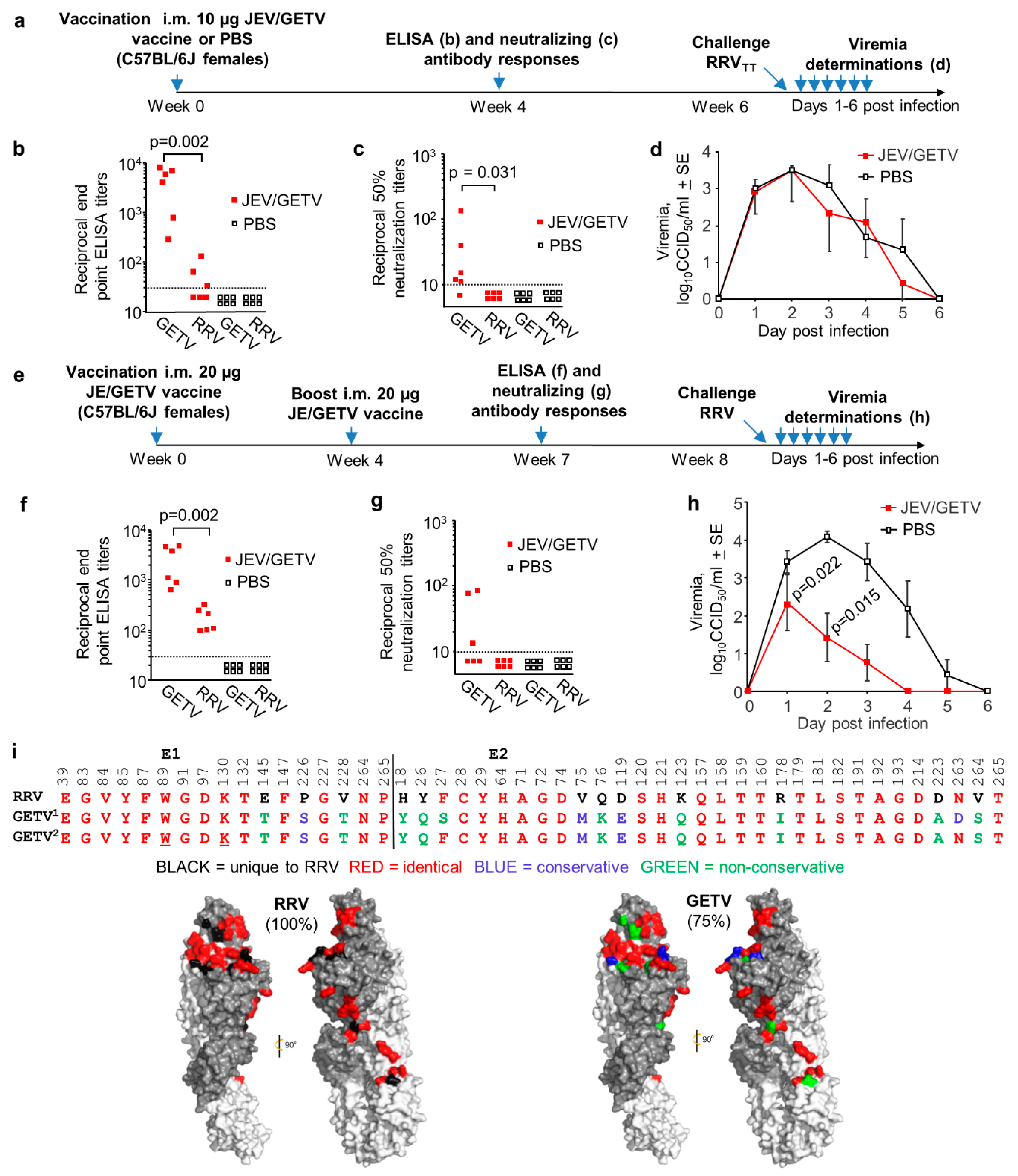
3.3. Conservation of Receptor Contact Residues in E1 and E2
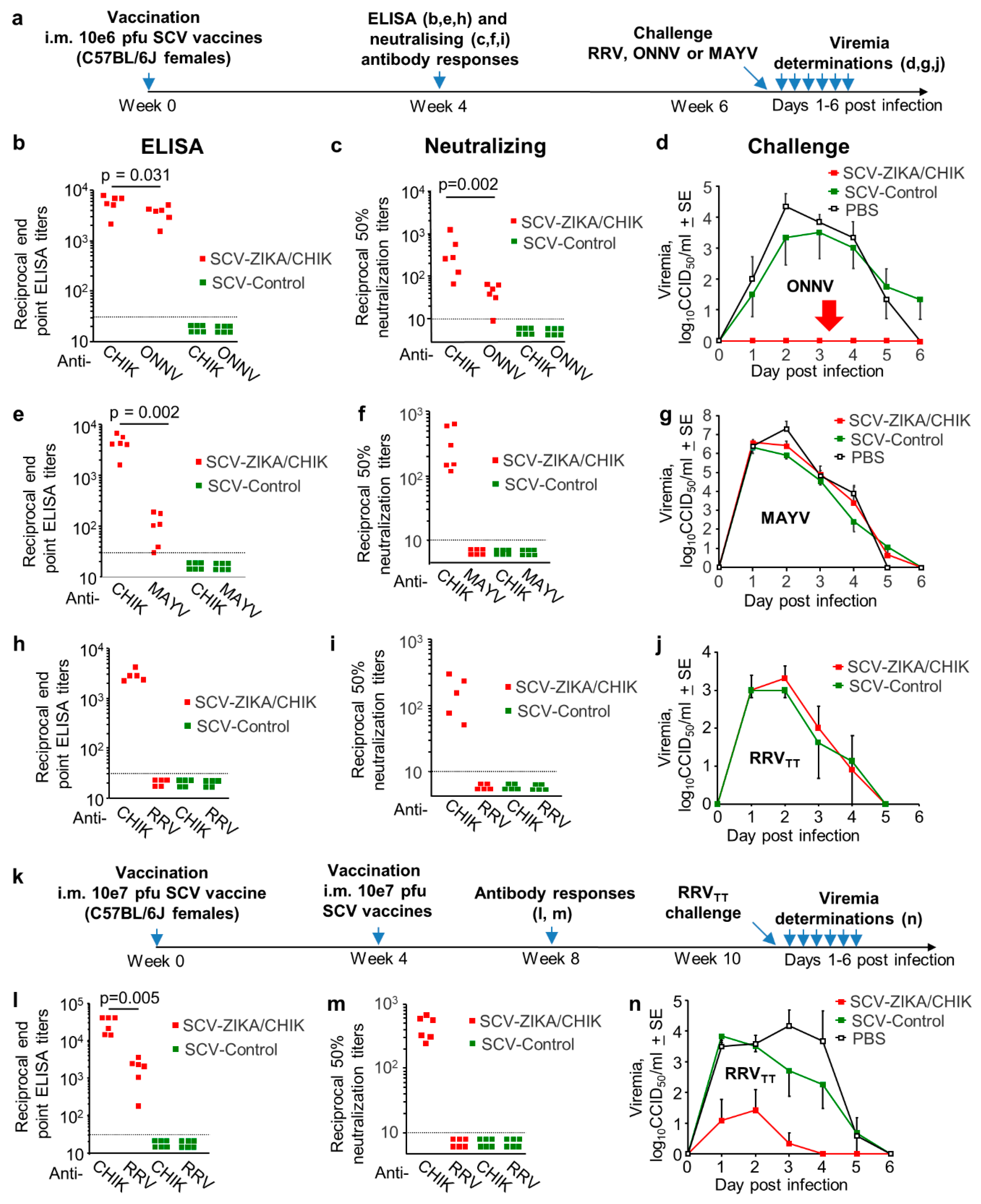
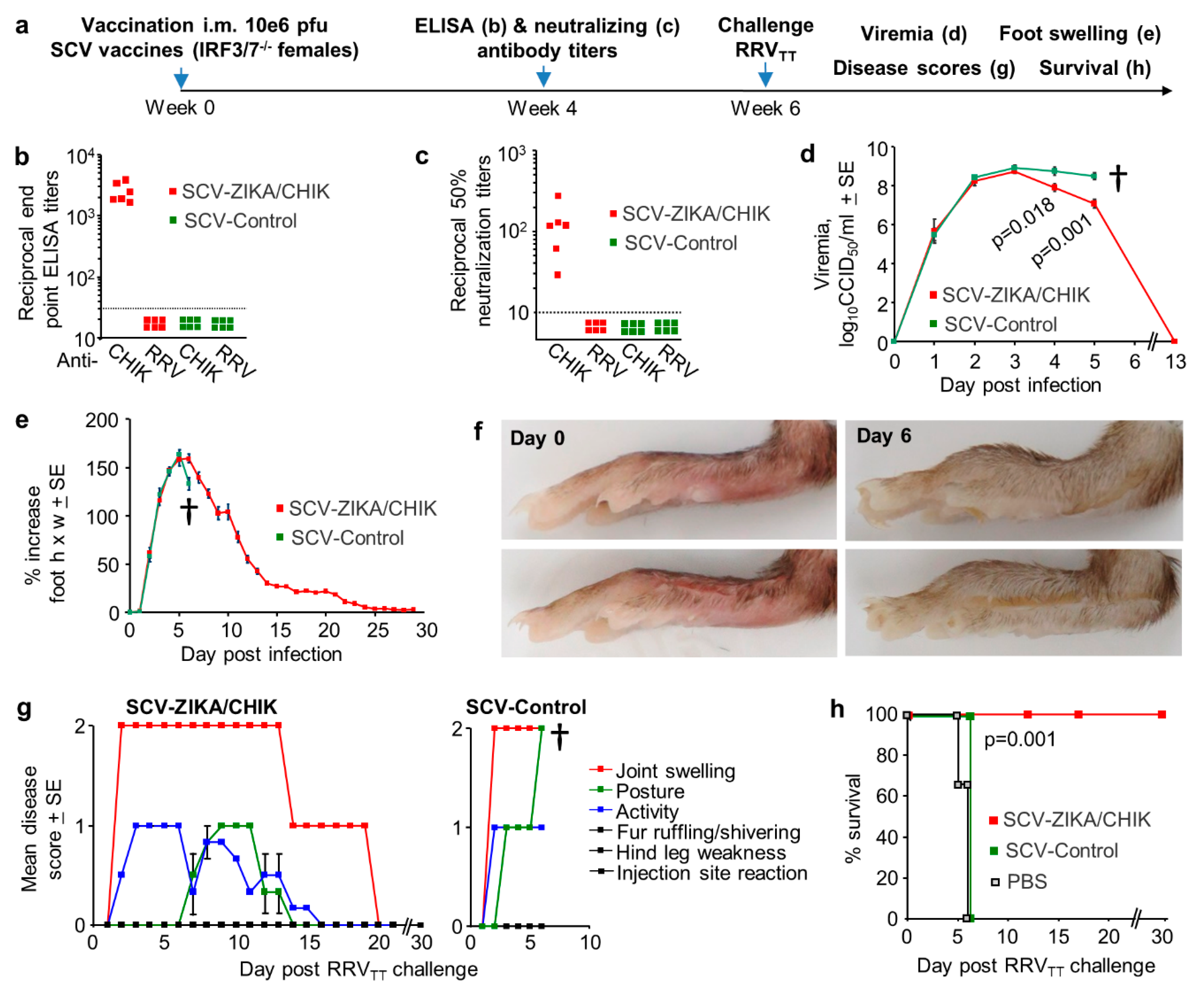
3.4. Cross-Protection Provided by SCV-ZIKA/CHIK Vaccination in Wild-Type Mice
3.5. Cross-Protection Against RRVTT Mediated by the JEV/GETV Vaccine in Wild-Type Mice
3.6. Cross-Protection Against RRVTT after SCV-ZIKA/CHIK Vaccination in IRF3/7-/- Mice
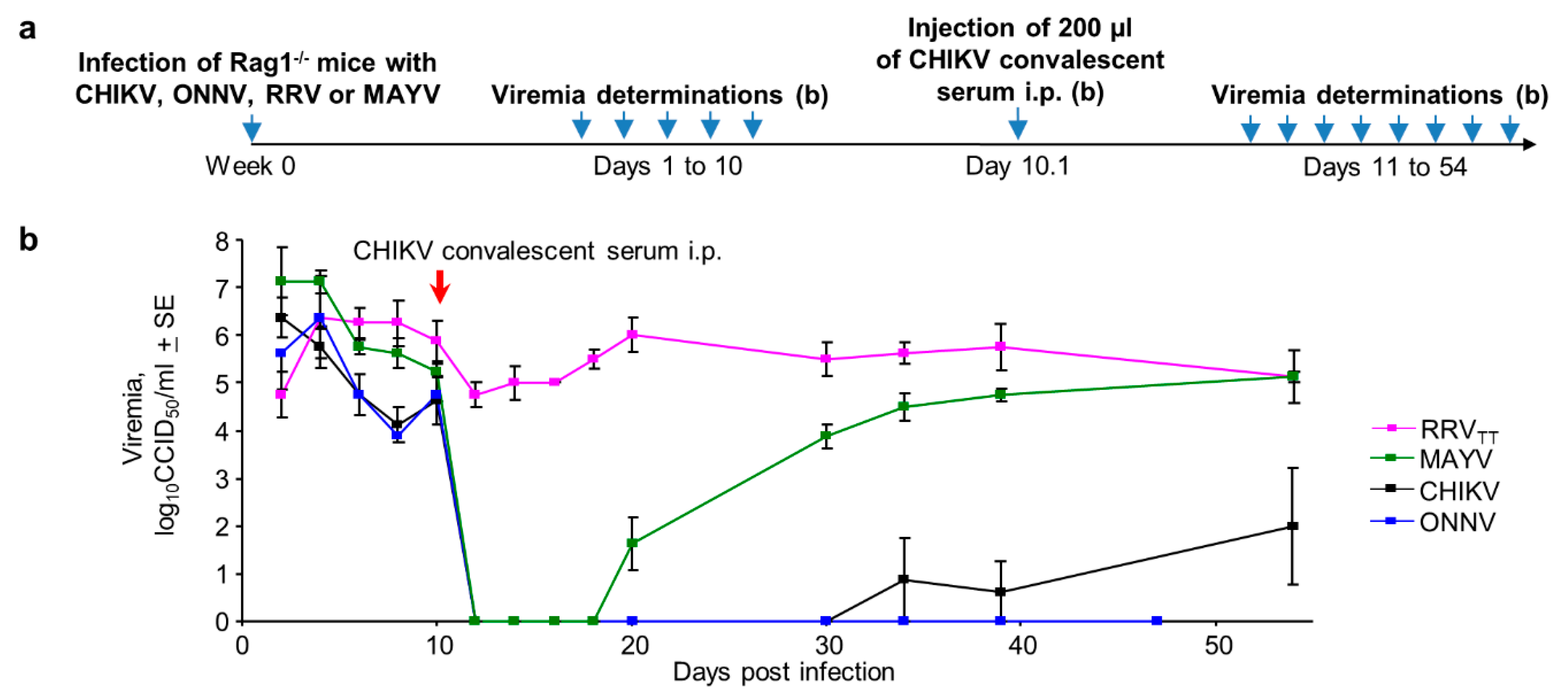
3.7. Serological Protection and Cross-Protection Evaluated in Rag-/- Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pezzi, L.; Rodriguez-Morales, A.J.; Reusken, C.B.; Ribeiro, G.S.; LaBeaud, A.D.; Lourenco-de-Oliveira, R.; Brasil, P.; Lecuit, M.; Failloux, A.B.; Gallian, P.; et al. GloPID-R report on chikungunya, o’nyong-nyong and Mayaro virus, part 3: Epidemiological distribution of Mayaro virus. Antivir. Res. 2019, 172, 104610. [Google Scholar] [CrossRef] [PubMed]
- Pezzi, L.; LaBeaud, A.D.; Reusken, C.B.; Drexler, J.F.; Vasilakis, N.; Diallo, M.; Simon, F.; Jaenisch, T.; Gallian, P.; Sall, A.; et al. GloPID-R report on chikungunya, o’nyong-nyong and Mayaro virus, part 2: Epidemiological distribution of o’nyong-nyong virus. Antivir. Res. 2019, 172, 104611. [Google Scholar] [CrossRef] [PubMed]
- Harley, D.; Suhrbier, A. Ross River Virus Disease. In Hunter’s Tropical Medicine and Emerging Infectious Diseases, 10th ed.; Ryan, E.T., Hill, D.R., Solomon, T., Eds.; Elsevier Health Sciences: Amsterdam, the Netherlands, 2019; Volume 3, pp. 342–344. [Google Scholar]
- Suhrbier, A.; Jaffar-Bandjee, M.C.; Gasque, P. Arthritogenic alphaviruses-an overview. Nat. Rev. Rheumatol. 2012, 8, 420–429. [Google Scholar] [CrossRef] [PubMed]
- Suhrbier, A. Rheumatic manifestations of chikungunya: Emerging concepts and interventions. Nat. Rev. Rheumatol. 2019, 15, 597–611. [Google Scholar] [CrossRef]
- Powers, A.M. Risks to the Americas associated with the continued expansion of chikungunya virus. J. Gen. Virol. 2015, 96, 1–5. [Google Scholar] [CrossRef]
- Farmer, J.F.; Suhrbier, A. Interpreting paired serology for Ross River virus and Barmah Forest virus diseases. Aust. J. Gen. Pract. 2019, 48, 645–649. [Google Scholar] [CrossRef]
- Harley, D.; Sleigh, A.; Ritchie, S. Ross River virus transmission, infection, and disease: A cross-disciplinary review. Clin. Microbiol. Rev. 2001, 14, 909–932. [Google Scholar] [CrossRef]
- Liu, W.; Kizu, J.R.; Le Grand, L.R.; Moller, C.G.; Carthew, T.L.; Mitchell, I.R.; Gubala, A.J.; Aaskov, J.G. Localized Outbreaks of Epidemic Polyarthritis among Military Personnel Caused by Different Sublineages of Ross River Virus, Northeastern Australia, 2016–2017. Emerg. Infect. Dis. 2019, 25, 1793–1801. [Google Scholar] [CrossRef]
- Shanks, G.D. Could Ross River Virus Be the Next Zika? J. Travel. Med. 2019, 26, taz003. [Google Scholar] [CrossRef]
- Lanciotti, R.S.; Ludwig, M.L.; Rwaguma, E.B.; Lutwama, J.J.; Kram, T.M.; Karabatsos, N.; Cropp, B.C.; Miller, B.R. Emergence of epidemic O’nyong-nyong fever in Uganda after a 35-year absence: Genetic characterization of the virus. Virology 1998, 252, 258–268. [Google Scholar] [CrossRef]
- Rezza, G.; Chen, R.; Weaver, S.C. O’nyong-nyong fever: A neglected mosquito-borne viral disease. Pathog. Global Health 2017, 111, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Pezzi, L.; Reusken, C.B.; Weaver, S.C.; Drexler, J.F.; Busch, M.; LaBeaud, A.D.; Diamond, M.S.; Vasilakis, N.; Drebot, M.A.; Siqueira, A.M.; et al. GloPID-R report on Chikungunya, O’nyong-nyong and Mayaro virus, part I: Biological diagnostics. Antivir. Res. 2019, 166, 66–81. [Google Scholar] [CrossRef] [PubMed]
- Acosta-Ampudia, Y.; Monsalve, D.M.; Rodriguez, Y.; Pacheco, Y.; Anaya, J.M.; Ramirez-Santana, C. Mayaro: An emerging viral threat? Emerg. Microbes Infect. 2018, 7, 163. [Google Scholar] [CrossRef] [PubMed]
- Mavian, C.; Rife, B.D.; Dollar, J.J.; Cella, E.; Ciccozzi, M.; Prosperi, M.C.F.; Lednicky, J.; Morris, J.G.; Capua, I.; Salemi, M. Emergence of recombinant Mayaro virus strains from the Amazon basin. Sci. Rep. 2017, 7, 8718. [Google Scholar] [CrossRef] [PubMed]
- Prow, N.A.; Liu, L.; Nakayama, E.; Cooper, T.H.; Yan, K.; Eldi, P.; Hazlewood, J.E.; Tang, B.; Le, T.T.; Setoh, Y.X.; et al. A vaccinia-based single vector construct multi-pathogen vaccine protects against both Zika and chikungunya viruses. Nat. Commun. 2018, 9, 1230. [Google Scholar] [CrossRef]
- DeFilippis, V.R. Chikungunya Virus Vaccines: Platforms, Progress, and Challenges. Curr. Top. Microbiol. Immunol. 2019, 364, 1–26. [Google Scholar]
- Reisinger, E.C.; Tschismarov, R.; Beubler, E.; Wiedermann, U.; Firbas, C.; Loebermann, M.; Pfeiffer, A.; Muellner, M.; Tauber, E.; Ramsauer, K. Immunogenicity, safety, and tolerability of the measles-vectored chikungunya virus vaccine MV-CHIK: A double-blind, randomised, placebo-controlled and active-controlled phase 2 trial. Lancet 2019, 392, 2718–2727. [Google Scholar] [CrossRef]
- Roques, P.; Ljungberg, K.; Kummerer, B.M.; Gosse, L.; Dereuddre-Bosquet, N.; Tchitchek, N.; Hallengard, D.; Garcia-Arriaza, J.; Meinke, A.; Esteban, M.; et al. Attenuated and vectored vaccines protect nonhuman primates against Chikungunya virus. JCI Insight 2017, 2, e83527. [Google Scholar] [CrossRef]
- Rezza, G.; Weaver, S.C. Chikungunya as a paradigm for emerging viral diseases: Evaluating disease impact and hurdles to vaccine development. PLoS Neglected Trop. Dis. 2019, 13, e0006919. [Google Scholar] [CrossRef]
- Powers, A.M. Licensed chikungunya virus vaccine: A possibility? Lancet 2019, 392, 2660–2661. [Google Scholar] [CrossRef]
- Wressnigg, N.; van der Velden, M.V.; Portsmouth, D.; Draxler, W.; O’Rourke, M.; Richmond, P.; Hall, S.; McBride, W.J.; Redfern, A.; Aaskov, J.; et al. An inactivated Ross River virus vaccine is well tolerated and immunogenic in an adult population in a randomized phase 3 trial. Clin. Vaccine Immunol. 2015, 22, 267–273. [Google Scholar] [CrossRef]
- Metz, S.W.; Gardner, J.; Geertsema, C.; Le, T.T.; Goh, L.; Vlak, J.M.; Suhrbier, A.; Pijlman, G.P. Effective chikungunya virus-like particle vaccine produced in insect cells. PLoS Neglected Trop. Dis. 2013, 7, e2124. [Google Scholar] [CrossRef]
- Wang, D.; Suhrbier, A.; Penn-Nicholson, A.; Woraratanadharm, J.; Gardner, J.; Luo, M.; Le, T.T.; Anraku, I.; Sakalian, M.; Einfeld, D.; et al. A complex adenovirus vaccine against chikungunya virus provides complete protection against viraemia and arthritis. Vaccine 2011, 29, 2803–2809. [Google Scholar] [CrossRef]
- Weger-Lucarelli, J.; Aliota, M.T.; Kamlangdee, A.; Osorio, J.E. Identifying the Role of E2 Domains on Alphavirus Neutralization and Protective Immune Responses. PLoS Neglected Trop. Dis. 2015, 9, e0004163. [Google Scholar] [CrossRef]
- Yap, M.L.; Klose, T.; Urakami, A.; Hasan, S.S.; Akahata, W.; Rossmann, M.G. Structural studies of Chikungunya virus maturation. Proc. Natl. Acad. Sci. USA 2017, 114, 13703–13707. [Google Scholar] [CrossRef]
- Jin, J.; Liss, N.M.; Chen, D.H.; Liao, M.; Fox, J.M.; Shimak, R.M.; Fong, R.H.; Chafets, D.; Bakkour, S.; Keating, S.; et al. Neutralizing Monoclonal Antibodies Block Chikungunya Virus Entry and Release by Targeting an Epitope Critical to Viral Pathogenesis. Cell Rep. 2015, 13, 2553–2564. [Google Scholar] [CrossRef]
- Eldi, P.; Cooper, T.H.; Liu, L.; Prow, N.A.; Diener, K.R.; Howley, P.M.; Suhrbier, A.; Hayball, J.D. Production of a Chikungunya Vaccine Using a CHO Cell and Attenuated Viral-Based Platform Technology. Mol. Ther. 2017, 25, 2332–2344. [Google Scholar] [CrossRef]
- Prow, N.A.; Liu, L. The vaccinia-virus based Sementis Copenhagen Vector vaccine against Zika and chikungunya is immunogenic in non-human primates. npj Vaccines, accepted.
- Gouglas, D.; Thanh Le, T.; Henderson, K.; Kaloudis, A.; Danielsen, T.; Hammersland, N.C.; Robinson, J.M.; Heaton, P.M.; Rottingen, J.A. Estimating the cost of vaccine development against epidemic infectious diseases: A cost minimisation study. Lancet Glob. Health 2018, 6, e1386–e1396. [Google Scholar] [CrossRef]
- Bannai, H.; Nemoto, M.; Ochi, A.; Kikuchi, T.; Kobayashi, M.; Tsujimura, K.; Yamanaka, T.; Kondo, T. Epizootiological Investigation of Getah Virus Infection among Racehorses in Japan in 2014. J. Clin. Microbiol. 2015, 53, 2286–2291. [Google Scholar] [CrossRef]
- Bannai, H.; Ochi, A.; Nemoto, M.; Tsujimura, K.; Yamanaka, T.; Kondo, T. A 2015 outbreak of Getah virus infection occurring among Japanese racehorses sequentially to an outbreak in 2014 at the same site. BMC Vet. Res. 2016, 12, 98. [Google Scholar] [CrossRef]
- Nemoto, M.; Bannai, H.; Tsujimura, K.; Kobayashi, M.; Kikuchi, T.; Yamanaka, T.; Kondo, T. Getah Virus Infection among Racehorses, Japan, 2014. Emerg. Infect. Dis. 2015, 21, 883–885. [Google Scholar] [CrossRef]
- Fukunaga, Y.; Kumanomido, T.; Kamada, M. Getah virus as an equine pathogen. Vet. Clin. North Am. Large Anim. Pract. 2000, 16, 605–617. [Google Scholar] [CrossRef]
- Lu, G.; Ou, J.; Ji, J.; Ren, Z.; Hu, X.; Wang, C.; Li, S. Emergence of Getah Virus Infection in Horse With Fever in China, 2018. Front. Microbiol. 2019, 10, 1416. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, X.; Li, L.X.; Shi, N.; Sun, X.T.; Liu, Q.; Jin, N.Y.; Si, X.K. First isolation and characterization of Getah virus from cattle in northeastern China. BMC Vet. Res. 2019, 15, 320. [Google Scholar] [CrossRef]
- Gummow, B.; Tan, R.; Joice, R.K.; Burgess, G.; Picard, J. Seroprevalence and associated risk factors of mosquito-borne alphaviruses in horses in northern Queensland. Aust. Vet. J. 2018, 96, 243–251. [Google Scholar] [CrossRef]
- Stephenson, E.B.; Peel, A.J.; Reid, S.A.; Jansen, C.C.; McCallum, H. The non-human reservoirs of Ross River virus: A systematic review of the evidence. Parasites Vectors 2018, 11, 188. [Google Scholar] [CrossRef]
- El-Hage, C.M.; McCluskey, M.J.; Azuolas, J.K. Disease suspected to be caused by Ross River virus infection of horses. Aust. Vet. J. 2008, 86, 367–370. [Google Scholar] [CrossRef]
- Barton, A.; Bielefeldt-Ohmann, H. Clinical Presentation, Progression, and Management of Five Cases of Ross River Virus Infection in Performance Horses Located in Southeast Queensland: A Longitudinal Case Series. J. Equine Vet. Sci. 2017, 51, 34–40. [Google Scholar] [CrossRef][Green Version]
- Calisher, C.H.; Shope, R.E.; Brandt, W.; Casals, J.; Karabatsos, N.; Murphy, F.A.; Tesh, R.B.; Wiebe, M.E. Proposed antigenic classification of registered arboviruses I. Togaviridae, Alphavirus. Intervirology 1980, 14, 229–232. [Google Scholar] [CrossRef]
- Gardner, J.; Anraku, I.; Le, T.T.; Larcher, T.; Major, L.; Roques, P.; Schroder, W.A.; Higgs, S.; Suhrbier, A. Chikungunya virus arthritis in adult wild-type mice. J. Virol. 2010, 84, 8021–8032. [Google Scholar] [CrossRef] [PubMed]
- Martins, K.A.; Gregory, M.K.; Valdez, S.M.; Sprague, T.R.; Encinales, L.; Pacheco, N.; Cure, C.; Porras-Ramirez, A.; Rico-Mendoza, A.; Chang, A.; et al. Neutralizing Antibodies from Convalescent Chikungunya Virus Patients Can Cross-Neutralize Mayaro and Una Viruses. Am. J. Trop. Med. Hyg. 2019, 100, 1541–1544. [Google Scholar] [CrossRef] [PubMed]
- Webb, E.M.; Azar, S.R.; Haller, S.L.; Langsjoen, R.M.; Cuthbert, C.E.; Ramjag, A.T.; Luo, H.; Plante, K.; Wang, T.; Simmons, G.; et al. Effects of Chikungunya virus immunity on Mayaro virus disease and epidemic potential. Sci. Rep. 2019, 9, 20399. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.M.; Long, F.; Edeling, M.A.; Lin, H.; van Duijl-Richter, M.K.S.; Fong, R.H.; Kahle, K.M.; Smit, J.M.; Jin, J.; Simmons, G.; et al. Broadly Neutralizing Alphavirus Antibodies Bind an Epitope on E2 and Inhibit Entry and Egress. Cell 2015, 163, 1095–1107. [Google Scholar] [CrossRef]
- Earnest, J.T.; Basore, K.; Roy, V.; Bailey, A.L.; Wang, D.; Alter, G.; Fremont, D.H.; Diamond, M.S. Neutralizing antibodies against Mayaro virus require Fc effector functions for protective activity. J. Exp. Med. 2019, 216, 2282–2301. [Google Scholar] [CrossRef]
- Partidos, C.D.; Paykel, J.; Weger, J.; Borland, E.M.; Powers, A.M.; Seymour, R.; Weaver, S.C.; Stinchcomb, D.T.; Osorio, J.E. Cross-protective immunity against o’nyong-nyong virus afforded by a novel recombinant chikungunya vaccine. Vaccine 2012, 30, 4638–4643. [Google Scholar] [CrossRef]
- Holzer, G.W.; Coulibaly, S.; Aichinger, G.; Savidis-Dacho, H.; Mayrhofer, J.; Brunner, S.; Schmid, K.; Kistner, O.; Aaskov, J.G.; Falkner, F.G.; et al. Evaluation of an inactivated Ross River virus vaccine in active and passive mouse immunization models and establishment of a correlate of protection. Vaccine 2011, 29, 4132–4141. [Google Scholar] [CrossRef]
- Fine, D.L.; Allen, W.P.; Wilkins, L.B. Features of cross protection between Sindbis and Venezuelan equine encephalitis viruses in mice--relationship of route of immunization to protection. J. Gen. Virol. 1974, 24, 401–408. [Google Scholar] [CrossRef]
- Wust, C.J.; Crombie, R.; Brown, A. Passive protection across subgroups of alphaviruses by hyperimmune non-cross-neutralizing anti-Sindbis serum. Proc. Soc. Exp. Biol. Med. 1987, 184, 56–63. [Google Scholar] [CrossRef]
- Peck, R.; Wust, C.J.; Brown, A. Adoptive transfer of cross-protection among alphaviruses in mice requires allogeneic stimulation. Infect. Immun. 1979, 25, 320–327. [Google Scholar] [CrossRef]
- Langsjoen, R.M.; Haller, S.L.; Roy, C.J.; Vinet-Oliphant, H.; Bergren, N.A.; Erasmus, J.H.; Livengood, J.A.; Powell, T.D.; Weaver, S.C.; Rossi, S.L. Chikungunya Virus Strains Show Lineage-Specific Variations in Virulence and Cross-Protective Ability in Murine and Nonhuman Primate Models. MBio 2018, 9, e02449-17. [Google Scholar] [CrossRef]
- Hoad, V.C.; Speers, D.J.; Keller, A.J.; Dowse, G.K.; Seed, C.R.; Lindsay, M.D.; Faddy, H.M.; Pink, J. First reported case of transfusion-transmitted Ross River virus infection. Med. J. Aust. 2015, 202, 267–270. [Google Scholar] [CrossRef]
- Prow, N.A.; Jimenez Martinez, R.; Hayball, J.D.; Howley, P.M.; Suhrbier, A. Poxvirus-based vector systems and the potential for multi-valent and multi-pathogen vaccines. Expert. Rev. Vaccines 2018, 17, 925–934. [Google Scholar] [CrossRef]
- Rudd, P.A.; Wilson, J.; Gardner, J.; Larcher, T.; Babarit, C.; Le, T.T.; Anraku, I.; Kumagai, Y.; Loo, Y.M.; Gale, M., Jr.; et al. Interferon response factors 3 and 7 protect against Chikungunya virus hemorrhagic fever and shock. J. Virol. 2012, 86, 9888–9898. [Google Scholar] [CrossRef]
- Poo, Y.S.; Rudd, P.A.; Gardner, J.; Wilson, J.A.; Larcher, T.; Colle, M.A.; Le, T.T.; Nakaya, H.I.; Warrilow, D.; Allcock, R.; et al. Multiple immune factors are involved in controlling acute and chronic chikungunya virus infection. PLoS Neglected Trop. Dis. 2014, 8, e3354. [Google Scholar] [CrossRef]
- Wilson, J.A.; Prow, N.A.; Schroder, W.A.; Ellis, J.J.; Cumming, H.E.; Gearing, L.J.; Poo, Y.S.; Taylor, A.; Hertzog, P.J.; Di Giallonardo, F.; et al. RNA-Seq analysis of chikungunya virus infection and identification of granzyme A as a major promoter of arthritic inflammation. PLoS Pathog. 2017, 13, e1006155. [Google Scholar] [CrossRef]
- Johnson, B.J.; Le, T.T.; Dobbin, C.A.; Banovic, T.; Howard, C.B.; Flores Fde, M.; Vanags, D.; Naylor, D.J.; Hill, G.R.; Suhrbier, A. Heat shock protein 10 inhibits lipopolysaccharide-induced inflammatory mediator production. J. Biol. Chem. 2005, 280, 4037–4047. [Google Scholar] [CrossRef]
- Doherty, R.L.; Carley, J.G.; Mackerras, M.J.; Marks, E.N. Studies of arthropod-borne virus infections in Queensland. III. Isolation and characterization of virus strains from wild-caught mosquitoes in North Queensland. Aust. J. Exp. Biol. Med. Sci. 1963, 41, 17–39. [Google Scholar] [CrossRef]
- Taylor, W.P.; Marshall, I.D. Adaptation studies with Ross River virus: Laboratory mice and cell cultures. J. Gen. Virol. 1975, 28, 59–72. [Google Scholar] [CrossRef]
- Parola, P.; de Lamballerie, X.; Jourdan, J.; Rovery, C.; Vaillant, V.; Minodier, P.; Brouqui, P.; Flahault, A.; Raoult, D.; Charrel, R.N. Novel chikungunya virus variant in travelers returning from Indian Ocean islands. Emerg. Infect. Dis. 2006, 12, 1493–1499. [Google Scholar] [CrossRef]
- Bessaud, M.; Peyrefitte, C.N.; Pastorino, B.A.; Gravier, P.; Tock, F.; Boete, F.; Tolou, H.J.; Grandadam, M. O’nyong-nyong Virus, Chad. Emerg. Infect. Dis. 2006, 12, 1248–1250. [Google Scholar] [CrossRef] [PubMed]
- Varghese, F.S.; Thaa, B.; Amrun, S.N.; Simarmata, D.; Rausalu, K.; Nyman, T.A.; Merits, A.; McInerney, G.M.; Ng, L.F.P.; Ahola, T. The Antiviral Alkaloid Berberine Reduces Chikungunya Virus-Induced Mitogen-Activated Protein Kinase Signaling. J. Virol. 2016, 90, 9743–9757. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, F.P.; Freitas, R.B.; Travassos da Rosa, J.F.; Gabbay, Y.B.; Mello, W.A.; LeDuc, J.W. An outbreak of Mayaro virus disease in Belterra, Brazil. I. Clinical and virological findings. Am. J. Trop. Med. Hyg. 1981, 30, 674–681. [Google Scholar] [CrossRef] [PubMed]
- La Linn, M.; Bellett, A.J.; Parsons, P.G.; Suhrbier, A. Complete removal of mycoplasma from viral preparations using solvent extraction. J. Virol. Methods 1995, 52, 51–54. [Google Scholar] [CrossRef]
- Warrilow, D.; Hall-Mendelin, S.; Hobson-Peters, J.; Prow, N.A.; Allcock, R.; Hall, R.A. Complete coding sequences of three members of the kokobera group of flaviviruses. Genome Announc. 2014, 2, e00890-14. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Faddy, H.M.; Tran, T.V.; Hoad, V.C.; Seed, C.R.; Viennet, E.; Chan, H.T.; Harley, R.; Hewlett, E.; Hall, R.A.; Bielefeldt-Ohmann, H.; et al. Ross River virus in Australian blood donors: Possible implications for blood transfusion safety. Transfusion 2018, 58, 485–492. [Google Scholar] [CrossRef]
- Schroder, W.A.; Le, T.T.; Major, L.; Street, S.; Gardner, J.; Lambley, E.; Markey, K.; MacDonald, K.P.; Fish, R.J.; Thomas, R.; et al. A physiological function of inflammation-associated SerpinB2 is regulation of adaptive immunity. J. Immunol. 2010, 184, 2663–2670. [Google Scholar] [CrossRef]
- Poo, Y.S.; Nakaya, H.; Gardner, J.; Larcher, T.; Schroder, W.A.; Le, T.T.; Major, L.D.; Suhrbier, A. CCR2 deficiency promotes exacerbated chronic erosive neutrophil-dominated chikungunya virus arthritis. J. Virol. 2014, 88, 6862–6872. [Google Scholar] [CrossRef]
- Zouache, K.; Fontaine, A.; Vega-Rua, A.; Mousson, L.; Thiberge, J.M.; Lourenco-De-Oliveira, R.; Caro, V.; Lambrechts, L.; Failloux, A.B. Three-way interactions between mosquito population, viral strain and temperature underlying chikungunya virus transmission potential. Proc. Biol. Sci. 2014, 281, 20141078. [Google Scholar] [CrossRef]
- Song, H.; Zhao, Z.; Chai, Y.; Jin, X.; Li, C.; Yuan, F.; Liu, S.; Gao, Z.; Wang, H.; Song, J.; et al. Molecular Basis of Arthritogenic Alphavirus Receptor MXRA8 Binding to Chikungunya Virus Envelope Protein. Cell 2019, 177, 1714–1724. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, N.M. Analysis of Ross River virus using monoclonal and polyclonal antibodies with applications in diagnostic and surveillance techniques. Ph.D. Thesis, University of Western Australia, Crawley, Australia, 1998. [Google Scholar]
- O’Brien, C.A.; Hobson-Peters, J.; Yam, A.W.; Colmant, A.M.; McLean, B.J.; Prow, N.A.; Watterson, D.; Hall-Mendelin, S.; Warrilow, D.; Ng, M.L.; et al. Viral RNA intermediates as targets for detection and discovery of novel and emerging mosquito-borne viruses. PLoS Neglected Trop. Dis. 2015, 9, e0003629. [Google Scholar] [CrossRef] [PubMed]
- Prow, N.A.; Tang, B.; Gardner, J.; Le, T.T.; Taylor, A.; Poo, Y.S.; Nakayama, E.; Hirata, T.D.C.; Nakaya, H.I.; Slonchak, A.; et al. Lower temperatures reduce type I interferon activity and promote alphaviral arthritis. PLoS Pathog. 2017, 13, e1006788. [Google Scholar] [CrossRef] [PubMed]
- Wolf, S.; Taylor, A.; Zaid, A.; Freitas, J.; Herrero, L.J.; Rao, S.; Suhrbier, A.; Forwood, M.R.; Bucala, R.; Mahalingam, S. Inhibition of Interleukin-1beta Signaling by Anakinra Demonstrates a Critical Role of Bone Loss in Experimental Arthritogenic Alphavirus Infections. Arthritis Rheumatol. 2019, 71, 1185–1190. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Foo, S.S.; Zaid, A.; Teng, T.S.; Herrero, L.J.; Wolf, S.; Tharmarajah, K.; Vu, L.D.; van Vreden, C.; Taylor, A.; et al. Specific inhibition of NLRP3 in chikungunya disease reveals a role for inflammasomes in alphavirus-induced inflammation. Nat. Microbiol. 2017, 2, 1435–1445. [Google Scholar] [CrossRef]
- Seay, A.R.; Griffin, D.E.; Johnson, R.T. Experimental viral polymyositis: Age dependency and immune responses to Ross River virus infection in mice. Neurology 1981, 31, 656–660. [Google Scholar] [CrossRef]
- Rulli, N.E.; Guglielmotti, A.; Mangano, G.; Rolph, M.S.; Apicella, C.; Zaid, A.; Suhrbier, A.; Mahalingam, S. Amelioration of alphavirus-induced arthritis and myositis in a mouse model by treatment with bindarit, an inhibitor of monocyte chemotactic proteins. Arthritis Rheumatol. 2009, 60, 2513–2523. [Google Scholar] [CrossRef]
- Herrero, L.; Nelson, M.; Bettadapura, J.; Gahan, M.E.; Mahalingam, S. Applications of animal models of infectious arthritis in drug discovery: A focus on alphaviral disease. Curr. Drug Targets 2011, 12, 1024–1036. [Google Scholar] [CrossRef]
- Jin, J.; Simmons, G. Antiviral Functions of Monoclonal Antibodies against Chikungunya Virus. Viruses 2019, 11, 305. [Google Scholar] [CrossRef]
- Burrack, K.S.; Montgomery, S.A.; Homann, D.; Morrison, T.E. CD8+ T cells control Ross River virus infection in musculoskeletal tissues of infected mice. J. Immunol. 2015, 194, 678–689. [Google Scholar] [CrossRef]
- Chanas, A.C.; Hubalek, Z.; Johnson, B.K.; Simpson, D.I. A comparative study of O’nyong nyong virus with Chikungunya virus and plaque variants. Arch. Virol. 1979, 59, 231–238. [Google Scholar] [CrossRef] [PubMed]
- La Linn, M.; Gardner, J.; Warrilow, D.; Darnell, G.A.; McMahon, C.R.; Field, I.; Hyatt, A.D.; Slade, R.W.; Suhrbier, A. Arbovirus of marine mammals: A new alphavirus isolated from the elephant seal louse, Lepidophthirus macrorhini. J. Virol. 2001, 75, 4103–4109. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.M.; Diamond, M.S. Immune-Mediated Protection and Pathogenesis of Chikungunya Virus. J. Immunol. 2016, 197, 4210–4218. [Google Scholar] [CrossRef] [PubMed]
- Miyata, T.; Miyazawa, S.; Yasunaga, T. Two types of amino acid substitutions in protein evolution. J. Mol. Evol. 1979, 12, 219–236. [Google Scholar] [CrossRef] [PubMed]
- Quintel, B.K.; Thomas, A.; Poer DeRaad, D.E.; Slifka, M.K.; Amanna, I.J. Advanced oxidation technology for the development of a next-generation inactivated West Nile virus vaccine. Vaccine 2019, 37, 4214–4221. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.C.; Chiu, H.C.; Chen, L.K.; Chang, G.J.; Chiou, S.S. Formalin Inactivation of Japanese Encephalitis Virus Vaccine Alters the Antigenicity and Immunogenicity of a Neutralization Epitope in Envelope Protein Domain III. PLoS Neglected Trop. Dis. 2015, 9, e0004167. [Google Scholar] [CrossRef] [PubMed]
- Vani, K.; Bogen, S.A.; Sompuram, S.R. A high throughput combinatorial library technique for identifying formalin-sensitive epitopes. J. Immunol. Methods 2006, 317, 80–89. [Google Scholar] [CrossRef][Green Version]
- Zhang, Y.N.; Deng, C.L.; Li, J.Q.; Li, N.; Zhang, Q.Y.; Ye, H.Q.; Yuan, Z.M.; Zhang, B. Infectious Chikungunya Virus (CHIKV) with a Complete Capsid Deletion: A New Approach for a CHIKV Vaccine. J. Virol. 2019, 93, e00504-19. [Google Scholar] [CrossRef]
- Brandler, S.; Ruffie, C.; Combredet, C.; Brault, J.B.; Najburg, V.; Prevost, M.C.; Habel, A.; Tauber, E.; Despres, P.; Tangy, F. A recombinant measles vaccine expressing chikungunya virus-like particles is strongly immunogenic and protects mice from lethal challenge with chikungunya virus. Vaccine 2013, 31, 3718–3725. [Google Scholar] [CrossRef]
- Akahata, W.; Yang, Z.Y.; Andersen, H.; Sun, S.; Holdaway, H.A.; Kong, W.P.; Lewis, M.G.; Higgs, S.; Rossmann, M.G.; Rao, S.; et al. A virus-like particle vaccine for epidemic Chikungunya virus protects nonhuman primates against infection. Nat. Med. 2010, 16, 334–338. [Google Scholar] [CrossRef]
- Gardner, J.; Rudd, P.A.; Prow, N.A.; Belarbi, E.; Roques, P.; Larcher, T.; Gresh, L.; Balmaseda, A.; Harris, E.; Schroder, W.A.; et al. Infectious Chikungunya Virus in the Saliva of Mice, Monkeys and Humans. PLoS ONE 2015, 10, e0139481. [Google Scholar] [CrossRef] [PubMed]
- Smith-Owirodu, A.; Wust, C.J.; Gates, D.; Brown, A. Cross-protection correlates with delayed antibody formation to challenge virus after immunization with Sindbis virus. J. Gen. Virol. 1980, 51, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Peck, R.D.; Brown, A.; Wust, C.J. Preliminary evidence for cell-mediated immunity in cross-protection among group A arboviruses. J. Immunol. 1975, 114, 581–584. [Google Scholar] [PubMed]
- Wolcott, J.A.; Wust, C.J.; Brown, A. Cross-reactive target antigen in cell-mediated cytolysis of cells infected with a temperature-sensitive mutant of Sindbis virus. J. Gen. Virol. 1985, 66, 1167–1170. [Google Scholar] [CrossRef] [PubMed]
- Latif, Z.; Gates, D.; Wust, C.J.; Brown, A. Cross protection among togaviruses in nude mice and littermates. J. Gen. Virol. 1979, 45, 89–98. [Google Scholar] [CrossRef]
- Mullbacher, A.; Marshall, I.D.; Blanden, R.V. Cross-reactive cytotoxic T cells to alphavirus infection. Scand. J. Immunol. 1979, 10, 291–296. [Google Scholar] [CrossRef]
- Wust, C.J.; Nicholas, J.A.; Fredin, D.; Dodd, D.C.; Brideau, R.J.; Levely, M.E.; Brown, A. Monoclonal antibodies that cross-react with the E1 glycoprotein of different alphavirus serogroups: Characterization including passive protection in vivo. Virus Res. 1989, 13, 101–112. [Google Scholar] [CrossRef]
- Herve, C.; Laupeze, B.; Del Giudice, G.; Didierlaurent, A.M.; Da Silva, F.T. The how’s and what’s of vaccine reactogenicity. npj Vaccines 2019, 4, 39. [Google Scholar] [CrossRef]
- Yoon, I.K.; Alera, M.T.; Lago, C.B.; Tac-An, I.A.; Villa, D.; Fernandez, S.; Thaisomboonsuk, B.; Klungthong, C.; Levy, J.W.; Velasco, J.M.; et al. High rate of subclinical chikungunya virus infection and association of neutralizing antibody with protection in a prospective cohort in the Philippines. PLoS Neglected Trop. Dis. 2015, 9, e0003764. [Google Scholar] [CrossRef]
- Milligan, G.N.; Schnierle, B.S.; McAuley, A.J.; Beasley, D.W.C. Defining a correlate of protection for chikungunya virus vaccines. Vaccine 2019, 37, 7427–7436. [Google Scholar] [CrossRef]
- Rudd, P.A.; Raphael, A.P.; Yamada, M.; Nufer, K.L.; Gardner, J.; Le, T.T.; Prow, N.A.; Dang, N.; Schroder, W.A.; Prow, T.W.; et al. Effective cutaneous vaccination using an inactivated chikungunya virus vaccine delivered by Foroderm. Vaccine 2015, 33, 5172–5180. [Google Scholar] [CrossRef] [PubMed]
- de Castro-Jorge, L.A.; de Carvalho, R.V.H.; Klein, T.M.; Hiroki, C.H.; Lopes, A.H.; Guimaraes, R.M.; Fumagalli, M.J.; Floriano, V.G.; Agostinho, M.R.; Slhessarenko, R.D.; et al. The NLRP3 inflammasome is involved with the pathogenesis of Mayaro virus. PLoS Pathog. 2019, 15, e1007934. [Google Scholar] [CrossRef] [PubMed]
- Michlmayr, D.; Pak, T.R.; Rahman, A.H.; Amir, E.D.; Kim, E.Y.; Kim-Schulze, S.; Suprun, M.; Stewart, M.G.; Thomas, G.P.; Balmaseda, A.; et al. Comprehensive innate immune profiling of chikungunya virus infection in pediatric cases. Mol. Syst. Biol. 2018, 14, e7862. [Google Scholar] [CrossRef] [PubMed]
- Soares-Schanoski, A.; Baptista Cruz, N.; de Castro-Jorge, L.A.; de Carvalho, R.V.H.; Santos, C.A.D.; Ros, N.D.; Oliveira, U.; Costa, D.D.; Santos, C.; Cunha, M.D.P.; et al. Systems analysis of subjects acutely infected with the Chikungunya virus. PLoS Pathog. 2019, 15, e1007880. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.G.; Steigerwald, R.; Kalla, M.; Volkmann, A.; Noll, D.; Nagata, L.P. Protective efficacy of monovalent and trivalent recombinant MVA-based vaccines against three encephalitic alphaviruses. Vaccine 2018, 36, 5194–5203. [Google Scholar] [CrossRef] [PubMed]
- Devine, G.J.; Overgaard, H.J.; Paul, R.E. Global Vector Control Guidelines—The Need For Co-Creation. Trends Parasitol. 2019, 35, 267–270. [Google Scholar] [CrossRef]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, W.; Nakayama, E.; Yan, K.; Tang, B.; Le, T.T.; Liu, L.; Cooper, T.H.; Hayball, J.D.; Faddy, H.M.; Warrilow, D.; et al. Arthritogenic Alphavirus Vaccines: Serogrouping Versus Cross-Protection in Mouse Models. Vaccines 2020, 8, 209. https://doi.org/10.3390/vaccines8020209
Nguyen W, Nakayama E, Yan K, Tang B, Le TT, Liu L, Cooper TH, Hayball JD, Faddy HM, Warrilow D, et al. Arthritogenic Alphavirus Vaccines: Serogrouping Versus Cross-Protection in Mouse Models. Vaccines. 2020; 8(2):209. https://doi.org/10.3390/vaccines8020209
Chicago/Turabian StyleNguyen, Wilson, Eri Nakayama, Kexin Yan, Bing Tang, Thuy T. Le, Liang Liu, Tamara H. Cooper, John D. Hayball, Helen M. Faddy, David Warrilow, and et al. 2020. "Arthritogenic Alphavirus Vaccines: Serogrouping Versus Cross-Protection in Mouse Models" Vaccines 8, no. 2: 209. https://doi.org/10.3390/vaccines8020209
APA StyleNguyen, W., Nakayama, E., Yan, K., Tang, B., Le, T. T., Liu, L., Cooper, T. H., Hayball, J. D., Faddy, H. M., Warrilow, D., Allcock, R. J. N., Hobson-Peters, J., Hall, R. A., Rawle, D. J., Lutzky, V. P., Young, P., Oliveira, N. M., Hartel, G., Howley, P. M., ... Suhrbier, A. (2020). Arthritogenic Alphavirus Vaccines: Serogrouping Versus Cross-Protection in Mouse Models. Vaccines, 8(2), 209. https://doi.org/10.3390/vaccines8020209